- ИОННЫЕ КАНАЛЫ

Содержание
- 2. БИОЛОГИЧЕСКИ ВАЖНЫЕ МЕТАЛЛЫ ИОННЫЙ ГОМЕОСТАЗ КЛЕТКИ
- 3. СОДЕРЖАНИЕ ОСНОВНЫХ ИОНОВ В КЛЕТКАХ И ВНЕКЛЕТОЧНОЙ ЖИДКОСТИ НЕКОТОРЫХ ЖИВОТНЫХ В СРАВНЕНИИ С ИОННЫМ СОСТАВОМ МОРСКОЙ
- 5. ХАРАКТЕРИСТИКА ИОНОВ НАТРИЯ И КАЛИЯ ШИРОКО РАСПРОСТРАНЕНЫ В НЕЖИВОЙ ПРИРОДЕ НИЗКИЙ ПОТЕНЦИАЛ ИОНИЗАЦИИ ВНЕШНЕГО ЭЛЕКТРОНА ОБРАЗОВАВШИЙСЯ
- 6. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА ИОНОВ НАТРИЯ И КАЛИЯ ЭНЕРГИЯ ГИДРАТАЦИИ (ПРИ КОМНАТНОЙ ТЕМПЕРАТУРЕ) ДЛЯ ИОНОВ НАТРИЯ +1,03 кДж/моль
- 7. СВОЙСТВА ИОННЫХ КАНАЛОВ СЕЛЕКТИВНОСТЬ НАСЫЩЕНИЕ КОНКУРЕНТНОЕ ПОДАВЛЕНИЕ ИОННЫХ КАНАЛОВ ИОНАМИ-БЛОКАТОРАМИ КАНАЛЫ – УПРАВЛЯЕМЫЕ СТРУКТУРЫ НЕЗАВИСИМОСТЬ РАБОТЫ
- 8. КЛАССИФИКАЦИОННЫЕ ПРИЗНАКИ КАНАЛОВ СЕЛЕКТИВНОСТЬ (Na-, K-, Ca- КАНАЛЫ и др.) ПРОВОДИМОСТЬ (КАНАЛЫ ВЫСОКОЙ, СРЕДНЕЙ, НИЗКОЙ ПРОВОДИМОСТИ)
- 9. ОСНОВНЫЕ ТИПЫ ИОННЫХ КАНАЛОВ ПО МЕХАНИЗМУ УПРАВЛЕНИЯ ЭЛЕКТРОУПРАВЛЯЕМЫЕ КАНАЛЫ ХЕМОУПРАВЛЯЕМЫЕ КАНАЛЫ МЕХАНОУПРАВЛЯЕМЫЕ КАНАЛЫ
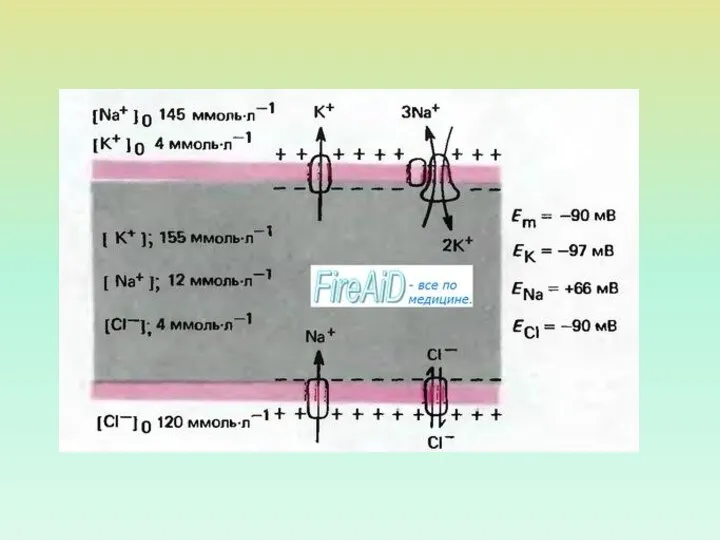
- 10. ПОТЕНЦИАЛОЗАВИСИМЫЕ ИОННЫЕ КАНАЛЫ
- 11. МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ Na-КАНАЛЫ К-КАНАЛЫ
- 12. МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ СТЕРИЧЕСКИЙ ФАКТОР ЭНЕРГЕТИЧЕСКИЙ ФАКТОР
- 13. ВЗАИМОДЕЙСТВИЕ ИОНОВ С ВОДОЙ ХАРАКТЕРИЗУЕТ ЭНЕРГИЯ ГИДРАТАЦИИ
- 14. РАДИУСЫ ИОНОВ И ЭНЕРГИЯ ГИДРАТАЦИЯ
- 15. СТЕРИЧЕСКИЙ ФАКТОР ГИПОТЕЗА МУЛЛИНЗА: ИОН МОЖЕТ ПРОЙТИ ЧЕРЕЗ ПОРУ, ЕСЛИ ЕГО РАДИУС С ОДНИМ СЛОЕМ ГИДРАТАЦИИ
- 16. ЭНЕРГЕТИЧЕСКИЙ ФАКТОР СЕЛЕКТИВНОСТИ ЭЛЕКТРОСТАТИЧЕСКАЯ ТЕОРИЯ ЭЙЗЕНМАНА Ионный обмен для катионселективных стеклянных электродов: катионы А+ и В+
- 17. РЕАКЦИЯ СДВИГАЕТСЯ В СТОРОНУ ВЗАИМОДЕЙСТВИЯ ИОНА А+ СО СТЕКЛОМ, ЕСЛИ ΔGA(ВОДНАЯ СРЕДА→СТЕКЛО)
- 18. Анионная группировка сферическая с радиусом rА Катион также сферический с радиусом rС Энергия взаимодействия катиона и
- 19. ЗНАЧЕНИЕ rА ВЕЛИКО АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СЛАБОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА U МАЛА ДЛЯ ВСЕХ КАТИОНОВ
- 20. ЗНАЧЕНИЕ rА МАЛО АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СИЛЬНОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА U ВЕЛИКА И ПРЕВЫШАЕТ ЭНЕРГИЮ
- 21. ПРОНИЦАЕМОСТЬ Na+КАНАЛА ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ PLi : PNa : PK : PRb = 1,1 : 1
- 22. МОДЕЛЬ ХИЛЛЕ ОСНОВАНА НА ИЗУЧЕНИИ ПРОНИЦАЕМОСТИ Na-КАНАЛА ДЛЯ ОРГАНИЧЕСКИХ ИОНОВ Na >ГИДРОКСИЛАМИН >ГИДРАЗИН >АММОНИЙ≈ФОРМАМИДИН ≈ГУАНИДИН >>МЕТИЛАММОНИЙ
- 23. СЕЛЕКТИВНЫЙ ФИЛЬТР НАТРИЕВОГО КАНАЛА ИОН НАТРИЯ В СЕЛЕКТИВНОМ ФИЛЬТРЕ С МОЛЕКУЛОЙ ВОДЫ Н2О Н2О
- 24. МОДЕЛЬ ХИЛЛЕ ВХОД В КАНАЛ – ПРЯМОУГОЛЬНАЯ ПОРА 0,3х0,5 нм СЕЛЕКТИВНЫЙ ФИЛЬТР В Nа-КАНАЛЕ СОДЕРЖИТ 8
- 25. СВОБОДНЫЙ ИОН (1) СВЯЗЫВАЕТСЯ (2) И ДЕСОЛЬВАТИРУЕТСЯ (23) ЗАТЕМ РЕСОЛЬВАТИРУЕТСЯ НА ВНУТРЕННЕЙ СТОРОНЕ МЕМБРАНЫ (3) И
- 26. МОДЕЛЬ НАТРИЕВОГО КАНАЛА 1 – БЕЛКИ КАНАЛА 2 – СЕЛЕКТИВНЫЙ ФИЛЬТР, СОДЕРЖАЩИЙ, ВИДИМО, КАРБОКСИЛЬНУЮ ГРУППУ 3
- 27. ВОЗМОЖНАЯ ТРАНСМЕМБРАННАЯ ОРГАНИЗАЦИЯ НАТРИЕВОГО КАНАЛА АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ИМЕЮТ ЧЕТЫРЕ ВЫСОКОГОМОЛОГИЧНЫХ ДОМЕНА, КАЖДЫЙ ИЗ НИХ СОДЕРЖИТ 6
- 28. ТРАНСМЕМБРАННАЯ СТРУКТУРА НАТРИЕВОГО КАНАЛА А – субъединица, формирующая канал Б – схема канала с селективным фильтром
- 29. БЛОКАТОРЫ НАТРИЕВОГО КАНАЛА
- 30. ТЕТРОДОТОКСИН
- 31. САКСИТОКСИН ДИНОФЛАГЕЛЛЯТЫ
- 32. ТЕТРОДОТОКСИН САКСИТОКСИН
- 33. МЕСТНЫЕ АНЕСТЕТИКИ КАК БЛОКАТОРЫ НАТРИЕВЫХ КАНАЛОВ
- 34. МЕХАНИЗМ ДЕЙСТВИЯ МЕСТНЫХ АНЕСТЕТИКОВ А – БЛОКИРУЮТ НАТРИЕВУЮ ПРОВОДИМОСТЬ ПОСРЕДСТВОМ ВЗАИМОДЕЙСТВИЯ С БЕЛКАМИ Б - БЛОКИРУЮТ
- 36. ВОЗДЕЙСТВИЯ, УСТРАНЯЮЩИЕ ИНАКТИВАЦИЮ НАТРИЕВОГО КАНАЛА ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ ПЕПТИДНЫЕ ТОКСИНЫ (ЯД СКОРПИОНА, ЯД АНЕМОН) АЛКОЛОИДНЫЕ НЕЙРОТОКСИНЫ (АКОНИТИН,
- 37. ТетродотоксинСакситоксин Блокируют транспорт Токсины скорпиона, морской анемоны Препятствуют инактивации, усиливают активацию Вератридин, аконитин и др. Вызывают
- 38. КАЛИЕВЫЙ КАНАЛ
- 39. Схематический энергетический профиль К-канала. По оси ординат кинетическая энергия, необходимая для прохождения канала; по оси абсцисс
- 40. ПРОНИЦАЕМОСТЬ КАЛИЕВОГО КАНАЛА ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ НЕ КОРРЕЛИРУЕТ С РАДИУСОМ ИОНА PK : PRb : PCs
- 41. СТРУКТУРА КАЛИЕВОГО КАНАЛА, УСТАНОВЛЕННАЯ С ПОМОЩЬЮ ТЕТРАЭТИЛАММОНИЯ КАЛИЕВЫЙ КАНАЛ АСИММЕТРИЧЕН, ИМЕЕТ ФОРМУ ВОРОНКИ ДИАМЕТР ВЫХОДНОГО ОТВЕРСТИЯ
- 43. ТРАНСМЕМБРАННАЯ ТОПОЛОГИЯ И АРХИТЕКТУРА КАЛИЕВОГО КАНАЛА
- 44. Р. Мак-Киннон Нобелевская премия по химии за 2003 г. присуждена Р. Мак-Киннону за открытия, касающиеся переноса
- 45. Мак-Киннон и его коллеги расшифровали структуру и механизм функционирования нескольких бактериальных белков, каждый из которых формирует
- 46. Рис.1Стереомодель полного KvAP канала и одной субъединицы
- 47. Рис. 3. Модели, иллюстрирующие перенос положительных зарядов канальным калиевым белком в ответ на изменение потенциала на
- 48. Рис. 4. Схема движения “лопастей” калиевого канала и их положение внутри мембраны, когда его ворота закрыты
- 49. ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ МЕХАНОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
- 50. ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
- 51. РЕЦЕПТОРУПРАВЛЯЕМЫЕ Примеры: АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР ГАМК- И ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ В ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ
- 52. АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР
- 55. Са2+-ЗАВИСИМЫЙ КАЛИЕВЫЙ КАНАЛ
- 57. АТФ-ЧУВСТВИТЕЛЬНЫЕ КАЛИЕВЫЕ КАНАЛЫ А Трансмембранная топология двух субъединиц КАТФ канала Б - Схематическое изображение канального комплекса
- 58. В Модель формирования поры КАТФ канала субъединицами (продольное сечение) с формированием селективного фильтра в наружном (1)
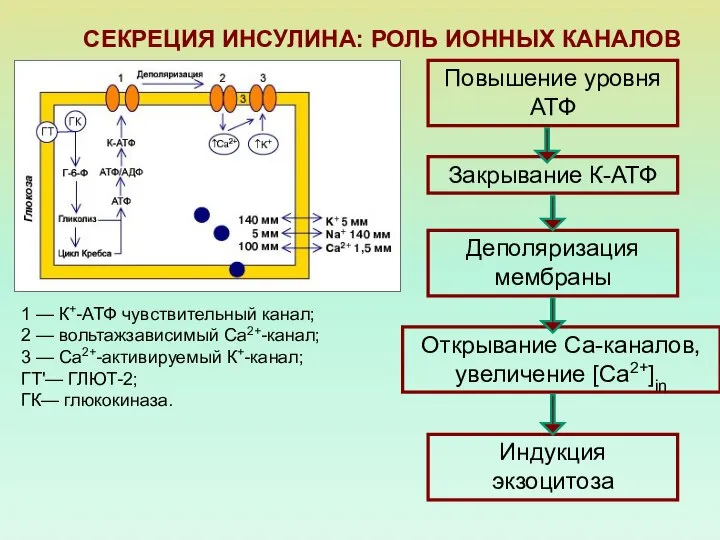
- 59. СЕКРЕЦИЯ ИНСУЛИНА: РОЛЬ ИОННЫХ КАНАЛОВ 1 — К+-АТФ чувствительный канал; 2 — вольтажзависимый Са2+-канал; 3 —
- 61. МЕХАНОЧУВСТВИТЕЛЬНЫЙ КАНАЛ
- 62. Восприятие звука и вибрации, гравитации, ускорения, скорости, давления, прикосновения, изменения формы и объема клетки, ее местоположения
- 63. Системы управления механочувствительным каналом
- 64. А - ориентация белка механочувствительного канала в мембране. Б - 6 субъединиц механочувствительного группируются, образуя компактный
- 65. 2. Управление молекулярными мостиками, которые связаны с одной стороны с цитоскелетом, а с другой – с
- 67. Скачать презентацию
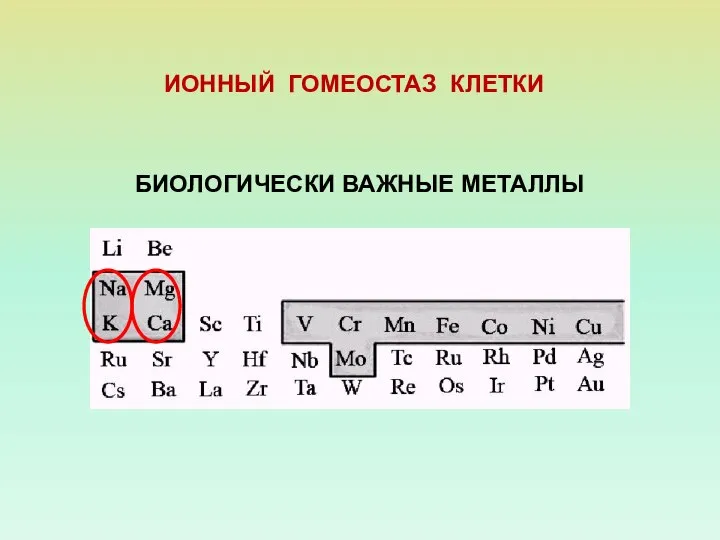
БИОЛОГИЧЕСКИ ВАЖНЫЕ МЕТАЛЛЫ
ИОННЫЙ ГОМЕОСТАЗ КЛЕТКИ
БИОЛОГИЧЕСКИ ВАЖНЫЕ МЕТАЛЛЫ
ИОННЫЙ ГОМЕОСТАЗ КЛЕТКИ
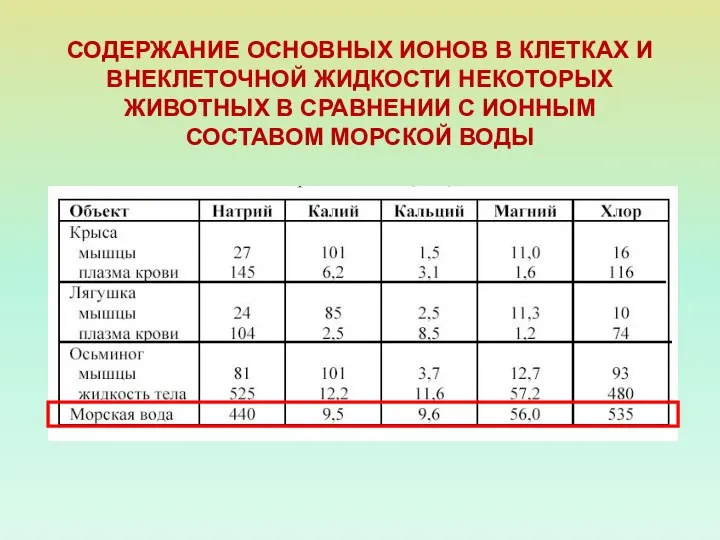
СОДЕРЖАНИЕ ОСНОВНЫХ ИОНОВ В КЛЕТКАХ И ВНЕКЛЕТОЧНОЙ ЖИДКОСТИ НЕКОТОРЫХ ЖИВОТНЫХ В
СОДЕРЖАНИЕ ОСНОВНЫХ ИОНОВ В КЛЕТКАХ И ВНЕКЛЕТОЧНОЙ ЖИДКОСТИ НЕКОТОРЫХ ЖИВОТНЫХ В

