- Биохимия крови

Содержание
- 2. Внутренняя среда организма формируется совокупностью биологических жидкостей (кровь, лимфа, тканевая жидкость), омывающих клетки и структуры тканей.
- 4. Химический состав плазмы
- 5. Функции крови Дыхательная -транспорт кислорода от легких к тканям и CO2 от тканей к легким Выделительная
- 6. Химический состав крови Большую часть этих функций выполняют компоненты плазмы крови. Плазма крови состоит на 90-93%
- 7. Характеристика белков плазмы крови содержатся в плазме крови синтезируются в печени или РЭС (реже в специализированных
- 8. Состав белков плазмы крови В плазме обнаружено более 100 разных белков соответствующих этим критериям, содержание которых
- 9. Электрофорез белков плазмы
- 10. Белковые фракции
- 11. Функциональная классификация белков плазмы Транспортная а. специф. Б. неспециф Резервная ~ 50% альбумина Регуляторная колл-осм давление
- 12. Остаточный азот Все азотсодержащие вещества плазмы образуют общий пул азота, состоящий из: Азота белкового – осаждаемого
- 13. Состав ОА Мочевина - 50% (главный компонент) АК - 25% (~ 10% ГЛУ и ГЛН) Ураты
- 14. Ds значение ОА Уровень ОА зависит от: Интенсивности катаболизма Травмы (ожоги, краш-синдром) Распад тканей (tbc, c-r,
- 15. Азотемия - повышение уровня ОА в крови Ретенционная – задержка компонентов ОА в организме из-за нарушения
- 16. Общие понятия КОС КОС – система гомеостаза рН внутри- и внеклеточной среды организма. Единицы измерения :рН
- 17. рН – производное метаболизма За сутки организм hs образует 50-100 мМ [Н+] на 15-20л ВКЖ. Весь
- 18. Стабильный рН - необходимое условие метаболизма Изменение рН приводит к изменению: заряда и функции белков (ферментов,
- 19. Оптимум рН разных ферментов
- 20. Изоосмолярность – осм. давление=310 осМ/л - const любые изменения должны поддерживать эту константу Электронейтральность – (по
- 21. Диаграмма Гэмбла
- 22. Механизмы регуляции КОС Физико-хим – действует в автоматическом режиме и представлен: разбавлением т.е. выходом Н+ или
- 23. Классификация нарушений КОС рНО =7.40±0.04 рН = 7.35 и ниже – ацидоз рН = 7.45 и
- 24. Механизм развития респираторных нарушений КОС ацидоз СО2 + Н2О Н2СО3 Н++ НСО3- Алкалоз Причины: изменение частоты
- 25. Межорганное взаимодействие в регуляции рН Если этих респираторных механизмов недостаточно, то активируются др.экскреторные системы. В печени
- 29. Эритропоэтин (Эпо) Эпо – цитокин, специфический регулятор эритропоэза в костном мозге Эпо человека – гликопротеид, состоит
- 30. Э общий обзор Кол-во Э у мужчин - 4.6-6.2 млн/мкл крови, а у женщин - 4.2-5.4
- 31. Цитоскелет Э
- 32. Белки цитоскелета Э α-спектрин Спектрин Анкирин Полоса 3 Полоса 4.1 Полоса 4.2 Полоса 4.9 Актин
- 33. Белки Э 1. Часть белков Э явл общими для мембран и цитоскелета: Спектрин Анкирин Актин Фракция
- 34. Структура цитоскелета Э Большинство мембран Э - интегральные Б, гликопротеиды. Б без углеводной части находятся на
- 35. Метаболизм глюкозы в Э Гликолиз (90-95%) – образование АТФ ПФП (10-5%) - образование NADPH (АОЗ) Особенностью
- 36. Особенности метаболизма Э (шунт Раппопорта)
- 37. Мет Hb редуктазная система Э Hb(Fe2+) Met-Hb(Fe3+) 2 G-SH G-S-S-G NAD(P)H + H+ NAD(P)+ Мет Hb
- 38. Синтез порфобилиногена и гема Первая реакция б/с гема происходит в Мх и происходит путем конденсации гли
- 40. Синтез порфобилиногена и гема (прод) Затем следует этап конденсации (голова-хвост) 4 молекул порфобилиногена с образованием линейного
- 41. Б/с гема
- 42. Б/с гема (прод)
- 43. SH-содержащие ферменты - Феррохелатаза, синтаза дАЛК и дАЛК дегидратаза высокочувствительны к действию тяжелых металлов Характерный признак
- 44. Экспрессии субъединиц глобина
- 45. Структура Hb
- 46. Кривая диссоциации HbО2
- 47. Распад гема Продолжительность жизни Э ~120 дней, ежедневный оборот Hb ~6 г/день. Гем из старых Э
- 48. Образование билирубина Следующий этап СН2 мост (между кольцами III и IV) биливердина восстанавливается биливердин редуктазой до
- 49. Деградация гема M= Метил, P=Пропионил V=Винил
- 50. Билирубин-диглюкуронид В гепатоцитах УДФ-глюкуронил трансфераза присоединяет 2 остатка ГК к билирубину с образованием гидрофильного билирубина-диглюкуронида, что
- 51. Клинические аспекты метаболизма гема представлены в виде: Дефектов ферментов и нарушения б/с гема – порфирий, которые
- 52. Желтухи Гипербилирубинемия проявляется в виде желтух – желтой пигментации кожи и слизистых. В норме в кишечнике
- 53. Нейтрофилы (гранулоциты) –полиморфноядерные лейкоциты: обладают выраженной способностью к фагоцитозу, Хорошо развитый, подвижный цитоскелет, Активное Мх и
- 54. 4O2 2O2 H+ 2H2O2 2H2O + O2 (pH↓) Цитоплазма 4e- Вакуоль 4K+ и/или 4H+ (pHнейтр) (pH↑)
- 56. Скачать презентацию

