- Биохимия печени

Содержание
- 2. Печень – посредник между кишечником и другими органами и тканями. У взрослого человека вес печени -
- 3. Схема гепатоцита
- 4. Функции печени
- 5. Функции печени
- 6. Химический состав печени
- 7. Роль печени в углеводном обмене Печень – депо гликогена. Печень обеспечивает постоянный уровень глюкозы в крови,
- 8. Регуляция синтеза и распада гликогена Увеличивают содержание гликогена в печени: АКТГ, глюкокортикоиды, инсулин. Стимулируют распад гликогена:
- 9. Гликогенолиз Нарушение активности ферментов фосфоролиза приводит к накоплению гликогена в печени и далее к гипогликемии. Это
- 10. Глюконеогенез в печени
- 11. Метаболизм фруктозы и галактозы
- 12. Роль печени в липидном обмене В печени осуществляется: синтез желчных кислот (желчь необходима для переваривания и
- 13. Обмен липидов в печени
- 14. Биосинтез кетоновых тел
- 15. Желчные кислоты
- 16. Метаболизм желчных кислот
- 17. Роль печени в обмене белков В печени протекает: синтез белков (за сутки обновляется около 9 %
- 18. Метаболизм гормонов в печени Инактивация: стероидных гормонов, тироксина, АДГ, альдостерона, эстрогенов, инсулина. Синтез: транскортина, дофамина.
- 19. Печень, витамины, микроэлементы Печень - депо витаминов А, Д, К, РР. В большом количестве содержатся витамины
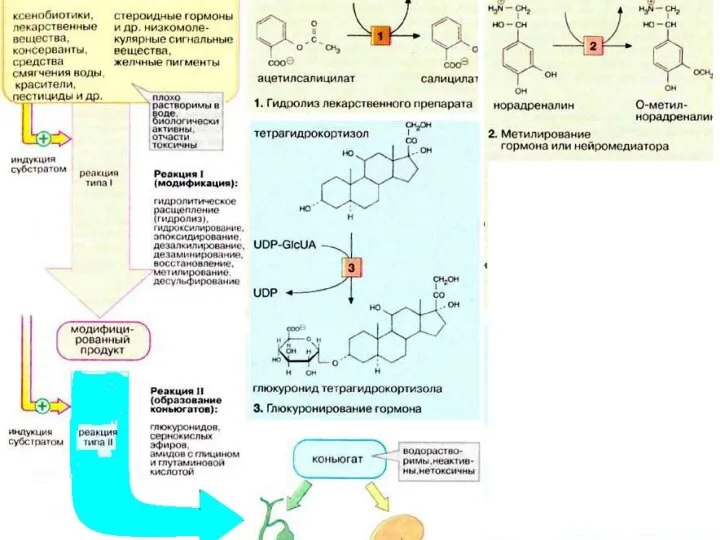
- 20. Роль печени в обезвреживании метаболитов и токсических веществ Обезвреживание происходит путем: окисления, восстановления, метилирования, ацетилирования, конъюгации.
- 21. Обезвреживание токсических веществ в печени В печени происходит синтез мочевины (обезвреживание аммиака). Путем образования парных соединений
- 23. Метаболизм этанола в печени
- 24. Жировая дистрофия печени
- 25. Микросомальное окисление Микросомы – морфологически замкнутые везикулы, в которые превращается эндоплазматический ретикулум при гомогенизации тканей. Функция
- 26. Монооксигеназная система состоит из трёх компонентов: НАДФ-специфичного ФАД - содержащего флавопротеина, железосерного белка, цитохрома Р450. Монооксигеназы
- 27. Реакции, катализируемые системой цитохром Р450
- 29. Цитохром Р450 выполняет двойную функцию: цитохром Р450 связывает субстрат гидроксилирования, на нём происходит активация молекулярного кислорода.
- 30. Цепь микросом печени - универсальная биологическая система, окисляющая неполярные соединения любого происхождения: эндогенные субстраты – стероидные
- 31. Элиминация. Окисление молекулярным кислородом приводит к увеличению гидрофильности чужеродных соединений. Детоксикация. Химическая модификация приводит к потере
- 32. Микросомальные гидроксилазы могут катализировать не только гидроксилирование, но и другие реакции: эпоксидирование, сульфоокисление, дезалкилирование, восстановление нитросоединений.
- 33. Цитохром Р450 инактивируется in vitro окисью углерода и тиоловыми ядами, реактивируется – тиоловыми антиоксидантами. В печени
- 34. Внутриклеточная локализация основных видов конъюгации
- 35. Метаболизм и выведение ксенобиотиков из организма
- 36. Субклеточная локализация ферментных систем в печени Ядро гепатоцита служит хранилищем информации и может быть источником генетических
- 37. Биохимические показатели при цитолитическом синдроме Повышение в сыворотке крови активности: АЛТ, АСТ, альдолазы, ГЛДГ, сорбитолдегидрогеназы, ЛДГ
- 38. Соотношение АСТ/АЛТ: АЛТ сосредоточен в цитозоле. АСТ – в цитозоле и митохондриях. При вирусном гепатите первично
- 39. Повышение активности: щелочной фосфатазы (стимуляция биосинтеза ЩФ на поверхности гепатоцитов поступление в синусоиды проникновение в кровь)
- 40. Биохимические показатели при мезенхимально-воспалительном синдроме (исследование сыворотки крови) Рост содержания иммуноглобулинов: IgM при острых воспалениях, IgG
- 41. Индикаторы гепато-депрессивного синдрома (малой недостаточности печени) Показатели выделительной функции: Скорость выведения бромсульфолеина 10-16 мг краски/мин. Тесты
- 42. Индикаторы гепатодепрессии, связанные с синтезом белка альбумины (35-50 г/л), фибронектин (333+8,6 мкг/мл), церулоплазмин (0,15-0,6 г/л), а1-АТ
- 43. Биохимические показатели при синдроме печеночно-клеточной недостаточности (исследования сыворотки крови) понижение активности холинэстеразы, гипопротеинемия и диспротеинемия с
- 44. Биохимические показатели при синдроме портокавального шунтирования Возникает этот синдром за счет развития мощных венозных коллатералей с
- 45. Взаимосвязь обменов осуществляется благодаря интегрирующим системам: нервной, эндокринной, сосудистой. Взаимосвязь обеспечивается различными уровнями: информационный уровень, структурный
- 46. Информационный уровень взаимосвязи В геноме клеток заложена информация о структуре и функциональной активности различных белков ,
- 47. Структурный уровень взаимосвязи мембранный аппарат клеток, рибосомы.
- 48. Общее энергетическое обеспечение АТФ – универсальная энергетическая валюта, образующаяся при окислении углеводов, жиров, аминокислот. НАДФН2 –
- 49. Окисляемые субстраты Продукты окисления Продукты биосинтезов Субстраты для биосинтезов НАДФ+ НАДФН+Н+ НАДН+Н+ НАД+ 2Н+ 2Н+ 2Н+
- 50. Взаимосвязь на уровне общих метаболитов Центральные метаболиты: ацетил-КоА, ПВК, ЩУК, ФГА.
- 51. ПВК ФГА ацетил-КоА ЩУК Белки Углеводы Жиры Глицерин Холестерин Ц.Т.К. Кетоновые тела Жирные кислоты АМК Лактат
- 52. Ацетил-КоА образуется при окислительном декарбоксилировании ПВК, при β-окислении жирных кислот, из аминокислот.
- 53. ПВК + СО2 ЩУК пируваткарбоксилаза Биотин ЩУК асп глюконеогенез Ц.Т.К. аспарагин Синтез мочевины Синтез пиримидиновых нуклеотидов
- 54. Взаимосвязь углеводного и липидного обменов осуществляется через ацетил-КоА, ФГА, НАДФН2 из пентозного цикла идёт на синтез
- 55. Взаимосвязь белкового и жирового обменов на уровне ПВК, ацетил-КоА, кетоновых тел. Из белков осуществляется синтез жира.
- 56. Взаимосвязь углеводного и белкового обменов на уровне ПВК, ЩУК, ацетил-КоА, пентоз, глюкопластичных и кетопластичных аминокислот. Углеродные
- 57. Сопряжение на уровне Ц.Т.К. углеродные скелеты АМК включаются в Ц.Т.К, все кислоты Ц.Т.К. превращаются в ЩУК
- 58. Связь липидного обмена с Ц.Т.К. осуществляется через цитрат - активатор ацетил-КоА-карбоксилазы, - перенос в цитоплазму ацетил-КоА,
- 59. Взаимосвязь обменов на уровне органов и тканей Печень и мышцы. В покоящихся мышцах субстрат энергетического обмена
- 60. Печень и мозг. Глюкоза из печени с кровью поступает в мозг. В ткани мозга содержится много
- 61. Почки и печень. Глюконеогенез происходит в почках и печени. Почки зависят от поступления из печени глутамина,
- 63. Скачать презентацию

