- макроэргические соединения

Содержание
- 2. Владимир Петрович Скулачев Академик РАН, директор Научно-исследовательского института физико-химической биологии им. А.Н.Белозерского и декан факультета биоинженерии
- 3. 1-й закон Живая клетка избегает прямого использования энергии внешних ресурсов для совершения полезной работы. Она сначала
- 4. 2-й закон Любая живая клетка всегда располагает как минимум двумя "энергетическими валютами": водорастворимой (АТФ ) и
- 5. Таким образом, Биологические виды энергии зависит от места локализации. Энергетические превращения в живой клетке подразделяют на
- 6. Макроэргические соединения
- 7. Макроэргические соединения (macroergic compounds) [греч. makros — большой и ergon — работа, действие] — органические соединения,
- 8. Термин «макроэргическая связь» используется исключительно для связей, энергия которых используется в метаболизме и не указывает на
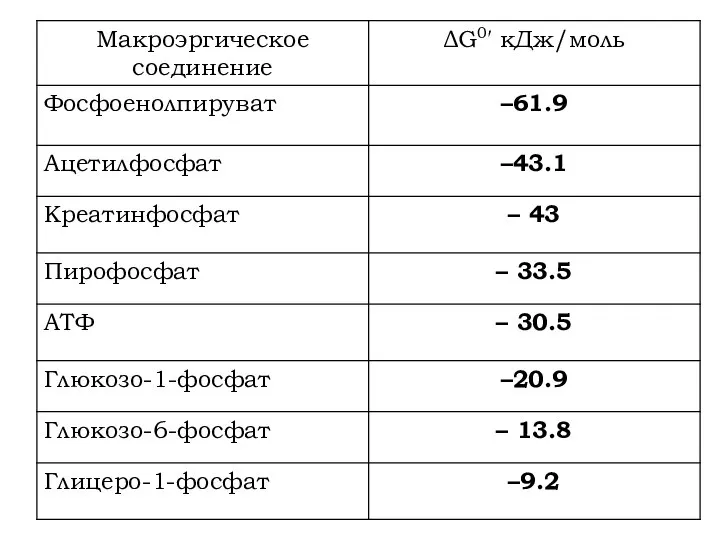
- 9. Виды макроэргических связей Енолфосфатная связь ΔG0′ = –61.9 кДж/моль Представители: фосфоенолпируват
- 11. Ацилфосфатная связь ΔG0′ = – 46 кДж/моль. Представители: 1,3-дифосфоглицерат
- 12. Гуанидинфосфатная связь ΔG0′ = – 42 кДж/моль. Представители: креатинфосфат
- 14. Тиоэфирная связь ΔG0′ = – 34 кДж/моль. Представители: ацетил-КоА, сукцинил-КоА.
- 15. Фосфоангидридная связь ΔG0′ = – 32 кДж/моль Представители: все нуклеозидтрифосфаты и нуклеозиддифосфаты (АТФ, ГДФ и их
- 17. Аденозинтрифосфорная кислота АТФ был открыт в 1929 году Карлом Ломанном. В 1940-41 гг. немецким биохимиком Фрицем
- 19. В клетках организмов животных есть три основных источника ~P для синтеза АТФ. 1) окислительное фосфорилирование –
- 20. Сумму всех адениловых нуклеотидов в клетке (АТФ, АДФ и АМФ) называют адениловой системой. Процессы гидролиза и
- 21. Причины макроэргичности АТФ: 1 причина. АТФ присутствует в клетках в диссоциированной форме: АТФ4- + H2O ----->
- 22. 2 причина: в структуре АТФ имеется 3 фосфата и 2 ангидридные связи, за счет этого на
- 24. Скачать презентацию

Владимир Петрович Скулачев
Академик РАН, директор Научно-исследовательского института физико-химической биологии им. А.Н.Белозерского
Владимир Петрович Скулачев
Академик РАН, директор Научно-исследовательского института физико-химической биологии им. А.Н.Белозерского

1-й закон
Живая клетка избегает прямого использования энергии внешних ресурсов для совершения
1-й закон
Живая клетка избегает прямого использования энергии внешних ресурсов для совершения

