- Цикл ди и три карбоновых кислот

Содержание
- 2. ИСТОЧНИКИ АЦЕТИЛ-КоА- основного субстрата цикла Кребса Преимущественным источником ацетил-SКоА является: пировиноградная кислота (ПВК, пируват). ПВК –
- 3. Химическая структура коэнзима А (SН-КоА) Ацетил-коэнзимА (ац-SКоА) – это комплекс ацетильного остатка и свободного кофермента А.
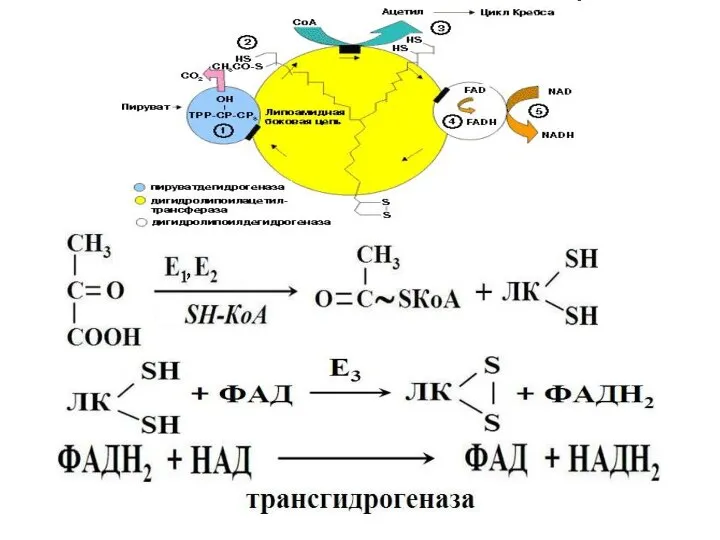
- 4. Превращение пирувата в ацетил-КоА осуществляется в ходе окислительного декарбоксилирования пируватдегидрогеназным комплексом. Происходит в матриксе митохондрий
- 5. Строение ПДГ В состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза.
- 7. Продукты полного окисления ПВК с участием ПДК:
- 8. РЕГУЛЯЦИЯ АКТИВНОСТИ ПИРУВАТДЕГИДРОГЕНАЗНОГО КОМПЛЕКСА ИНГИБИТОРЫ: -ацетил-КоА -НАДН -АТФ АКТИВАТОРЫ: + АДФ +пируват +НАД+ +КоА
- 9. Цикл трикарбоновых кислот, цикл Кребса, цикл лимонной кислоты, цитратный цикл Ганс Адольф Кребс ацетил-КоА + 3НАД