ХАРАКТЕРИСТИКА ИОНОВ НАТРИЯ И КАЛИЯ
ШИРОКО РАСПРОСТРАНЕНЫ В НЕЖИВОЙ ПРИРОДЕ
НИЗКИЙ ПОТЕНЦИАЛ ИОНИЗАЦИИ
ХАРАКТЕРИСТИКА ИОНОВ НАТРИЯ И КАЛИЯ
ШИРОКО РАСПРОСТРАНЕНЫ В НЕЖИВОЙ ПРИРОДЕ
НИЗКИЙ ПОТЕНЦИАЛ ИОНИЗАЦИИ

ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА ИОНОВ НАТРИЯ И КАЛИЯ
ЭНЕРГИЯ ГИДРАТАЦИИ (ПРИ КОМНАТНОЙ ТЕМПЕРАТУРЕ)
ДЛЯ ИОНОВ
ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА ИОНОВ НАТРИЯ И КАЛИЯ
ЭНЕРГИЯ ГИДРАТАЦИИ (ПРИ КОМНАТНОЙ ТЕМПЕРАТУРЕ)
ДЛЯ ИОНОВ
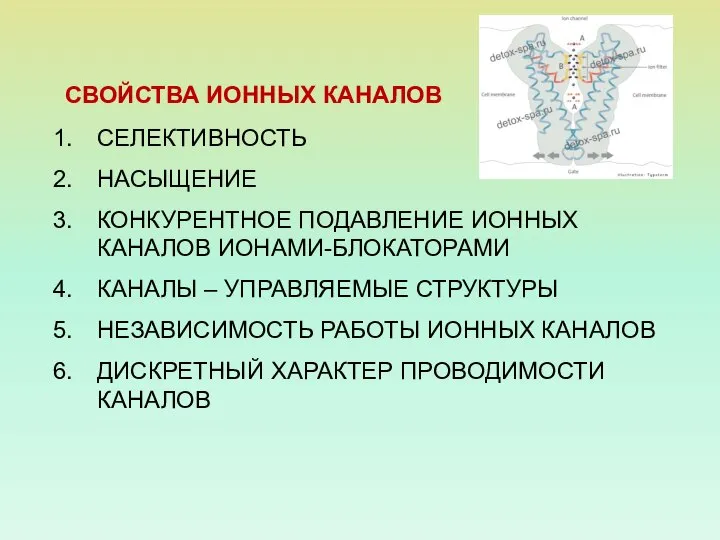
СВОЙСТВА ИОННЫХ КАНАЛОВ
СЕЛЕКТИВНОСТЬ
НАСЫЩЕНИЕ
КОНКУРЕНТНОЕ ПОДАВЛЕНИЕ ИОННЫХ КАНАЛОВ ИОНАМИ-БЛОКАТОРАМИ
КАНАЛЫ – УПРАВЛЯЕМЫЕ СТРУКТУРЫ
НЕЗАВИСИМОСТЬ РАБОТЫ
СВОЙСТВА ИОННЫХ КАНАЛОВ
СЕЛЕКТИВНОСТЬ
НАСЫЩЕНИЕ
КОНКУРЕНТНОЕ ПОДАВЛЕНИЕ ИОННЫХ КАНАЛОВ ИОНАМИ-БЛОКАТОРАМИ
КАНАЛЫ – УПРАВЛЯЕМЫЕ СТРУКТУРЫ
НЕЗАВИСИМОСТЬ РАБОТЫ

