- БИОФИЗИКА АНАЛИЗАТОРОВ

Содержание
- 2. МОДАЛЬНОСТЬ – КАЧЕСТВО ОЩУЩЕНИЙ, ВОЗНИКАЮЩИХ ПОД ДЕЙСТВИЕМ ОПРЕДЕЛЕННЫХ РАЗДРАЖИТЕЛЕЙ. 5 КЛАССИЧЕСКИХ МОДАЛЬНОСТЕЙ: ЗРИТЕЛЬНАЯ СЛУХОВАЯ ТАКТИЛЬНАЯ ОБОНЯТЕЛЬНАЯ
- 3. ЗАКОН СПЕЦИФИЧЕСКИХ ЭНЕРГИЙ И.МЮЛЛЕРА (I-я половина XIX века) 1801-1858 Иоганнес Мюллер
- 4. ОДНО И ТО ЖЕ РАЗДРАЖЕНИЕ, ДЕЙСТВУЯ НА РАЗНЫЕ ОРГАНЫ ЧУВСТВ, ВЫЗЫВАЕТ ОЩУЩЕНИЯ РАЗЛИЧНОГО КАЧЕСТВА. РАЗНЫЕ РАЗДРАЖИТЕЛИ,
- 5. ПОРОГИ РАЗДРАЖЕНИЯ
- 6. АБСОЛЮТНЫЙ ПОРОГ – минимальная величина раздражителя, вызывающая ответ анализатора ПОРОГ определяет возбудимость E – возбудимость J
- 7. ДИФФЕРЕНЦИАЛЬНЫЙ ПОРОГ – ОТНОШЕНИЕ МИНИМАЛЬНО ВОСПРИНИМАЕМОГО ПРИРОСТА СТИМУЛА К ИСХОДНОЙ ВЕЛИЧИНЕ. ЭТО ВЕЛИЧИНА ПОСТОЯННАЯ
- 8. Г.Т. Фехнер (1801-1887) Основатель психофизики Э.Г. Вебер (1795-1878) Сформулировал закон различной чувствительности
- 9. ОЩУЩЕНИЕ изменяется пропорционально логарифму раздражителя. Закон справедлив для любых раздражителей. Закон Вебера- Фехнера S – ощущение,
- 10. ЗАКОН С. СТИВЕНСА Интенсивность ощущения описывается степенной функцией I – интенсивность ощущения S0 - порог n
- 11. n определяется углом наклона α
- 12. ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ СЕНСОРНЫХ СИСТЕМ рецептор Проводниковый отдел Центральный отдел (кора больших полушарий)
- 13. РЕЦЕПТОР:воспринимает и преобразует определенный стимул внешней среды
- 14. МЕХАНИЗМ ПРЕОБРАЗОВАНИЯ В СЕНСОРНОЙ КЛЕТКЕ: ЭНЕРГИЯ ДЕЙСТВУЮЩЕГО СТИМУЛА ПРЕОБРАЗУЕТСЯ В ЭНЕРГИЮ ЭЛЕКТРИЧЕСКОГО ИМПУЛЬСА. ВОЗДЕЙСТВИЕ ФАКТОРА НА
- 15. МЕХАНИЗМ СЕНСОРНОЙ ТРАНСДУКЦИИ РЕЦЕПТОР + СТИМУЛ (свет, химический агент и др.) Активация G-белка G-белок активирует специфический
- 16. Схема основных путей сенсорной трансдукции с участием внутриклеточных посредников.
- 17. БИОФИЗИКА СЛУХОВОГО АНАЛИЗАТОРА
- 18. АДЕКВАТНЫЙ РАЗДРАЖИТЕЛЬ – ЗВУК ДИАПАЗОН ВОСПРИНИМАЕМЫХ ЗВУКОВ от 15 – 16 Гц до 20 – 22
- 19. наружное среднее внутреннее
- 20. ФУНКЦИИ НАРУЖНОГО УХА АКУСТИЧЕСКАЯ ЗАЩИТНАЯ ЭСТЕТИЧЕСКАЯ
- 21. 1 – БАРАБАННАЯ ПЕРЕПОНКА 2 – ЕВСТАХИЕВА ТРУБА 3 – 5 СЛУХОВЫЕ КОСТОЧКИ 3 – МОЛОТОЧЕК
- 22. ФУНКЦИИ СРЕДНЕГО УХА ТРАНСФОРМАЦИЯ ЗВУКОВЫХ КОЛЕБАНИЙ ВОЗДУХА В КОЛЕБАНИЯ ЖИДКОСТИ В УЛИТКЕ УСИЛЕНИЕ СИГНАЛА ЗАЩИТНАЯ РОЛЬ
- 23. УСИЛЕНИЕ СИГНАЛА ДВА МЕХАНИЗМА: ПЛОЩАДЬ БАРАБАННОЙ ПЕРЕПОНКИ БОЛЬШЕ ПЛОЩАДИ ОВАЛЬНОГО ОКНА в 14 – 18 раз
- 24. ДВА ТИПА КОЛЕБАНИЯ СТРЕМЕЧКА А- вращение стремечка вокруг вертикальной оси (слабая, средняя сила звука) Б- вращение
- 25. БИОФИЗИКА ВНУТРЕННЕГО УХА
- 27. ВЕСТИБУЛЯРНЫЙ И БАРАБАННЫЙ КАНАЛЫ ЗАПОЛНЕНЫ ПЕРИЛИМФОЙ, УЛИТКОВЫЙ КАНАЛ – ЭНДОЛИМФОЙ ПЕРИЛИМФА: 140 ммоль/л Na+ ЭНДОЛИМФА: 155
- 28. Схематическое изображение улитки ( А ) и ее поперечный разрез ( Б ). СОСУДИСТАЯ ПОЛОСКА Вестибулярный
- 29. Особенности основной мембраны
- 30. ОСОБЕННОСТЬ ОСНОВНОЙ (БАЗИЛЯРНОЙ МЕМБРАНЫ) НАРАСТАНИЕ ЖЕСТКОСТИ ОТ ВЕРШИНЫ УЛИТКИ К ОСНОВАНИЮ
- 31. КОРТИЕВ ОРГАН
- 32. ВОЛОСКОВАЯ КЛЕТКА СТЕРЕОЦИЛИИ ЦИТОСКЕЛЕТ ВОЛОСКОВЫХ КЛЕТОК ЭЛЕКТРОНОГРАММА ВОЛОСКОВОЙ КЛЕТКИ МОРСКОЙ СВИНКИ 1- стереоцилии 2- кутикулярная пластинка
- 34. ТЕОРИИ КОДИРОВАНИЯ ЗВУКА В УЛИТКЕ РЕЗОНАНСНАЯ ТЕОРИЯ Г.ГЕЛЬМГОЛЬЦА ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ Г.БЕКЕШИ
- 35. Г. ГЕЛЬМГОЛЬЦ РЕЗОНАНСНАЯ ТЕОРИЯ ОСНОВНАЯ МЕМБРАНА СОСТОИТ ИЗ МНОЖЕСТВА НАТЯНУТЫХ СТРУН, НАСТРОЕННЫХ В РЕЗОНАНС К ОПРЕДЕЛЕННЫМ
- 36. ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА ОСНОВНАЯ МЕМБРАНА – ЖЕСТКАЯ ПЛАСТИНКА ПОСТОЯННЫЙ ТОН ЗАСТАВЛЯЕТ ВСЮ МЕМБРАНУ КОЛЕБАТЬСЯ В ФАЗЕ
- 37. Г. БЕКЕШИ ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ Нобелевская премия по физиологии и медицине, 1961 г. 1899 –1972
- 38. Улитка помещена под водой и открыта на своей вершине. Маленькая металлическая трубочка R укрепляется вместо стремечка
- 39. Когда прикладывается давление (см. направление стрелки), то улиточный ход вдавливается в барабанный ход, что показано пунктиром
- 41. А – бегущая волна в базилярной мембране, Б – амплитуда колебаний базилярной мембраны (бегущей волны) при
- 42. АМПЛИТУДЫ СМЕЩЕНИЯ ВДОЛЬ ОСНОВНОЙ МЕМБРАНЫ ДЛЯ РАЗНЫХ ЧАСТОТ
- 43. ЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ УЛИТКИ
- 44. МИКРОФОННЫЙ ПОТЕНЦИАЛ (МФ) ОБНАРУЖЕН УИВЕРОМ И БРЕЕМ В 1930 ГОДУ. ПРИЗНАКИ: НЕ ИМЕЕТ ПОРОГА ПАРАМЕТРЫ МФ
- 46. ЭНДОКОХЛЕАРНЫЙ ПОТЕНЦИАЛ ПЕРИЛИМФА ЭНДОЛИМФА + + + ВЕСТИБУЛЯРНЫЙ КАНАЛ УЛИТКОВЫЙ КАНАЛ -2 Мв - 40 Мв
- 47. ОСНОВНАЯ МЕМБРАНА ЭНДОЛИМФА СТЕРЕОЦИЛИИ - + До150 мВ эндолимфа +60…+80 перилимфа 0 Цитоплазма волосковой клетки -
- 48. СМЕЩЕНИЕ ОСНОВНОЙ МЕМБРАНЫ МОДУЛЯЦИЯ ЭНДОКОХЛЕАРНОГО ПОТЕНЦИАЛА ИЗМЕНЕНИЕ СОПРОТИВЛЕНИЯ ВОЛОСКОВЫХ КЛЕТОК МИКРОФОННЫЙ ПОТЕНЦИАЛ
- 49. Электрические реакции слухового рецептора на раздражение. 2 – стереоцилии, 3 – киноцилия, 5 – афферентное волокно.
- 50. БИОФИЗИКА ЗРИТЕЛЬНОГО АНАЛИЗАТОРА
- 51. ВИДИМЫЙ СВЕТ ОТ 400 ДО 750 НМ ЗРЕНИЕ БАЗИРУЕТСЯ НА ВОСПРИЯТИИ КОНТРАСТОВ СВЕТЛОГО И ТЕМНОГО, ЦВЕТОВЫХ
- 52. ЯДРО ГЛАЗНОГО ЯБЛОКА: ХРУСТАЛИК, ВОДЯНИСТАЯ ВЛАГА, СТЕКЛОВИДНОЕ ТЕЛО КАПСУЛА ГЛАЗНОГО ЯБЛОКА: ОБОЛОЧКИ ГЛАЗА
- 53. ЯДРО ГЛАЗНОГО ЯБЛОКА ХРУСТАЛИК: ДВОЯКОВЫПКУЛАЯ ЛИНЗА ПРОЗРАЧНЫЙ И БЕСЦВЕТНЫЙ НАХОДИТСЯ В ПРОЗРАЧНОЙ КАПСУЛЕ ВОЛОКНА ХРУСТАЛИКА ОБРАЗОВАНЫ
- 54. КАПСУЛА ГЛАЗНОГО ЯБЛОКА БЕЛОЧНАЯ ОБОЛОЧКА: СКЛЕРА + РОГОВИЦА СОСУДИСТАЯ ОБОЛОЧКА: СОБСТВЕННО СОСУДИСТАЯ ОБОЛОЧКА + РЕСНИЧНОЕ ТЕЛО
- 55. ФОРМИРОВАНИЕ ИЗОБРАЖЕНИЯ НА СЕТЧАТКЕ
- 56. РЕДУЦИРОВАННЫЙ ГЛАЗ
- 57. ИЗОБРАЖЕНИЕ НА СЕТЧАТКЕ РЕДУЦИРОВАННОГО ГЛАЗА: ДЕЙСТВИТЕЛЬНОЕ УМЕНЬШЕННОЕ ПЕРЕВЕРНУТОЕ
- 58. СТРОЕНИЕ СЕТЧАТКИ
- 59. СВЕТ
- 60. ФУНКЦИИ ПИГМЕНТНОГО СЛОЯ: ПИТАНИЕ ДЛЯ ФОТОРЕЦЕПТОРОВ ОБНОВЛЕНИЕ ФОТОРЕЦЕПТОРОВ ПОГЛОЩЕНИЕ СВЕТА, НЕ ПОГЛОТИВШЕГОСЯ ФОТОРЕЦЕПТОРАМИ ФУНКЦИЯ СЕТЧАТКИ: ВОСПРИЯТИЕ
- 61. СТРОЕНИЕ ПАЛОЧЕК И КОЛБОЧЕК 7млн. сосредоточены в центре сетчатки Цветное зрение 120 млн. Сосредоточены на периферии
- 62. У ЧЕЛОВЕКА 1000 ФОТОРЕЦЕПТОРНЫХ ДИСКОВ НА ОДИН НАРУЖНЫЙ СЕГМЕНТ
- 63. СОСТАВ ФОТОРЕЦЕПТОРНЫХ ДИСКОВ БЕЛКИ – ОКОЛО 60%, ЛИПИДЫ – ОКОЛО 40%, УГЛЕВОДЫ – МЕНЕЕ 4% ЛИПИДНЫЙ
- 64. БЕЛКОВЫЙ СОСТАВ ИНТЕГРАЛЬНЫЕ МЕМБРАННЫЕ: РОДОПСИН (95% ) ЭКСТРАГИРУЕМЫЕ: G-БЕЛОК ТРАНСДУЦИН ФОСФОДИЭСТЕРАЗА цГМФ РОДОПСИНКИНАЗА ГУАНИЛАТЦИКЛАЗА
- 65. ЭЛЕКТРОФИЗИОЛОГИЯ ФОТОРЕЦЕПТОРОВ
- 66. ИОННЫЕ КАНАЛЫ цГМФ-ЗАВИСИМЫЕ, ОТКРЫТЫ ПРИ ВЗАИМОДЕЙСТВИИ С цГМФ В ТЕМНОТЕ: ВЫСОКИЙ УРОВЕНЬ цГМФ, КАНАЛЫ ОТКРЫТЫ НА
- 68. БЕЛКИ ЗРИТЕЛЬНОГО КАСКАДА РОДОПСИН G-белок ТРАНСДУЦИН ЦГМФ-ФОСФОДИЭСТЕРАЗА
- 69. РОДОПСИН – АКЦЕПТОР ФОТОНА ИНТЕГРАЛЬНЫЙ БЕЛОК ОПСИН + 11-ЦИС-РЕТИНАЛЬ (ПРОИЗВОДНОЕ ВИТАМИНА А, СЛУЖИТ ЛОВУШКОЙ ДЛЯ ФОТОНА)
- 70. 11-ЦИС-РЕТИНАЛЬ ПОД ДЕЙСТВИЕМ СВЕТА ИЗОМЕРИРУЕТСЯ В ТРАНС-РЕТИНАЛЬ ЭТО ИЗМЕНЯЕТ ПРОСТРАНСТ-ВЕННУЮ СТРУКТУРУ ОПСИНА
- 74. НАДЕЮСЬ, ЧТО КУРС ЛЕКЦИЙ ПО БИОФИЗИКЕ ПОМОЖЕТ В ВАШЕЙ ПРОФЕССИОНАЛЬНОЙ ДЕЯТЕЛЬНОСТИ
- 76. Скачать презентацию
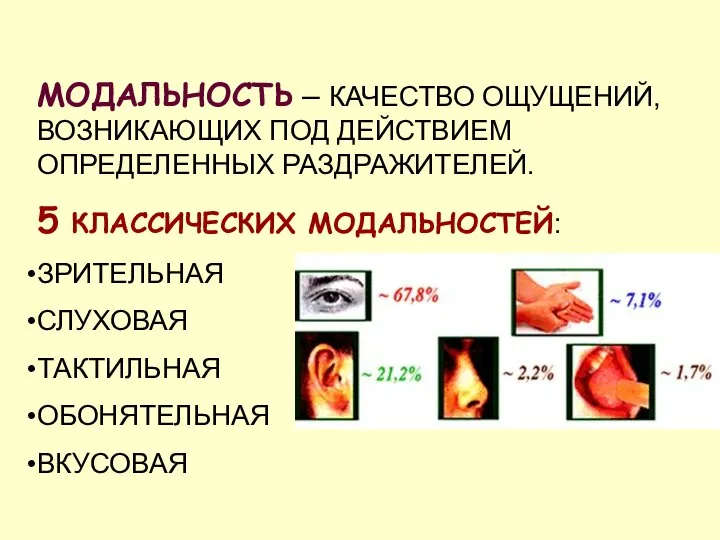
МОДАЛЬНОСТЬ – КАЧЕСТВО ОЩУЩЕНИЙ, ВОЗНИКАЮЩИХ ПОД ДЕЙСТВИЕМ ОПРЕДЕЛЕННЫХ РАЗДРАЖИТЕЛЕЙ.
5 КЛАССИЧЕСКИХ МОДАЛЬНОСТЕЙ:
ЗРИТЕЛЬНАЯ
СЛУХОВАЯ
ТАКТИЛЬНАЯ
ОБОНЯТЕЛЬНАЯ
ВКУСОВАЯ
МОДАЛЬНОСТЬ – КАЧЕСТВО ОЩУЩЕНИЙ, ВОЗНИКАЮЩИХ ПОД ДЕЙСТВИЕМ ОПРЕДЕЛЕННЫХ РАЗДРАЖИТЕЛЕЙ.
5 КЛАССИЧЕСКИХ МОДАЛЬНОСТЕЙ:
ЗРИТЕЛЬНАЯ
СЛУХОВАЯ
ТАКТИЛЬНАЯ
ОБОНЯТЕЛЬНАЯ
ВКУСОВАЯ