- 10. ОБРАЗОВАНИЕ ЦИТРАТА В реакции образования цитрата углеродный атом метильной труппы ацетил-КоА связывается с карбонильной группой оксалоацетата;
- 11. ПРЕВРАЩЕНИЕ ЦИТРАТА В ИЗОЦИТРАТ Фермент, катализирующий эту реакцию, назван аконитазой по промежуточному продукту, цис-аконитовой кислоте, которая
- 12. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ИЗОЦИТРАТА Изолимонная кислота (ИЗОЦИТРАТ) дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы. В ходе изоцитратдегидрогеназной реакции изолимонная
- 13. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ α -КЕТОГЛУТАРАТА α-кетоглутарат подвергается окислительному декарбоксилированию с образованием в качестве конечных продуктов сукцинил-КоА, СО2
- 14. ПРЕВРАЩЕНИЕ СУКЦИНИЛ-КоА В СУКЦИНАТ Сукцинил-КоА - высокоэнергетическое соединение. В митохондриях разрыв тиоэфирной связи сукцинил-КоА сопряжён с
- 15. ДЕГИДРИРОВАНИЕ СУКЦИНАТА Сукцинат превращается в фумарат под действием сукцинатдегидрогеназы. Этот фермент - флавопротеин, молекула которого содержит
- 16. ОБРАЗОВАНИЕ МАЛАТА ИЗ ФУМАРАТА Образование малата происходит при участии фермента фумаразы, расположенном в матриксе митохондрий. Фумаразу
- 17. ДЕГИДРИРОВАНИЕ МАЛАТА В заключительной стадии цитратного цикла малат дегидрируется с образованием оксалоацетата. Реакцию катализирует NAD-зависимая малатдегидрогеназа,
- 19. ФУНКЦИИ ЦИКЛА ТРИКАРБОНОВЫХ КИСЛОТ ИНТЕГРАТИВНАЯ КАТАБОЛИЧЕСКАЯ АНАБОЛИЧЕСКАЯ ЭНЕРГЕТИЧЕСКАЯ
- 21. Скачать презентацию
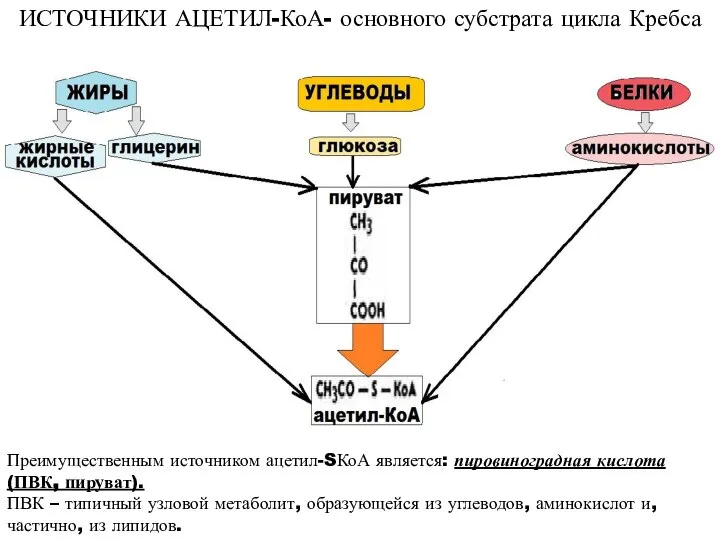
ИСТОЧНИКИ АЦЕТИЛ-КоА- основного субстрата цикла Кребса
Преимущественным источником ацетил-SКоА является: пировиноградная кислота
ИСТОЧНИКИ АЦЕТИЛ-КоА- основного субстрата цикла Кребса
Преимущественным источником ацетил-SКоА является: пировиноградная кислота