КЛАССИФИКАЦИОННЫЕ ПРИЗНАКИ КАНАЛОВ
СЕЛЕКТИВНОСТЬ (Na-, K-, Ca- КАНАЛЫ и др.)
ПРОВОДИМОСТЬ (КАНАЛЫ
КЛАССИФИКАЦИОННЫЕ ПРИЗНАКИ КАНАЛОВ
СЕЛЕКТИВНОСТЬ (Na-, K-, Ca- КАНАЛЫ и др.)
ПРОВОДИМОСТЬ (КАНАЛЫ
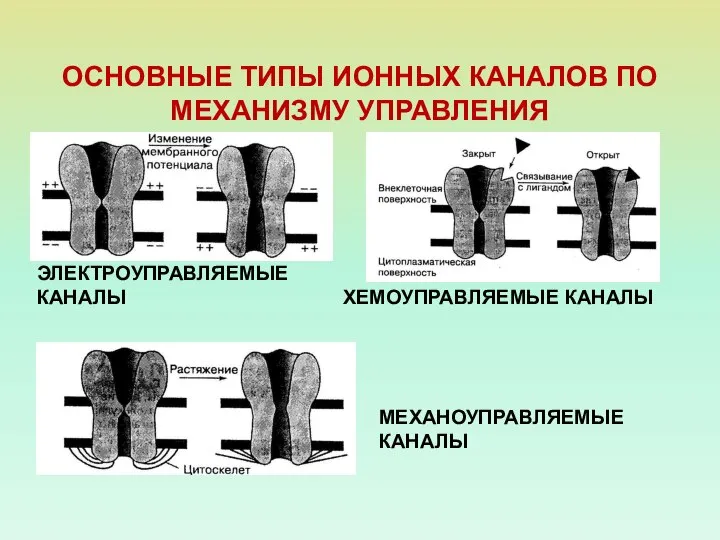
ОСНОВНЫЕ ТИПЫ ИОННЫХ КАНАЛОВ ПО МЕХАНИЗМУ УПРАВЛЕНИЯ
ЭЛЕКТРОУПРАВЛЯЕМЫЕ КАНАЛЫ
ХЕМОУПРАВЛЯЕМЫЕ КАНАЛЫ
МЕХАНОУПРАВЛЯЕМЫЕ КАНАЛЫ
ОСНОВНЫЕ ТИПЫ ИОННЫХ КАНАЛОВ ПО МЕХАНИЗМУ УПРАВЛЕНИЯ
ЭЛЕКТРОУПРАВЛЯЕМЫЕ КАНАЛЫ
ХЕМОУПРАВЛЯЕМЫЕ КАНАЛЫ
МЕХАНОУПРАВЛЯЕМЫЕ КАНАЛЫ

ПОТЕНЦИАЛОЗАВИСИМЫЕ ИОННЫЕ КАНАЛЫ
ПОТЕНЦИАЛОЗАВИСИМЫЕ ИОННЫЕ КАНАЛЫ

МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ
Na-КАНАЛЫ
К-КАНАЛЫ
МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ
Na-КАНАЛЫ
К-КАНАЛЫ

МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ
СТЕРИЧЕСКИЙ ФАКТОР
ЭНЕРГЕТИЧЕСКИЙ ФАКТОР
МЕХАНИЗМЫ СЕЛЕКТИВНОСТИ ИОННЫХ КАНАЛОВ
СТЕРИЧЕСКИЙ ФАКТОР
ЭНЕРГЕТИЧЕСКИЙ ФАКТОР
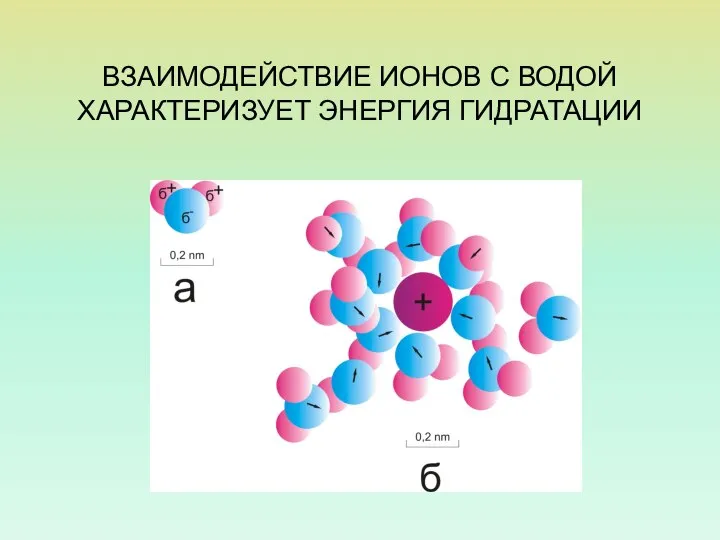
ВЗАИМОДЕЙСТВИЕ ИОНОВ С ВОДОЙ
ХАРАКТЕРИЗУЕТ ЭНЕРГИЯ ГИДРАТАЦИИ
ВЗАИМОДЕЙСТВИЕ ИОНОВ С ВОДОЙ
ХАРАКТЕРИЗУЕТ ЭНЕРГИЯ ГИДРАТАЦИИ
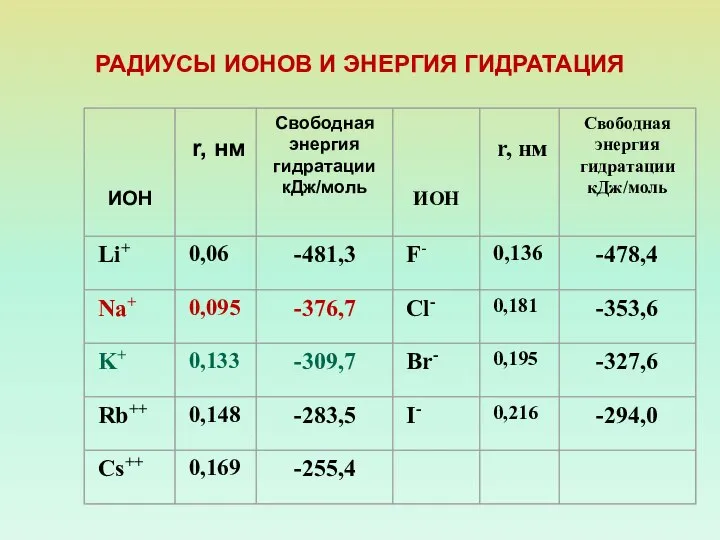
РАДИУСЫ ИОНОВ И ЭНЕРГИЯ ГИДРАТАЦИЯ
РАДИУСЫ ИОНОВ И ЭНЕРГИЯ ГИДРАТАЦИЯ

СТЕРИЧЕСКИЙ ФАКТОР
ГИПОТЕЗА МУЛЛИНЗА:
ИОН МОЖЕТ ПРОЙТИ ЧЕРЕЗ ПОРУ, ЕСЛИ ЕГО РАДИУС
СТЕРИЧЕСКИЙ ФАКТОР
ГИПОТЕЗА МУЛЛИНЗА:
ИОН МОЖЕТ ПРОЙТИ ЧЕРЕЗ ПОРУ, ЕСЛИ ЕГО РАДИУС

ЭНЕРГЕТИЧЕСКИЙ ФАКТОР СЕЛЕКТИВНОСТИ
ЭЛЕКТРОСТАТИЧЕСКАЯ ТЕОРИЯ ЭЙЗЕНМАНА
Ионный обмен для катионселективных стеклянных электродов: катионы
ЭНЕРГЕТИЧЕСКИЙ ФАКТОР СЕЛЕКТИВНОСТИ
ЭЛЕКТРОСТАТИЧЕСКАЯ ТЕОРИЯ ЭЙЗЕНМАНА
Ионный обмен для катионселективных стеклянных электродов: катионы
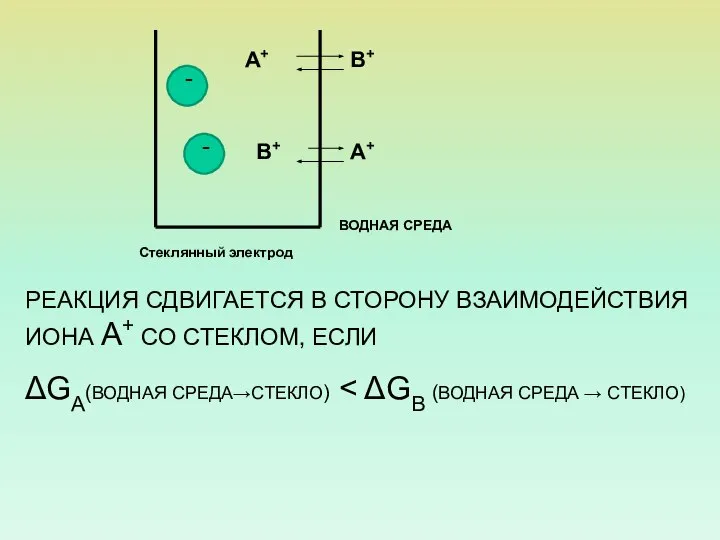
РЕАКЦИЯ СДВИГАЕТСЯ В СТОРОНУ ВЗАИМОДЕЙСТВИЯ ИОНА А+ СО СТЕКЛОМ, ЕСЛИ
ΔGA(ВОДНАЯ
РЕАКЦИЯ СДВИГАЕТСЯ В СТОРОНУ ВЗАИМОДЕЙСТВИЯ ИОНА А+ СО СТЕКЛОМ, ЕСЛИ
ΔGA(ВОДНАЯ