2-й закон
Любая живая клетка всегда располагает как минимум двумя "энергетическими валютами": водорастворимой (АТФ
2-й закон
Любая живая клетка всегда располагает как минимум двумя "энергетическими валютами": водорастворимой (АТФ

Таким образом,
Биологические виды энергии зависит от места локализации.
Энергетические превращения в
Таким образом,
Биологические виды энергии зависит от места локализации.
Энергетические превращения в

Макроэргические соединения
Макроэргические соединения

Макроэргические соединения (macroergic compounds) [греч. makros — большой и ergon —
Макроэргические соединения (macroergic compounds) [греч. makros — большой и ergon —

Термин «макроэргическая связь» используется исключительно для связей, энергия которых используется в
Термин «макроэргическая связь» используется исключительно для связей, энергия которых используется в
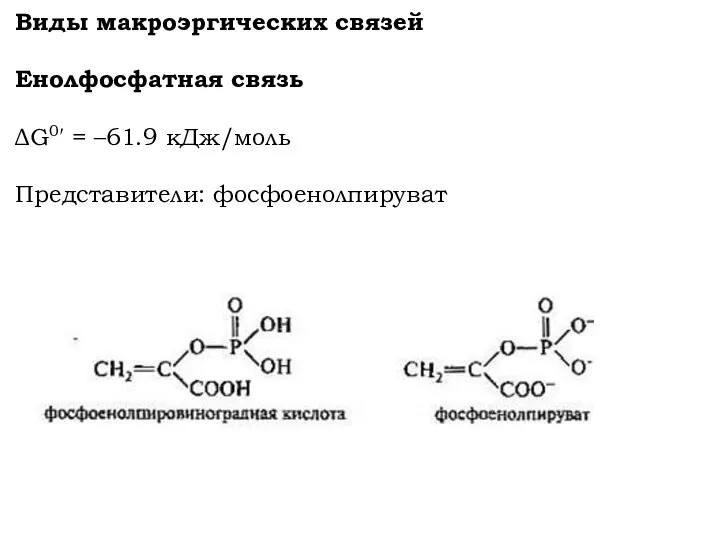
Виды макроэргических связей
Енолфосфатная связь
ΔG0′ = –61.9 кДж/моль
Представители: фосфоенолпируват
Виды макроэргических связей
Енолфосфатная связь
ΔG0′ = –61.9 кДж/моль
Представители: фосфоенолпируват

