ФИЗИОЛОГИЯ КЛЕТКИ наука, изучающая закономерности и свойства функционирования клеток животных, растений, простейших независ
-
ФИЗИОЛОГИЯ КЛЕТКИ наука, изучающая закономерности и свойства функционирования клеток животных, растений, простейших независ

Содержание
- 2. Аспекты изучения «Физиологии клетки» ДВИГАТЕЛЬНАЯ ФУНКЦИЯ КЛЕТОК РЕЦЕПТОРНАЯ ФУНКЦИЯ КЛЕТОК ФУНКЦИИ ИЗМЕНЕНИЯ рН В КЛЕТКЕ. ПЕРЕКИСНОЕ
- 3. ДВИГАТЕЛЬНАЯ ФУНКЦИЯ КЛЕТОК
- 4. Контрактильные механизмы (приводящие к сокращению или движению): мышечное сокращение движение ресничек и жгутиков расхождение хромосом при
- 5. Цитоскелет клетки Микротрубочки (~ 25 нм) Тубулин микрофиламенты (6-7 нм) Актин промежуточные филаменты (8-10 нм) Кератин,
- 6. Микрофиламенты (Актиновые филаменты)
- 7. Актин имеет участки связывания с: Mg2+ АТФ 3 типа актинов: α- актин характерен для мышечных клеток
- 8. Процесс полимеризации-деполимеризации G-актина Нуклеация – образование затравок (тримеров) Элонгация – рост полимеров засчет присоединения к обоим
- 9. Полимеризация и деполимеризация актина G - актин F - актин _ + профиллин G–актин – глобулярный
- 10. Типы АСБ Белки, ингибирующие полимеризацию актина: профиллин Кэпирующие белки. Кэпирование «+»-конца F–актина : гельзолин, виллин, фрагмин.
- 11. Некоторые соединения имитируют действие АСБ, ингибируя полимеризацию и деполимеризацию актина Цитохалазины (низкомолекулярные гетероциклические соединения, вторичные метаболиты
- 12. Функции микрофиламентов Образование сократимого кольца при цитотомии Перемещение клетки Эндо- и экзоцитоз Участие в свертывании крови
- 13. Структура миозина палочковидная хвостовая часть две глобулярные головки: -тяжелые цепи (200 кДа) - легкие цепи (18
- 14. МЫШЕЧНОЕ ВОЛОКНО
- 15. ВИД САРКОМЕРА
- 16. САРКОМЕР
- 17. Сократимый аппарат Миозин Актин
- 18. СОКРАЩЕНИЕ САРКОМЕРА
- 19. Действие Са2+ во время активации миофибриллы 1- «шейка» миозина 2- «головка» миозина 3-актиновый мономер 4-тропонин 5-тропомиозин
- 20. Микротрубочки
- 21. Микротрубочки Основной белок микротрубочек – тубулин. У всех эукариотических клеток он представляет собой гетеродимер, состоящий из
- 22. Полимеризация тубулина Нуклеация - образование затравок – олигомеры тубулина, содержащие несколько десятков молекул; образуются нитевидные структуры
- 23. ЗАРИСУЙТЕ СХЕМУ ОБРАЗОВАНИЯ МИКРОТРУБОЧЕК
- 24. Полимеризация тубулина
- 25. Соединения, блокирующие полимеризацию-деполимеризацию МТ колхицин, (растительный алкалоид), связывается с тубулиновым димером и «+»-концом микротрубочек, препятствуя полимеризации;
- 26. Строение реснички (жгутика)
- 27. Центриоли клеточного центра - Состоят из 9-ти триплетов микротрубочек, располагающихся строго по переферии. - Триплеты микротрубочек
- 28. ЗАРИСУЙТЕ КЛЕТОЧНЫЙ ЦЕНТР
- 29. Центриоли клеточного центра
- 30. Моторные белки МТ Динеины и кинезины - эти молекулы одним концом прикрепляются сбоку к микротрубочке и
- 31. Взаимодействие МТ с моторными белками митохондрия лизосома МТ МТ кинезин динеин _ _ + +
- 32. Промежуточные филаменты
- 33. Представляют собой фибриллы диаметром 8-12 нм. В клетке локализуются в виде трехмерной сети преимущественно в околоядерной
- 34. Промежуточные филаменты Кератины (эпителии) Виментин (соединит. ткани) Десмин (мышечние ткани) Нейрофибриллы (нервная ткань)
- 35. Промежуточные филаменты в составе различают: - центральный консервативный (одинаковый у всех) домен. Имеет палочковидную форму и
- 36. Этапы полимеризации белков промежуточных филаментов 1 — отдельная молекула; 2 — димер; 3 — тетрамер-протофиламент; 4,
- 37. Циклоз (течение цитоплазмы) лучше всего выражено в растительных клетках, но его можно наблюдать и у простейших,
- 38. Циклоз В основе циклоза лежит функционирование сократительных белков, обладающих АТФ-азной активностью, например может быть обусловлено сокращением
- 39. Амебоидное движение Амебоидным движением обладают самые разнообразные клетки – простейшие из класса саркодовых, зооспоры, некоторые сперматозоиды
- 40. Амебоидное движение состоит в медленном перетекании тела клетки по субстрату и осуществляется благодаря внутриклеточному течению цитоплазмы
- 41. Поляризованный движущийся фибробласт Красным цветом окрашены микрофиламенты и их пучки, связанные с флуоресцирующими антителами к актину,
- 42. Мерцательное движение Мерцательное движение обусловлено деятельностью специализированных органелл – выростов клетки, называемых ресничками (многочисленны и короткие,
- 43. Реснички и жгутики Основной тип движения жгутиков – ундулирующее или волнообразное (синусоидальное, распространяющееся в одном направлении,
- 45. Скачать презентацию