Анионная группировка сферическая с радиусом rА
Катион также сферический с радиусом rС
Энергия
Анионная группировка сферическая с радиусом rА
Катион также сферический с радиусом rС
Энергия

ЗНАЧЕНИЕ rА ВЕЛИКО
АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СЛАБОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА U
ЗНАЧЕНИЕ rА ВЕЛИКО
АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СЛАБОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА U

ЗНАЧЕНИЕ rА МАЛО
АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СИЛЬНОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА
ЗНАЧЕНИЕ rА МАЛО
АНИОННАЯ ГРУППИРОВКА СОЗДАЕТ СИЛЬНОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ, ПОЭТОМУ ВЕЛИЧИНА

ПРОНИЦАЕМОСТЬ Na+КАНАЛА ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ
PLi : PNa : PK : PRb
ПРОНИЦАЕМОСТЬ Na+КАНАЛА ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ
PLi : PNa : PK : PRb

МОДЕЛЬ ХИЛЛЕ ОСНОВАНА НА ИЗУЧЕНИИ ПРОНИЦАЕМОСТИ Na-КАНАЛА ДЛЯ ОРГАНИЧЕСКИХ ИОНОВ
Na >ГИДРОКСИЛАМИН
МОДЕЛЬ ХИЛЛЕ ОСНОВАНА НА ИЗУЧЕНИИ ПРОНИЦАЕМОСТИ Na-КАНАЛА ДЛЯ ОРГАНИЧЕСКИХ ИОНОВ
Na >ГИДРОКСИЛАМИН
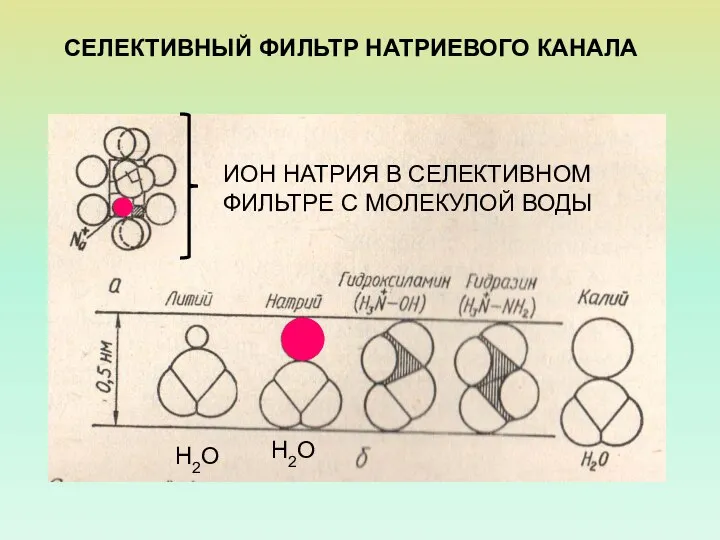
СЕЛЕКТИВНЫЙ ФИЛЬТР НАТРИЕВОГО КАНАЛА
ИОН НАТРИЯ В СЕЛЕКТИВНОМ ФИЛЬТРЕ С МОЛЕКУЛОЙ ВОДЫ
Н2О
Н2О
СЕЛЕКТИВНЫЙ ФИЛЬТР НАТРИЕВОГО КАНАЛА
ИОН НАТРИЯ В СЕЛЕКТИВНОМ ФИЛЬТРЕ С МОЛЕКУЛОЙ ВОДЫ
Н2О
Н2О

МОДЕЛЬ ХИЛЛЕ
ВХОД В КАНАЛ – ПРЯМОУГОЛЬНАЯ ПОРА 0,3х0,5 нм
СЕЛЕКТИВНЫЙ ФИЛЬТР В
МОДЕЛЬ ХИЛЛЕ
ВХОД В КАНАЛ – ПРЯМОУГОЛЬНАЯ ПОРА 0,3х0,5 нм
СЕЛЕКТИВНЫЙ ФИЛЬТР В
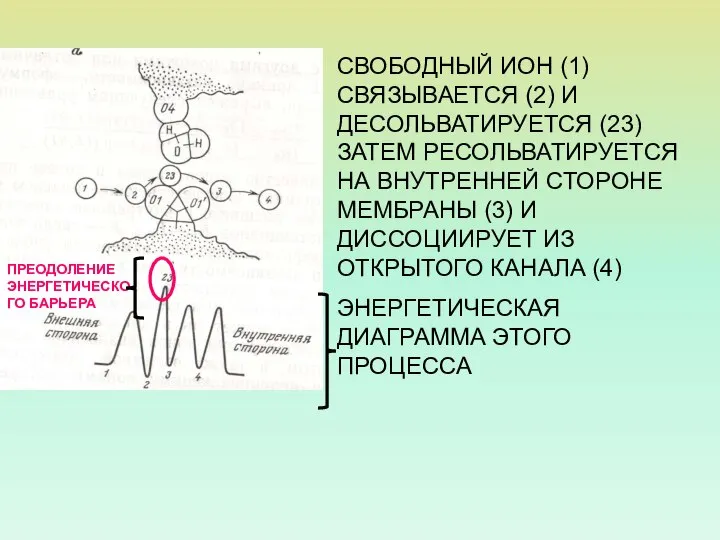
СВОБОДНЫЙ ИОН (1) СВЯЗЫВАЕТСЯ (2) И ДЕСОЛЬВАТИРУЕТСЯ (23) ЗАТЕМ РЕСОЛЬВАТИРУЕТСЯ НА
СВОБОДНЫЙ ИОН (1) СВЯЗЫВАЕТСЯ (2) И ДЕСОЛЬВАТИРУЕТСЯ (23) ЗАТЕМ РЕСОЛЬВАТИРУЕТСЯ НА
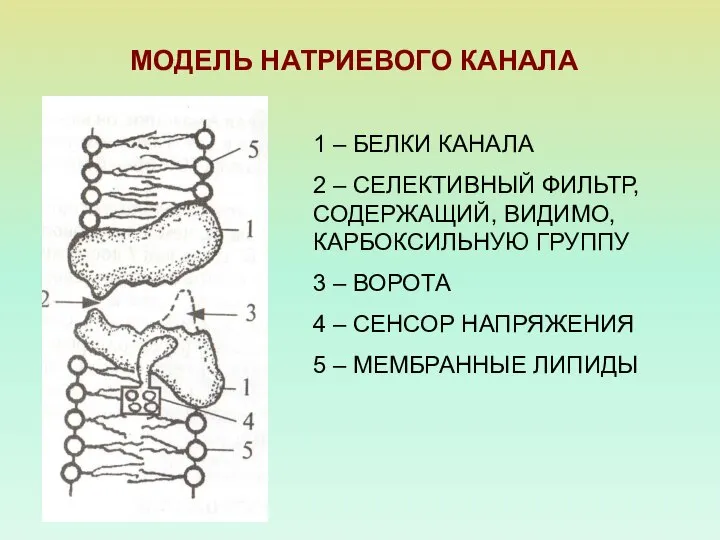
МОДЕЛЬ НАТРИЕВОГО КАНАЛА
1 – БЕЛКИ КАНАЛА
2 – СЕЛЕКТИВНЫЙ ФИЛЬТР, СОДЕРЖАЩИЙ, ВИДИМО,
МОДЕЛЬ НАТРИЕВОГО КАНАЛА
1 – БЕЛКИ КАНАЛА
2 – СЕЛЕКТИВНЫЙ ФИЛЬТР, СОДЕРЖАЩИЙ, ВИДИМО,
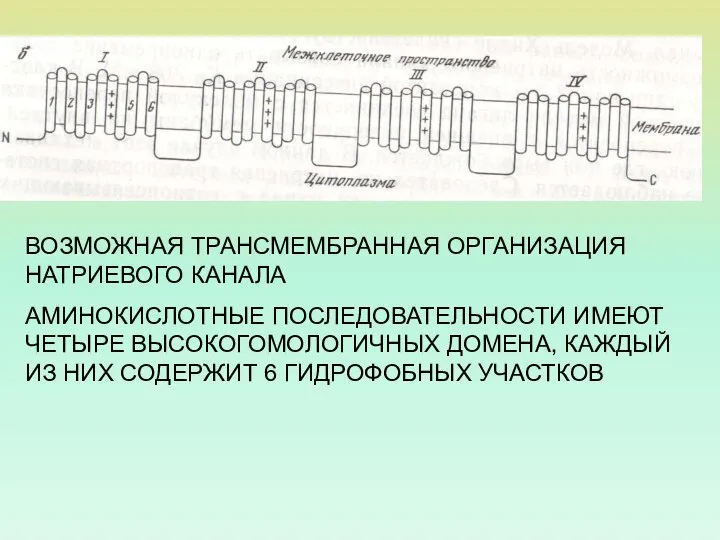
ВОЗМОЖНАЯ ТРАНСМЕМБРАННАЯ ОРГАНИЗАЦИЯ НАТРИЕВОГО КАНАЛА
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ИМЕЮТ ЧЕТЫРЕ ВЫСОКОГОМОЛОГИЧНЫХ ДОМЕНА, КАЖДЫЙ
ВОЗМОЖНАЯ ТРАНСМЕМБРАННАЯ ОРГАНИЗАЦИЯ НАТРИЕВОГО КАНАЛА
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ИМЕЮТ ЧЕТЫРЕ ВЫСОКОГОМОЛОГИЧНЫХ ДОМЕНА, КАЖДЫЙ
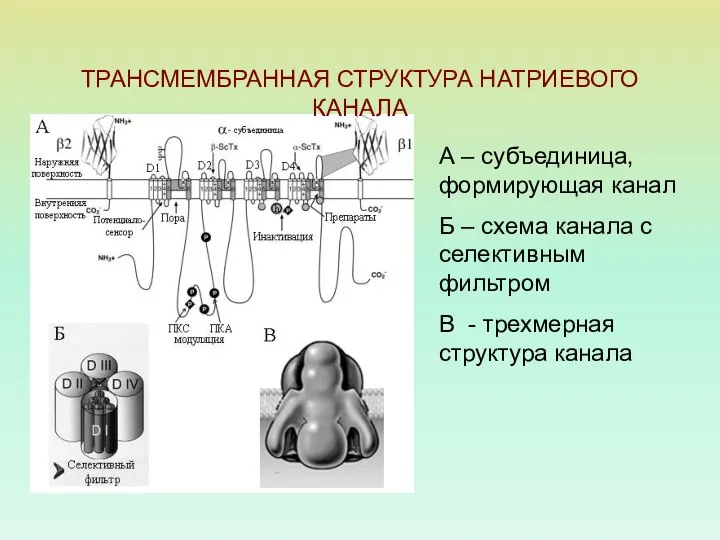
ТРАНСМЕМБРАННАЯ СТРУКТУРА НАТРИЕВОГО КАНАЛА
А – субъединица, формирующая канал
Б – схема канала
ТРАНСМЕМБРАННАЯ СТРУКТУРА НАТРИЕВОГО КАНАЛА
А – субъединица, формирующая канал
Б – схема канала