Внутренняя среда организма формируется совокупностью биологических жидкостей (кровь, лимфа, тканевая
Внутренняя среда организма формируется совокупностью биологических жидкостей (кровь, лимфа, тканевая

Химический состав плазмы
Химический состав плазмы

Функции крови
Дыхательная -транспорт кислорода от легких к тканям и CO2 от
Функции крови
Дыхательная -транспорт кислорода от легких к тканям и CO2 от

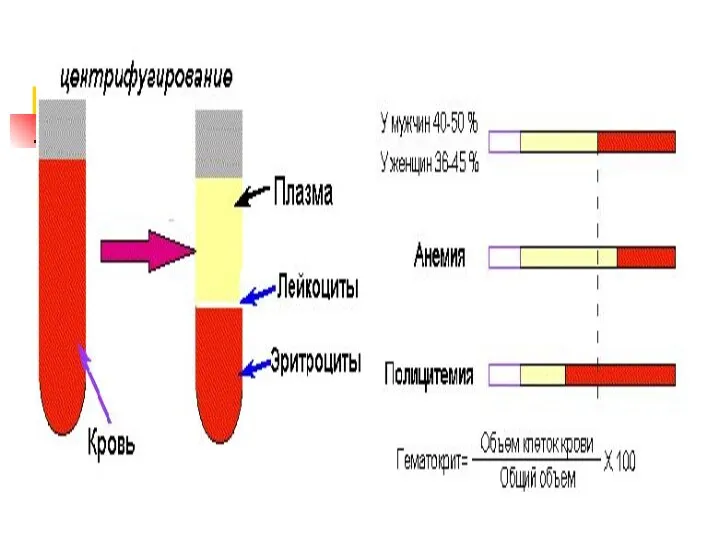
Химический состав крови
Большую часть этих функций выполняют компоненты плазмы крови.
Плазма
Химический состав крови
Большую часть этих функций выполняют компоненты плазмы крови.
Плазма

Характеристика белков плазмы крови
содержатся в плазме крови
синтезируются в печени или РЭС
Характеристика белков плазмы крови
содержатся в плазме крови
синтезируются в печени или РЭС

Состав белков плазмы крови
В плазме обнаружено более 100 разных белков соответствующих
Состав белков плазмы крови
В плазме обнаружено более 100 разных белков соответствующих
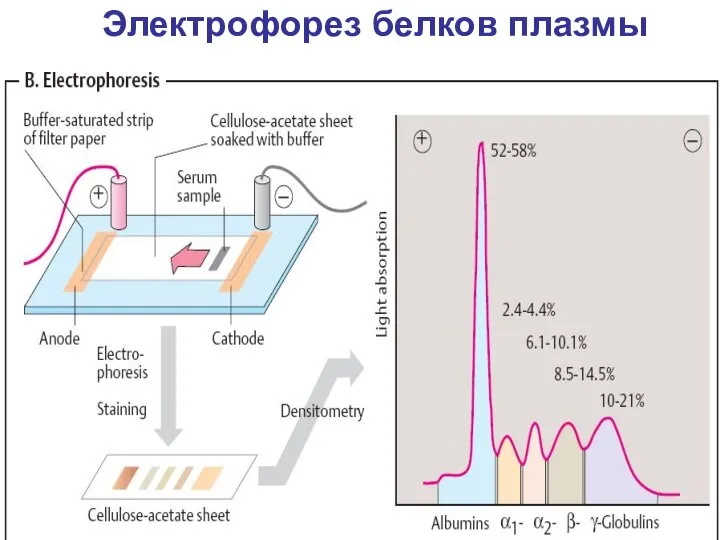
Электрофорез белков плазмы
Электрофорез белков плазмы
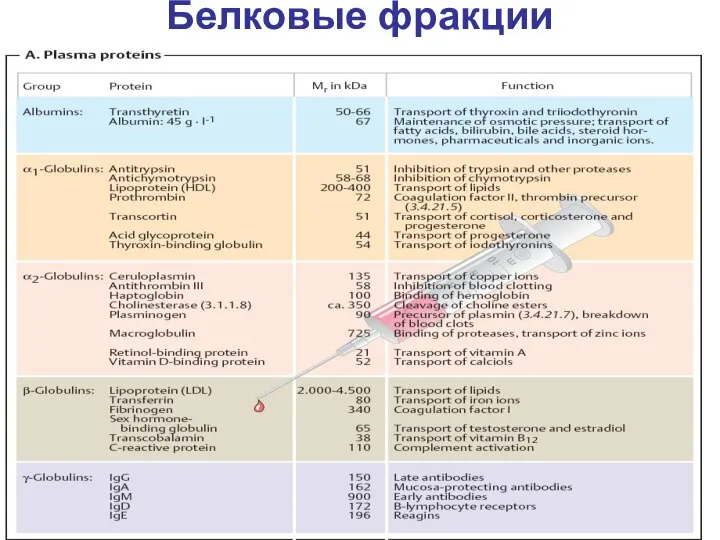
Белковые фракции
Белковые фракции

Функциональная классификация белков плазмы
Транспортная а. специф. Б. неспециф
Резервная ~ 50% альбумина
Регуляторная
Функциональная классификация белков плазмы
Транспортная а. специф. Б. неспециф
Резервная ~ 50% альбумина
Регуляторная