ЗАКОН СПЕЦИФИЧЕСКИХ ЭНЕРГИЙ И.МЮЛЛЕРА
(I-я половина XIX века)
1801-1858
Иоганнес Мюллер
ЗАКОН СПЕЦИФИЧЕСКИХ ЭНЕРГИЙ И.МЮЛЛЕРА
(I-я половина XIX века)
1801-1858
Иоганнес Мюллер

ОДНО И ТО ЖЕ РАЗДРАЖЕНИЕ, ДЕЙСТВУЯ НА РАЗНЫЕ ОРГАНЫ ЧУВСТВ, ВЫЗЫВАЕТ

ПОРОГИ РАЗДРАЖЕНИЯ
ПОРОГИ РАЗДРАЖЕНИЯ

АБСОЛЮТНЫЙ ПОРОГ – минимальная величина раздражителя, вызывающая ответ анализатора
ПОРОГ определяет возбудимость
E
АБСОЛЮТНЫЙ ПОРОГ – минимальная величина раздражителя, вызывающая ответ анализатора
ПОРОГ определяет возбудимость
E

ДИФФЕРЕНЦИАЛЬНЫЙ ПОРОГ – ОТНОШЕНИЕ МИНИМАЛЬНО ВОСПРИНИМАЕМОГО ПРИРОСТА СТИМУЛА К ИСХОДНОЙ ВЕЛИЧИНЕ.
ДИФФЕРЕНЦИАЛЬНЫЙ ПОРОГ – ОТНОШЕНИЕ МИНИМАЛЬНО ВОСПРИНИМАЕМОГО ПРИРОСТА СТИМУЛА К ИСХОДНОЙ ВЕЛИЧИНЕ.

Г.Т. Фехнер
(1801-1887)
Основатель психофизики
Э.Г. Вебер
(1795-1878)
Сформулировал закон различной
Г.Т. Фехнер
(1801-1887)
Основатель психофизики
Э.Г. Вебер
(1795-1878)
Сформулировал закон различной
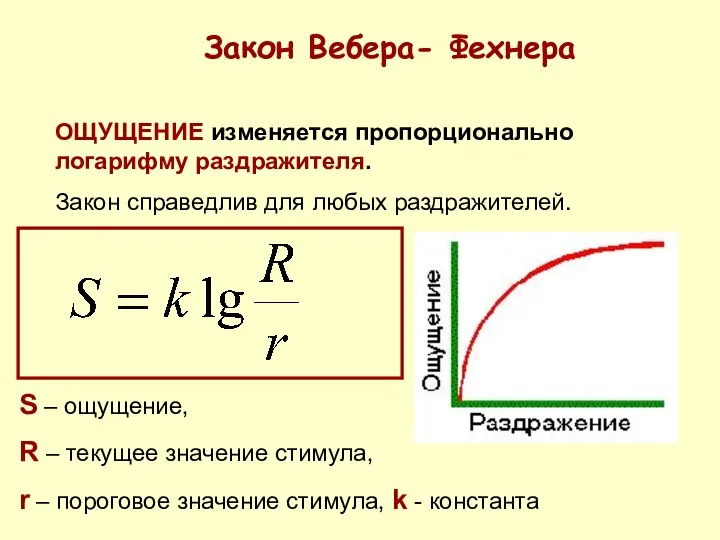
ОЩУЩЕНИЕ изменяется пропорционально логарифму раздражителя.
Закон справедлив для любых раздражителей.
Закон Вебера-
ОЩУЩЕНИЕ изменяется пропорционально логарифму раздражителя.
Закон справедлив для любых раздражителей.
Закон Вебера-