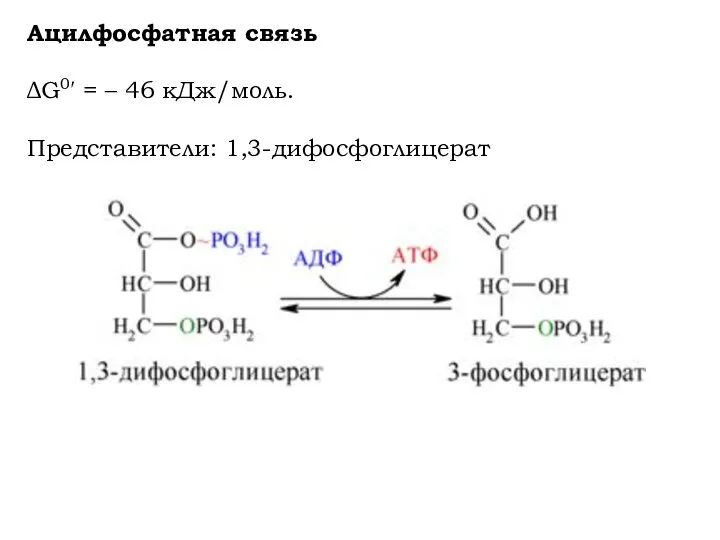
Ацилфосфатная связь
ΔG0′ = – 46 кДж/моль.
Представители: 1,3-дифосфоглицерат
Ацилфосфатная связь
ΔG0′ = – 46 кДж/моль.
Представители: 1,3-дифосфоглицерат
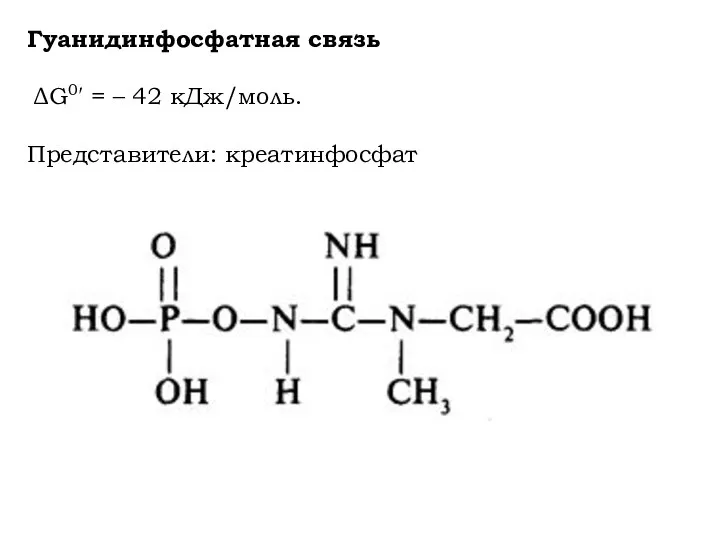
Гуанидинфосфатная связь
ΔG0′ = – 42 кДж/моль.
Представители: креатинфосфат
Гуанидинфосфатная связь
ΔG0′ = – 42 кДж/моль.
Представители: креатинфосфат
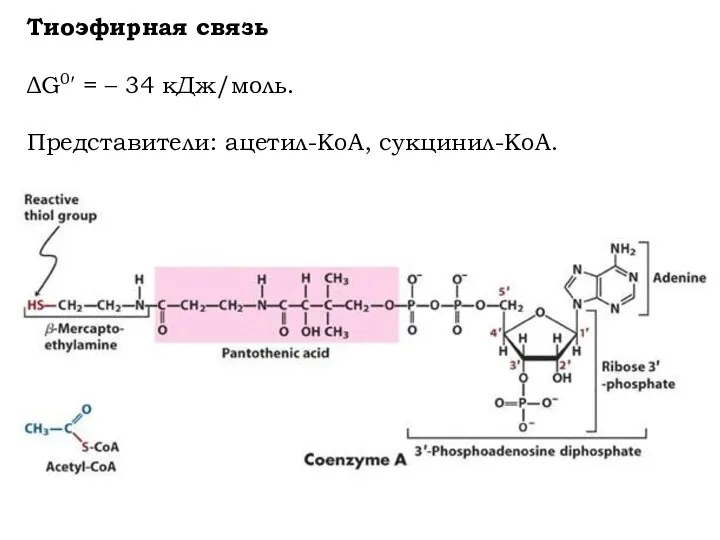
Тиоэфирная связь
ΔG0′ = – 34 кДж/моль.
Представители: ацетил-КоА, сукцинил-КоА.
Тиоэфирная связь
ΔG0′ = – 34 кДж/моль.
Представители: ацетил-КоА, сукцинил-КоА.
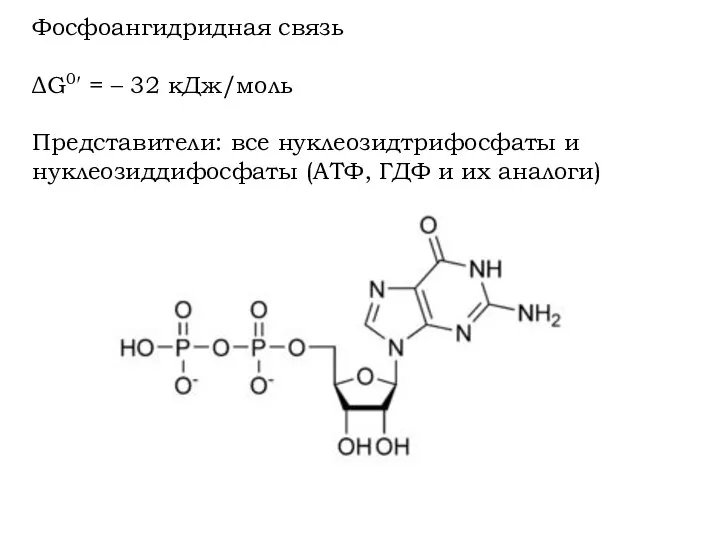
Фосфоангидридная связь
ΔG0′ = – 32 кДж/моль
Представители: все нуклеозидтрифосфаты и нуклеозиддифосфаты (АТФ,
Фосфоангидридная связь
ΔG0′ = – 32 кДж/моль
Представители: все нуклеозидтрифосфаты и нуклеозиддифосфаты (АТФ,