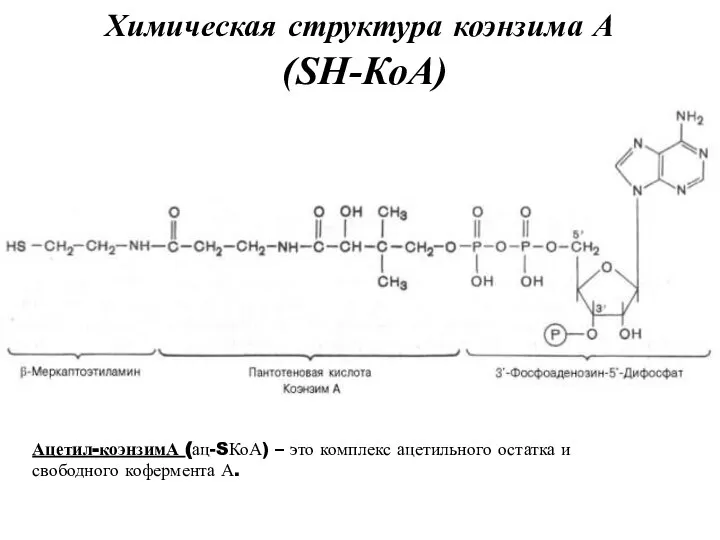
Химическая структура коэнзима А
(SН-КоА)
Ацетил-коэнзимА (ац-SКоА) – это комплекс ацетильного остатка
Химическая структура коэнзима А
(SН-КоА)
Ацетил-коэнзимА (ац-SКоА) – это комплекс ацетильного остатка
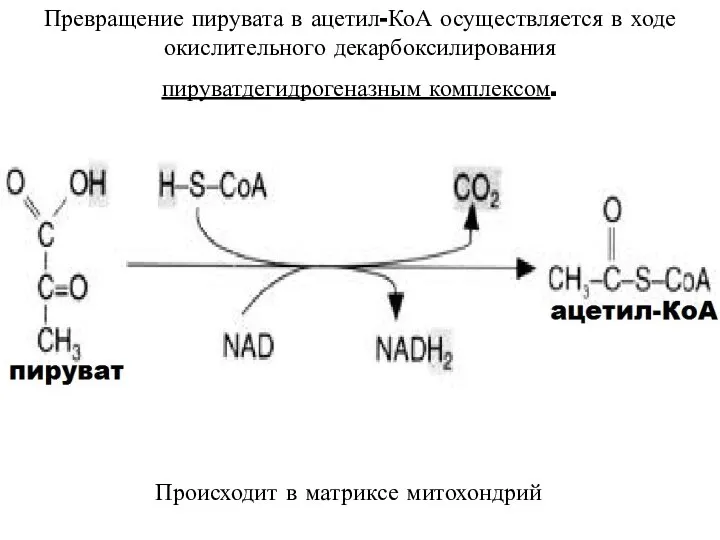
Превращение пирувата в ацетил-КоА осуществляется в ходе окислительного декарбоксилирования
пируватдегидрогеназным комплексом.
Превращение пирувата в ацетил-КоА осуществляется в ходе окислительного декарбоксилирования
пируватдегидрогеназным комплексом.
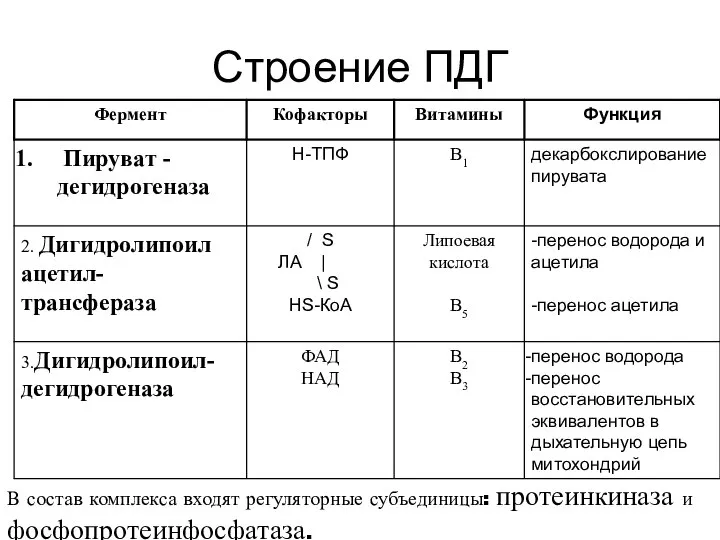
Строение ПДГ
В состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза.
Строение ПДГ
В состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза.
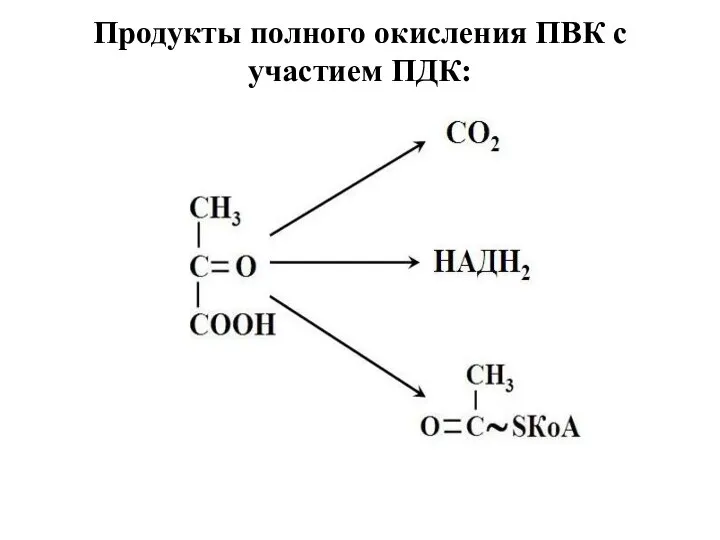
Продукты полного окисления ПВК с участием ПДК:
Продукты полного окисления ПВК с участием ПДК:
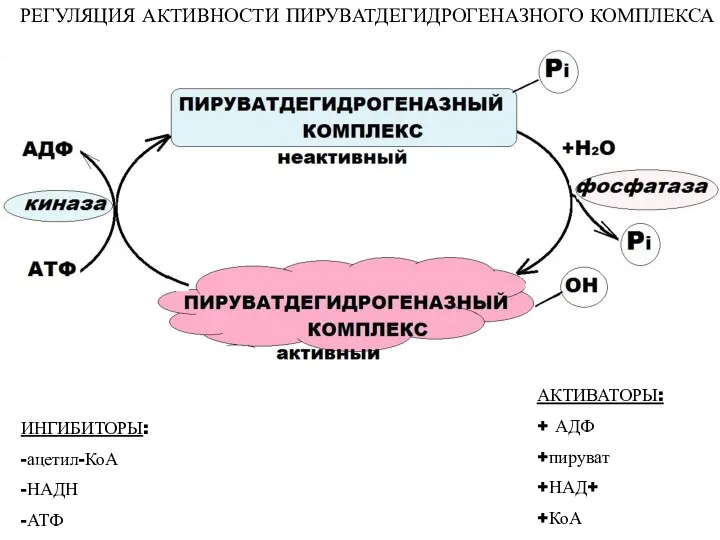