Остаточный азот
Все азотсодержащие вещества плазмы образуют общий пул азота, состоящий из:
Азота
Остаточный азот
Все азотсодержащие вещества плазмы образуют общий пул азота, состоящий из:
Азота

Состав ОА
Мочевина - 50% (главный компонент)
АК - 25% (~ 10% ГЛУ
Состав ОА
Мочевина - 50% (главный компонент)
АК - 25% (~ 10% ГЛУ
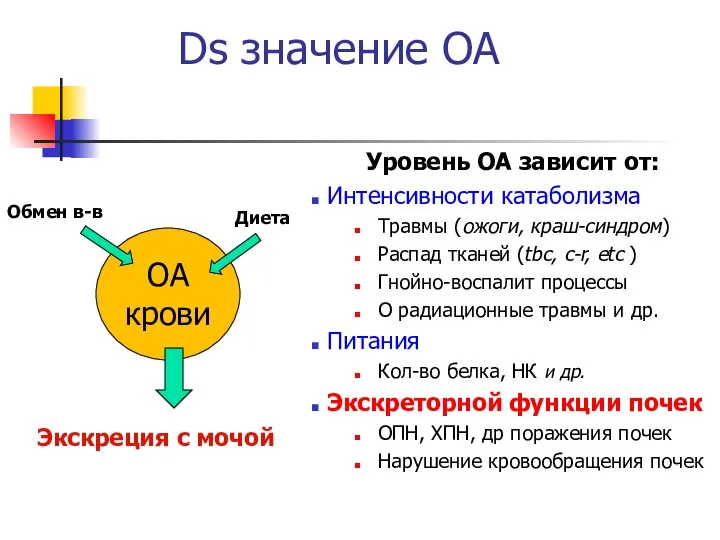
Ds значение ОА
Уровень ОА зависит от:
Интенсивности катаболизма
Травмы (ожоги, краш-синдром)
Распад тканей (tbc,
Ds значение ОА
Уровень ОА зависит от:
Интенсивности катаболизма
Травмы (ожоги, краш-синдром)
Распад тканей (tbc,

Азотемия - повышение уровня ОА в крови
Ретенционная – задержка компонентов
Азотемия - повышение уровня ОА в крови
Ретенционная – задержка компонентов

Общие понятия КОС
КОС – система гомеостаза рН внутри-
и внеклеточной среды
Общие понятия КОС
КОС – система гомеостаза рН внутри-
и внеклеточной среды

рН – производное метаболизма
За сутки организм hs образует 50-100 мМ [Н+]
рН – производное метаболизма
За сутки организм hs образует 50-100 мМ [Н+]

Стабильный рН - необходимое условие метаболизма
Изменение рН приводит к изменению:
заряда и
Стабильный рН - необходимое условие метаболизма
Изменение рН приводит к изменению:
заряда и
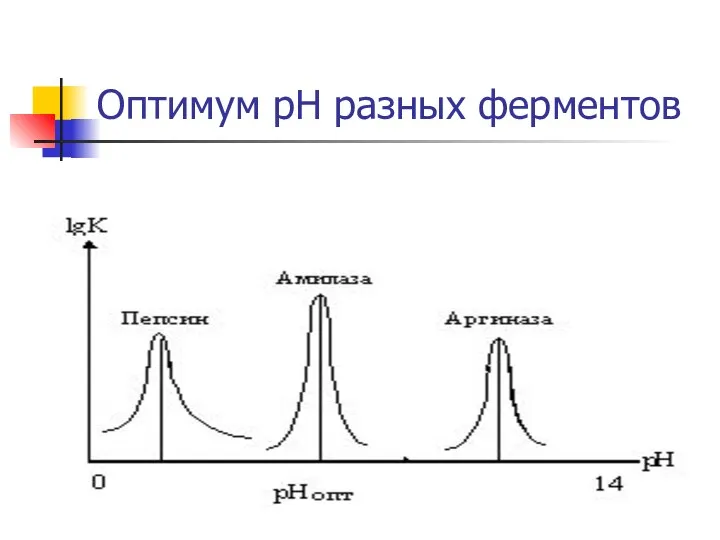
Оптимум рН разных ферментов
Оптимум рН разных ферментов

Изоосмолярность – осм. давление=310 осМ/л - const
любые изменения должны поддерживать эту
любые изменения должны поддерживать эту

Диаграмма Гэмбла
Диаграмма Гэмбла

Механизмы регуляции КОС
Физико-хим – действует в автоматическом режиме и представлен:
разбавлением т.е.
Механизмы регуляции КОС
Физико-хим – действует в автоматическом режиме и представлен:
разбавлением т.е.

Классификация нарушений КОС
рНО =7.40±0.04
рН = 7.35 и ниже – ацидоз
рН =
Классификация нарушений КОС
рНО =7.40±0.04
рН = 7.35 и ниже – ацидоз
рН =
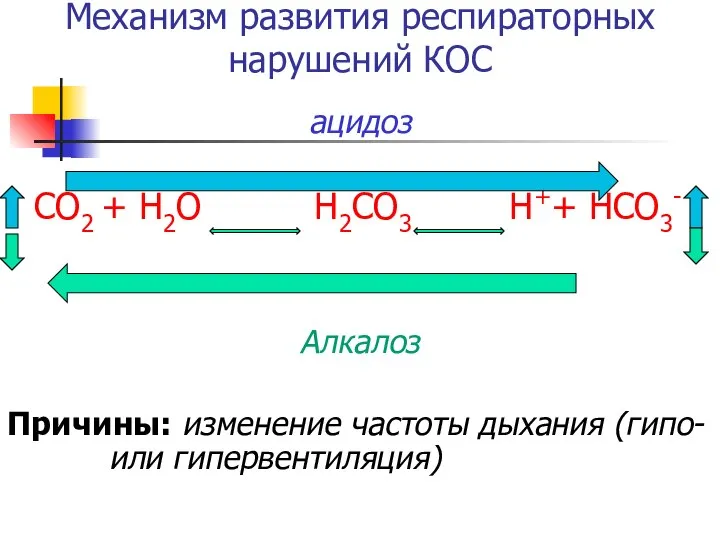
Механизм развития респираторных нарушений КОС
ацидоз
СО2 + Н2О Н2СО3 Н++ НСО3-
Алкалоз
Причины: изменение
Механизм развития респираторных нарушений КОС
ацидоз
СО2 + Н2О Н2СО3 Н++ НСО3-
Алкалоз
Причины: изменение