ЗАКОН С. СТИВЕНСА
Интенсивность ощущения описывается степенной функцией
I – интенсивность ощущения
S0 -
ЗАКОН С. СТИВЕНСА
Интенсивность ощущения описывается степенной функцией
I – интенсивность ощущения
S0 -
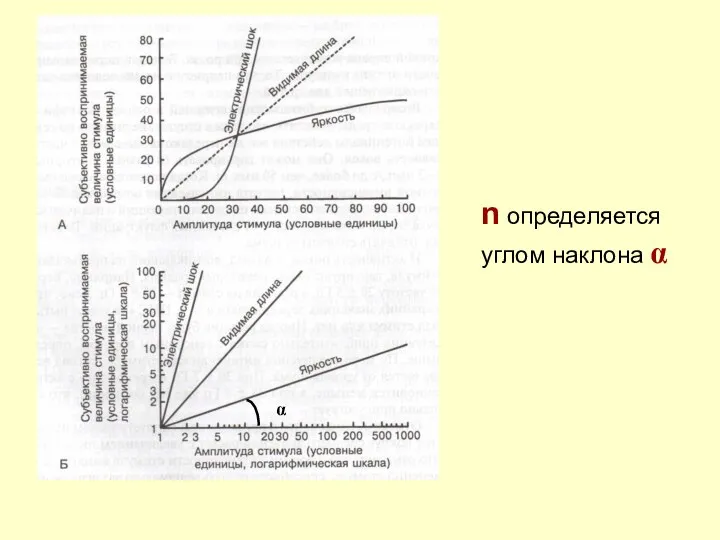
n определяется углом наклона α
n определяется углом наклона α
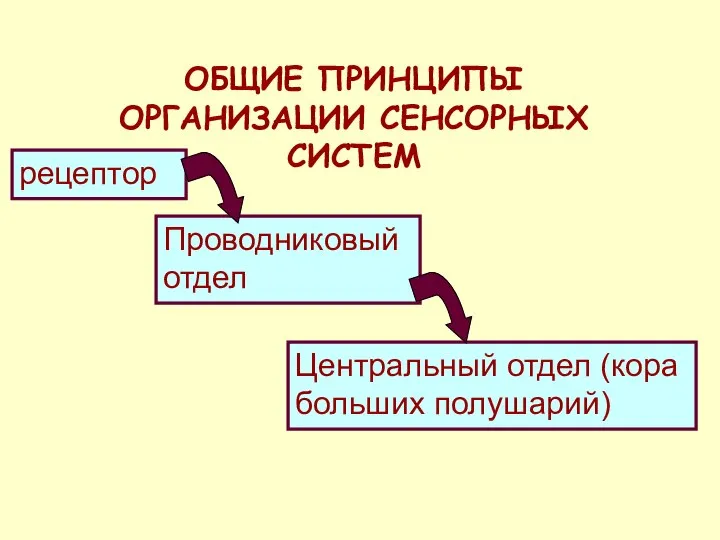
ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ СЕНСОРНЫХ СИСТЕМ
рецептор
Проводниковый отдел
Центральный отдел (кора больших полушарий)
ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ СЕНСОРНЫХ СИСТЕМ
рецептор
Проводниковый отдел
Центральный отдел (кора больших полушарий)
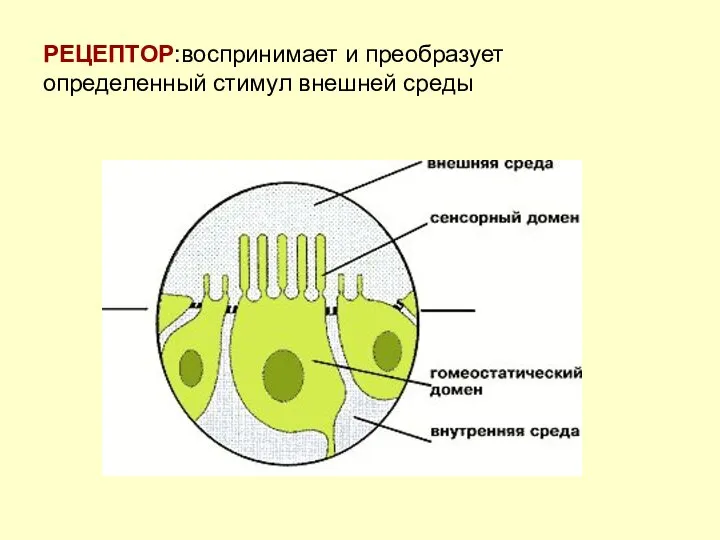
РЕЦЕПТОР:воспринимает и преобразует определенный стимул внешней среды
РЕЦЕПТОР:воспринимает и преобразует определенный стимул внешней среды

МЕХАНИЗМ ПРЕОБРАЗОВАНИЯ В СЕНСОРНОЙ КЛЕТКЕ: ЭНЕРГИЯ ДЕЙСТВУЮЩЕГО СТИМУЛА ПРЕОБРАЗУЕТСЯ В ЭНЕРГИЮ
МЕХАНИЗМ ПРЕОБРАЗОВАНИЯ В СЕНСОРНОЙ КЛЕТКЕ: ЭНЕРГИЯ ДЕЙСТВУЮЩЕГО СТИМУЛА ПРЕОБРАЗУЕТСЯ В ЭНЕРГИЮ
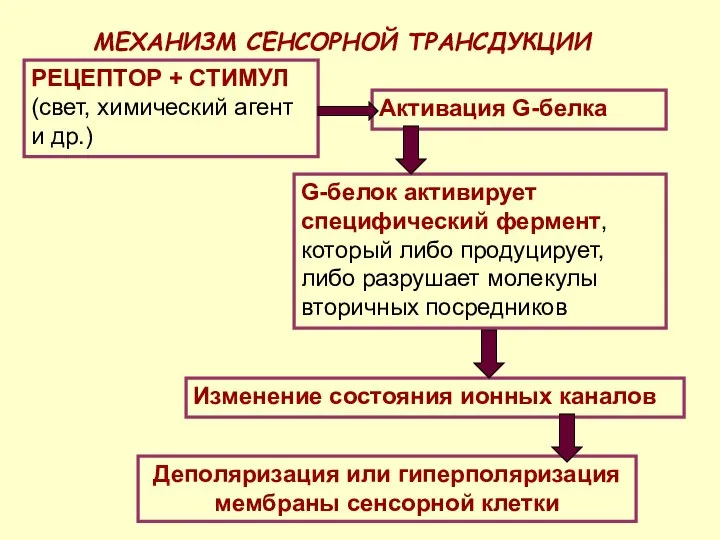
МЕХАНИЗМ СЕНСОРНОЙ ТРАНСДУКЦИИ
РЕЦЕПТОР + СТИМУЛ (свет, химический агент и др.)
Активация G-белка
G-белок
МЕХАНИЗМ СЕНСОРНОЙ ТРАНСДУКЦИИ
РЕЦЕПТОР + СТИМУЛ (свет, химический агент и др.)
Активация G-белка
G-белок
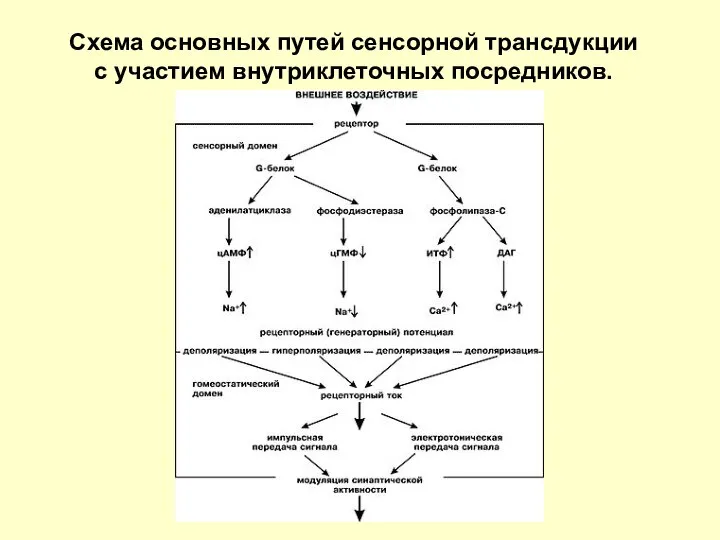
Схема основных путей сенсорной трансдукции
с участием внутриклеточных посредников.
Схема основных путей сенсорной трансдукции
с участием внутриклеточных посредников.

БИОФИЗИКА СЛУХОВОГО АНАЛИЗАТОРА
БИОФИЗИКА СЛУХОВОГО АНАЛИЗАТОРА

АДЕКВАТНЫЙ РАЗДРАЖИТЕЛЬ – ЗВУК
ДИАПАЗОН ВОСПРИНИМАЕМЫХ ЗВУКОВ
от 15 – 16
АДЕКВАТНЫЙ РАЗДРАЖИТЕЛЬ – ЗВУК
ДИАПАЗОН ВОСПРИНИМАЕМЫХ ЗВУКОВ
от 15 – 16
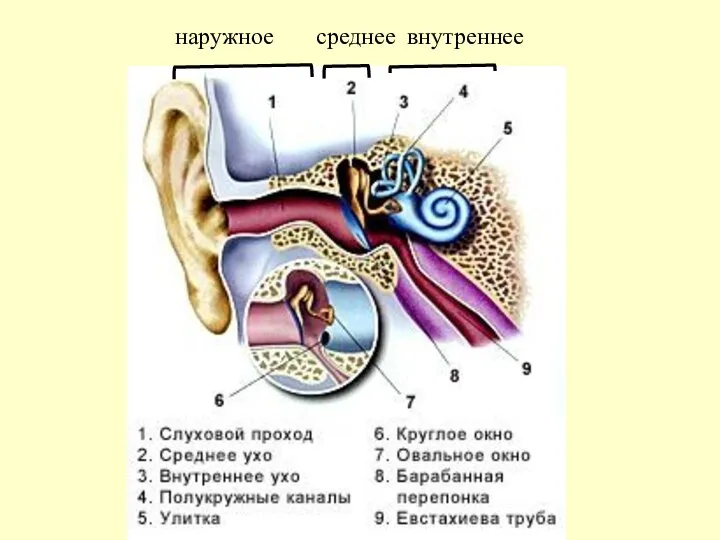
наружное среднее внутреннее
наружное среднее внутреннее

ФУНКЦИИ НАРУЖНОГО УХА
АКУСТИЧЕСКАЯ
ЗАЩИТНАЯ
ЭСТЕТИЧЕСКАЯ
ФУНКЦИИ НАРУЖНОГО УХА
АКУСТИЧЕСКАЯ
ЗАЩИТНАЯ
ЭСТЕТИЧЕСКАЯ

1 – БАРАБАННАЯ ПЕРЕПОНКА
2 – ЕВСТАХИЕВА ТРУБА
3 – 5 СЛУХОВЫЕ КОСТОЧКИ
3
1 – БАРАБАННАЯ ПЕРЕПОНКА
2 – ЕВСТАХИЕВА ТРУБА
3 – 5 СЛУХОВЫЕ КОСТОЧКИ
3

ФУНКЦИИ СРЕДНЕГО УХА
ТРАНСФОРМАЦИЯ ЗВУКОВЫХ КОЛЕБАНИЙ ВОЗДУХА В КОЛЕБАНИЯ ЖИДКОСТИ В УЛИТКЕ
УСИЛЕНИЕ
ФУНКЦИИ СРЕДНЕГО УХА
ТРАНСФОРМАЦИЯ ЗВУКОВЫХ КОЛЕБАНИЙ ВОЗДУХА В КОЛЕБАНИЯ ЖИДКОСТИ В УЛИТКЕ
УСИЛЕНИЕ

УСИЛЕНИЕ СИГНАЛА
ДВА МЕХАНИЗМА:
ПЛОЩАДЬ БАРАБАННОЙ ПЕРЕПОНКИ БОЛЬШЕ ПЛОЩАДИ ОВАЛЬНОГО ОКНА в 14
УСИЛЕНИЕ СИГНАЛА
ДВА МЕХАНИЗМА:
ПЛОЩАДЬ БАРАБАННОЙ ПЕРЕПОНКИ БОЛЬШЕ ПЛОЩАДИ ОВАЛЬНОГО ОКНА в 14
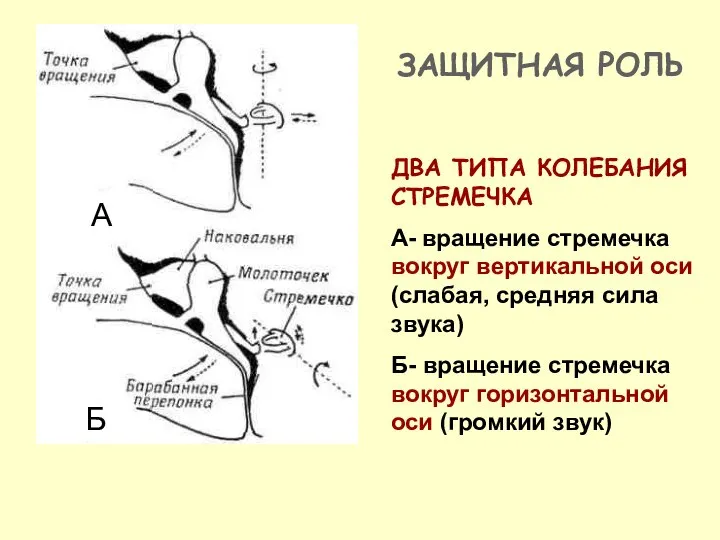
ДВА ТИПА КОЛЕБАНИЯ СТРЕМЕЧКА
А- вращение стремечка вокруг вертикальной оси (слабая, средняя
ДВА ТИПА КОЛЕБАНИЯ СТРЕМЕЧКА
А- вращение стремечка вокруг вертикальной оси (слабая, средняя