БЛОКАТОРЫ НАТРИЕВОГО КАНАЛА
БЛОКАТОРЫ НАТРИЕВОГО КАНАЛА

ТЕТРОДОТОКСИН
ТЕТРОДОТОКСИН
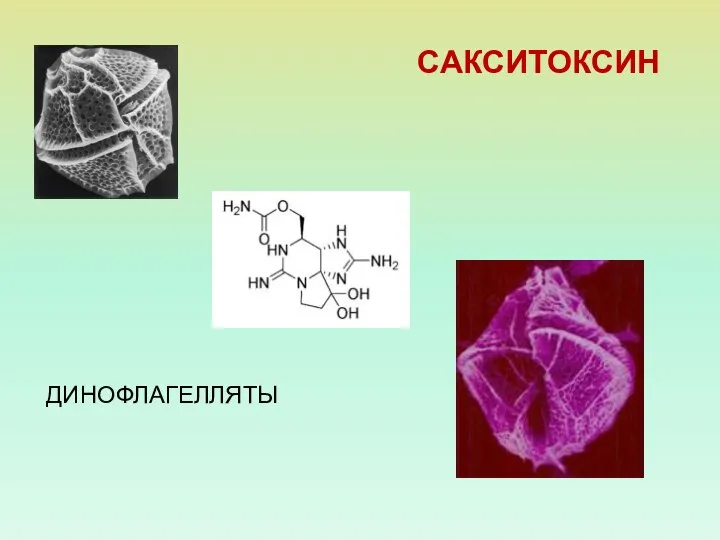
САКСИТОКСИН
ДИНОФЛАГЕЛЛЯТЫ
САКСИТОКСИН
ДИНОФЛАГЕЛЛЯТЫ
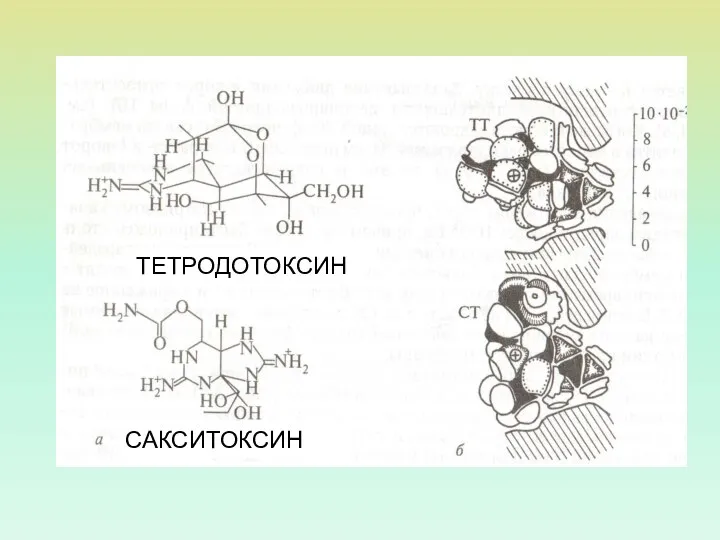
ТЕТРОДОТОКСИН
САКСИТОКСИН
ТЕТРОДОТОКСИН
САКСИТОКСИН
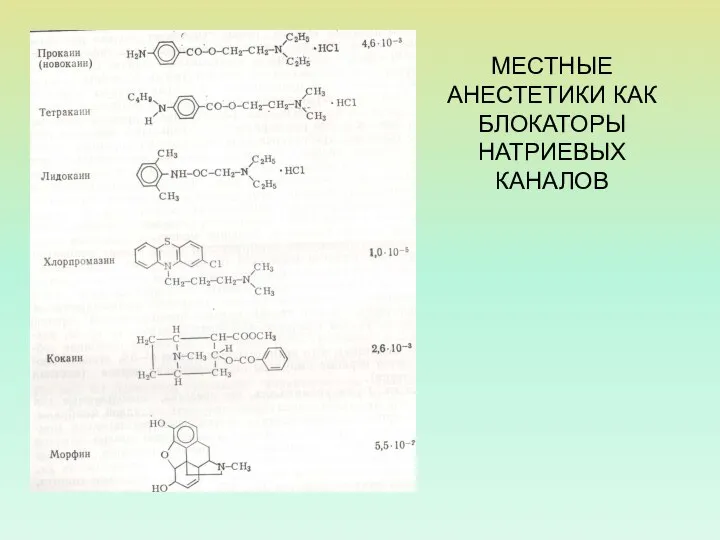
МЕСТНЫЕ АНЕСТЕТИКИ КАК БЛОКАТОРЫ НАТРИЕВЫХ КАНАЛОВ
МЕСТНЫЕ АНЕСТЕТИКИ КАК БЛОКАТОРЫ НАТРИЕВЫХ КАНАЛОВ
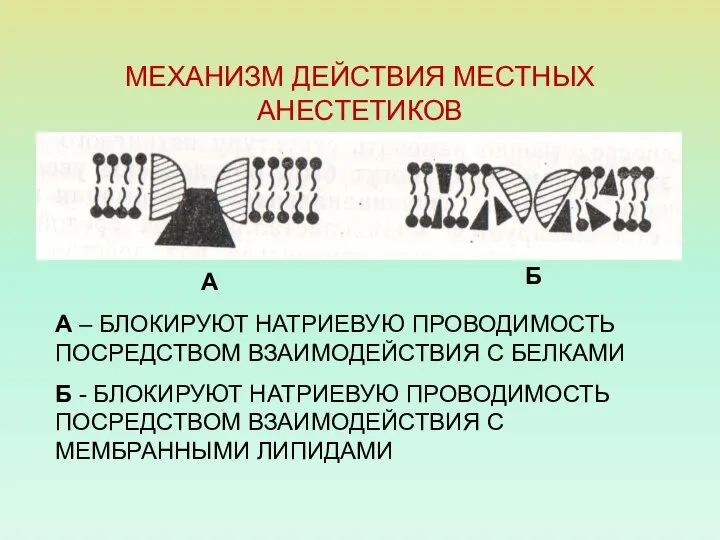
МЕХАНИЗМ ДЕЙСТВИЯ МЕСТНЫХ АНЕСТЕТИКОВ
А – БЛОКИРУЮТ НАТРИЕВУЮ ПРОВОДИМОСТЬ ПОСРЕДСТВОМ ВЗАИМОДЕЙСТВИЯ С
МЕХАНИЗМ ДЕЙСТВИЯ МЕСТНЫХ АНЕСТЕТИКОВ
А – БЛОКИРУЮТ НАТРИЕВУЮ ПРОВОДИМОСТЬ ПОСРЕДСТВОМ ВЗАИМОДЕЙСТВИЯ С
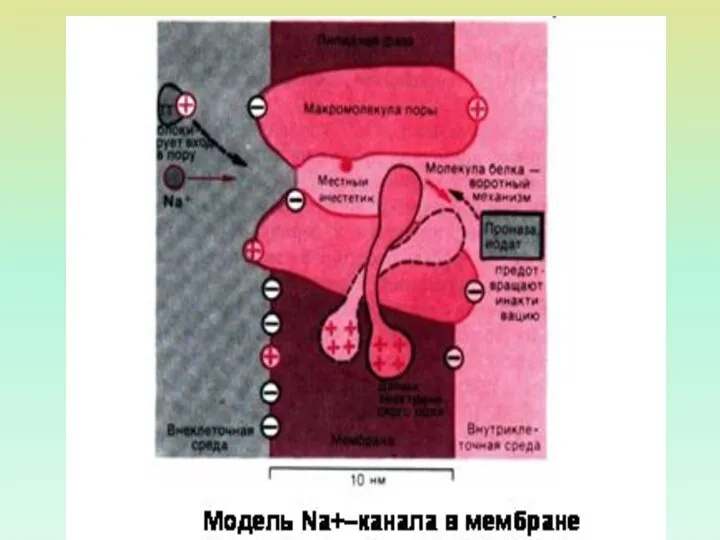