Печень –
посредник между кишечником и другими
органами и тканями.
У взрослого
Печень –
посредник между кишечником и другими
органами и тканями.
У взрослого
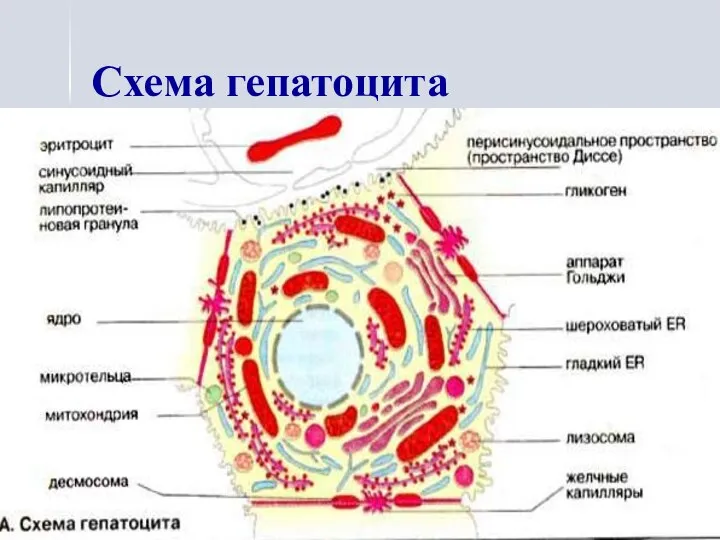
Схема гепатоцита
Схема гепатоцита

Функции печени
Функции печени
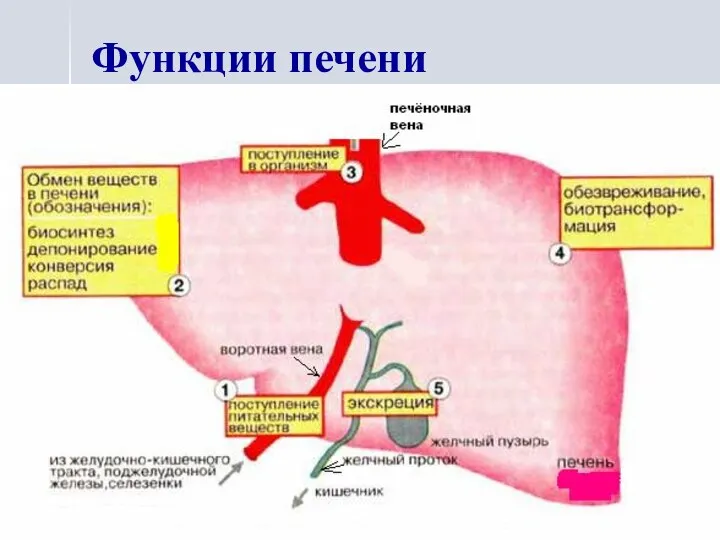
Функции печени
Функции печени
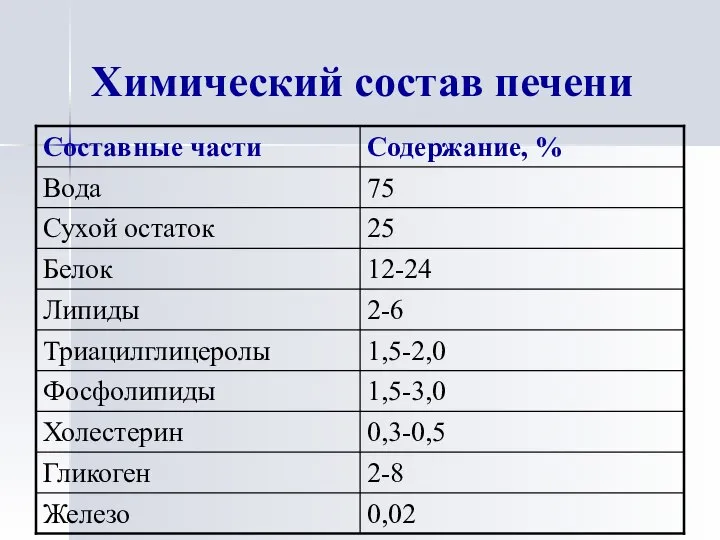
Химический состав печени
Химический состав печени