БИОФИЗИКА ВНУТРЕННЕГО УХА
БИОФИЗИКА ВНУТРЕННЕГО УХА
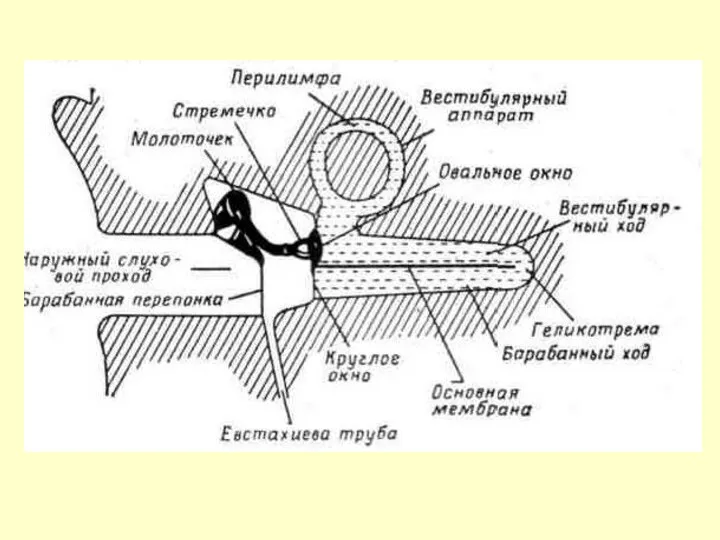

ВЕСТИБУЛЯРНЫЙ И БАРАБАННЫЙ КАНАЛЫ
ЗАПОЛНЕНЫ ПЕРИЛИМФОЙ,
УЛИТКОВЫЙ КАНАЛ – ЭНДОЛИМФОЙ
ПЕРИЛИМФА: 140
ВЕСТИБУЛЯРНЫЙ И БАРАБАННЫЙ КАНАЛЫ
ЗАПОЛНЕНЫ ПЕРИЛИМФОЙ,
УЛИТКОВЫЙ КАНАЛ – ЭНДОЛИМФОЙ
ПЕРИЛИМФА: 140
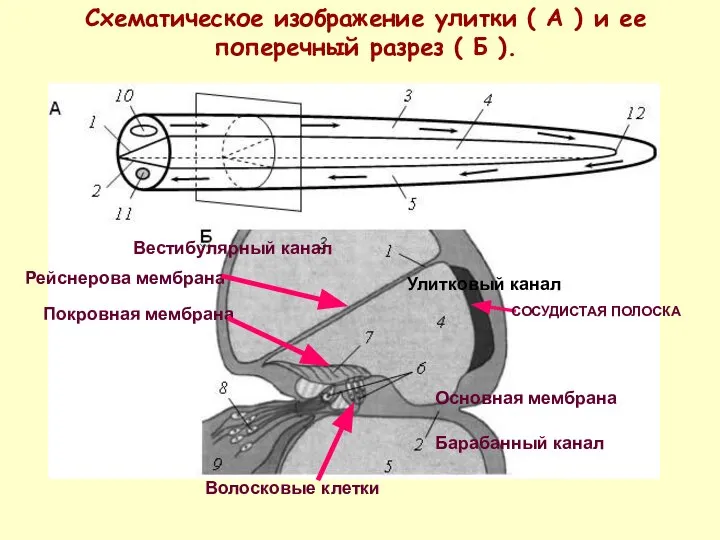
Схематическое изображение улитки ( А ) и ее поперечный разрез (
Схематическое изображение улитки ( А ) и ее поперечный разрез (
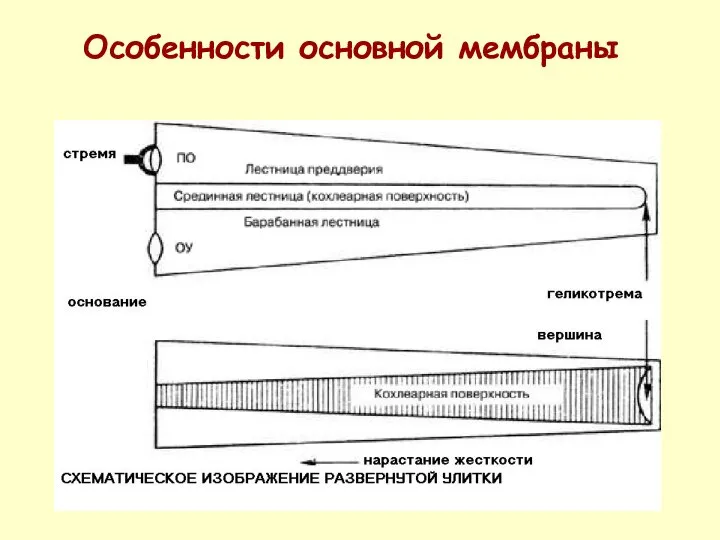
Особенности основной мембраны
Особенности основной мембраны
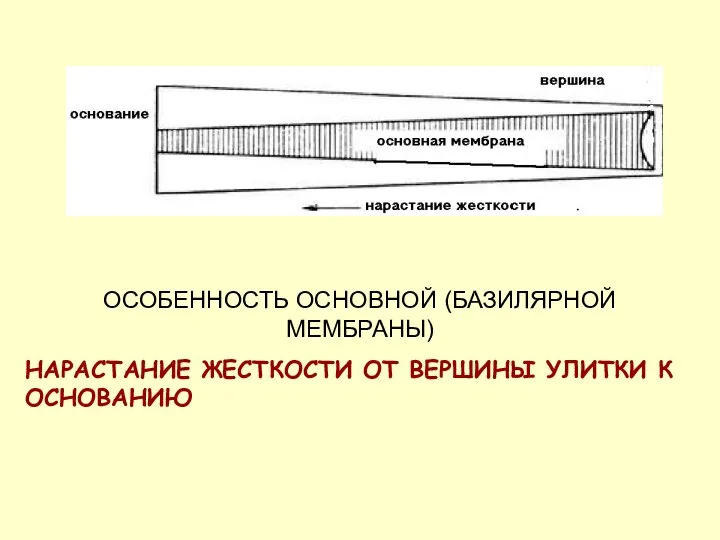
ОСОБЕННОСТЬ ОСНОВНОЙ (БАЗИЛЯРНОЙ МЕМБРАНЫ)
НАРАСТАНИЕ ЖЕСТКОСТИ ОТ ВЕРШИНЫ УЛИТКИ К ОСНОВАНИЮ
ОСОБЕННОСТЬ ОСНОВНОЙ (БАЗИЛЯРНОЙ МЕМБРАНЫ)
НАРАСТАНИЕ ЖЕСТКОСТИ ОТ ВЕРШИНЫ УЛИТКИ К ОСНОВАНИЮ
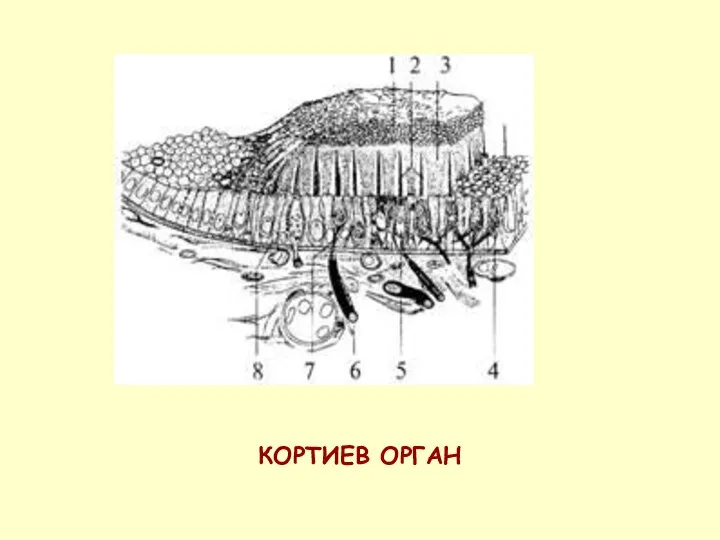
КОРТИЕВ ОРГАН
КОРТИЕВ ОРГАН
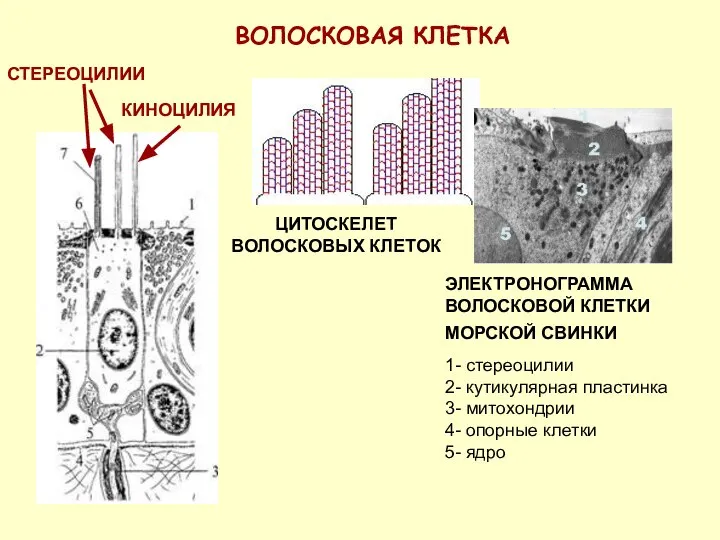
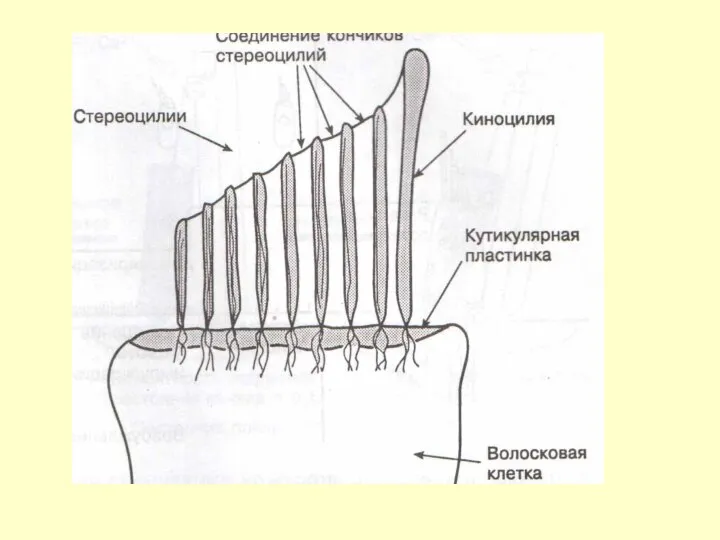
ВОЛОСКОВАЯ КЛЕТКА
СТЕРЕОЦИЛИИ
ЦИТОСКЕЛЕТ ВОЛОСКОВЫХ КЛЕТОК
ЭЛЕКТРОНОГРАММА ВОЛОСКОВОЙ КЛЕТКИ МОРСКОЙ СВИНКИ
1- стереоцилии
2- кутикулярная
ВОЛОСКОВАЯ КЛЕТКА
СТЕРЕОЦИЛИИ
ЦИТОСКЕЛЕТ ВОЛОСКОВЫХ КЛЕТОК
ЭЛЕКТРОНОГРАММА ВОЛОСКОВОЙ КЛЕТКИ МОРСКОЙ СВИНКИ
1- стереоцилии
2- кутикулярная