ВОЗДЕЙСТВИЯ, УСТРАНЯЮЩИЕ ИНАКТИВАЦИЮ НАТРИЕВОГО КАНАЛА
ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
ПЕПТИДНЫЕ ТОКСИНЫ (ЯД СКОРПИОНА,
ЯД АНЕМОН)
АЛКОЛОИДНЫЕ
ВОЗДЕЙСТВИЯ, УСТРАНЯЮЩИЕ ИНАКТИВАЦИЮ НАТРИЕВОГО КАНАЛА
ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
ПЕПТИДНЫЕ ТОКСИНЫ (ЯД СКОРПИОНА,
ЯД АНЕМОН)
АЛКОЛОИДНЫЕ
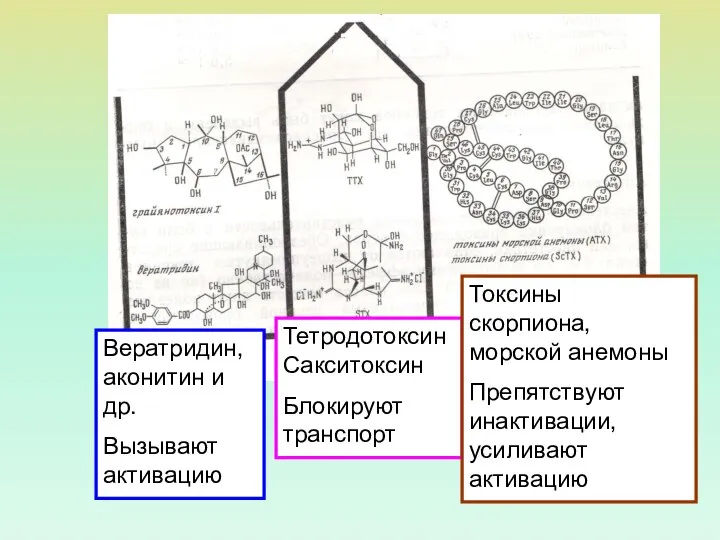
ТетродотоксинСакситоксин
Блокируют транспорт
Токсины скорпиона, морской анемоны
Препятствуют инактивации, усиливают активацию
Вератридин, аконитин и др.
Вызывают
ТетродотоксинСакситоксин
Блокируют транспорт
Токсины скорпиона, морской анемоны
Препятствуют инактивации, усиливают активацию
Вератридин, аконитин и др.
Вызывают

КАЛИЕВЫЙ КАНАЛ
КАЛИЕВЫЙ КАНАЛ
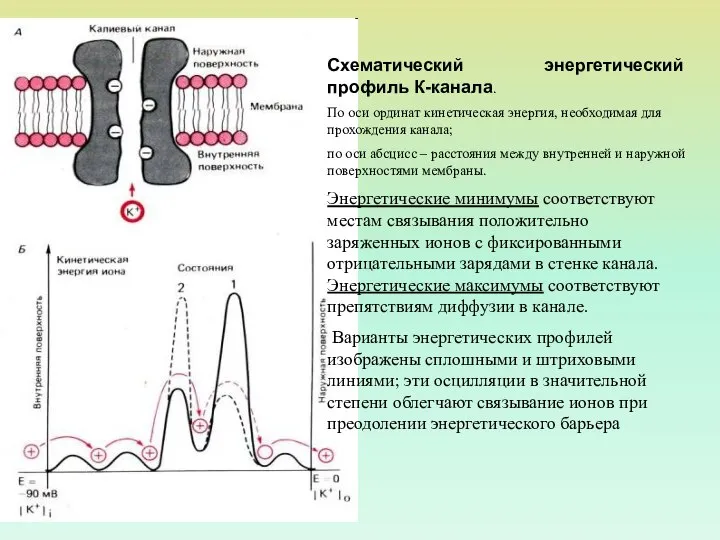
Схематический энергетический профиль К-канала.
По оси ординат кинетическая энергия, необходимая для
Схематический энергетический профиль К-канала.
По оси ординат кинетическая энергия, необходимая для

ПРОНИЦАЕМОСТЬ КАЛИЕВОГО КАНАЛА
ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ
НЕ КОРРЕЛИРУЕТ С РАДИУСОМ ИОНА
PK
ПРОНИЦАЕМОСТЬ КАЛИЕВОГО КАНАЛА
ДЛЯ ОДНОВАЛЕНТНЫХ КАТИОНОВ
НЕ КОРРЕЛИРУЕТ С РАДИУСОМ ИОНА
PK
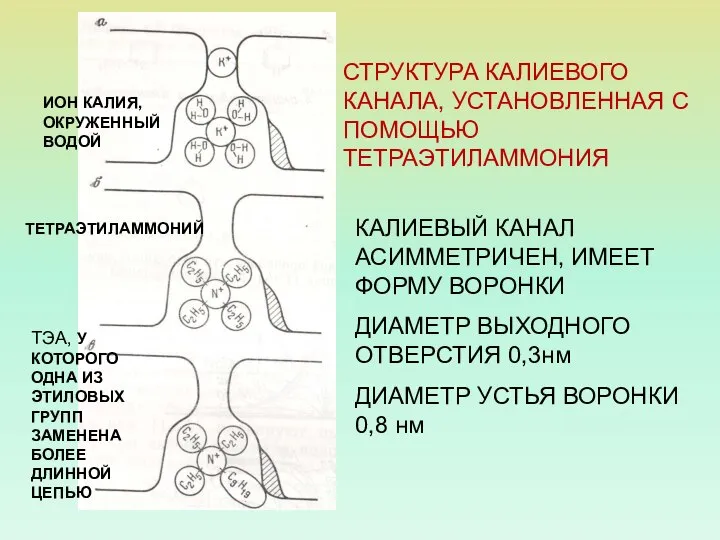
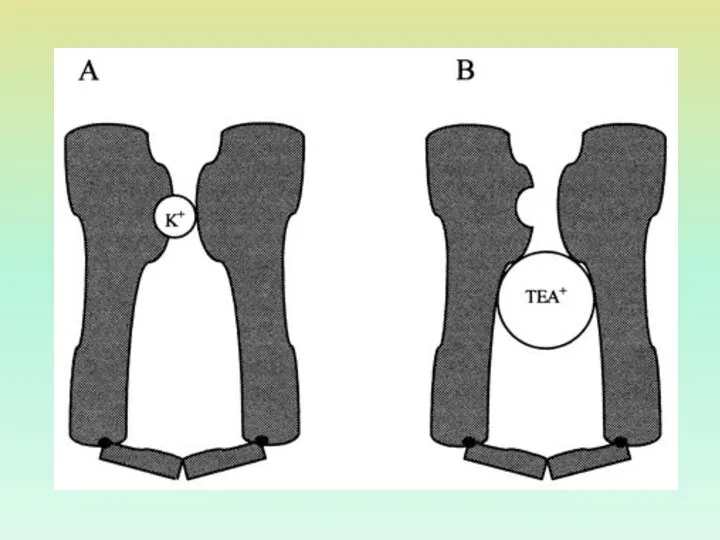
СТРУКТУРА КАЛИЕВОГО КАНАЛА, УСТАНОВЛЕННАЯ С ПОМОЩЬЮ ТЕТРАЭТИЛАММОНИЯ
КАЛИЕВЫЙ КАНАЛ АСИММЕТРИЧЕН, ИМЕЕТ ФОРМУ
СТРУКТУРА КАЛИЕВОГО КАНАЛА, УСТАНОВЛЕННАЯ С ПОМОЩЬЮ ТЕТРАЭТИЛАММОНИЯ
КАЛИЕВЫЙ КАНАЛ АСИММЕТРИЧЕН, ИМЕЕТ ФОРМУ
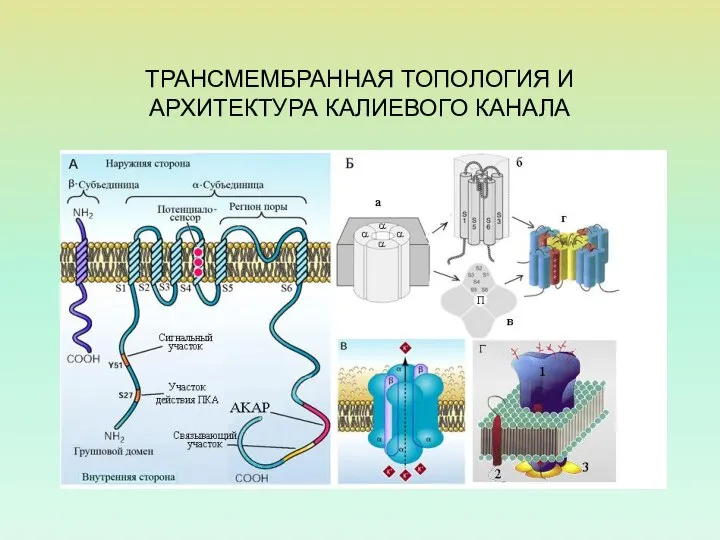
ТРАНСМЕМБРАННАЯ ТОПОЛОГИЯ И АРХИТЕКТУРА КАЛИЕВОГО КАНАЛА
ТРАНСМЕМБРАННАЯ ТОПОЛОГИЯ И АРХИТЕКТУРА КАЛИЕВОГО КАНАЛА

Р. Мак-Киннон
Нобелевская премия по химии за 2003 г. присуждена
Р. Мак-Киннону
Р. Мак-Киннон
Нобелевская премия по химии за 2003 г. присуждена Р. Мак-Киннону

Мак-Киннон и его коллеги расшифровали структуру и механизм функционирования нескольких бактериальных
Мак-Киннон и его коллеги расшифровали структуру и механизм функционирования нескольких бактериальных
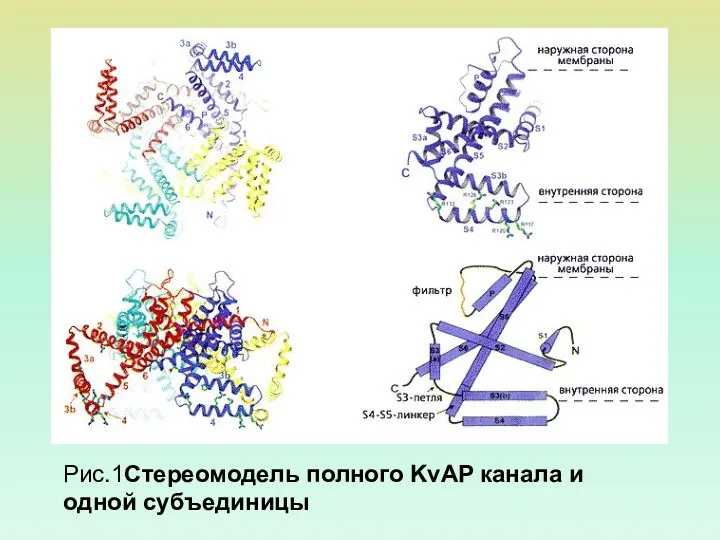
Рис.1Стереомодель полного KvAP канала и одной субъединицы
Рис.1Стереомодель полного KvAP канала и одной субъединицы