Роль печени в углеводном обмене
Печень – депо гликогена.
Печень обеспечивает постоянный
Роль печени в углеводном обмене
Печень – депо гликогена.
Печень обеспечивает постоянный

Регуляция синтеза и распада гликогена
Увеличивают содержание гликогена в печени:
АКТГ,
Регуляция синтеза и распада гликогена
Увеличивают содержание гликогена в печени:
АКТГ,

Гликогенолиз
Нарушение активности ферментов фосфоролиза приводит к накоплению гликогена в печени
Гликогенолиз
Нарушение активности ферментов фосфоролиза приводит к накоплению гликогена в печени
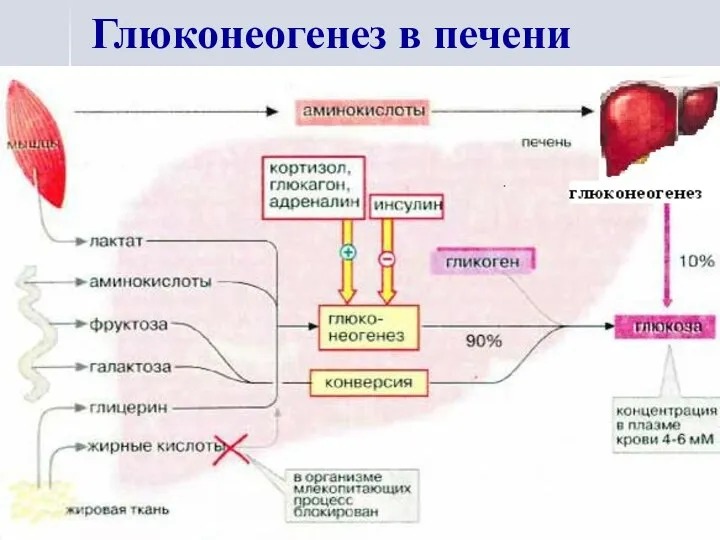
Глюконеогенез в печени
Глюконеогенез в печени
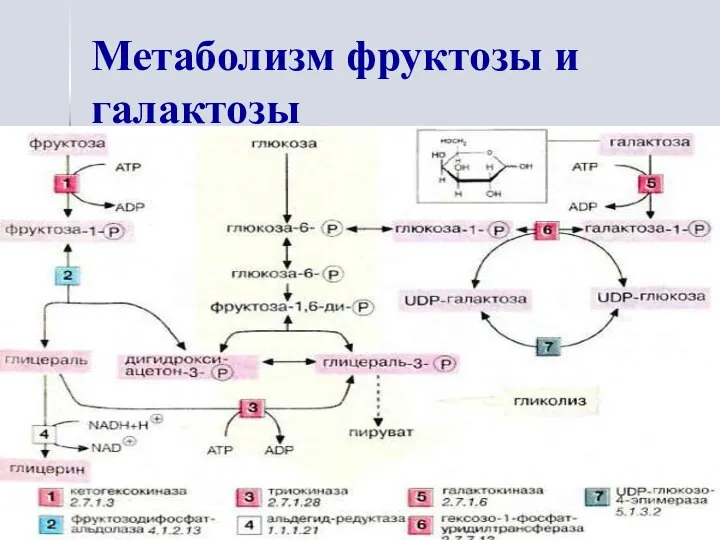
Метаболизм фруктозы и галактозы
Метаболизм фруктозы и галактозы