Аспекты изучения «Физиологии клетки»
ДВИГАТЕЛЬНАЯ ФУНКЦИЯ КЛЕТОК
РЕЦЕПТОРНАЯ ФУНКЦИЯ КЛЕТОК
ФУНКЦИИ
Аспекты изучения «Физиологии клетки»
ДВИГАТЕЛЬНАЯ ФУНКЦИЯ КЛЕТОК
РЕЦЕПТОРНАЯ ФУНКЦИЯ КЛЕТОК
ФУНКЦИИ

ДВИГАТЕЛЬНАЯ
ФУНКЦИЯ КЛЕТОК
ДВИГАТЕЛЬНАЯ
ФУНКЦИЯ КЛЕТОК

Контрактильные механизмы
(приводящие к сокращению или движению):
мышечное сокращение
движение ресничек и
Контрактильные механизмы
(приводящие к сокращению или движению):
мышечное сокращение
движение ресничек и
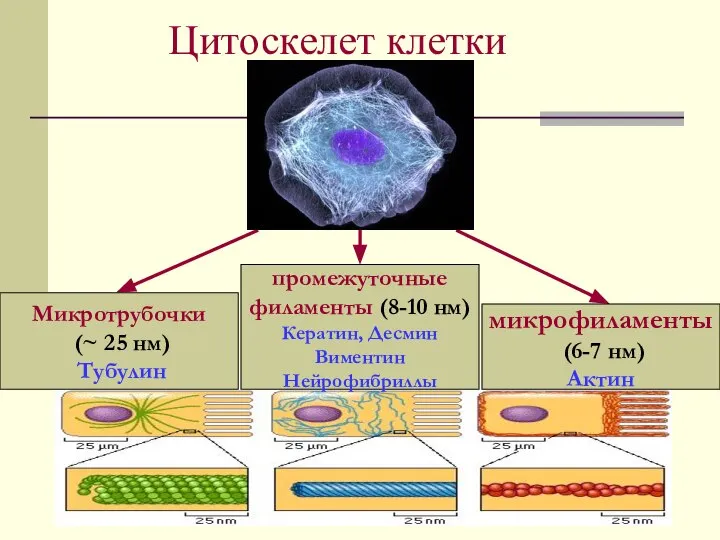
Цитоскелет клетки
Микротрубочки
(~ 25 нм)
Тубулин
микрофиламенты
(6-7 нм)
Актин
промежуточные
филаменты (8-10
Цитоскелет клетки
Микротрубочки
(~ 25 нм)
Тубулин
микрофиламенты
(6-7 нм)
Актин
промежуточные
филаменты (8-10

Микрофиламенты
(Актиновые филаменты)
Микрофиламенты
(Актиновые филаменты)
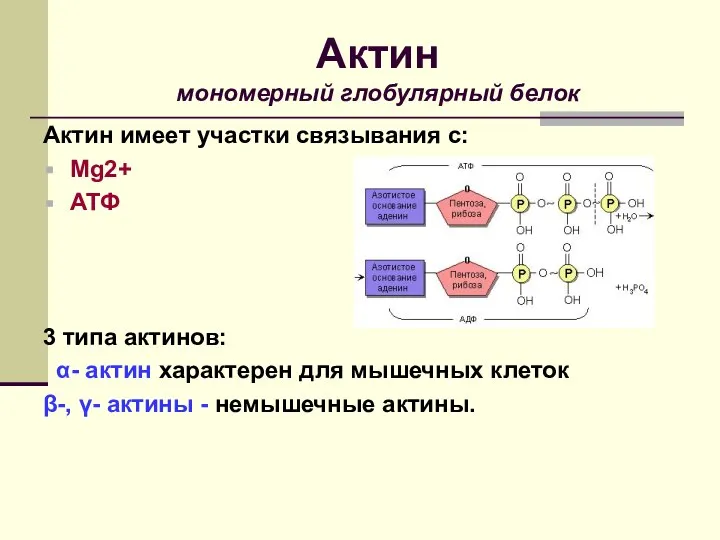
Актин имеет участки связывания с:
Mg2+
АТФ
3 типа актинов:
α- актин
Актин имеет участки связывания с:
Mg2+
АТФ
3 типа актинов:
α- актин