Межорганное взаимодействие в регуляции рН
Если этих респираторных механизмов недостаточно, то активируются
Межорганное взаимодействие в регуляции рН
Если этих респираторных механизмов недостаточно, то активируются


Эритропоэтин (Эпо)
Эпо – цитокин, специфический регулятор эритропоэза в костном
Эритропоэтин (Эпо)
Эпо – цитокин, специфический регулятор эритропоэза в костном

Э общий обзор
Кол-во Э у мужчин - 4.6-6.2 млн/мкл крови, а
Э общий обзор
Кол-во Э у мужчин - 4.6-6.2 млн/мкл крови, а
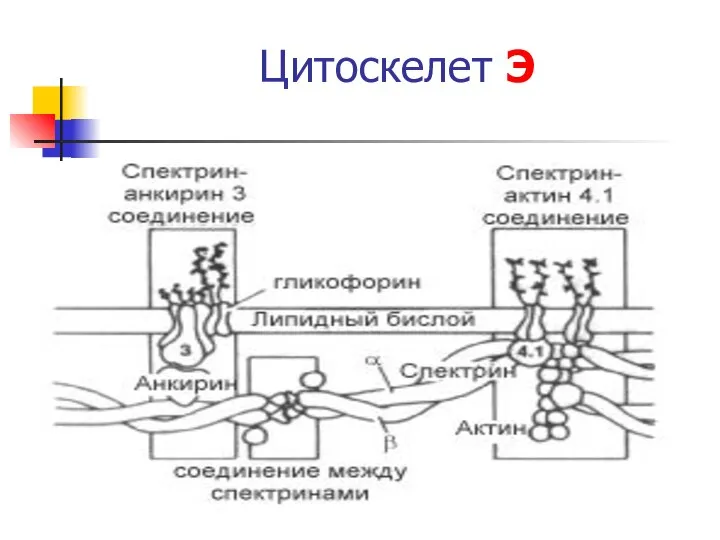
Цитоскелет Э
Цитоскелет Э
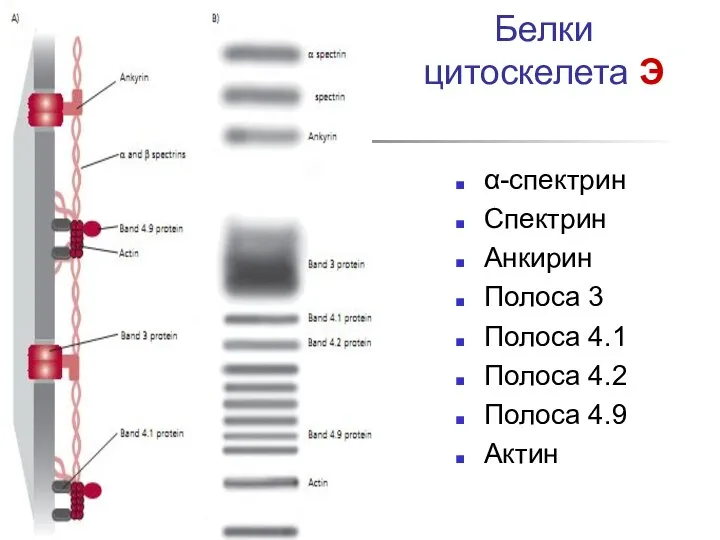
Белки цитоскелета Э
α-спектрин
Спектрин
Анкирин
Полоса 3
Полоса 4.1
Полоса 4.2
Полоса 4.9
Актин
Белки цитоскелета Э
α-спектрин
Спектрин
Анкирин
Полоса 3
Полоса 4.1
Полоса 4.2
Полоса 4.9
Актин
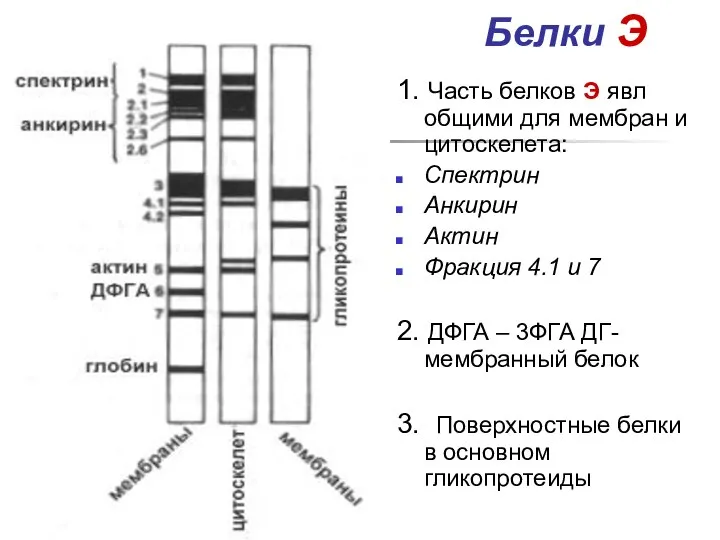
Белки Э
1. Часть белков Э явл общими для мембран и цитоскелета:
Спектрин
Белки Э
1. Часть белков Э явл общими для мембран и цитоскелета:
Спектрин