Роль печени в липидном обмене
В печени осуществляется:
синтез желчных кислот
Роль печени в липидном обмене
В печени осуществляется:
синтез желчных кислот
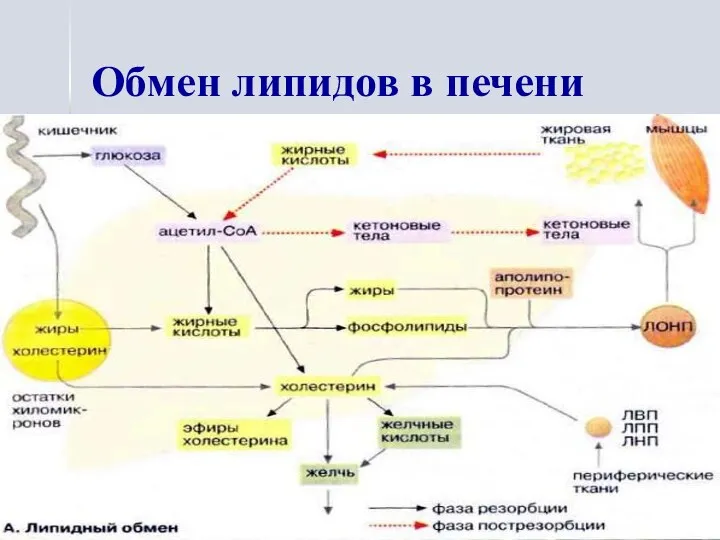
Обмен липидов в печени
Обмен липидов в печени
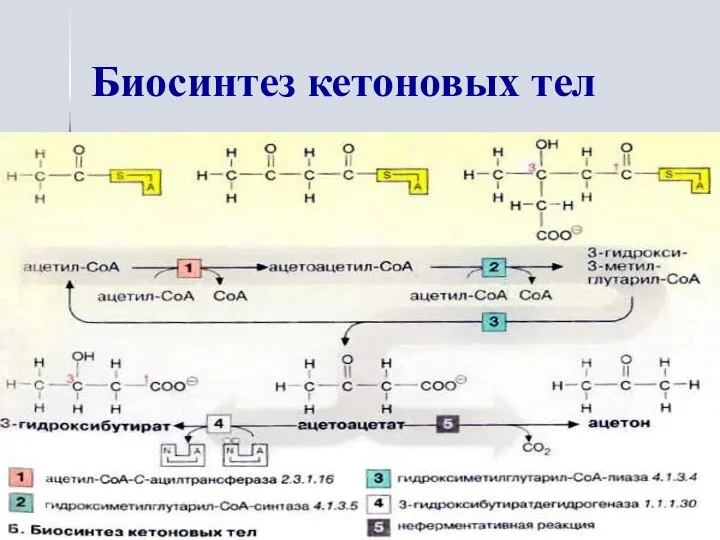
Биосинтез кетоновых тел
Биосинтез кетоновых тел
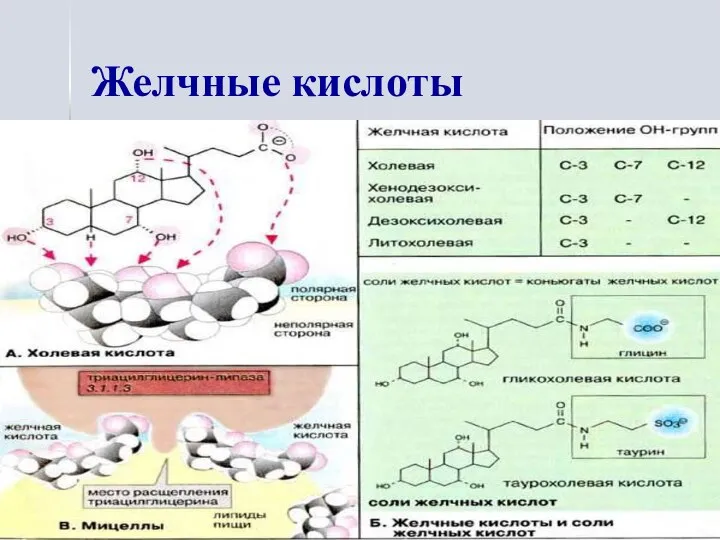
Желчные кислоты
Желчные кислоты
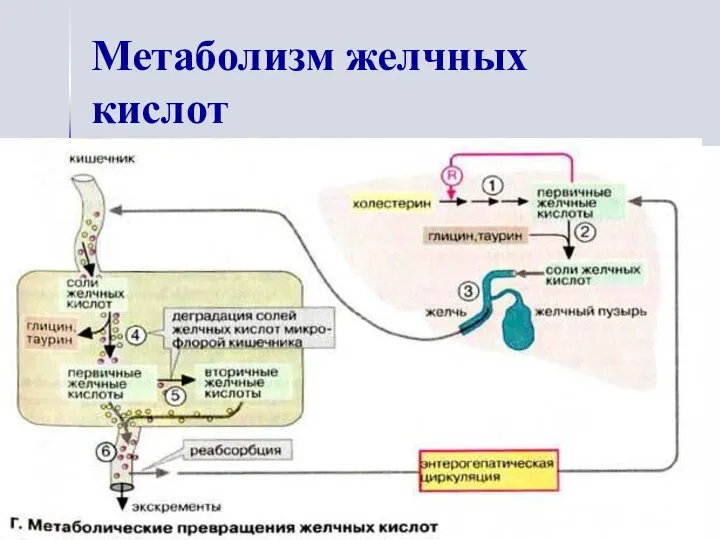
Метаболизм желчных кислот
Метаболизм желчных кислот

Роль печени в обмене белков
В печени протекает:
синтез белков (за сутки
Роль печени в обмене белков
В печени протекает:
синтез белков (за сутки

Метаболизм гормонов в печени
Инактивация:
стероидных гормонов,
тироксина,
АДГ,
альдостерона,
эстрогенов,
инсулина.
Синтез:
Метаболизм гормонов в печени
Инактивация:
стероидных гормонов,
тироксина,
АДГ,
альдостерона,
эстрогенов,
инсулина.
Синтез:

Печень, витамины, микроэлементы
Печень - депо витаминов А, Д, К, РР.
В
Печень, витамины, микроэлементы
Печень - депо витаминов А, Д, К, РР.
В

Роль печени в обезвреживании метаболитов и токсических веществ
Обезвреживание происходит путем:
окисления,
Роль печени в обезвреживании метаболитов и токсических веществ
Обезвреживание происходит путем:
окисления,

Обезвреживание токсических веществ в печени
В печени происходит синтез мочевины (обезвреживание аммиака).
Путем
Обезвреживание токсических веществ в печени
В печени происходит синтез мочевины (обезвреживание аммиака).
Путем
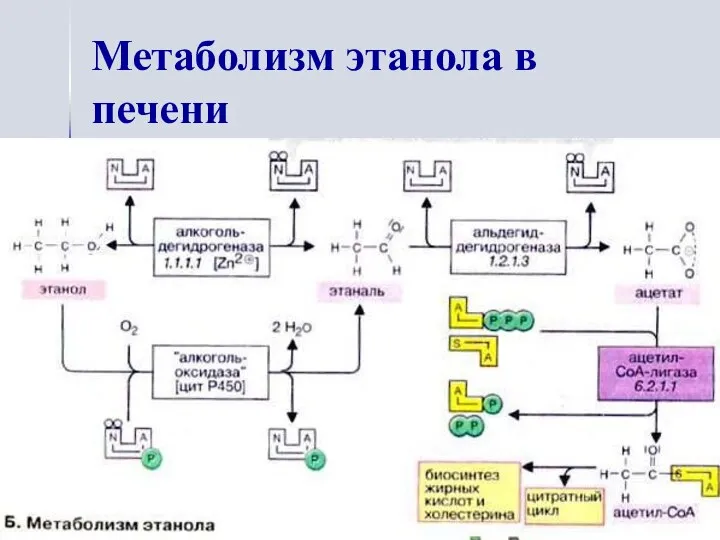
Метаболизм этанола в печени
Метаболизм этанола в печени
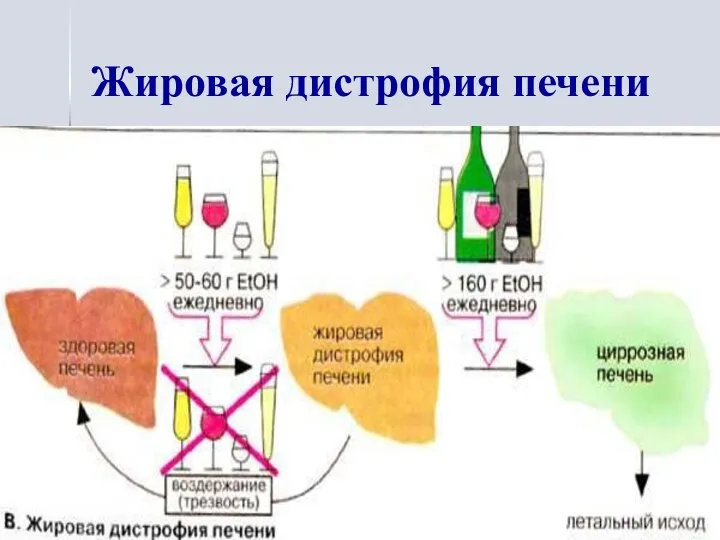
Жировая дистрофия печени
Жировая дистрофия печени