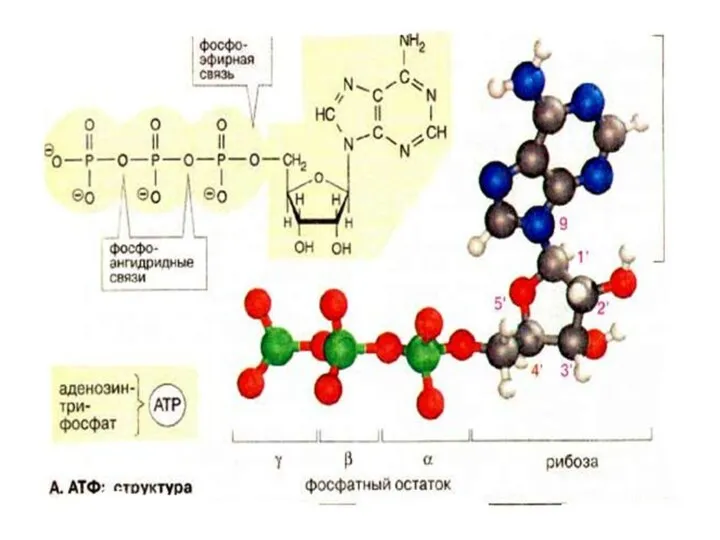
Аденозинтрифосфорная кислота
АТФ был открыт в 1929 году Карлом Ломанном.
В 1940-41 гг.
Аденозинтрифосфорная кислота
АТФ был открыт в 1929 году Карлом Ломанном.
В 1940-41 гг.

В клетках организмов животных есть три основных источника ~P для синтеза
В клетках организмов животных есть три основных источника ~P для синтеза
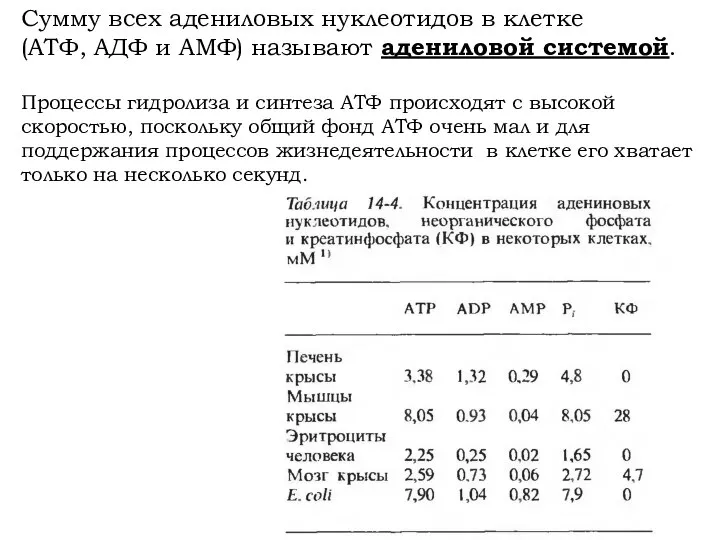
Сумму всех адениловых нуклеотидов в клетке
(АТФ, АДФ и АМФ) называют адениловой
Сумму всех адениловых нуклеотидов в клетке
(АТФ, АДФ и АМФ) называют адениловой

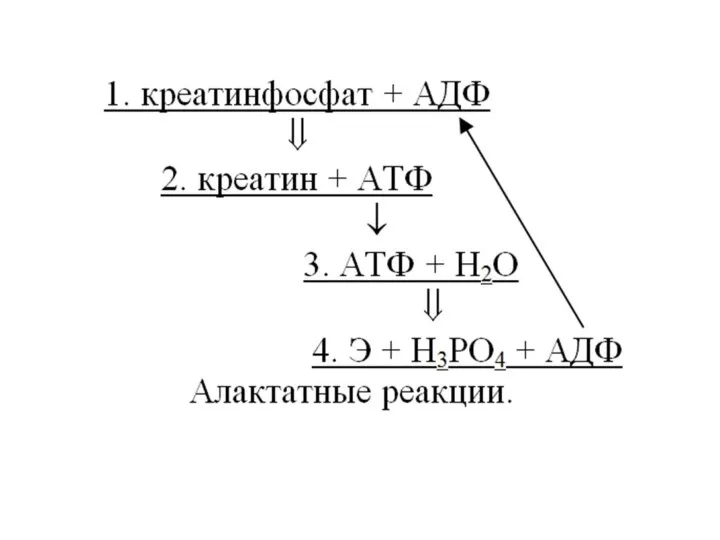
Причины макроэргичности АТФ:
1 причина.
АТФ присутствует в клетках в диссоциированной форме:
АТФ4-
Причины макроэргичности АТФ:
1 причина.
АТФ присутствует в клетках в диссоциированной форме:
АТФ4-