РЕГУЛЯЦИЯ АКТИВНОСТИ ПИРУВАТДЕГИДРОГЕНАЗНОГО КОМПЛЕКСА
ИНГИБИТОРЫ:
-ацетил-КоА
-НАДН
-АТФ
АКТИВАТОРЫ:
+ АДФ
+пируват
+НАД+
+КоА
РЕГУЛЯЦИЯ АКТИВНОСТИ ПИРУВАТДЕГИДРОГЕНАЗНОГО КОМПЛЕКСА
ИНГИБИТОРЫ:
-ацетил-КоА
-НАДН
-АТФ
АКТИВАТОРЫ:
+ АДФ
+пируват
+НАД+
+КоА
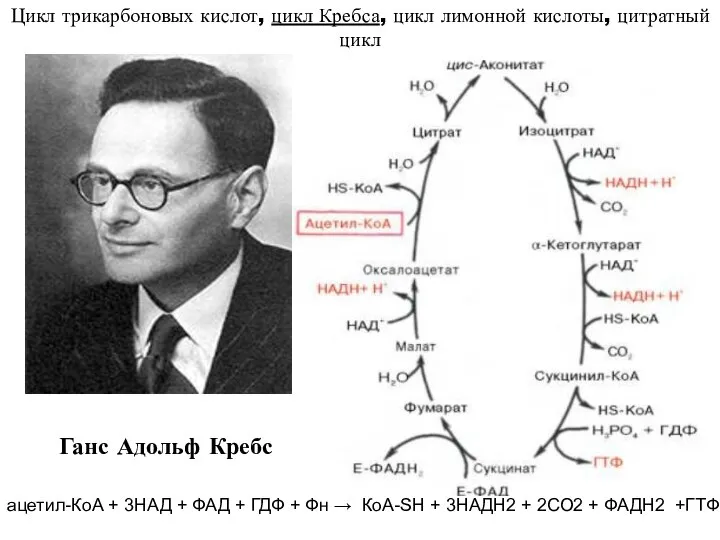
Цикл трикарбоновых кислот, цикл Кребса, цикл лимонной кислоты, цитратный цикл
Ганс Адольф
Цикл трикарбоновых кислот, цикл Кребса, цикл лимонной кислоты, цитратный цикл
Ганс Адольф
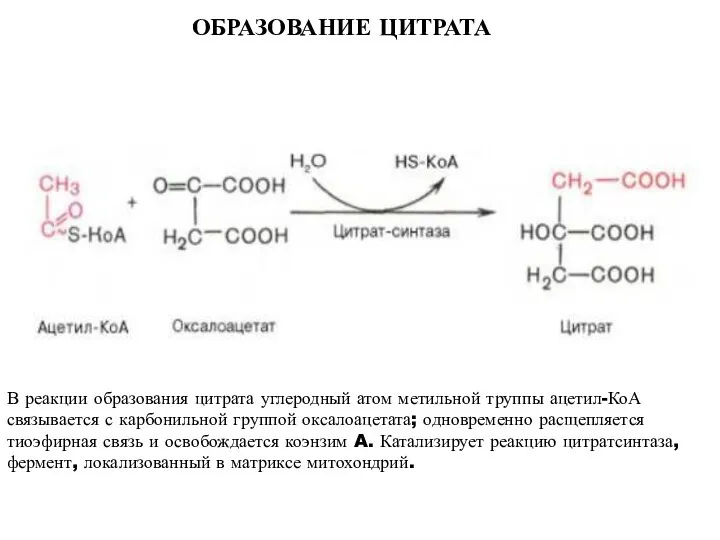
ОБРАЗОВАНИЕ ЦИТРАТА
В реакции образования цитрата углеродный атом метильной труппы ацетил-КоА связывается
ОБРАЗОВАНИЕ ЦИТРАТА
В реакции образования цитрата углеродный атом метильной труппы ацетил-КоА связывается
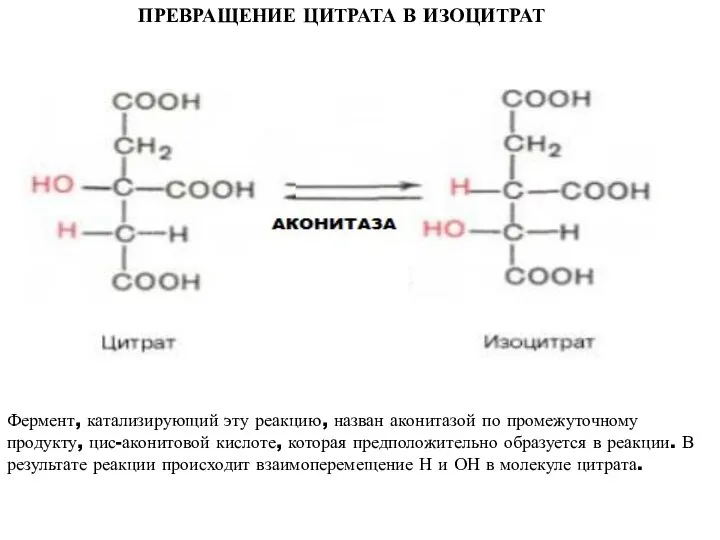
ПРЕВРАЩЕНИЕ ЦИТРАТА В ИЗОЦИТРАТ
Фермент, катализирующий эту реакцию, назван аконитазой по промежуточному
ПРЕВРАЩЕНИЕ ЦИТРАТА В ИЗОЦИТРАТ