ТЕОРИИ КОДИРОВАНИЯ ЗВУКА В УЛИТКЕ
РЕЗОНАНСНАЯ ТЕОРИЯ Г.ГЕЛЬМГОЛЬЦА
ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА
ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ
ТЕОРИИ КОДИРОВАНИЯ ЗВУКА В УЛИТКЕ
РЕЗОНАНСНАЯ ТЕОРИЯ Г.ГЕЛЬМГОЛЬЦА
ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА
ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ

Г. ГЕЛЬМГОЛЬЦ
РЕЗОНАНСНАЯ ТЕОРИЯ
ОСНОВНАЯ МЕМБРАНА СОСТОИТ
ИЗ МНОЖЕСТВА НАТЯНУТЫХ
СТРУН, НАСТРОЕННЫХ В
Г. ГЕЛЬМГОЛЬЦ
РЕЗОНАНСНАЯ ТЕОРИЯ
ОСНОВНАЯ МЕМБРАНА СОСТОИТ
ИЗ МНОЖЕСТВА НАТЯНУТЫХ
СТРУН, НАСТРОЕННЫХ В

ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА
ОСНОВНАЯ МЕМБРАНА – ЖЕСТКАЯ ПЛАСТИНКА
ПОСТОЯННЫЙ ТОН ЗАСТАВЛЯЕТ ВСЮ МЕМБРАНУ
ТЕЛЕФОННАЯ ТЕОРИЯ Э.РЕЗЕРФОРДА
ОСНОВНАЯ МЕМБРАНА – ЖЕСТКАЯ ПЛАСТИНКА
ПОСТОЯННЫЙ ТОН ЗАСТАВЛЯЕТ ВСЮ МЕМБРАНУ

Г. БЕКЕШИ
ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ
Нобелевская премия по физиологии и медицине, 1961
Г. БЕКЕШИ
ТЕОРИЯ БЕГУЩЕЙ ВОЛНЫ
Нобелевская премия по физиологии и медицине, 1961
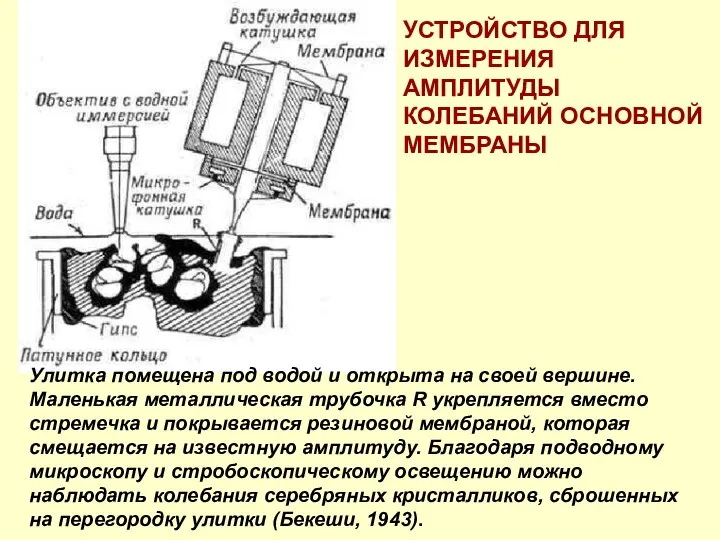
Улитка помещена под водой и открыта на своей вершине. Маленькая металлическая
Улитка помещена под водой и открыта на своей вершине. Маленькая металлическая
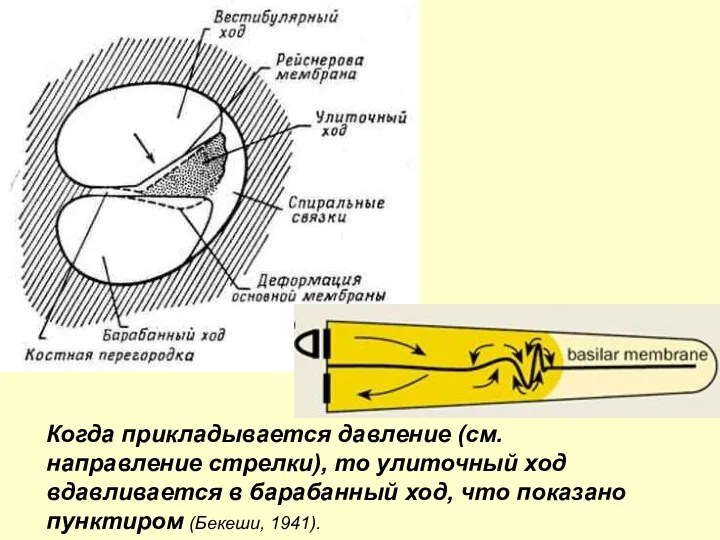
Когда прикладывается давление (см. направление стрелки), то улиточный ход вдавливается в
Когда прикладывается давление (см. направление стрелки), то улиточный ход вдавливается в
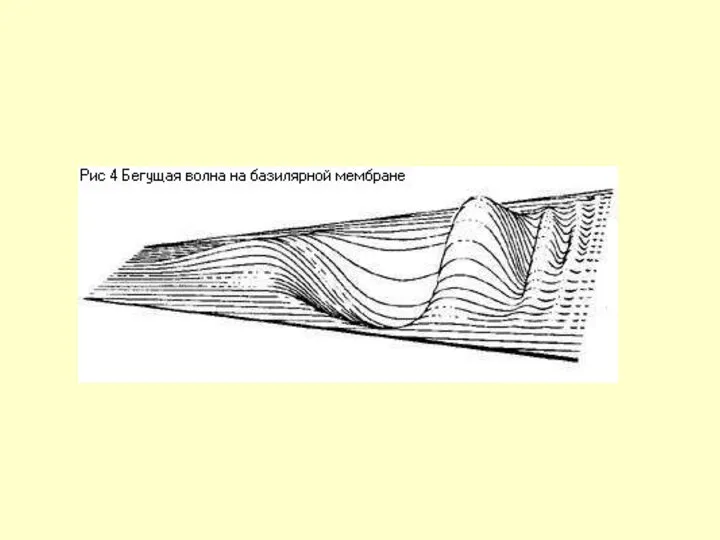
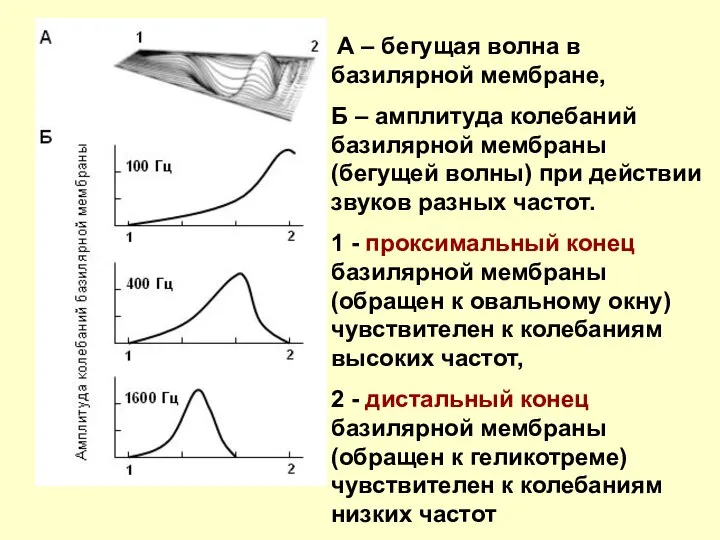
А – бегущая волна в базилярной мембране,
Б – амплитуда
А – бегущая волна в базилярной мембране,
Б – амплитуда
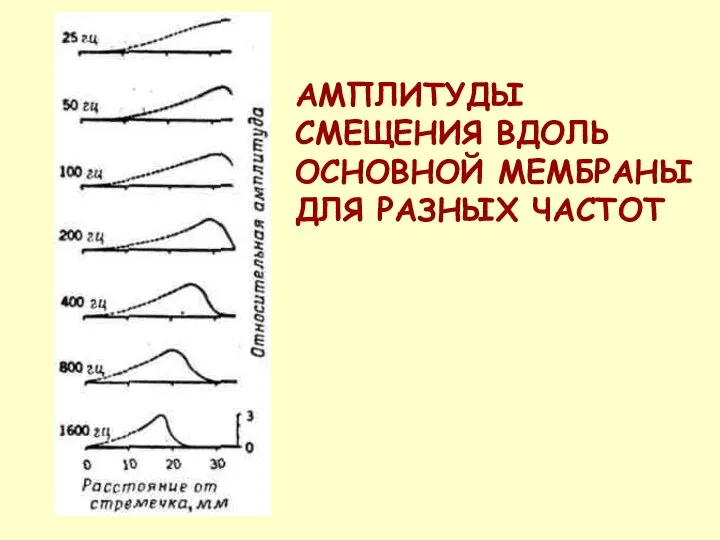
АМПЛИТУДЫ СМЕЩЕНИЯ ВДОЛЬ ОСНОВНОЙ МЕМБРАНЫ ДЛЯ РАЗНЫХ ЧАСТОТ
АМПЛИТУДЫ СМЕЩЕНИЯ ВДОЛЬ ОСНОВНОЙ МЕМБРАНЫ ДЛЯ РАЗНЫХ ЧАСТОТ

ЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ УЛИТКИ
ЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ УЛИТКИ