Рис. 3. Модели, иллюстрирующие перенос положительных зарядов канальным калиевым белком в
Рис. 3. Модели, иллюстрирующие перенос положительных зарядов канальным калиевым белком в

Рис. 4. Схема движения “лопастей” калиевого канала и их положение внутри
Рис. 4. Схема движения “лопастей” калиевого канала и их положение внутри

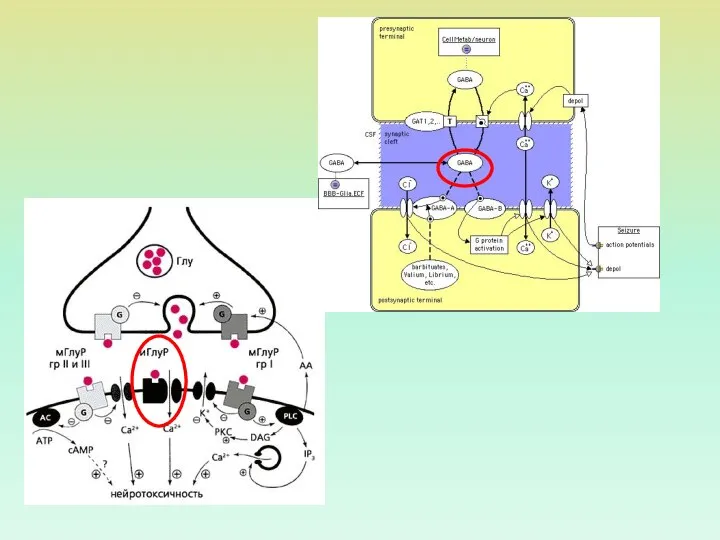
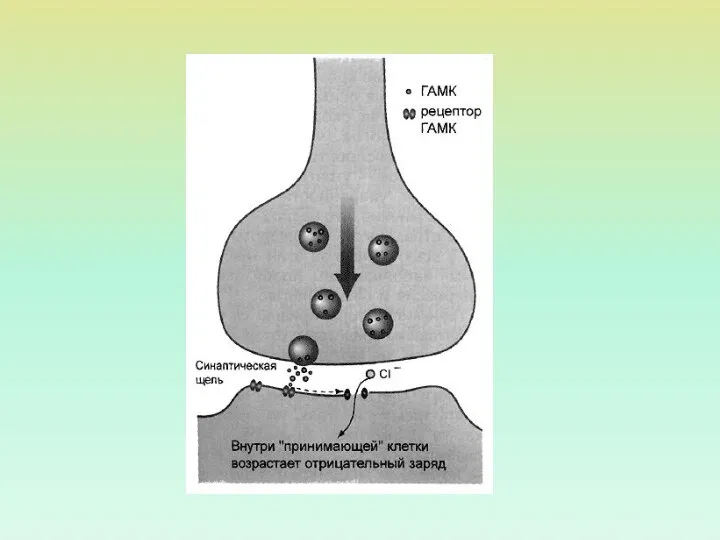
ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
МЕХАНОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
МЕХАНОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ

ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
ХЕМОЧУВСТВИТЕЛЬНЫЕ ИОННЫЕ КАНАЛЫ
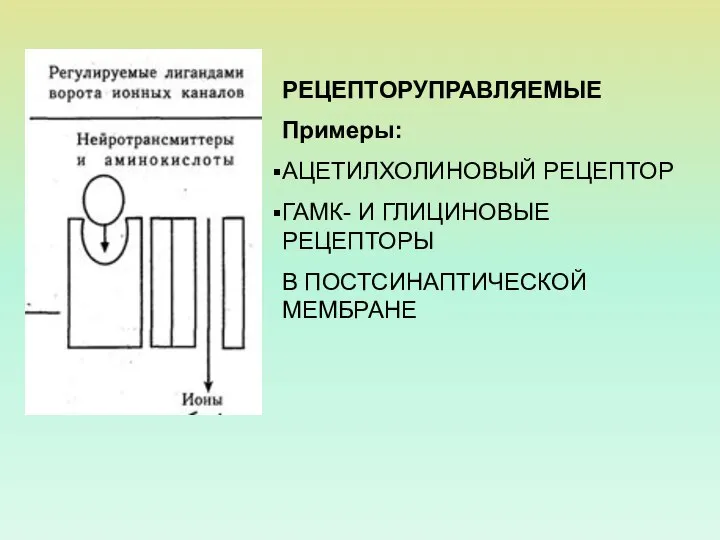
РЕЦЕПТОРУПРАВЛЯЕМЫЕ
Примеры:
АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР
ГАМК- И ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ
В ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ
РЕЦЕПТОРУПРАВЛЯЕМЫЕ
Примеры:
АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР
ГАМК- И ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ
В ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ
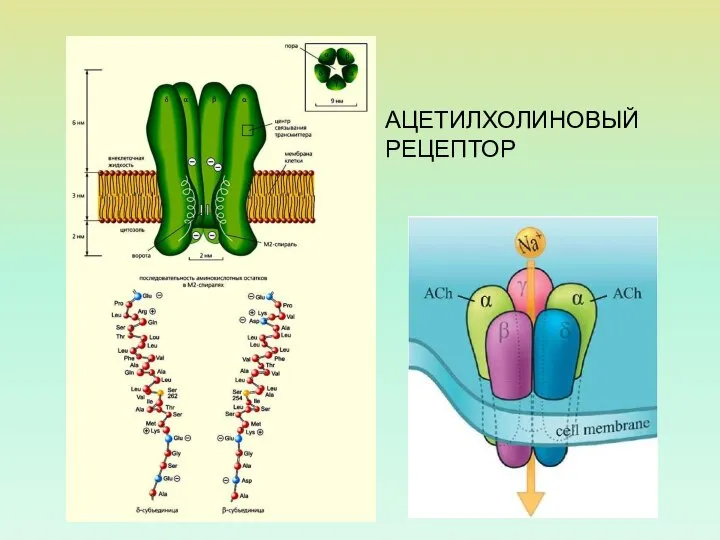
АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР
АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР
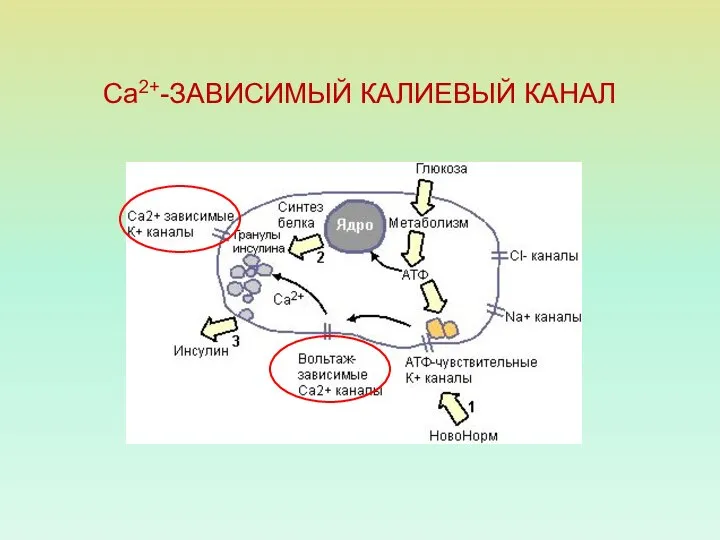
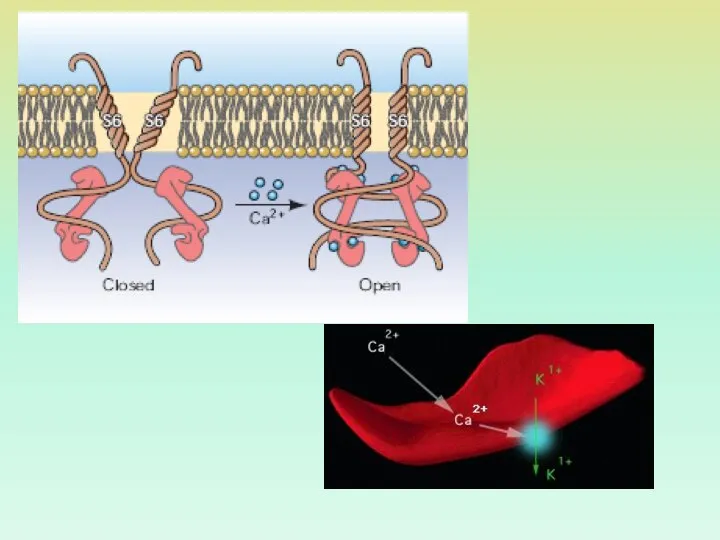
Са2+-ЗАВИСИМЫЙ КАЛИЕВЫЙ КАНАЛ
Са2+-ЗАВИСИМЫЙ КАЛИЕВЫЙ КАНАЛ
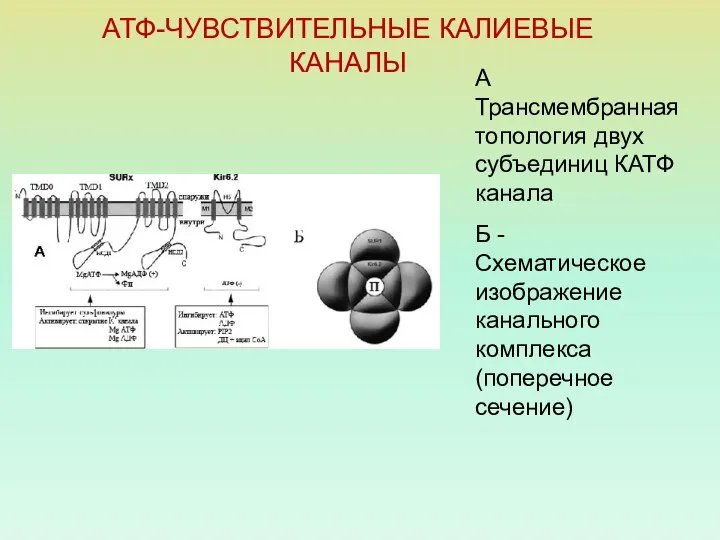
АТФ-ЧУВСТВИТЕЛЬНЫЕ КАЛИЕВЫЕ КАНАЛЫ
А Трансмембранная топология двух субъединиц КАТФ канала
Б - Схематическое
АТФ-ЧУВСТВИТЕЛЬНЫЕ КАЛИЕВЫЕ КАНАЛЫ
А Трансмембранная топология двух субъединиц КАТФ канала
Б - Схематическое
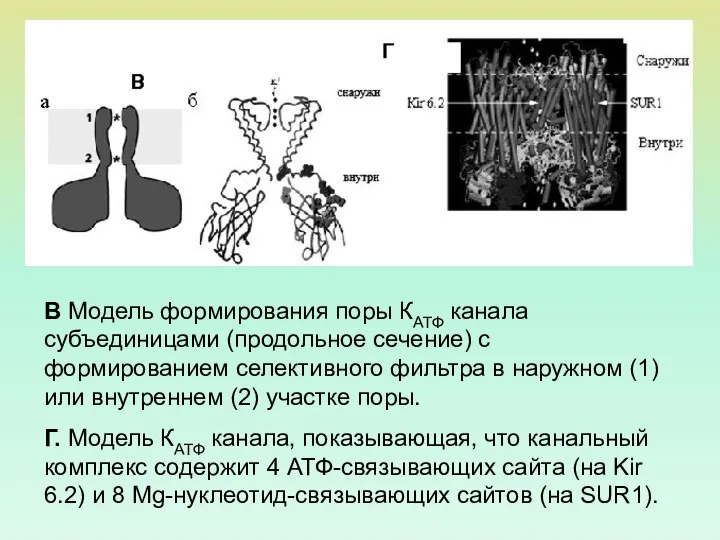
В Модель формирования поры КАТФ канала субъединицами (продольное сечение) с формированием
В Модель формирования поры КАТФ канала субъединицами (продольное сечение) с формированием
СЕКРЕЦИЯ ИНСУЛИНА: РОЛЬ ИОННЫХ КАНАЛОВ
1 — К+-АТФ чувствительный канал;
2 —
СЕКРЕЦИЯ ИНСУЛИНА: РОЛЬ ИОННЫХ КАНАЛОВ
1 — К+-АТФ чувствительный канал;
2 —