Структура цитоскелета Э
Большинство мембран Э - интегральные Б,
гликопротеиды.
Б
Структура цитоскелета Э
Большинство мембран Э - интегральные Б,
гликопротеиды.
Б

Метаболизм глюкозы в Э
Гликолиз (90-95%) – образование АТФ
ПФП (10-5%) -
Метаболизм глюкозы в Э
Гликолиз (90-95%) – образование АТФ
ПФП (10-5%) -
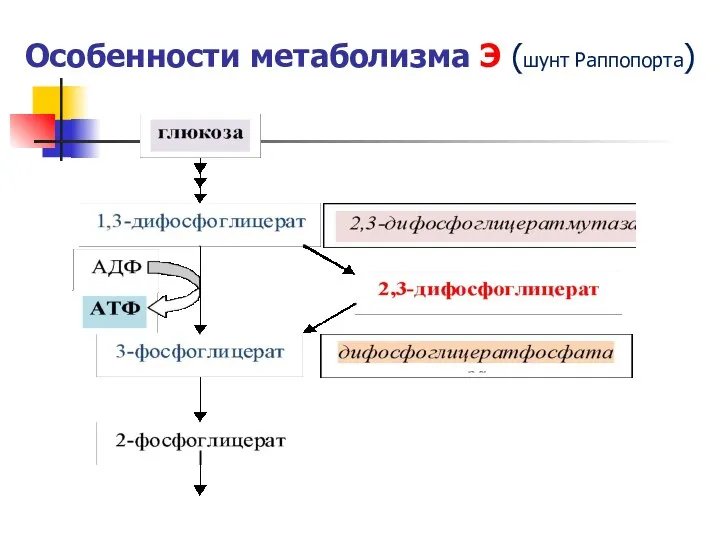
Особенности метаболизма Э (шунт Раппопорта)
Особенности метаболизма Э (шунт Раппопорта)
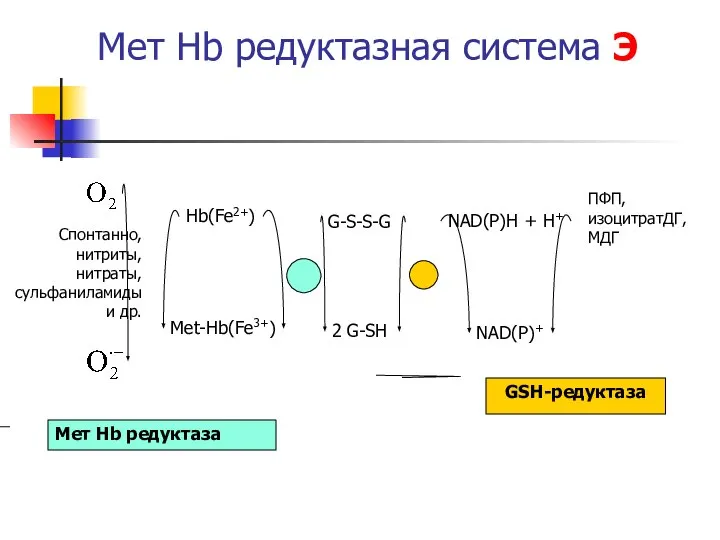
Мет Hb редуктазная система Э
Hb(Fe2+)
Met-Hb(Fe3+)
2 G-SH
G-S-S-G
NAD(P)H + H+
NAD(P)+
Мет Hb редуктаза
GSH-редуктаза
ПФП,
изоцитратДГ,
МДГ
Спонтанно, нитриты,
Мет Hb редуктазная система Э
Hb(Fe2+)
Met-Hb(Fe3+)
2 G-SH
G-S-S-G
NAD(P)H + H+
NAD(P)+
Мет Hb редуктаза
GSH-редуктаза
ПФП,
изоцитратДГ,
МДГ
Спонтанно, нитриты,

Синтез порфобилиногена и гема
Первая реакция б/с гема происходит в Мх и
Синтез порфобилиногена и гема
Первая реакция б/с гема происходит в Мх и