Микросомальное окисление
Микросомы – морфологически замкнутые везикулы, в которые превращается эндоплазматический
Микросомальное окисление
Микросомы – морфологически замкнутые везикулы, в которые превращается эндоплазматический

Монооксигеназная система состоит из трёх компонентов:
НАДФ-специфичного ФАД - содержащего
Монооксигеназная система состоит из трёх компонентов:
НАДФ-специфичного ФАД - содержащего
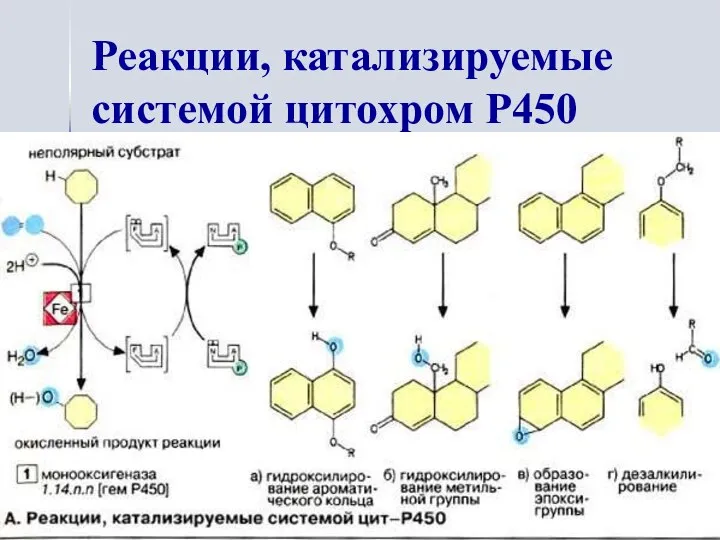
Реакции, катализируемые системой цитохром Р450
Реакции, катализируемые системой цитохром Р450
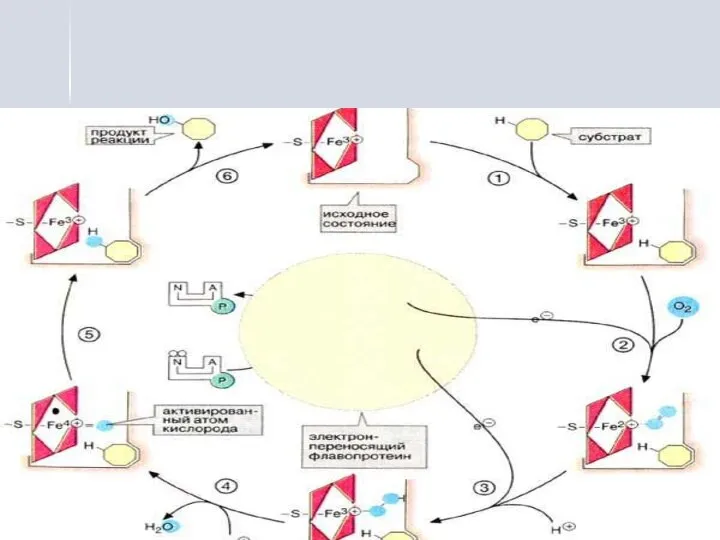

Цитохром Р450 выполняет двойную функцию:
цитохром Р450 связывает субстрат гидроксилирования,
на
Цитохром Р450 выполняет двойную функцию:
цитохром Р450 связывает субстрат гидроксилирования,
на

Цепь микросом печени
- универсальная биологическая система, окисляющая неполярные соединения любого происхождения:
Цепь микросом печени
- универсальная биологическая система, окисляющая неполярные соединения любого происхождения:

Элиминация.
Окисление молекулярным кислородом приводит к увеличению гидрофильности чужеродных соединений.
Детоксикация.
Химическая модификация
Элиминация.
Окисление молекулярным кислородом приводит к увеличению гидрофильности чужеродных соединений.
Детоксикация.
Химическая модификация

Микросомальные гидроксилазы могут
катализировать не только гидроксилирование,
но и другие реакции:
эпоксидирование,
сульфоокисление,
Микросомальные гидроксилазы могут
катализировать не только гидроксилирование,
но и другие реакции:
эпоксидирование,
сульфоокисление,