МЕХАНОЧУВСТВИТЕЛЬНЫЙ КАНАЛ
МЕХАНОЧУВСТВИТЕЛЬНЫЙ КАНАЛ
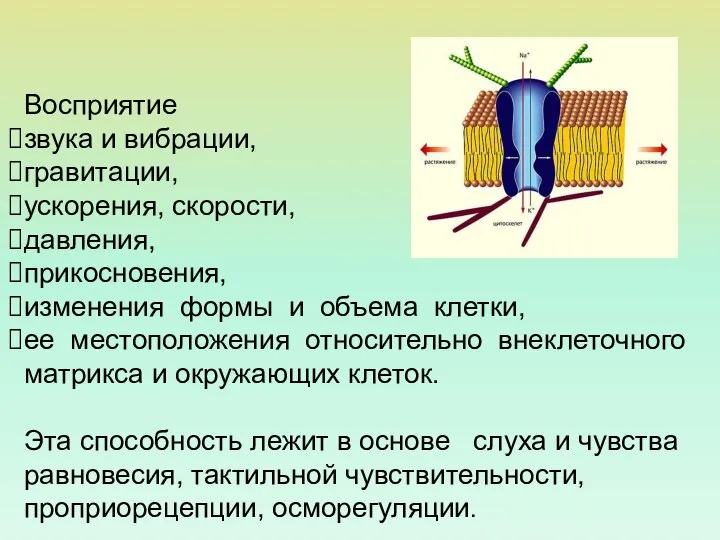
Восприятие
звука и вибрации,
гравитации,
ускорения, скорости,
давления,
прикосновения,
изменения формы
Восприятие
звука и вибрации,
гравитации,
ускорения, скорости,
давления,
прикосновения,
изменения формы

Системы управления механочувствительным каналом
Системы управления механочувствительным каналом
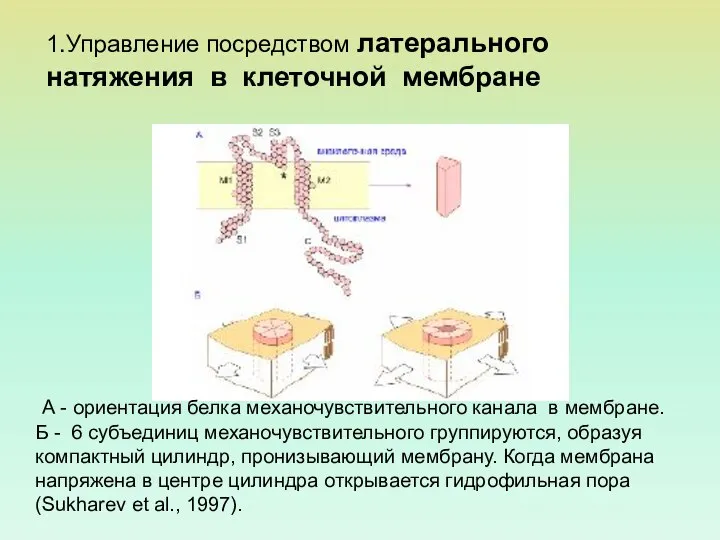
А - ориентация белка механочувствительного канала в мембране.
Б -
А - ориентация белка механочувствительного канала в мембране.
Б -
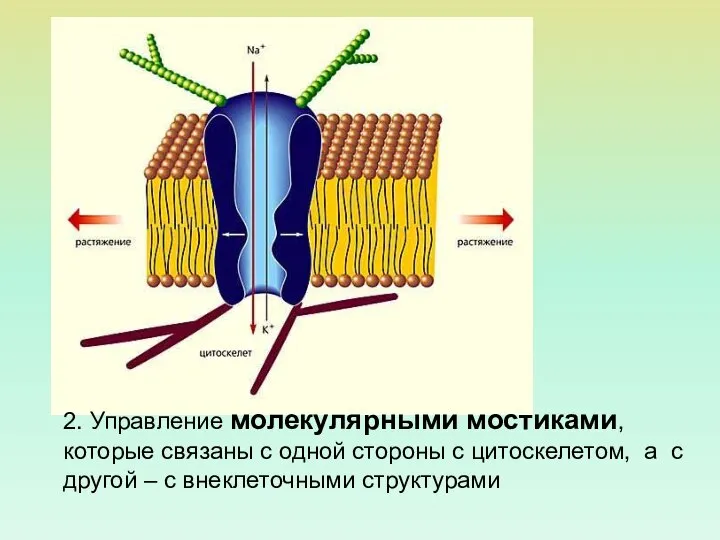
2. Управление молекулярными мостиками, которые связаны с одной стороны с цитоскелетом,
2. Управление молекулярными мостиками, которые связаны с одной стороны с цитоскелетом,
 Простые, но важные способы продвижения бизнеса в интернете
Простые, но важные способы продвижения бизнеса в интернете Презентация по образовательной программе внеурочной деятельности «Планета Здоровья» по курсу «Земля- наш дом» Составитель: учи
Презентация по образовательной программе внеурочной деятельности «Планета Здоровья» по курсу «Земля- наш дом» Составитель: учи Система управления качеством на предприятии и пути ее совершенствования с целью повышения конкурентоспособности
Система управления качеством на предприятии и пути ее совершенствования с целью повышения конкурентоспособности Крепость Нарын-Кала в истории Дагестана
Крепость Нарын-Кала в истории Дагестана マーケティング講座資料
マーケティング講座資料 Центрифуга
Центрифуга Презентация на тему "13 признаков учителя" - скачать презентации по Педагогике
Презентация на тему "13 признаков учителя" - скачать презентации по Педагогике Портрет. Изучение нового материала
Портрет. Изучение нового материала Характеристики систем видеоэлектроники. Импульсная реакция или функция рассеяния точки
Характеристики систем видеоэлектроники. Импульсная реакция или функция рассеяния точки Жозеф Луи Гей-Люссак. Французский физик и химик (1778 - 1850)
Жозеф Луи Гей-Люссак. Французский физик и химик (1778 - 1850) Арбитраж. 9 поток
Арбитраж. 9 поток Показания свидетелей
Показания свидетелей Ресторан мечты. Начало. Организационные моменты
Ресторан мечты. Начало. Организационные моменты Презентация Социальное страхование в России
Презентация Социальное страхование в России Классы радиоуправляемых автомоделей
Классы радиоуправляемых автомоделей Презентация Костяной фарфор
Презентация Костяной фарфор Понятие, виды и структура норм права
Понятие, виды и структура норм права Порядок формирования палат Федерального Собрания
Порядок формирования палат Федерального Собрания Етика та естетика. Виникнення етики та її розвиток
Етика та естетика. Виникнення етики та її розвиток Қазақстан Республикасы Президенті Н.Назарбаевтың Қазақстан халқына «Нұрлы жол - болашаққа бастар жол». Жолдауы
Қазақстан Республикасы Президенті Н.Назарбаевтың Қазақстан халқына «Нұрлы жол - болашаққа бастар жол». Жолдауы Искусство Гжели. Истоки и современное развитие промысла. Урок- презентация в 5 классе по ИЗО программа Б.М.Неменского Автор : Баз
Искусство Гжели. Истоки и современное развитие промысла. Урок- презентация в 5 классе по ИЗО программа Б.М.Неменского Автор : Баз Служебное расследование
Служебное расследование  Общие сведения о компьютерах
Общие сведения о компьютерах Отчет Богу
Отчет Богу Разработка модели государственно-общественного управления
Разработка модели государственно-общественного управления Трудовое право
Трудовое право Prefer class hierarchies to tagged classes. (Item 20, 21, 22)
Prefer class hierarchies to tagged classes. (Item 20, 21, 22) Решение задач в младших класах - презентация для начальной школы
Решение задач в младших класах - презентация для начальной школы