Синтез порфобилиногена и гема (прод)
Затем следует этап конденсации (голова-хвост) 4 молекул
Синтез порфобилиногена и гема (прод)
Затем следует этап конденсации (голова-хвост) 4 молекул
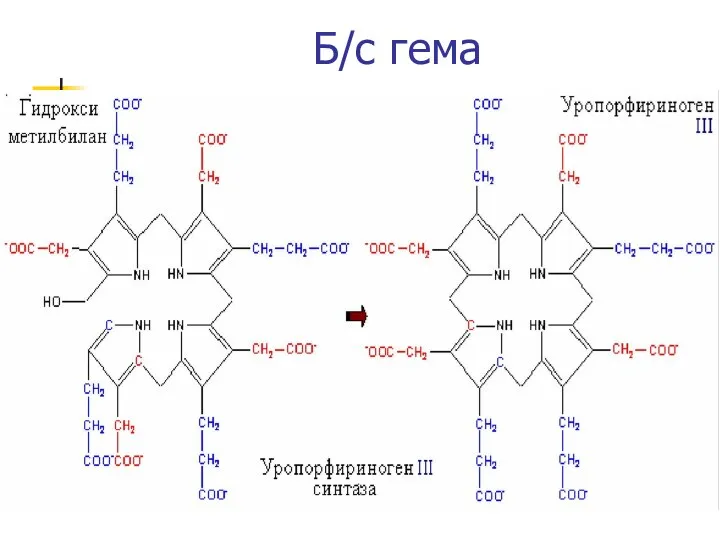
Б/с гема
Б/с гема
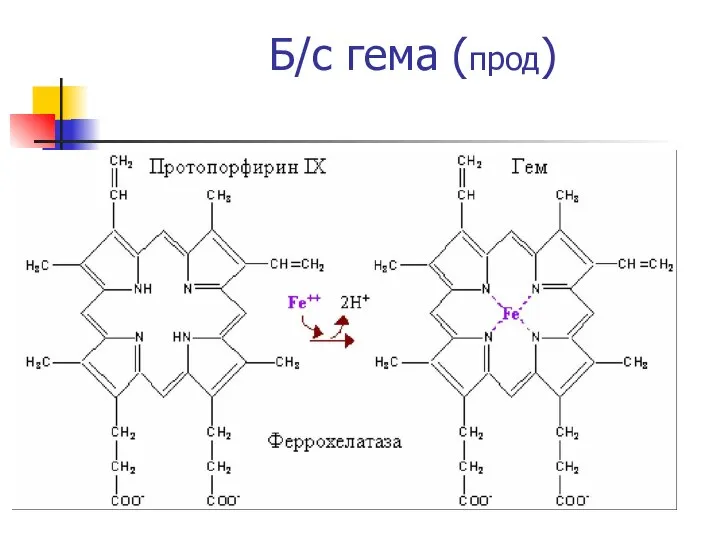
Б/с гема (прод)
Б/с гема (прод)
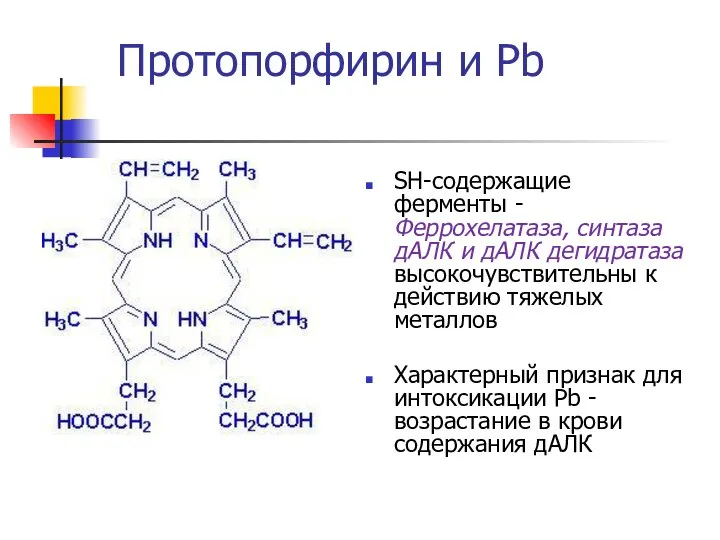
SH-содержащие ферменты - Феррохелатаза, синтаза дАЛК и дАЛК дегидратаза высокочувствительны к
SH-содержащие ферменты - Феррохелатаза, синтаза дАЛК и дАЛК дегидратаза высокочувствительны к
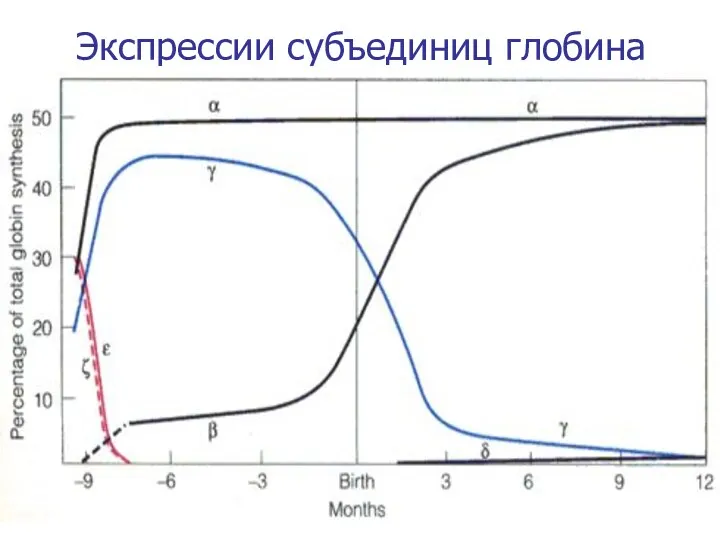
Экспрессии субъединиц глобина
Экспрессии субъединиц глобина
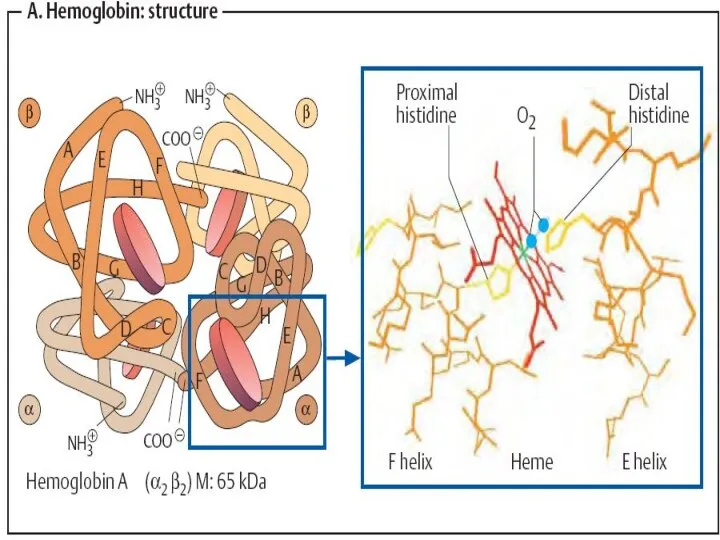
Структура Hb
Структура Hb
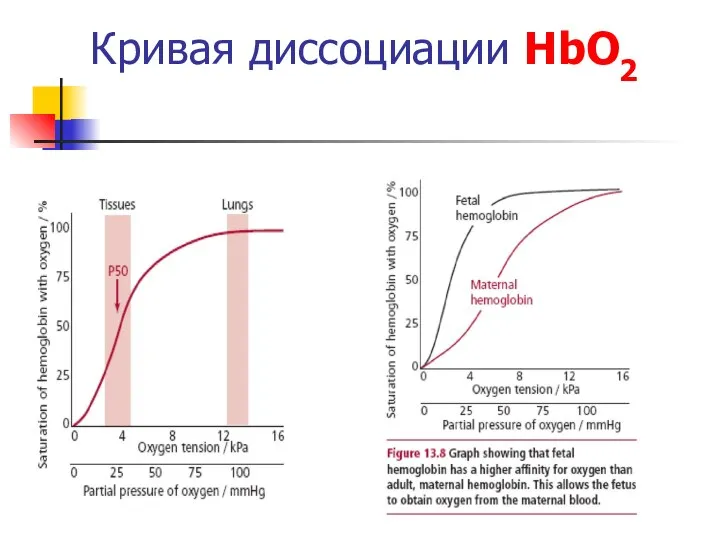
Кривая диссоциации HbО2
Кривая диссоциации HbО2