Цитохром Р450
инактивируется in vitro окисью углерода и тиоловыми ядами,
реактивируется
Цитохром Р450
инактивируется in vitro окисью углерода и тиоловыми ядами,
реактивируется
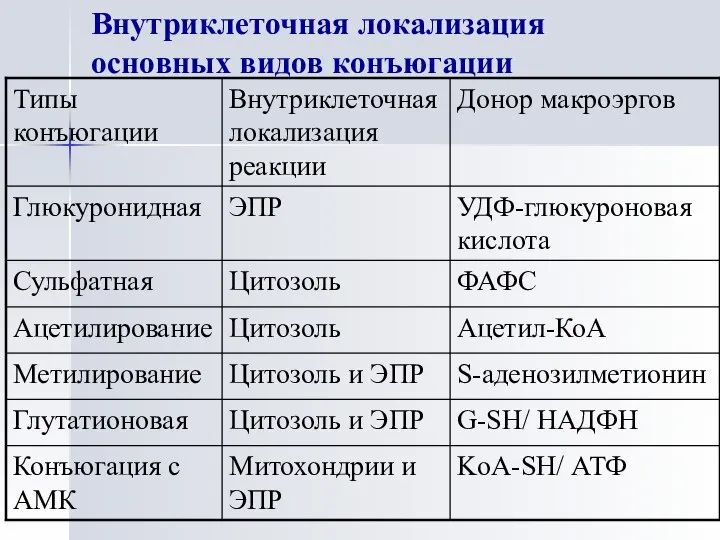
Внутриклеточная локализация основных видов конъюгации
Внутриклеточная локализация основных видов конъюгации
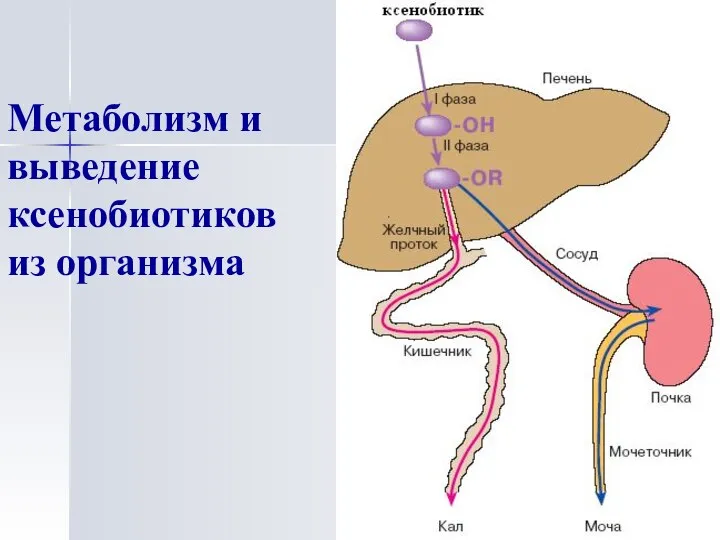
Метаболизм и выведение ксенобиотиков из организма
Метаболизм и выведение ксенобиотиков из организма

Субклеточная локализация ферментных систем в печени
Ядро гепатоцита служит хранилищем информации и
Субклеточная локализация ферментных систем в печени
Ядро гепатоцита служит хранилищем информации и

Биохимические показатели при цитолитическом синдроме
Повышение в сыворотке крови активности:
АЛТ,
Биохимические показатели при цитолитическом синдроме
Повышение в сыворотке крови активности:
АЛТ,

Соотношение АСТ/АЛТ:
АЛТ сосредоточен в цитозоле.
АСТ – в цитозоле и
Соотношение АСТ/АЛТ:
АЛТ сосредоточен в цитозоле.
АСТ – в цитозоле и

Повышение активности:
щелочной фосфатазы (стимуляция биосинтеза ЩФ на поверхности гепатоцитов поступление в
Повышение активности:
щелочной фосфатазы (стимуляция биосинтеза ЩФ на поверхности гепатоцитов поступление в

Биохимические показатели при мезенхимально-воспалительном синдроме (исследование сыворотки крови)
Рост содержания иммуноглобулинов:
IgM
Биохимические показатели при мезенхимально-воспалительном синдроме (исследование сыворотки крови)
Рост содержания иммуноглобулинов:
IgM

Индикаторы гепато-депрессивного синдрома
(малой недостаточности печени)
Показатели выделительной функции:
Скорость выведения бромсульфолеина 10-16
Индикаторы гепато-депрессивного синдрома
(малой недостаточности печени)
Показатели выделительной функции:
Скорость выведения бромсульфолеина 10-16

Индикаторы гепатодепрессии, связанные с синтезом белка
альбумины (35-50 г/л),
фибронектин (333+8,6 мкг/мл),
церулоплазмин (0,15-0,6
Индикаторы гепатодепрессии, связанные с синтезом белка
альбумины (35-50 г/л),
фибронектин (333+8,6 мкг/мл),
церулоплазмин (0,15-0,6

Биохимические показатели при синдроме печеночно-клеточной недостаточности
(исследования сыворотки крови)
понижение активности
Биохимические показатели при синдроме печеночно-клеточной недостаточности
(исследования сыворотки крови)
понижение активности

Биохимические показатели при синдроме портокавального шунтирования
Возникает этот синдром за счет развития
Биохимические показатели при синдроме портокавального шунтирования
Возникает этот синдром за счет развития

Взаимосвязь обменов
осуществляется благодаря интегрирующим системам:
нервной,
эндокринной,
сосудистой.
Взаимосвязь обеспечивается различными
Взаимосвязь обменов
осуществляется благодаря интегрирующим системам:
нервной,
эндокринной,
сосудистой.
Взаимосвязь обеспечивается различными

Информационный уровень взаимосвязи
В геноме клеток заложена информация о структуре и функциональной
Информационный уровень взаимосвязи
В геноме клеток заложена информация о структуре и функциональной
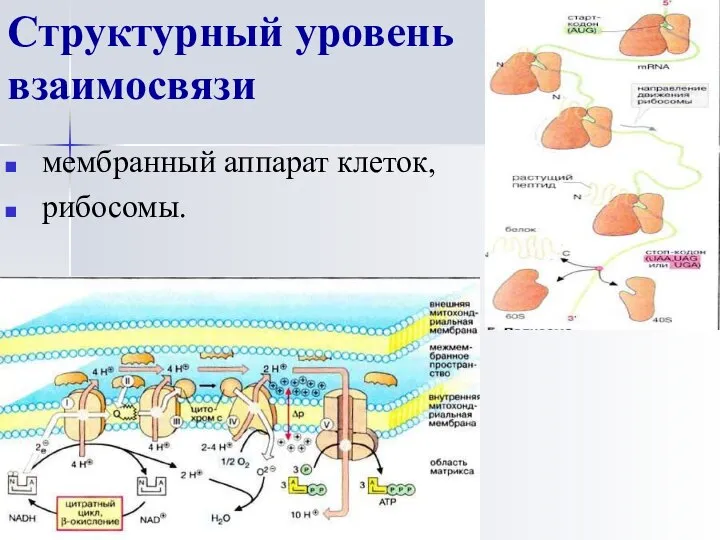
Структурный уровень взаимосвязи
мембранный аппарат клеток,
рибосомы.
Структурный уровень взаимосвязи
мембранный аппарат клеток,
рибосомы.