2 причина:
в структуре АТФ имеется 3 фосфата и 2 ангидридные
2 причина:
в структуре АТФ имеется 3 фосфата и 2 ангидридные
 Соломенный бычок-смоляной бочок
Соломенный бычок-смоляной бочок  Баға экономикалық категориясы: мағынасы, жүйелері, жіктелуі. Дәрілік заттардың бағасының түрлері
Баға экономикалық категориясы: мағынасы, жүйелері, жіктелуі. Дәрілік заттардың бағасының түрлері Великие математики - презентация по Алгебре_
Великие математики - презентация по Алгебре_ Бизнес и общество Лекций 32 Практических Занятий 16 экзамен
Бизнес и общество Лекций 32 Практических Занятий 16 экзамен  Охрана западной границы России регулярными войсками Выполнила студентка 2-го курса группы Э101 Овсянникова Надежда
Охрана западной границы России регулярными войсками Выполнила студентка 2-го курса группы Э101 Овсянникова Надежда Nike компаниясы
Nike компаниясы Europol
Europol Актуальность хохломской росписи в современном мире
Актуальность хохломской росписи в современном мире Умножение многозначного на однозначное число
Умножение многозначного на однозначное число  Модернизация на стопанския живот и българското общество до средата на XX век Изготвено от: Катерина К. Кирова
Модернизация на стопанския живот и българското общество до средата на XX век Изготвено от: Катерина К. Кирова ROUTING-5
ROUTING-5  Роль Интернет-ресурсов в продвижении имиджа региона
Роль Интернет-ресурсов в продвижении имиджа региона Собственные имена существительные (Обобщающий урок)
Собственные имена существительные (Обобщающий урок) Генплан детской площадки
Генплан детской площадки Аттестационная работа: Диалог культур во имя гражданского мира и согласия
Аттестационная работа: Диалог культур во имя гражданского мира и согласия Резьба. Изображение резьбы
Резьба. Изображение резьбы Шығыс мәдениетіндегі философия феномені
Шығыс мәдениетіндегі философия феномені Презентация на тему "Остеохондроз шейного отдела позвоночника" - скачать презентации по Медицине
Презентация на тему "Остеохондроз шейного отдела позвоночника" - скачать презентации по Медицине Соотношение, общие характеристики и принципиальные отличия тарифных и нетарифных мер регулирования внешнеторговой деятельности
Соотношение, общие характеристики и принципиальные отличия тарифных и нетарифных мер регулирования внешнеторговой деятельности  Социология политики
Социология политики Шаблони функцій. Шаблони класів
Шаблони функцій. Шаблони класів Складні випадки правопису знака м’якшення
Складні випадки правопису знака м’якшення Школа фольклорного искусства «Моя Русь». Русский народный костюм
Школа фольклорного искусства «Моя Русь». Русский народный костюм Как правильно составить резюме
Как правильно составить резюме Белорусские народные ремесла
Белорусские народные ремесла В мире басен
В мире басен Актуальные проблемы развития высшего образования и задачи психолого-педагогической подготовки преподавателя вуза
Актуальные проблемы развития высшего образования и задачи психолого-педагогической подготовки преподавателя вуза  * * * * * * * * * * * * *1. Якщо трапеція має прямий кут, її називають …. 2. Середньою лінією трапеції називається …. 3.На малюнку 1 трапеція ABCD - …, - презентация
* * * * * * * * * * * * *1. Якщо трапеція має прямий кут, її називають …. 2. Середньою лінією трапеції називається …. 3.На малюнку 1 трапеція ABCD - …, - презентация