Распад гема
Продолжительность жизни Э ~120 дней, ежедневный оборот Hb ~6 г/день.
Гем
Распад гема
Продолжительность жизни Э ~120 дней, ежедневный оборот Hb ~6 г/день.
Гем

Образование билирубина
Следующий этап СН2 мост (между кольцами III и IV) биливердина
Образование билирубина
Следующий этап СН2 мост (между кольцами III и IV) биливердина
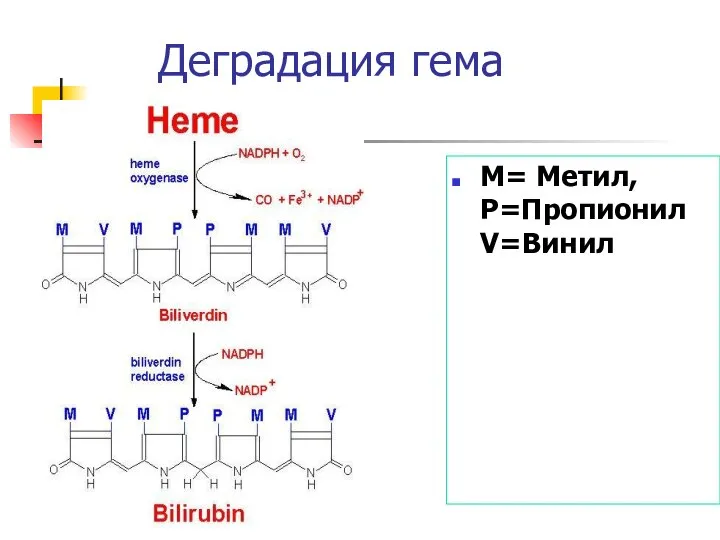
Деградация гема
M= Метил, P=Пропионил V=Винил
Деградация гема
M= Метил, P=Пропионил V=Винил
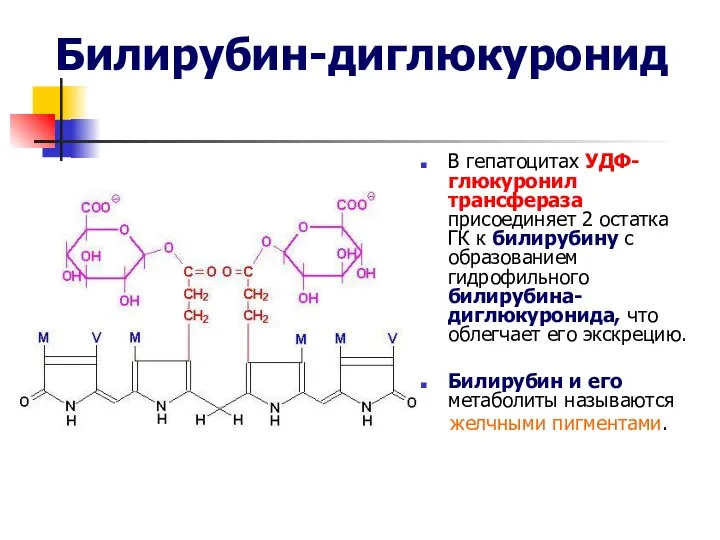
Билирубин-диглюкуронид
В гепатоцитах УДФ-глюкуронил трансфераза присоединяет 2 остатка ГК к билирубину с
Билирубин-диглюкуронид
В гепатоцитах УДФ-глюкуронил трансфераза присоединяет 2 остатка ГК к билирубину с

Клинические аспекты метаболизма гема
представлены в виде:
Дефектов ферментов и нарушения б/с гема
Клинические аспекты метаболизма гема
представлены в виде:
Дефектов ферментов и нарушения б/с гема

Желтухи
Гипербилирубинемия проявляется в виде желтух – желтой пигментации кожи и слизистых.
В
Желтухи
Гипербилирубинемия проявляется в виде желтух – желтой пигментации кожи и слизистых.
В