Фермент, катализирующий эту реакцию, назван аконитазой по промежуточному
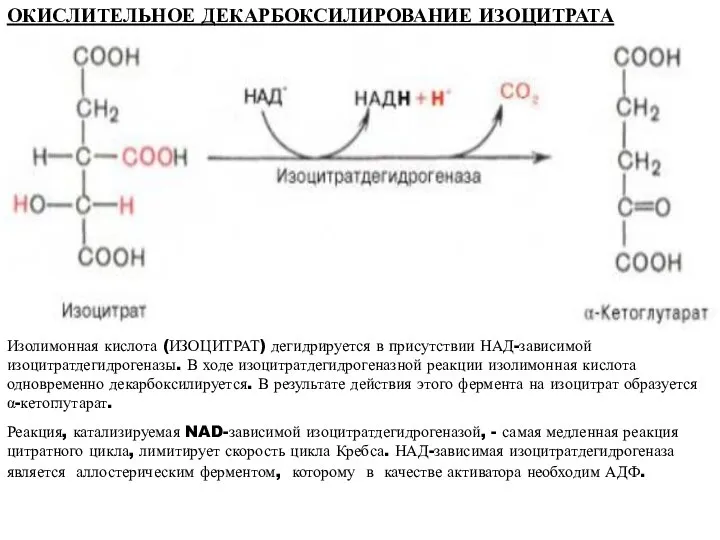
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ИЗОЦИТРАТА
Изолимонная кислота (ИЗОЦИТРАТ) дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы. В
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ИЗОЦИТРАТА
Изолимонная кислота (ИЗОЦИТРАТ) дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы. В
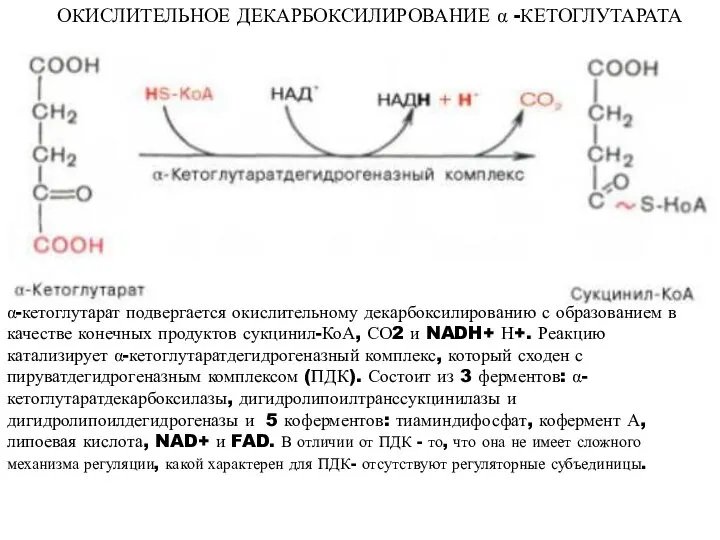
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ α -КЕТОГЛУТАРАТА
α-кетоглутарат подвергается окислительному декарбоксилированию с образованием в качестве
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ α -КЕТОГЛУТАРАТА
α-кетоглутарат подвергается окислительному декарбоксилированию с образованием в качестве

ПРЕВРАЩЕНИЕ СУКЦИНИЛ-КоА В СУКЦИНАТ
Сукцинил-КоА - высокоэнергетическое соединение. В митохондриях разрыв тиоэфирной