Процесс полимеризации-деполимеризации
G-актина
Нуклеация – образование затравок (тримеров)
Элонгация – рост полимеров
Процесс полимеризации-деполимеризации
G-актина
Нуклеация – образование затравок (тримеров)
Элонгация – рост полимеров
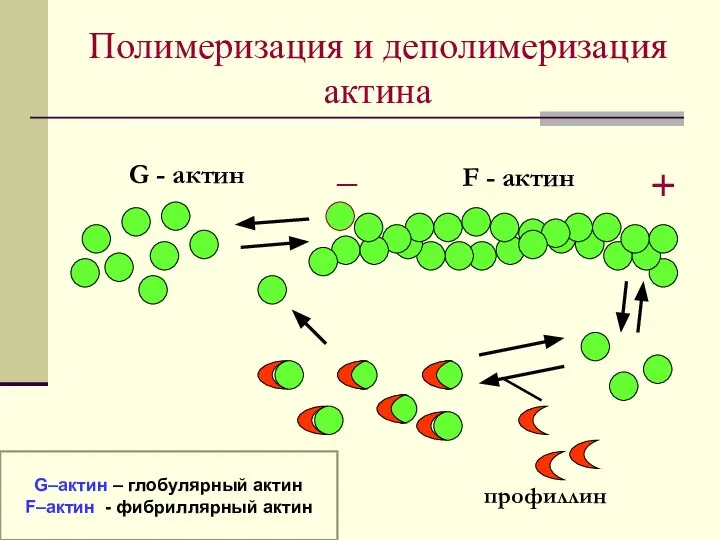
Полимеризация и деполимеризация актина
G - актин
F - актин
_
+
профиллин
G–актин – глобулярный
Полимеризация и деполимеризация актина
G - актин
F - актин
_
+
профиллин
G–актин – глобулярный

Типы АСБ
Белки, ингибирующие полимеризацию актина: профиллин
Кэпирующие белки. Кэпирование «+»-конца F–актина :
Типы АСБ
Белки, ингибирующие полимеризацию актина: профиллин
Кэпирующие белки. Кэпирование «+»-конца F–актина :

Некоторые соединения имитируют действие АСБ, ингибируя полимеризацию и деполимеризацию актина
Цитохалазины (низкомолекулярные
Некоторые соединения имитируют действие АСБ, ингибируя полимеризацию и деполимеризацию актина
Цитохалазины (низкомолекулярные

Функции микрофиламентов
Образование сократимого кольца при цитотомии
Перемещение клетки
Эндо- и экзоцитоз
Участие в свертывании
Функции микрофиламентов
Образование сократимого кольца при цитотомии
Перемещение клетки
Эндо- и экзоцитоз
Участие в свертывании