Нейтрофилы (гранулоциты) –полиморфноядерные лейкоциты:
обладают выраженной способностью к фагоцитозу,
Хорошо развитый, подвижный
Нейтрофилы (гранулоциты) –полиморфноядерные лейкоциты:
обладают выраженной способностью к фагоцитозу,
Хорошо развитый, подвижный
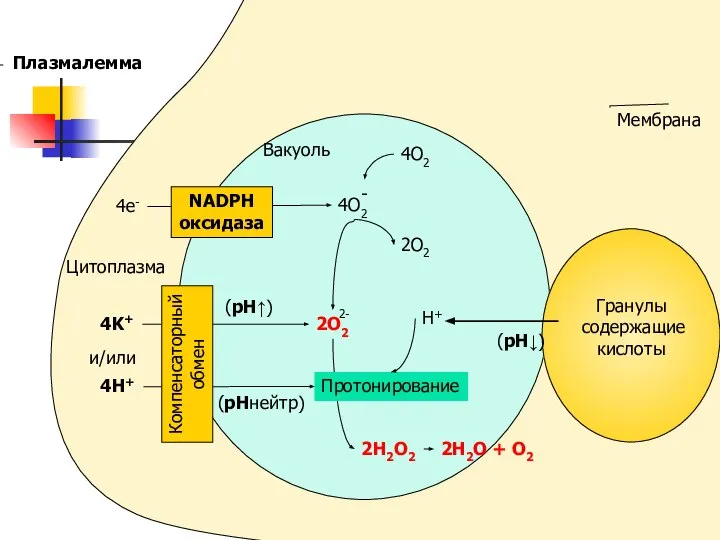
4O2
2O2
H+
2H2O2
2H2O + O2
(pH↓)
Цитоплазма
4e-
Вакуоль
4K+
и/или
4H+
(pHнейтр)
(pH↑)
NADPH
оксидаза
Компенсаторный
обмен
Плазмалемма
Мембрана
Протонирование
4O2
2O2
H+
2H2O2
2H2O + O2
(pH↓)
Цитоплазма
4e-
Вакуоль
4K+
и/или
4H+
(pHнейтр)
(pH↑)
NADPH
оксидаза
Компенсаторный
обмен
Плазмалемма
Мембрана
Протонирование
 Понятие алгоритма и способы записи алгоритмов. (урок 1)
Понятие алгоритма и способы записи алгоритмов. (урок 1) Sony Bravia R5. Инструкция по настройке Fork плеера
Sony Bravia R5. Инструкция по настройке Fork плеера Химическая безопасность
Химическая безопасность Тренажер «Состав чисел первого десятка». Чекалина Наталья Анатольевна. Учитель начальных классов МОУ СОШ №12 г.Волгограда. 2012г.
Тренажер «Состав чисел первого десятка». Чекалина Наталья Анатольевна. Учитель начальных классов МОУ СОШ №12 г.Волгограда. 2012г. Методы принятия управленческих решений Подготовила студентка группы МЭ-102 Швейкина Алёна
Методы принятия управленческих решений Подготовила студентка группы МЭ-102 Швейкина Алёна  Презентация "Ключевые показатели эффективности деятельности персонала банка" - скачать презентации по Экономике
Презентация "Ключевые показатели эффективности деятельности персонала банка" - скачать презентации по Экономике Декор одежды куклы Купавки . Куклы в народной традиции
Декор одежды куклы Купавки . Куклы в народной традиции ОБЩИЕ ПОНЯТИЯ ТЕОРИИ ОШИБОК ИЗМЕРЕНИЙ
ОБЩИЕ ПОНЯТИЯ ТЕОРИИ ОШИБОК ИЗМЕРЕНИЙ Электронные таблицы
Электронные таблицы Купола над «белым безмолвием». Остров Ватерлоо
Купола над «белым безмолвием». Остров Ватерлоо Мовленнєва компетенція
Мовленнєва компетенція Бытовые электроинструменты
Бытовые электроинструменты Беркли и Юм субъективный идеализм
Беркли и Юм субъективный идеализм  Разделительные знаки Ъ и Ь - презентация
Разделительные знаки Ъ и Ь - презентация Где логика. Игра
Где логика. Игра Презентация Рекреационные ресурсы мира
Презентация Рекреационные ресурсы мира  Разработка электронных часов, на микроконтроллере с динамической индикацией
Разработка электронных часов, на микроконтроллере с динамической индикацией Создание планово-высотного обоснования и съемка карьера тахеометром Nikon NPL 332
Создание планово-высотного обоснования и съемка карьера тахеометром Nikon NPL 332 Alfresco ECM (Enterprise Content Management)
Alfresco ECM (Enterprise Content Management) Федя Клюков
Федя Клюков Твердотільна електроніка. Лекция 4. Лавинно-пролітні діоди
Твердотільна електроніка. Лекция 4. Лавинно-пролітні діоди Искусство рассказывает о красоте Земли 8 класс
Искусство рассказывает о красоте Земли 8 класс История государственного гимна России
История государственного гимна России Лекция 3 Электропроводность растворов электролитов
Лекция 3 Электропроводность растворов электролитов Презентация "Первобытное искусство (6 класс)" - скачать презентации по МХК
Презентация "Первобытное искусство (6 класс)" - скачать презентации по МХК Типы соединения костей. Строение и классификация суставов. Биомеханика суставов
Типы соединения костей. Строение и классификация суставов. Биомеханика суставов Классификация тепловых электростанций и их энергоэкономические особенности
Классификация тепловых электростанций и их энергоэкономические особенности Монтаж высотных инженерных сооружений
Монтаж высотных инженерных сооружений