ПРЕВРАЩЕНИЕ СУКЦИНИЛ-КоА В СУКЦИНАТ
Сукцинил-КоА - высокоэнергетическое соединение. В митохондриях разрыв тиоэфирной
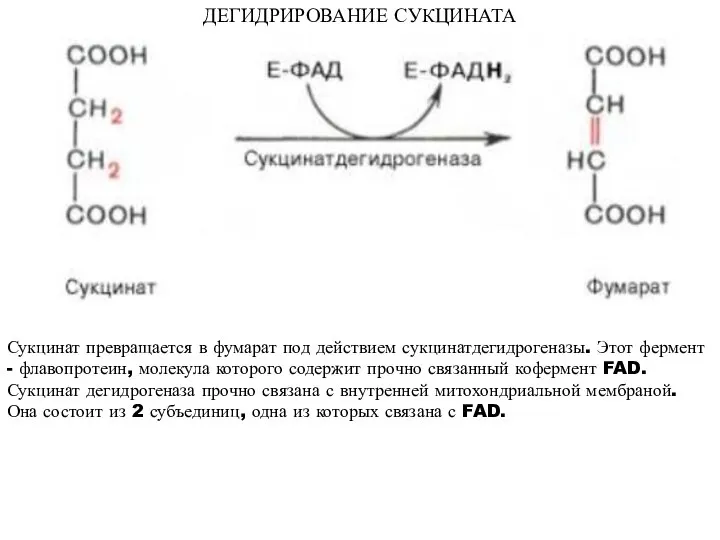
ДЕГИДРИРОВАНИЕ СУКЦИНАТА
Сукцинат превращается в фумарат под действием сукцинатдегидрогеназы. Этот фермент -
ДЕГИДРИРОВАНИЕ СУКЦИНАТА
Сукцинат превращается в фумарат под действием сукцинатдегидрогеназы. Этот фермент -
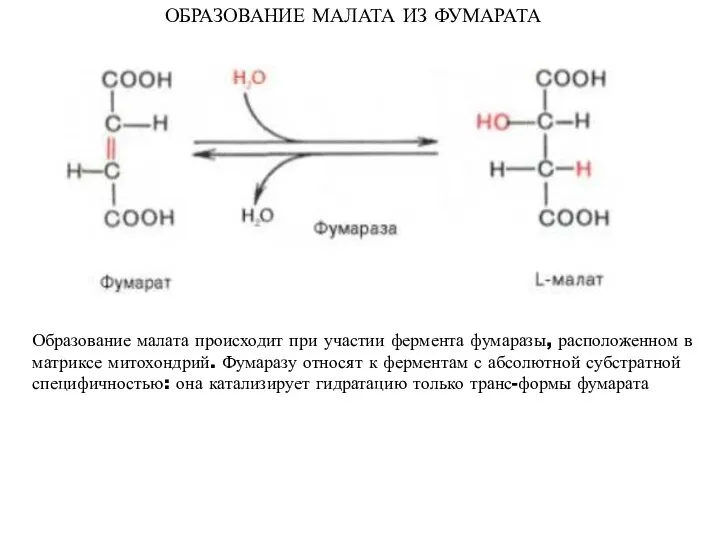
ОБРАЗОВАНИЕ МАЛАТА ИЗ ФУМАРАТА
Образование малата происходит при участии фермента фумаразы, расположенном
ОБРАЗОВАНИЕ МАЛАТА ИЗ ФУМАРАТА
Образование малата происходит при участии фермента фумаразы, расположенном
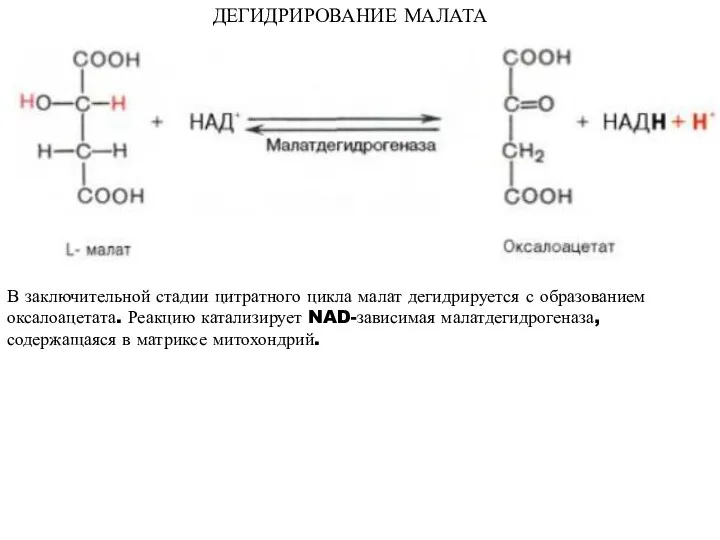
ДЕГИДРИРОВАНИЕ МАЛАТА
В заключительной стадии цитратного цикла малат дегидрируется с образованием оксалоацетата.
ДЕГИДРИРОВАНИЕ МАЛАТА
В заключительной стадии цитратного цикла малат дегидрируется с образованием оксалоацетата.