Структура миозина
палочковидная хвостовая часть
две глобулярные головки:
-тяжелые
Структура миозина
палочковидная хвостовая часть
две глобулярные головки:
-тяжелые
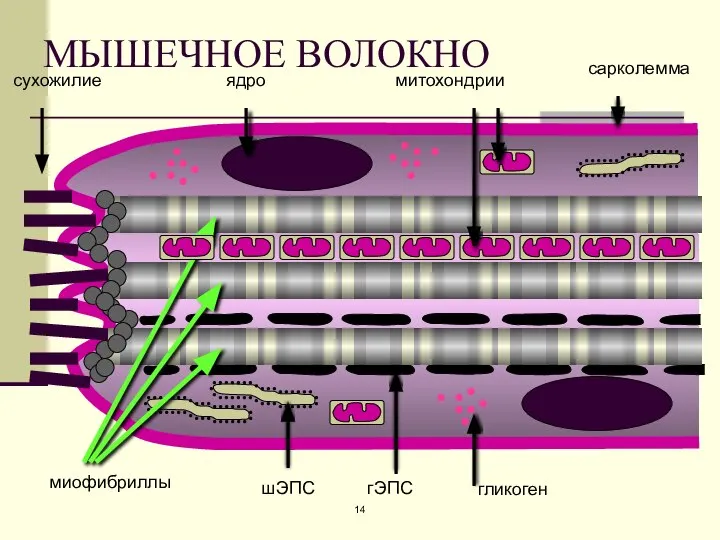
МЫШЕЧНОЕ ВОЛОКНО
МЫШЕЧНОЕ ВОЛОКНО
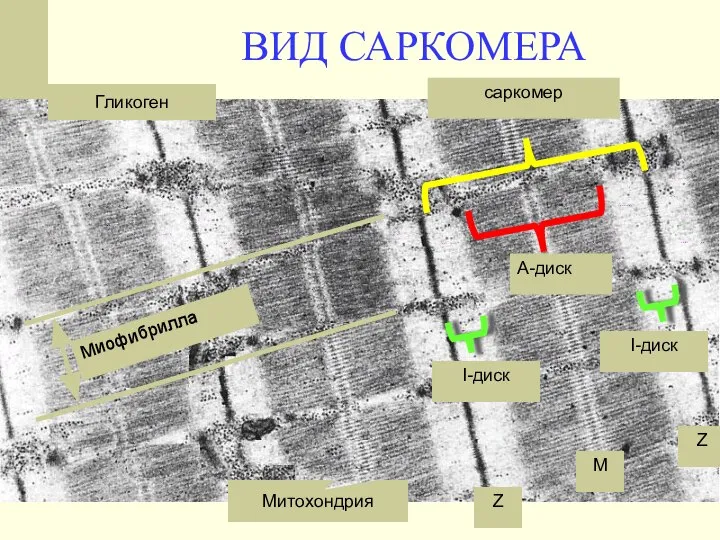
ВИД САРКОМЕРА
ВИД САРКОМЕРА
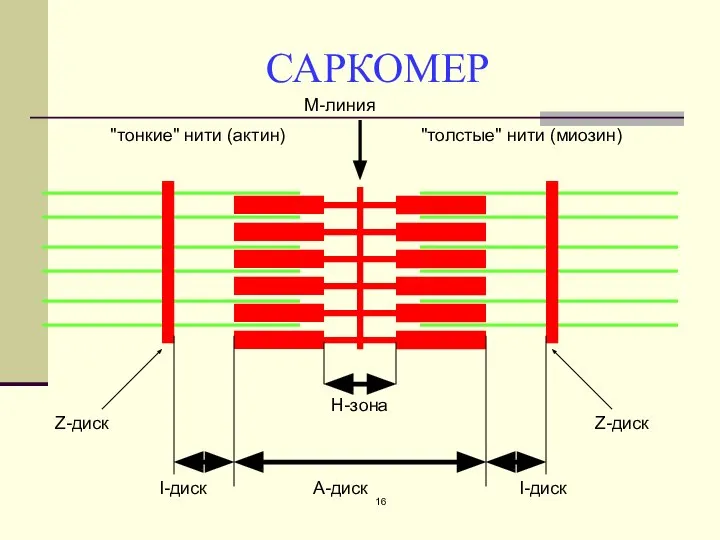
САРКОМЕР
САРКОМЕР
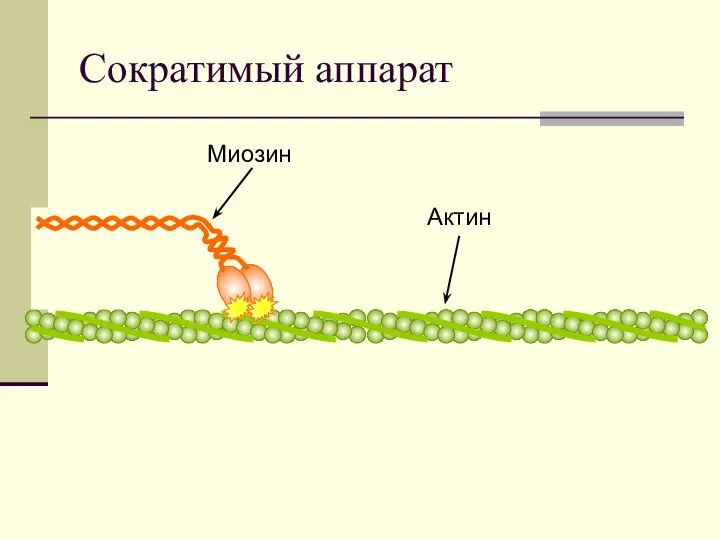
Сократимый аппарат
Миозин
Актин
Сократимый аппарат
Миозин
Актин
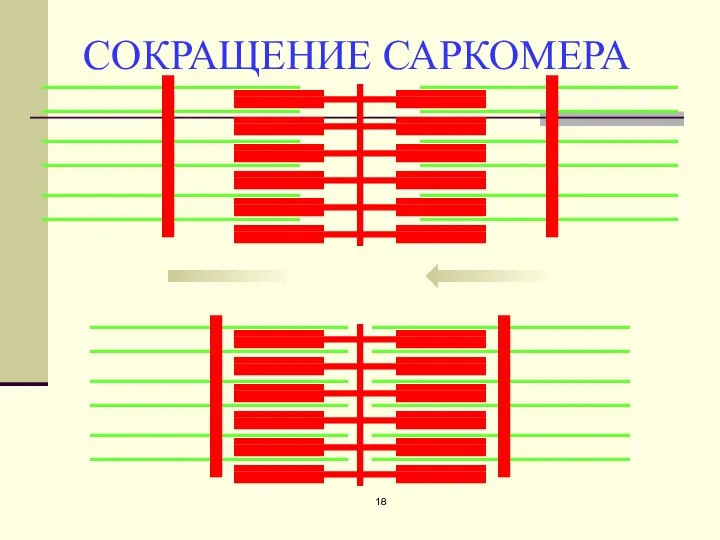
СОКРАЩЕНИЕ САРКОМЕРА
СОКРАЩЕНИЕ САРКОМЕРА
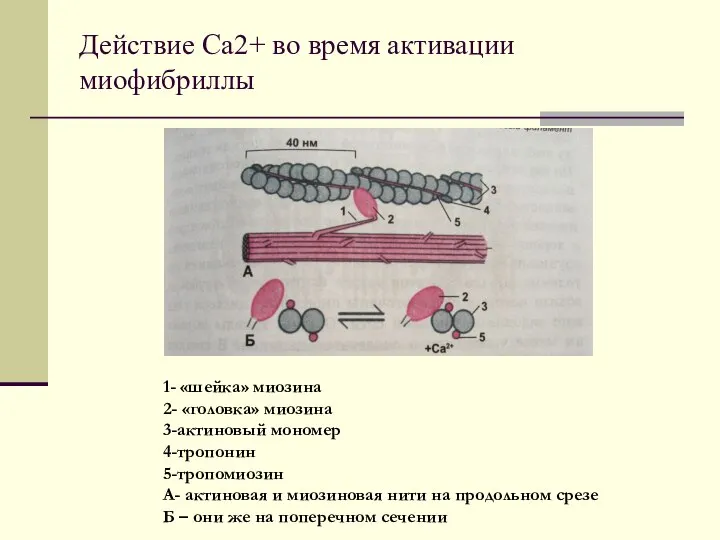
Действие Са2+ во время активации миофибриллы
1- «шейка» миозина
2- «головка» миозина
3-актиновый мономер
4-тропонин
5-тропомиозин
А-
Действие Са2+ во время активации миофибриллы
1- «шейка» миозина
2- «головка» миозина
3-актиновый мономер
4-тропонин
5-тропомиозин
А-