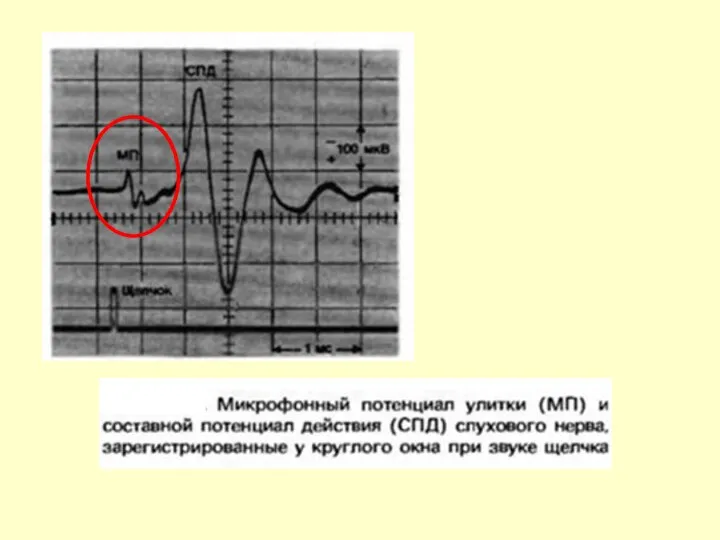
МИКРОФОННЫЙ ПОТЕНЦИАЛ (МФ)
ОБНАРУЖЕН УИВЕРОМ И БРЕЕМ В 1930 ГОДУ.
ПРИЗНАКИ:
НЕ ИМЕЕТ ПОРОГА
ПАРАМЕТРЫ
МИКРОФОННЫЙ ПОТЕНЦИАЛ (МФ)
ОБНАРУЖЕН УИВЕРОМ И БРЕЕМ В 1930 ГОДУ.
ПРИЗНАКИ:
НЕ ИМЕЕТ ПОРОГА
ПАРАМЕТРЫ
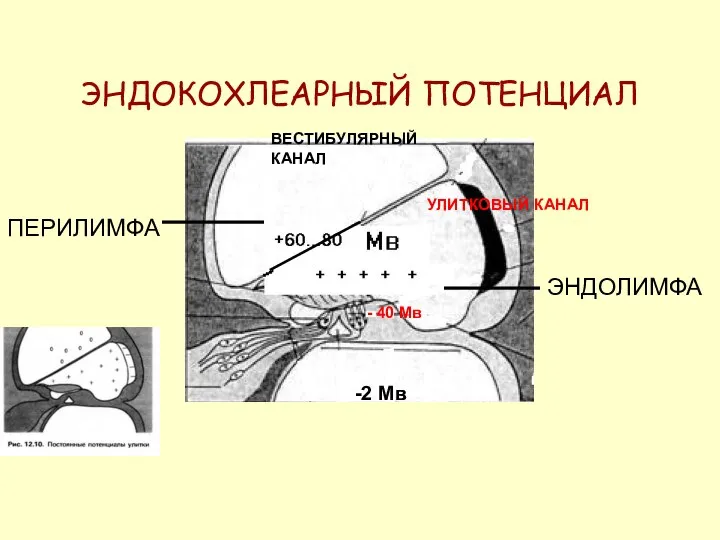
ЭНДОКОХЛЕАРНЫЙ ПОТЕНЦИАЛ
ПЕРИЛИМФА
ЭНДОЛИМФА
+ + +
ВЕСТИБУЛЯРНЫЙ КАНАЛ
УЛИТКОВЫЙ КАНАЛ
-2 Мв
- 40 Мв
ЭНДОКОХЛЕАРНЫЙ ПОТЕНЦИАЛ
ПЕРИЛИМФА
ЭНДОЛИМФА
+ + +
ВЕСТИБУЛЯРНЫЙ КАНАЛ
УЛИТКОВЫЙ КАНАЛ
-2 Мв
- 40 Мв
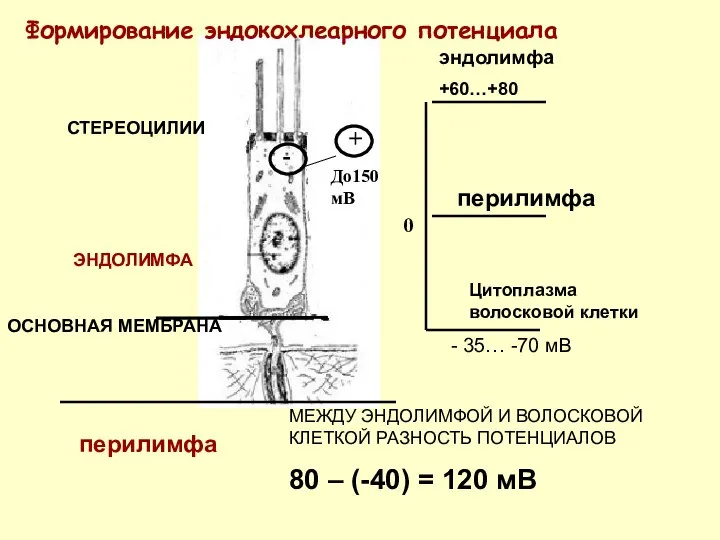
ОСНОВНАЯ МЕМБРАНА
ЭНДОЛИМФА
СТЕРЕОЦИЛИИ
-
+
До150 мВ
эндолимфа
+60…+80
перилимфа
0
Цитоплазма волосковой клетки
- 35… -70 мВ
перилимфа
Формирование эндокохлеарного потенциала
МЕЖДУ ЭНДОЛИМФОЙ
ОСНОВНАЯ МЕМБРАНА
ЭНДОЛИМФА
СТЕРЕОЦИЛИИ
-
+
До150 мВ
эндолимфа
+60…+80
перилимфа
0
Цитоплазма волосковой клетки
- 35… -70 мВ
перилимфа
Формирование эндокохлеарного потенциала
МЕЖДУ ЭНДОЛИМФОЙ
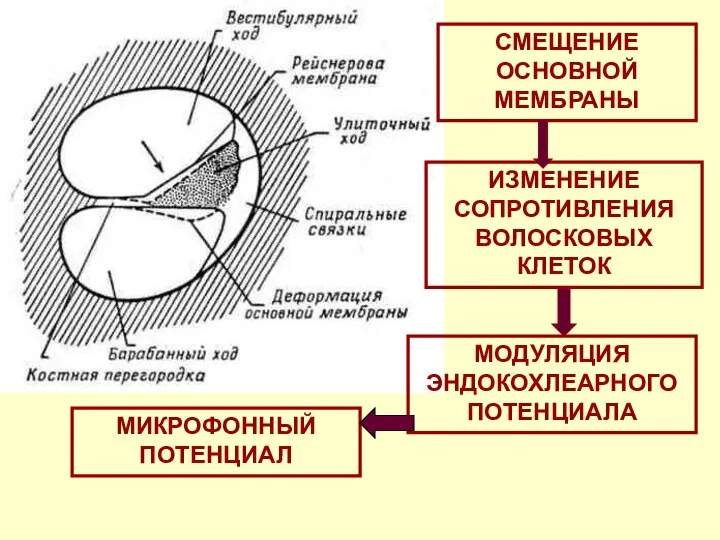
СМЕЩЕНИЕ ОСНОВНОЙ МЕМБРАНЫ
МОДУЛЯЦИЯ ЭНДОКОХЛЕАРНОГО ПОТЕНЦИАЛА
ИЗМЕНЕНИЕ СОПРОТИВЛЕНИЯ ВОЛОСКОВЫХ КЛЕТОК
МИКРОФОННЫЙ ПОТЕНЦИАЛ
СМЕЩЕНИЕ ОСНОВНОЙ МЕМБРАНЫ
МОДУЛЯЦИЯ ЭНДОКОХЛЕАРНОГО ПОТЕНЦИАЛА
ИЗМЕНЕНИЕ СОПРОТИВЛЕНИЯ ВОЛОСКОВЫХ КЛЕТОК
МИКРОФОННЫЙ ПОТЕНЦИАЛ
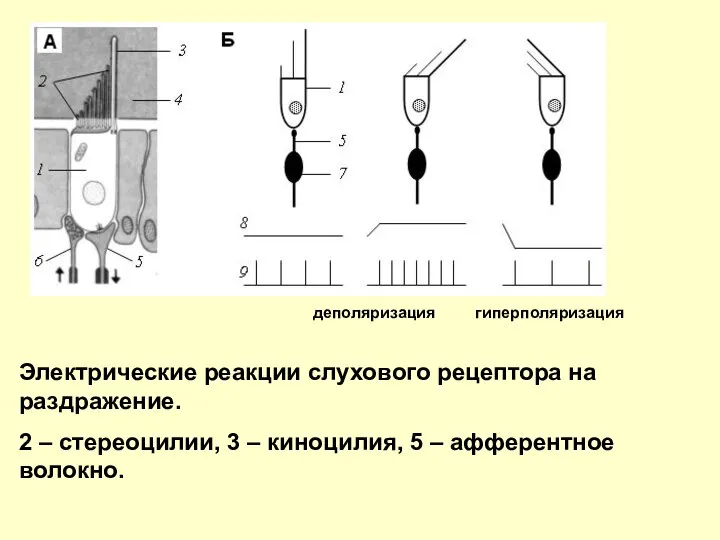
Электрические реакции слухового рецептора на раздражение.
2 – стереоцилии, 3 – киноцилия,
Электрические реакции слухового рецептора на раздражение.
2 – стереоцилии, 3 – киноцилия,
БИОФИЗИКА ЗРИТЕЛЬНОГО АНАЛИЗАТОРА
БИОФИЗИКА ЗРИТЕЛЬНОГО АНАЛИЗАТОРА

ВИДИМЫЙ СВЕТ ОТ 400 ДО 750 НМ
ЗРЕНИЕ БАЗИРУЕТСЯ НА ВОСПРИЯТИИ
КОНТРАСТОВ
ВИДИМЫЙ СВЕТ ОТ 400 ДО 750 НМ
ЗРЕНИЕ БАЗИРУЕТСЯ НА ВОСПРИЯТИИ
КОНТРАСТОВ
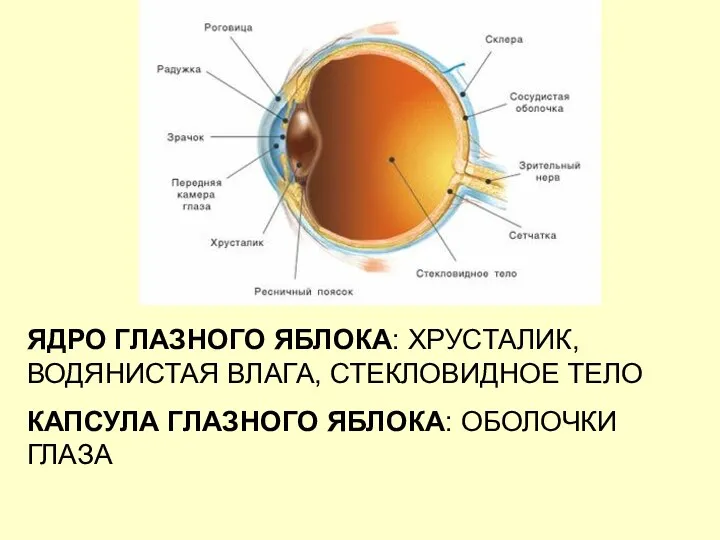
ЯДРО ГЛАЗНОГО ЯБЛОКА: ХРУСТАЛИК, ВОДЯНИСТАЯ ВЛАГА, СТЕКЛОВИДНОЕ ТЕЛО
КАПСУЛА ГЛАЗНОГО ЯБЛОКА: ОБОЛОЧКИ
ЯДРО ГЛАЗНОГО ЯБЛОКА: ХРУСТАЛИК, ВОДЯНИСТАЯ ВЛАГА, СТЕКЛОВИДНОЕ ТЕЛО
КАПСУЛА ГЛАЗНОГО ЯБЛОКА: ОБОЛОЧКИ

ЯДРО ГЛАЗНОГО ЯБЛОКА
ХРУСТАЛИК:
ДВОЯКОВЫПКУЛАЯ ЛИНЗА
ПРОЗРАЧНЫЙ И БЕСЦВЕТНЫЙ
НАХОДИТСЯ В ПРОЗРАЧНОЙ КАПСУЛЕ
ВОЛОКНА ХРУСТАЛИКА
ЯДРО ГЛАЗНОГО ЯБЛОКА
ХРУСТАЛИК:
ДВОЯКОВЫПКУЛАЯ ЛИНЗА
ПРОЗРАЧНЫЙ И БЕСЦВЕТНЫЙ
НАХОДИТСЯ В ПРОЗРАЧНОЙ КАПСУЛЕ
ВОЛОКНА ХРУСТАЛИКА

КАПСУЛА ГЛАЗНОГО ЯБЛОКА
БЕЛОЧНАЯ ОБОЛОЧКА: СКЛЕРА + РОГОВИЦА
СОСУДИСТАЯ ОБОЛОЧКА: СОБСТВЕННО СОСУДИСТАЯ ОБОЛОЧКА
КАПСУЛА ГЛАЗНОГО ЯБЛОКА
БЕЛОЧНАЯ ОБОЛОЧКА: СКЛЕРА + РОГОВИЦА
СОСУДИСТАЯ ОБОЛОЧКА: СОБСТВЕННО СОСУДИСТАЯ ОБОЛОЧКА