Общее энергетическое обеспечение
АТФ – универсальная энергетическая валюта, образующаяся при окислении углеводов,
Общее энергетическое обеспечение
АТФ – универсальная энергетическая валюта, образующаяся при окислении углеводов,
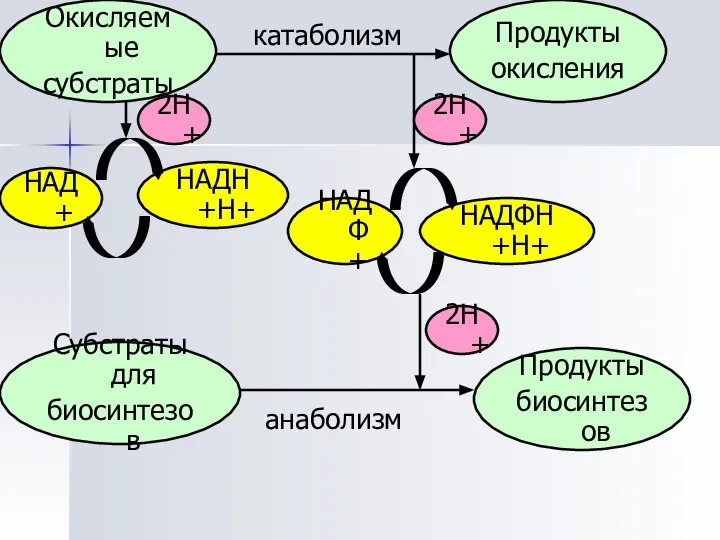
Окисляемые
субстраты
Продукты
окисления
Продукты
биосинтезов
Субстраты для
биосинтезов
НАДФ+
НАДФН+Н+
НАДН+Н+
НАД+
2Н+
2Н+
2Н+
анаболизм
катаболизм
Окисляемые
субстраты
Продукты
окисления
Продукты
биосинтезов
Субстраты для
биосинтезов
НАДФ+
НАДФН+Н+
НАДН+Н+
НАД+
2Н+
2Н+
2Н+
анаболизм
катаболизм

Взаимосвязь на уровне общих метаболитов
Центральные метаболиты:
ацетил-КоА,
ПВК,
ЩУК,
ФГА.
Взаимосвязь на уровне общих метаболитов
Центральные метаболиты:
ацетил-КоА,
ПВК,
ЩУК,
ФГА.
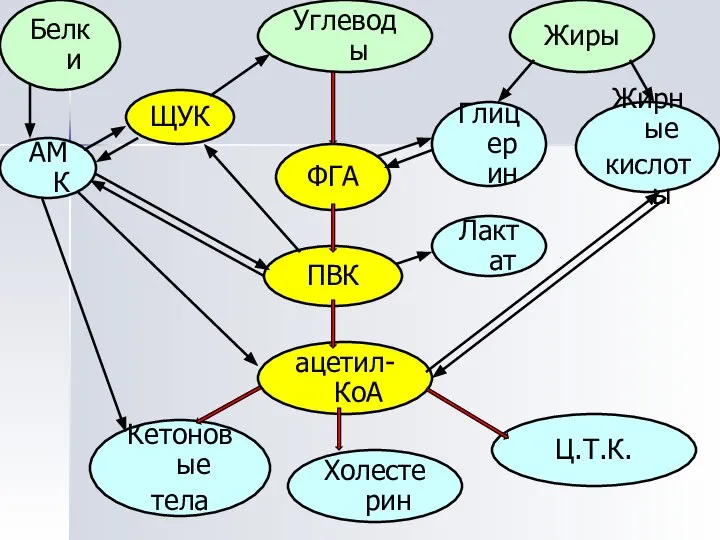
ПВК
ФГА
ацетил-КоА
ЩУК
Белки
Углеводы
Жиры
Глицерин
Холестерин
Ц.Т.К.
Кетоновые
тела
Жирные
кислоты
АМК
Лактат
ПВК
ФГА
ацетил-КоА
ЩУК
Белки
Углеводы
Жиры
Глицерин
Холестерин
Ц.Т.К.
Кетоновые
тела
Жирные
кислоты
АМК
Лактат

Ацетил-КоА образуется
при окислительном декарбоксилировании ПВК,
при β-окислении жирных кислот,
из
Ацетил-КоА образуется
при окислительном декарбоксилировании ПВК,
при β-окислении жирных кислот,
из
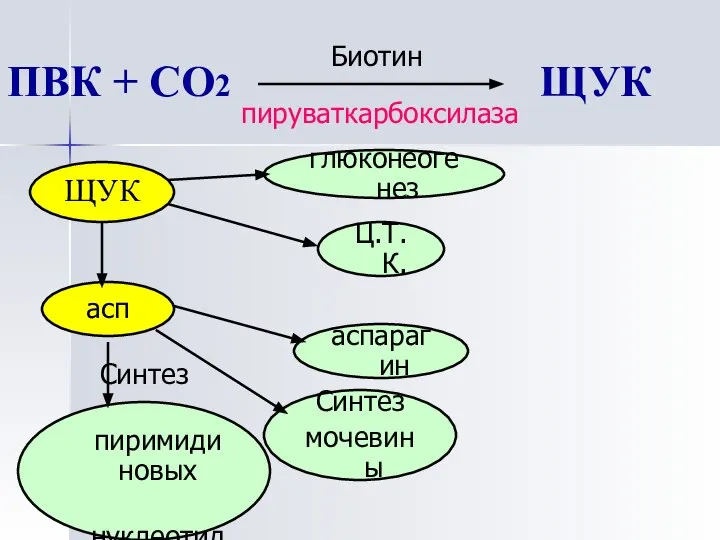
ПВК + СО2 ЩУК
пируваткарбоксилаза
Биотин
ЩУК
асп
глюконеогенез
Ц.Т.К.
аспарагин
Синтез
мочевины
Синтез
пиримидиновых
нуклеотидов
ПВК + СО2 ЩУК
пируваткарбоксилаза
Биотин
ЩУК
асп
глюконеогенез
Ц.Т.К.
аспарагин
Синтез
мочевины
Синтез
пиримидиновых
нуклеотидов