Микротрубочки
Микротрубочки
Микротрубочки
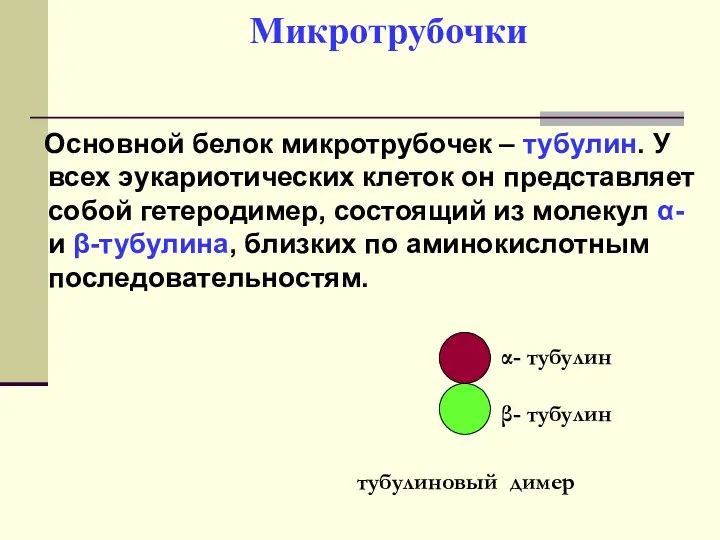
Основной белок микротрубочек – тубулин. У всех эукариотических клеток
Микротрубочки
Основной белок микротрубочек – тубулин. У всех эукариотических клеток

Полимеризация тубулина
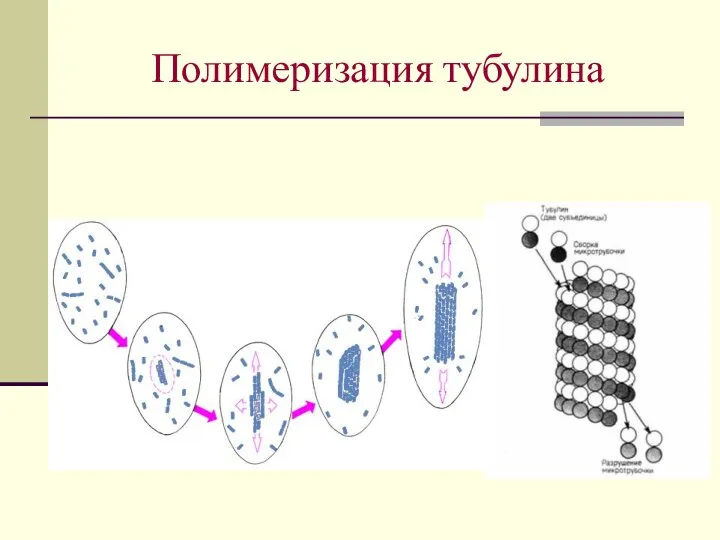
Нуклеация - образование затравок – олигомеры тубулина, содержащие несколько десятков
Полимеризация тубулина
Нуклеация - образование затравок – олигомеры тубулина, содержащие несколько десятков

ЗАРИСУЙТЕ СХЕМУ ОБРАЗОВАНИЯ МИКРОТРУБОЧЕК
ЗАРИСУЙТЕ СХЕМУ ОБРАЗОВАНИЯ МИКРОТРУБОЧЕК
Полимеризация тубулина
Полимеризация тубулина

Соединения, блокирующие полимеризацию-деполимеризацию МТ
колхицин, (растительный алкалоид), связывается с тубулиновым димером
Соединения, блокирующие полимеризацию-деполимеризацию МТ
колхицин, (растительный алкалоид), связывается с тубулиновым димером
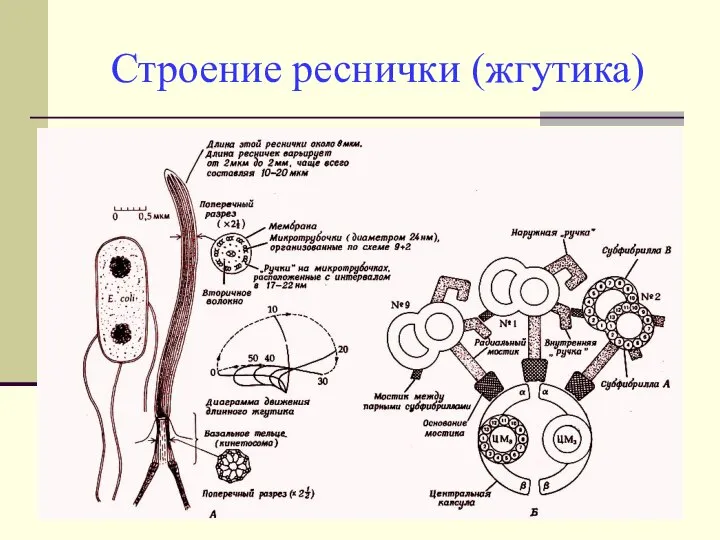
Строение реснички (жгутика)
Строение реснички (жгутика)