ФОРМИРОВАНИЕ ИЗОБРАЖЕНИЯ НА СЕТЧАТКЕ
ФОРМИРОВАНИЕ ИЗОБРАЖЕНИЯ НА СЕТЧАТКЕ
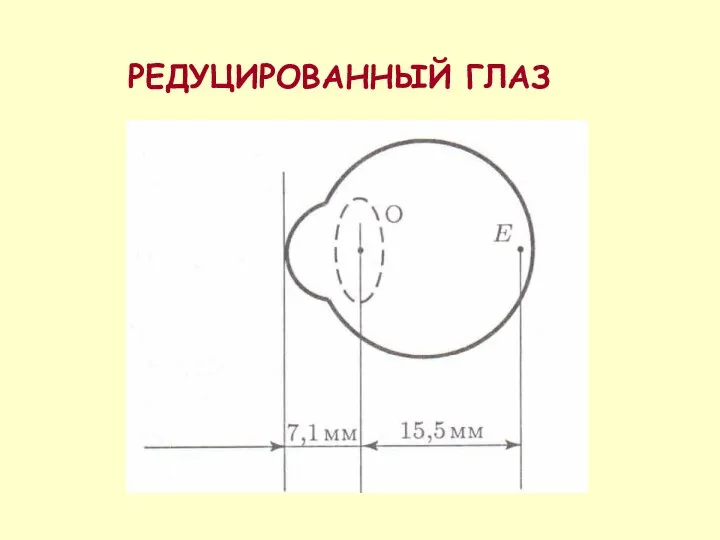
РЕДУЦИРОВАННЫЙ ГЛАЗ
РЕДУЦИРОВАННЫЙ ГЛАЗ
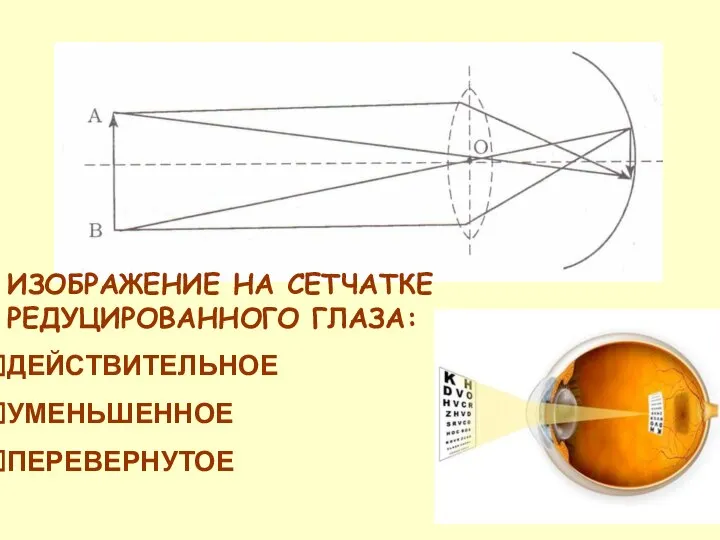
ИЗОБРАЖЕНИЕ НА СЕТЧАТКЕ РЕДУЦИРОВАННОГО ГЛАЗА:
ДЕЙСТВИТЕЛЬНОЕ
УМЕНЬШЕННОЕ
ПЕРЕВЕРНУТОЕ
ИЗОБРАЖЕНИЕ НА СЕТЧАТКЕ РЕДУЦИРОВАННОГО ГЛАЗА:
ДЕЙСТВИТЕЛЬНОЕ
УМЕНЬШЕННОЕ
ПЕРЕВЕРНУТОЕ

СТРОЕНИЕ СЕТЧАТКИ
СТРОЕНИЕ СЕТЧАТКИ
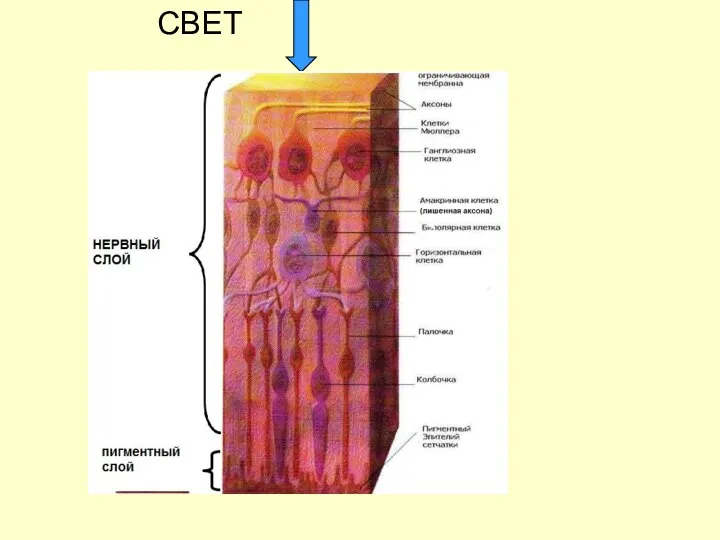
СВЕТ
СВЕТ

ФУНКЦИИ ПИГМЕНТНОГО СЛОЯ:
ПИТАНИЕ ДЛЯ ФОТОРЕЦЕПТОРОВ
ОБНОВЛЕНИЕ ФОТОРЕЦЕПТОРОВ
ПОГЛОЩЕНИЕ СВЕТА, НЕ ПОГЛОТИВШЕГОСЯ ФОТОРЕЦЕПТОРАМИ
ФУНКЦИЯ СЕТЧАТКИ:
ВОСПРИЯТИЕ
ФУНКЦИИ ПИГМЕНТНОГО СЛОЯ:
ПИТАНИЕ ДЛЯ ФОТОРЕЦЕПТОРОВ
ОБНОВЛЕНИЕ ФОТОРЕЦЕПТОРОВ
ПОГЛОЩЕНИЕ СВЕТА, НЕ ПОГЛОТИВШЕГОСЯ ФОТОРЕЦЕПТОРАМИ
ФУНКЦИЯ СЕТЧАТКИ:
ВОСПРИЯТИЕ
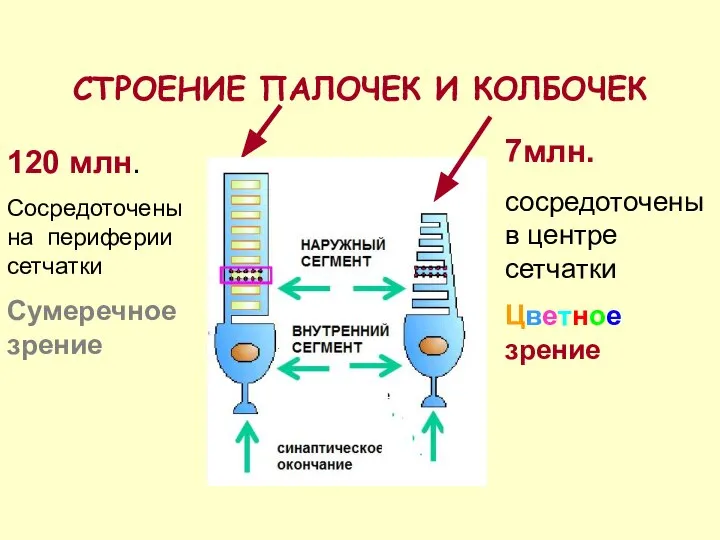
СТРОЕНИЕ ПАЛОЧЕК И КОЛБОЧЕК
7млн.
сосредоточены в центре сетчатки
Цветное зрение
120 млн.
Сосредоточены на
СТРОЕНИЕ ПАЛОЧЕК И КОЛБОЧЕК
7млн.
сосредоточены в центре сетчатки
Цветное зрение
120 млн.
Сосредоточены на
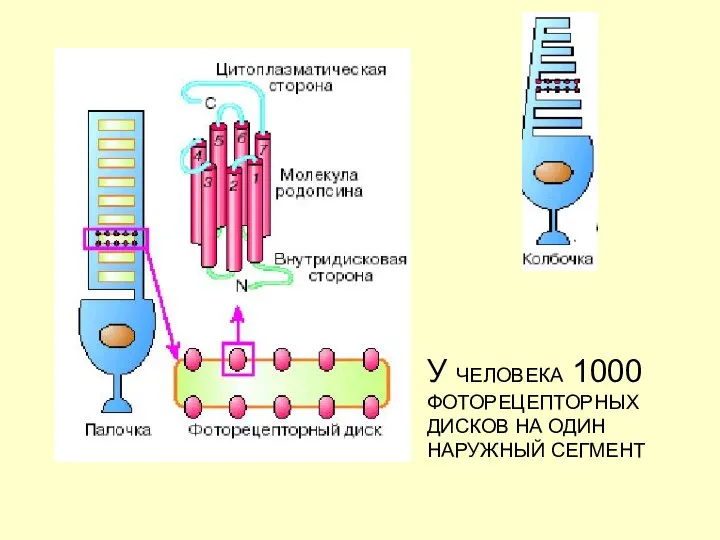
У ЧЕЛОВЕКА 1000 ФОТОРЕЦЕПТОРНЫХ ДИСКОВ НА ОДИН НАРУЖНЫЙ СЕГМЕНТ
У ЧЕЛОВЕКА 1000 ФОТОРЕЦЕПТОРНЫХ ДИСКОВ НА ОДИН НАРУЖНЫЙ СЕГМЕНТ

СОСТАВ ФОТОРЕЦЕПТОРНЫХ ДИСКОВ
БЕЛКИ – ОКОЛО 60%, ЛИПИДЫ – ОКОЛО 40%, УГЛЕВОДЫ
СОСТАВ ФОТОРЕЦЕПТОРНЫХ ДИСКОВ
БЕЛКИ – ОКОЛО 60%, ЛИПИДЫ – ОКОЛО 40%, УГЛЕВОДЫ

БЕЛКОВЫЙ СОСТАВ
ИНТЕГРАЛЬНЫЕ МЕМБРАННЫЕ: РОДОПСИН (95% )
ЭКСТРАГИРУЕМЫЕ:
G-БЕЛОК ТРАНСДУЦИН
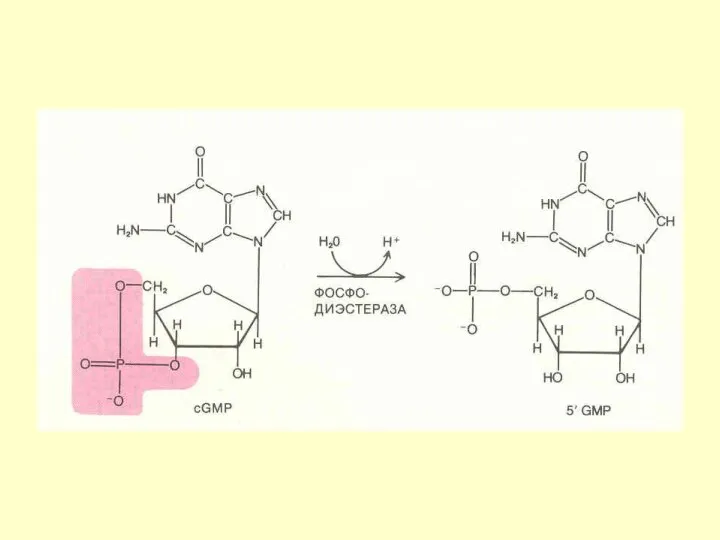
ФОСФОДИЭСТЕРАЗА цГМФ
РОДОПСИНКИНАЗА
ГУАНИЛАТЦИКЛАЗА
БЕЛКОВЫЙ СОСТАВ
ИНТЕГРАЛЬНЫЕ МЕМБРАННЫЕ: РОДОПСИН (95% )
ЭКСТРАГИРУЕМЫЕ:
G-БЕЛОК ТРАНСДУЦИН
ФОСФОДИЭСТЕРАЗА цГМФ
РОДОПСИНКИНАЗА
ГУАНИЛАТЦИКЛАЗА