Взаимосвязь углеводного и липидного обменов осуществляется через
ацетил-КоА,
ФГА,
НАДФН2 из пентозного цикла идёт
Взаимосвязь углеводного и липидного обменов осуществляется через
ацетил-КоА,
ФГА,
НАДФН2 из пентозного цикла идёт

Взаимосвязь белкового и жирового обменов на уровне
ПВК,
ацетил-КоА,
кетоновых тел.
Из
Взаимосвязь белкового и жирового обменов на уровне
ПВК,
ацетил-КоА,
кетоновых тел.
Из

Взаимосвязь углеводного и белкового обменов на уровне
ПВК,
ЩУК,
ацетил-КоА,
пентоз,
глюкопластичных и кетопластичных
Взаимосвязь углеводного и белкового обменов на уровне
ПВК,
ЩУК,
ацетил-КоА,
пентоз,
глюкопластичных и кетопластичных
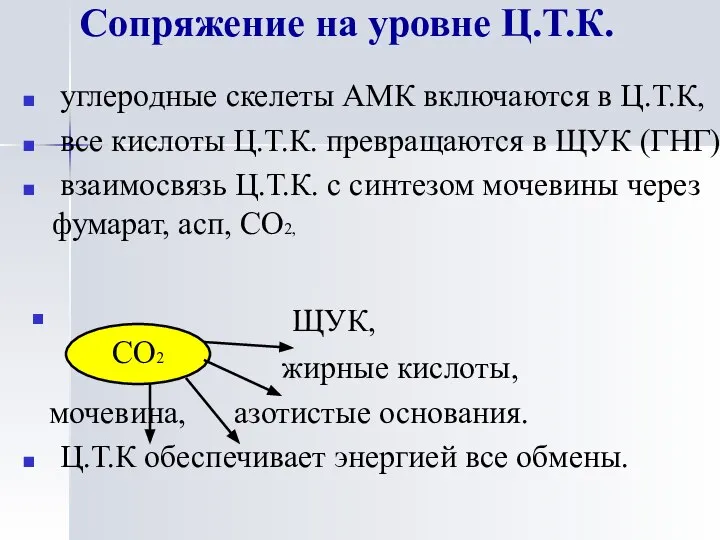
Сопряжение на уровне Ц.Т.К.
углеродные скелеты АМК включаются в Ц.Т.К,
все
Сопряжение на уровне Ц.Т.К.
углеродные скелеты АМК включаются в Ц.Т.К,
все

Связь липидного обмена с Ц.Т.К. осуществляется через
цитрат
- активатор ацетил-КоА-карбоксилазы,
-
Связь липидного обмена с Ц.Т.К. осуществляется через
цитрат
- активатор ацетил-КоА-карбоксилазы,
-

Взаимосвязь обменов
на уровне органов и тканей
Печень и мышцы.
В покоящихся
Взаимосвязь обменов
на уровне органов и тканей
Печень и мышцы.
В покоящихся

Печень и мозг.
Глюкоза из печени с кровью поступает в мозг.
В
Печень и мозг.
Глюкоза из печени с кровью поступает в мозг.
В

Почки и печень.
Глюконеогенез происходит в почках и печени.
Почки зависят от поступления
Почки и печень.
Глюконеогенез происходит в почках и печени.
Почки зависят от поступления
 Написание итогового сочинения.
Написание итогового сочинения. MapInfo
MapInfo  Фундаменты малоэтажных зданий
Фундаменты малоэтажных зданий Творческие работы учащихся художественного отделения МОУ школы № 124 г.о.Самара на тему «Батик» Автор: © Куприна Евгения Владимировна учитель МХК и истории изобразительного искусства МОУ школы № 124 г.о. Самара
Творческие работы учащихся художественного отделения МОУ школы № 124 г.о.Самара на тему «Батик» Автор: © Куприна Евгения Владимировна учитель МХК и истории изобразительного искусства МОУ школы № 124 г.о. Самара Облицовка - фреска
Облицовка - фреска Проект по продвижению комплекса ГТО
Проект по продвижению комплекса ГТО Пример оформления экспликации
Пример оформления экспликации Олимпйские игры современности
Олимпйские игры современности ТЕПЛОИЗОЛЯЦИОННЫЕ МАТЕРИАЛЫ И ИЗДЕЛИЯ Неорганические
ТЕПЛОИЗОЛЯЦИОННЫЕ МАТЕРИАЛЫ И ИЗДЕЛИЯ Неорганические  Практическое занятие 1. Пароли.
Практическое занятие 1. Пароли. Общественное движение в 80-90-ые годы XIX века
Общественное движение в 80-90-ые годы XIX века Информационная система по управлению складскими запасами предприятия
Информационная система по управлению складскими запасами предприятия Презентация Рынок недвижимости
Презентация Рынок недвижимости  Презентация на тему "Кленчинг (стискивание зубов) и парафункция" - скачать презентации по Медицине
Презентация на тему "Кленчинг (стискивание зубов) и парафункция" - скачать презентации по Медицине Годы священной войны Великая Отечественная война 1941-1945 гг.
Годы священной войны Великая Отечественная война 1941-1945 гг. Патология белой крови
Патология белой крови Деятельность учителя в рамках школы информационной компетенции. Конюхова О.А. учитель английского языка
Деятельность учителя в рамках школы информационной компетенции. Конюхова О.А. учитель английского языка Усилители электрических сигналов
Усилители электрических сигналов ДИСТОЦИЯ ПЛЕЧИКОВ ПЛОДА
ДИСТОЦИЯ ПЛЕЧИКОВ ПЛОДА Сцепление. Назначение сцепления
Сцепление. Назначение сцепления история бух учета
история бух учета Аудит бизнес плана и порядок проведения
Аудит бизнес плана и порядок проведения  Дорога жизни.
Дорога жизни.  Engetronics. Internet datacenter
Engetronics. Internet datacenter Рабочая тетрадь
Рабочая тетрадь Аукцион в электронной форме
Аукцион в электронной форме Стратегическое управление в образовании
Стратегическое управление в образовании Математическое моделирование транспортных процессов и принципы их построения
Математическое моделирование транспортных процессов и принципы их построения