Центриоли клеточного центра
- Состоят из 9-ти триплетов микротрубочек, располагающихся строго по
Центриоли клеточного центра
- Состоят из 9-ти триплетов микротрубочек, располагающихся строго по

ЗАРИСУЙТЕ КЛЕТОЧНЫЙ ЦЕНТР
ЗАРИСУЙТЕ КЛЕТОЧНЫЙ ЦЕНТР
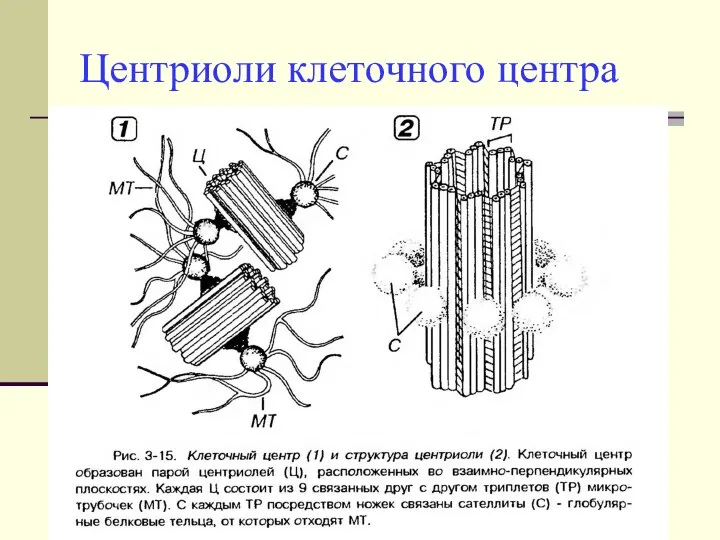
Центриоли клеточного центра
Центриоли клеточного центра
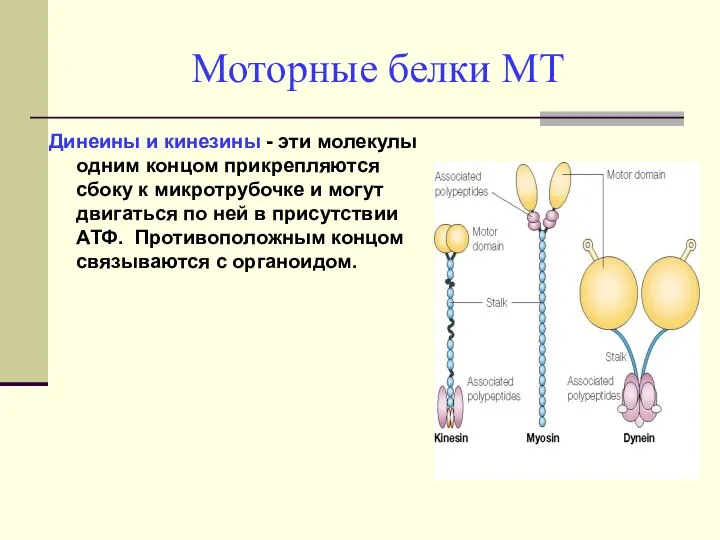
Моторные белки МТ
Динеины и кинезины - эти молекулы одним концом прикрепляются
Моторные белки МТ
Динеины и кинезины - эти молекулы одним концом прикрепляются
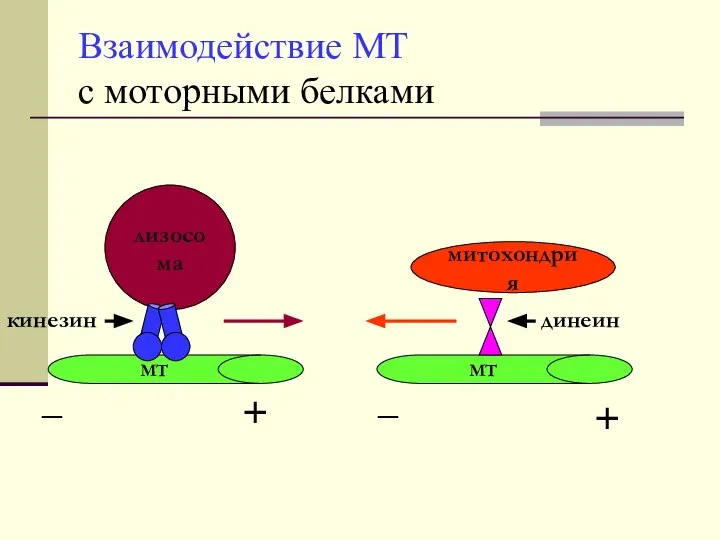
Взаимодействие МТ
с моторными белками
митохондрия
лизосома
МТ
МТ
кинезин
динеин
_
_
+
+
Взаимодействие МТ
с моторными белками
митохондрия
лизосома
МТ
МТ
кинезин
динеин
_
_
+
+