ЭЛЕКТРОФИЗИОЛОГИЯ ФОТОРЕЦЕПТОРОВ
ЭЛЕКТРОФИЗИОЛОГИЯ ФОТОРЕЦЕПТОРОВ
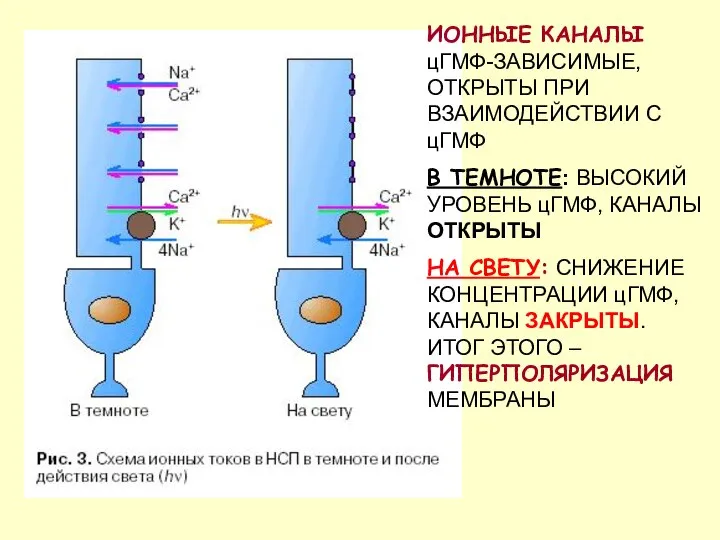
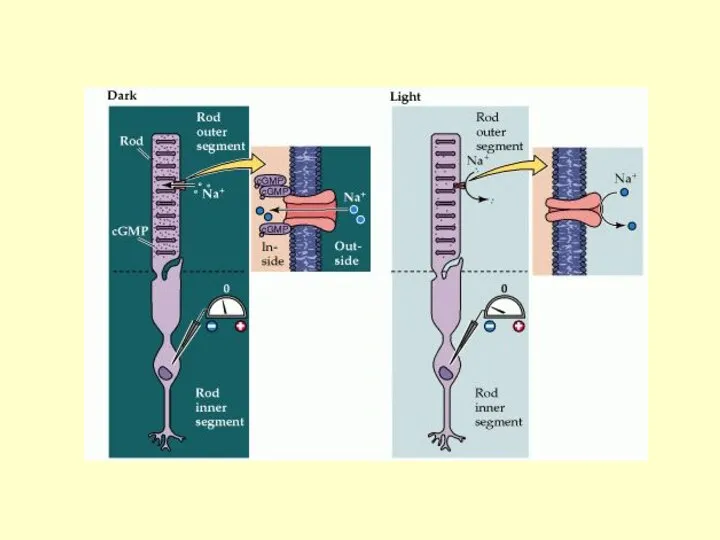
ИОННЫЕ КАНАЛЫ цГМФ-ЗАВИСИМЫЕ, ОТКРЫТЫ ПРИ ВЗАИМОДЕЙСТВИИ С цГМФ
В ТЕМНОТЕ: ВЫСОКИЙ УРОВЕНЬ
ИОННЫЕ КАНАЛЫ цГМФ-ЗАВИСИМЫЕ, ОТКРЫТЫ ПРИ ВЗАИМОДЕЙСТВИИ С цГМФ
В ТЕМНОТЕ: ВЫСОКИЙ УРОВЕНЬ

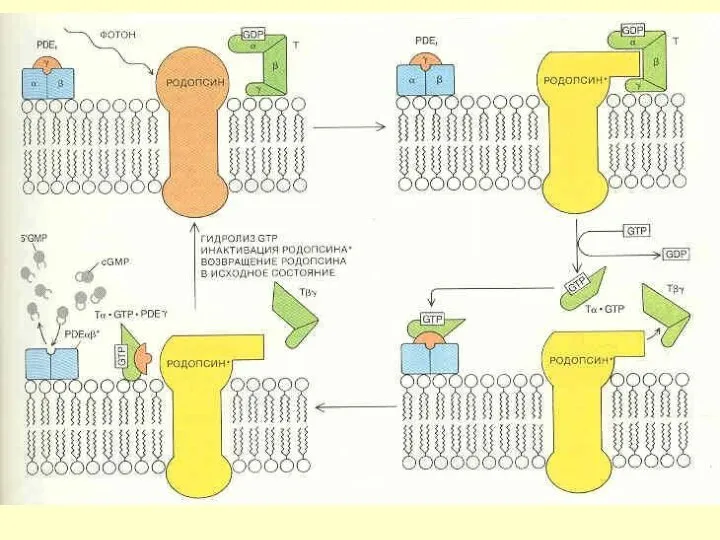
БЕЛКИ ЗРИТЕЛЬНОГО КАСКАДА
РОДОПСИН
G-белок ТРАНСДУЦИН
ЦГМФ-ФОСФОДИЭСТЕРАЗА
БЕЛКИ ЗРИТЕЛЬНОГО КАСКАДА
РОДОПСИН
G-белок ТРАНСДУЦИН
ЦГМФ-ФОСФОДИЭСТЕРАЗА
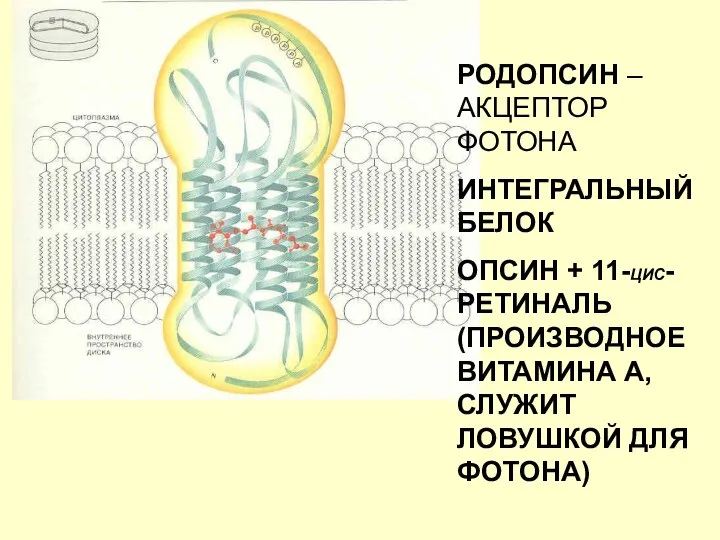
РОДОПСИН – АКЦЕПТОР ФОТОНА
ИНТЕГРАЛЬНЫЙ БЕЛОК
ОПСИН + 11-ЦИС-РЕТИНАЛЬ (ПРОИЗВОДНОЕ ВИТАМИНА А, СЛУЖИТ
РОДОПСИН – АКЦЕПТОР ФОТОНА
ИНТЕГРАЛЬНЫЙ БЕЛОК
ОПСИН + 11-ЦИС-РЕТИНАЛЬ (ПРОИЗВОДНОЕ ВИТАМИНА А, СЛУЖИТ
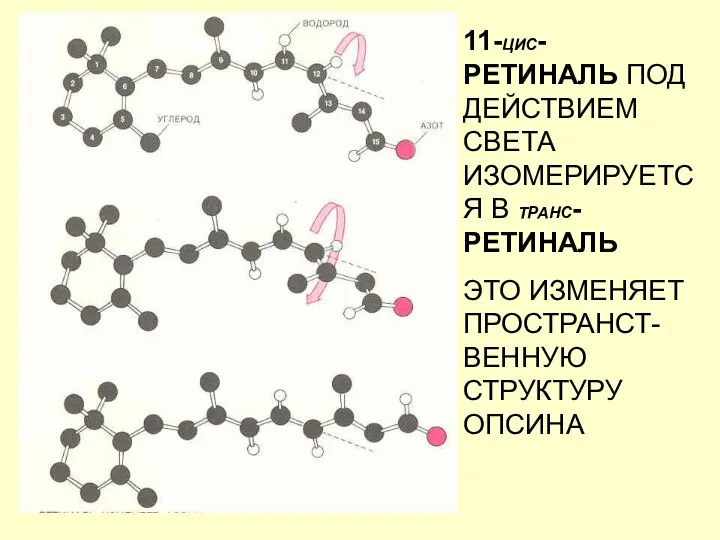
11-ЦИС-РЕТИНАЛЬ ПОД ДЕЙСТВИЕМ СВЕТА ИЗОМЕРИРУЕТСЯ В ТРАНС-РЕТИНАЛЬ
ЭТО ИЗМЕНЯЕТ ПРОСТРАНСТ-ВЕННУЮ СТРУКТУРУ ОПСИНА
11-ЦИС-РЕТИНАЛЬ ПОД ДЕЙСТВИЕМ СВЕТА ИЗОМЕРИРУЕТСЯ В ТРАНС-РЕТИНАЛЬ
ЭТО ИЗМЕНЯЕТ ПРОСТРАНСТ-ВЕННУЮ СТРУКТУРУ ОПСИНА

НАДЕЮСЬ, ЧТО КУРС ЛЕКЦИЙ ПО БИОФИЗИКЕ ПОМОЖЕТ В ВАШЕЙ ПРОФЕССИОНАЛЬНОЙ ДЕЯТЕЛЬНОСТИ
НАДЕЮСЬ, ЧТО КУРС ЛЕКЦИЙ ПО БИОФИЗИКЕ ПОМОЖЕТ В ВАШЕЙ ПРОФЕССИОНАЛЬНОЙ ДЕЯТЕЛЬНОСТИ
 Звукозапись. Носители звукозаписи
Звукозапись. Носители звукозаписи Система управления рисками
Система управления рисками  Интимный гель для женщин содержит L-аргинин способствует образованию оксида азота восстанавливает жизненно-важные функции органи
Интимный гель для женщин содержит L-аргинин способствует образованию оксида азота восстанавливает жизненно-важные функции органи Риск выбора и надежности внешнеторгового контрагента
Риск выбора и надежности внешнеторгового контрагента Документоведение и делопроизводство
Документоведение и делопроизводство Система команд МП х86
Система команд МП х86 Презентация Цена и общая стоимость контракта Виды цен Определение валюты цены
Презентация Цена и общая стоимость контракта Виды цен Определение валюты цены  Власть. Роль политики в жизни общества
Власть. Роль политики в жизни общества Особенности планирования и организации уроков рисования. Основы цветоведения. Методика ознакомления с живописью
Особенности планирования и организации уроков рисования. Основы цветоведения. Методика ознакомления с живописью Эпидемический паротит
Эпидемический паротит  Обработка ошибок
Обработка ошибок Лев Николаевич Толстой (1828 – 1910)
Лев Николаевич Толстой (1828 – 1910) Изображение и обозначение резьбы
Изображение и обозначение резьбы Эвристические комбинаторные алгоритмы
Эвристические комбинаторные алгоритмы Установка и стабилизация управляемых колес
Установка и стабилизация управляемых колес Уход за больными пожилого возраста
Уход за больными пожилого возраста География туризма
География туризма История развития физической культуры
История развития физической культуры BUENA ONDA Интернет портали за Туризъм & Резервации на хотели Здравей, Свят!
BUENA ONDA Интернет портали за Туризъм & Резервации на хотели Здравей, Свят! Универсальный спортивный комплекс «Победа»
Универсальный спортивный комплекс «Победа» История развития компьютерной техники (первое поколение ЭВМ)
История развития компьютерной техники (первое поколение ЭВМ) Модели деловой карьеры менеджера
Модели деловой карьеры менеджера Ренуар, Пьер Огюст (25 февраля 1841-2 декабря 1919)
Ренуар, Пьер Огюст (25 февраля 1841-2 декабря 1919) Имя существительное
Имя существительное Физиологические и биохимические основы физической культуры
Физиологические и биохимические основы физической культуры Схема климат. установки ПАНАСОНИК
Схема климат. установки ПАНАСОНИК Жиындар (массивтер
Жиындар (массивтер Travlling. sightseeings in Kazakhstan and the Uk
Travlling. sightseeings in Kazakhstan and the Uk