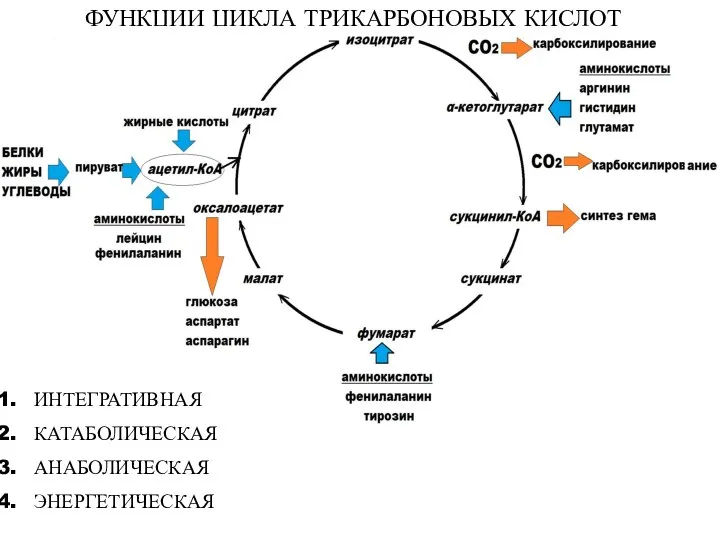
ФУНКЦИИ ЦИКЛА ТРИКАРБОНОВЫХ КИСЛОТ
ИНТЕГРАТИВНАЯ
КАТАБОЛИЧЕСКАЯ
АНАБОЛИЧЕСКАЯ
ЭНЕРГЕТИЧЕСКАЯ
ФУНКЦИИ ЦИКЛА ТРИКАРБОНОВЫХ КИСЛОТ
ИНТЕГРАТИВНАЯ
КАТАБОЛИЧЕСКАЯ
АНАБОЛИЧЕСКАЯ
ЭНЕРГЕТИЧЕСКАЯ
 Стратегическое планирование
Стратегическое планирование  Внеклассное занятие - презентация для начальной школы_
Внеклассное занятие - презентация для начальной школы_ Экономико-правовые аспекты рынка ПО
Экономико-правовые аспекты рынка ПО Презентация на тему "Профессия врач" - скачать презентации по Медицине
Презентация на тему "Профессия врач" - скачать презентации по Медицине Фильмы по мотивам Карфагена
Фильмы по мотивам Карфагена Қазіргі қазақ өлеңінің тілі
Қазіргі қазақ өлеңінің тілі Формы и методы информационной войны в современных условиях
Формы и методы информационной войны в современных условиях АКСЕЛЕРАЦИЯ ДЕЦЕЛЕРАЦИЯ
АКСЕЛЕРАЦИЯ ДЕЦЕЛЕРАЦИЯ  Возможности с Rejuvital
Возможности с Rejuvital Соединение болтом
Соединение болтом Презентация
Презентация  Природные ресурсы 11 класс power
Природные ресурсы 11 класс power Структурированные типы данных-Pascal Записи - C-С++
Структурированные типы данных-Pascal Записи - C-С++ Кинематика передач
Кинематика передач Париж
Париж Tajemnica szczęścia
Tajemnica szczęścia Предприниматель -специалист на все руки. Выполнила: Зайченко Елена, ученица 9 «в» класса; Преподаватель: Сизоненко О.В.
Предприниматель -специалист на все руки. Выполнила: Зайченко Елена, ученица 9 «в» класса; Преподаватель: Сизоненко О.В. ФГОС НОО - презентация_
ФГОС НОО - презентация_ Нормы права и правовые отношения
Нормы права и правовые отношения программы на языке Pascal
программы на языке Pascal Операционный (производственный, хозяйственный) рычаг (леверидж) Подготовила ст. преподаватель А.М.Лопарева
Операционный (производственный, хозяйственный) рычаг (леверидж) Подготовила ст. преподаватель А.М.Лопарева  Актуальные вопросы гастроэнтерологии
Актуальные вопросы гастроэнтерологии  Доклад про монастырь "Оптина Пустынь"
Доклад про монастырь "Оптина Пустынь" 2008-2009 учебный год Анализ учебно-воспитательного процесса в начальной школе
2008-2009 учебный год Анализ учебно-воспитательного процесса в начальной школе DConf 2016, Berlin Ethan Watson, Senior Generalist Programmer
DConf 2016, Berlin Ethan Watson, Senior Generalist Programmer Бог и человек
Бог и человек Технико-экономические параметры эффективных кровельных покрытий для Республики Башкортостан
Технико-экономические параметры эффективных кровельных покрытий для Республики Башкортостан Доказательство правильности программ. Структурное программирование
Доказательство правильности программ. Структурное программирование