Промежуточные филаменты
Промежуточные филаменты

Представляют собой фибриллы диаметром 8-12 нм.
В клетке локализуются в
Представляют собой фибриллы диаметром 8-12 нм.
В клетке локализуются в
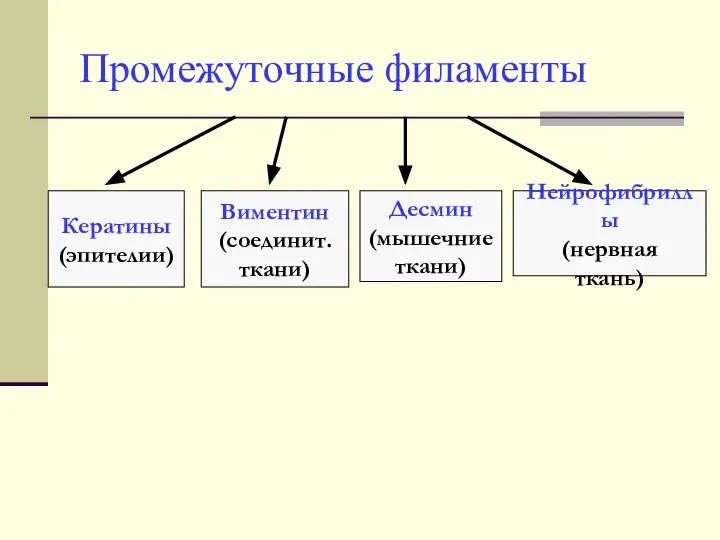
Промежуточные филаменты
Кератины
(эпителии)
Виментин
(соединит.
ткани)
Десмин
(мышечние
ткани)
Нейрофибриллы
(нервная
ткань)
Промежуточные филаменты
Кератины
(эпителии)
Виментин
(соединит.
ткани)
Десмин
(мышечние
ткани)
Нейрофибриллы
(нервная
ткань)

Промежуточные филаменты
в составе различают:
- центральный консервативный (одинаковый у всех) домен. Имеет
Промежуточные филаменты
в составе различают:
- центральный консервативный (одинаковый у всех) домен. Имеет
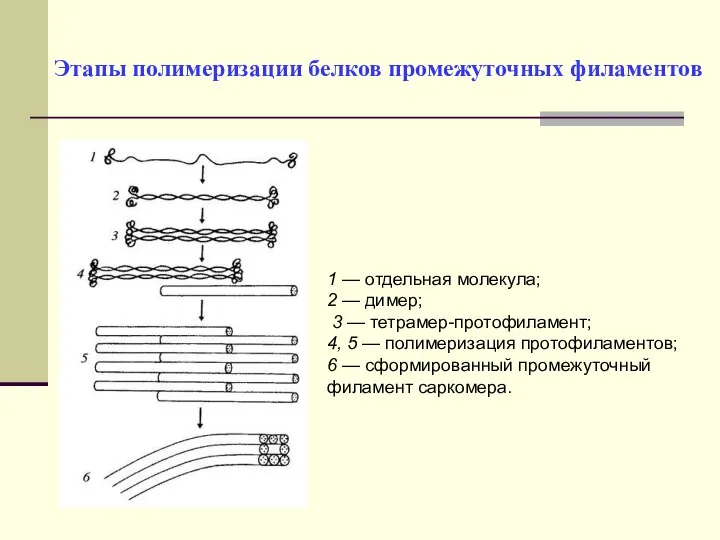
Этапы полимеризации белков промежуточных филаментов
1 — отдельная молекула;
2 — димер;
Этапы полимеризации белков промежуточных филаментов
1 — отдельная молекула;
2 — димер;

Циклоз (течение цитоплазмы) лучше всего выражено в растительных клетках, но его
Циклоз (течение цитоплазмы) лучше всего выражено в растительных клетках, но его

Циклоз
В основе циклоза лежит функционирование сократительных белков, обладающих АТФ-азной активностью, например
Циклоз
В основе циклоза лежит функционирование сократительных белков, обладающих АТФ-азной активностью, например

Амебоидное движение
Амебоидным движением обладают самые разнообразные клетки – простейшие из
Амебоидное движение
Амебоидным движением обладают самые разнообразные клетки – простейшие из

Амебоидное движение состоит в медленном перетекании тела клетки по субстрату
Амебоидное движение состоит в медленном перетекании тела клетки по субстрату
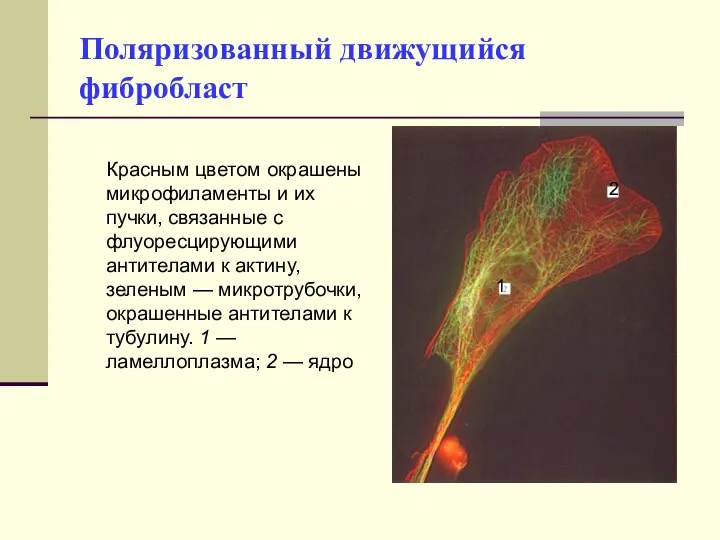
Поляризованный движущийся фибробласт
Красным цветом окрашены микрофиламенты и их пучки, связанные
Поляризованный движущийся фибробласт
Красным цветом окрашены микрофиламенты и их пучки, связанные
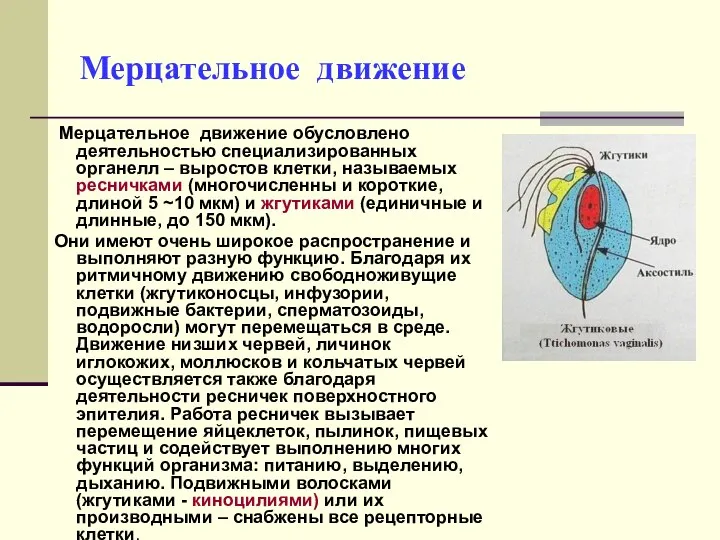
Мерцательное движение
Мерцательное движение обусловлено деятельностью специализированных органелл – выростов клетки,
Мерцательное движение
Мерцательное движение обусловлено деятельностью специализированных органелл – выростов клетки,

Реснички и жгутики
Основной тип движения жгутиков – ундулирующее или волнообразное
Реснички и жгутики
Основной тип движения жгутиков – ундулирующее или волнообразное
 Разоблачение культа личности Сталина
Разоблачение культа личности Сталина  Социально –экономические проблемы, стоящие перед ДНР, в современных условиях
Социально –экономические проблемы, стоящие перед ДНР, в современных условиях Презентация на тему "право на образование" - скачать презентации по Педагогике
Презентация на тему "право на образование" - скачать презентации по Педагогике Ученья корень горек, да плод сладок - презентация для начальной школы_
Ученья корень горек, да плод сладок - презентация для начальной школы_ Презентация Население и трудовые ресурсы России
Презентация Население и трудовые ресурсы России Презентация Классификация товаров в соответствии с ТН ВЭД на единой территории таможенного союза
Презентация Классификация товаров в соответствии с ТН ВЭД на единой территории таможенного союза Презентация Оценка маштабов теневой экономики
Презентация Оценка маштабов теневой экономики Лондон и его достопримечательности
Лондон и его достопримечательности Жизнь пресного водоема - презентация для начальной школы_
Жизнь пресного водоема - презентация для начальной школы_ Հայաստանյան ֆուտբոլի զարգացման Ռազմավարություն 2016 - 2025
Հայաստանյան ֆուտբոլի զարգացման Ռազմավարություն 2016 - 2025 Презентация Ценообразующие факторы сельскохозяйственной продукции
Презентация Ценообразующие факторы сельскохозяйственной продукции  BoardDiag Test Environment & Coverage
BoardDiag Test Environment & Coverage Наглядное пособие к уроку окружающего мира Тема: «Путешествие по миру» 3 класс
Наглядное пособие к уроку окружающего мира Тема: «Путешествие по миру» 3 класс Cпасибо, тебе Азбука!
Cпасибо, тебе Азбука! Потоки, анімація, багатовіконність на Java
Потоки, анімація, багатовіконність на Java Карпова Елена Геннадиевна, учитель математики ГОУСОШ №562
Карпова Елена Геннадиевна, учитель математики ГОУСОШ №562 Многофайловые проекты
Многофайловые проекты Методика технико-тактической подготовки таэквондистов-юниоров с использованием комплекса специальных упражнений
Методика технико-тактической подготовки таэквондистов-юниоров с использованием комплекса специальных упражнений Необходимость государственного регулирования образования. Лекция 3
Необходимость государственного регулирования образования. Лекция 3 Общество и государство. Гражданское общество в Российской Федерации
Общество и государство. Гражданское общество в Российской Федерации Телеоло́гия
Телеоло́гия  Презентация Система источников международного права : понятие, виды, применение в практике международных отношений
Презентация Система источников международного права : понятие, виды, применение в практике международных отношений  Звук [а] Буквы А,а - презентация для начальной школы_
Звук [а] Буквы А,а - презентация для начальной школы_ квадратичная форма
квадратичная форма  Кейнсианская теория Выполнила: Лазарюк А.С. Группа Т-104
Кейнсианская теория Выполнила: Лазарюк А.С. Группа Т-104 Понятие и сущность логистики
Понятие и сущность логистики Развитие исследований: от гена до функции
Развитие исследований: от гена до функции The middle east. Religion and ethnic groups
The middle east. Religion and ethnic groups