- Обмен углеводов

Содержание
- 2. План лекции 5: Гликоген. Особенности строения Синтез и распад гликогена Глюконеогенез Пентозофосфатный путь Дополнение к лекции
- 3. Метаболизм глюкозы
- 4. Использование метаболитов ЦТК в синтезе различных соединений
- 5. Ключевая роль глюкозо – 6-фосфата
- 6. Карта основных путей метаболизма
- 7. Обмен углеводов
- 8. 1. Обмен гликогена Гликогенез и гликогенолиз
- 9. Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид). У растений эту же функцию
- 10. В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение при распаде гликогена большого количества
- 11. Линейные участки молекулы гликогена образованы остатками глюкозы, связанными альфа-1,4-гликозидными связями, а в местах разветвления – альфа
- 12. Синтез и распад гликогена Синтез и распад гликогена гормонозависимый процессы и регулируются инсулином и глюкагоном (и
- 13. Синтез гликогена - гликогенез Синтез гликогена ускоряется в период пищеварения (абсорбтивный период – 1-2 часа после
- 14. Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент ветвления переносит фрагмент из 5-6 остатков
- 15. Синтез гликогена - гликогенез
- 16. Образование УДФ-глюкозы
- 17. Далее подробно расписаны реакции гликогенеза:
- 18. 1. Получение активной формы глюкозы реакцией фосфорилирования:
- 19. 2. Изомеризация активной формы глюкозы:
- 20. 3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
- 21. Строение УДФ-глюкозы Нуклеозид-фосфосахара используются как переносчики звеньев моноз в реакциях синтеза полимерных молекул
- 22. 4. Перенос остатка глюкозы с УДФ-глюкозы на затравочную цепь гликогена, которая содержит не менее 4-х звеньев
- 23. Ветвление гликогена 5. Синтез α-(1,6)-гликозидных связей происходит при участии гликоген-ветвящегося фермента – гликоген-трансферазы, которая переносит концевой
- 24. Подводим итог: Гликоген синтезируется в период пищеварения (в течение 1-2 часов после приема углеводной пищи). Гликогенез
- 25. Для подготовки и включения глюкозы в растущую полисахаридную цепь требуется энергия 1 моль АТФ и 1
- 26. Необходимость превращения глюкозы в гликоген связана с тем, что накопление значительного количества глюкозы в клетке привело
- 27. Распад гликогена - гликогенолиз или мобилизация гликогена – происходит в основном в период между приёмами пищи
- 28. Гликогенолиз - распад гликогена
- 29. 1. Освобождение глюкозы в виде глюкозо-1-фосфата (реакция 5) происходит в результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет
- 30. Рис. Схема гликогенолиза
- 31. Мобилизация гликогена в печени и мыщцах отличаются В печени есть фермент глюкозо-6-фосфорилаза, в мышцах нет. Этот
- 32. Постоянный уровень глюкозы в крови в постабсорбтивный период = 3,5-5,5 мМ/л Органы мозг и эритроциты потребляют
- 33. Функция мышечного гликогена заключается в высвобождении глюкозо-6фосфата, используемого в самой мышце для окисления и получения энергии.
- 34. Рис.Особенности мобилизации гликогена в печени и мышцах
- 35. Гормоны регулирующие обмен гликогена Переключение процессов синтеза и мобилизации гликогена в печени происходит при переходе состояния
- 36. Рис. Изменение активности гликогенсинтетазы и гликогенфосфорилазы
- 37. Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в Крови. Инсулин и глюкагон
- 38. В период пищеварения преобладающим является влияние инсулина, и инсулин/глюкагоновый индекс в этом случае повышается . Инсулин
- 39. При интенсивной мышечной работе и стрессе в кровь из мозгового вещества надпочечников секретируется гормон адреналин. Он
- 40. Болезни углеводного обмена Причины: нарушения переваривания и всасывания углеводов, синтеза и распада гликогена. - ферментопатии, -
- 41. Нарушения переваривания дисахаридов
- 42. Гликогеновые болезни их причины
- 43. 2. ГЛЮКОНЕОГЕНЕЗ
- 44. Карта основных путей метаболизма
- 45. Обмен углеводов
- 46. Глюконеогенез Глюконеогенез – процесс синтеза глюкозы из веществ неуглеводной природы. Субстраты глюконеогенеза: пируват, лактат, глицерол, аминокислоты.
- 47. Процесс протекает главным образом в печени и менее интенсивно в корковом веществе почек и слизистой кишечника.
- 48. Большинство реакций гликолиза и гликонеогенеза являются обратимыми и катализируются одними и теми же ферментами. Четыре реакции
- 49. Схема Гликолиза (слева) и глюконеогенеза (справа) I, II, III – необратимые циклы, катализируются разными ферментами.
- 50. В митохондриях под действием ферментов малатдегидроеназы и аминотрансферазы образуется малат и аспартат из оксалоацетата (ЩУК), которые
- 51. Превращение пирувата (ПВК) в оксалоацетат 1 - транспорт ПВК из цитозоля в митохондрю; 2 – превращение
- 52. Рис.Суммарное уравнение образования глюкозы из пирувата Видно, что синтез глюкозы энергозатратный процесс
- 53. Использование лактата в качестве субстрата в глюконеогенезе связано с транспортом его в печень и превращением в
- 54. Цикл Кори 1 – поступление лактата из сокращающейся мышцы и эритроцитов с током крови в печень;
- 55. Часть ПВК, образовавшегося из лактата, окисляется в печени до СО2 и Н2О. Энергия выделяющаяся при окислении,
- 56. Снижение использования лактата в качестве субстрата в синтезе глюкозы, вызванное дефектом ферментов глюконеогенеза, и может приводить
- 57. Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие: тканевой гипоксии, вызывающей активацию анаэробного
- 58. 3. Пентозофосфатный путь в метаболизме глюкозы (ПФП) Это альтернативный путь окиления глюкозы. К синтезу АТФ этот
- 59. Кроме того, пентозофосфатный путь поставляет клетке пентозофосфаты (пентозы) , необходимые для синтеза нуклеиновых кислот и коферментов
- 60. В пентозофосфатном пути превращения глюкозы можно выделить две части: А – окислительный этап и Б –
- 61. Пентозофосфатный путь превращений глюкозы
- 62. ПФП превращения глюкозы А – окислительный этап; Б – неокислительный этап; В – пентозофосфатний цикл
- 63. Пентозофосфатный путь является альтернативным путем окисления глюкозы. Он включает несколько циклов, в результате функционирования которых из
- 64. 3глюкозо-6-фосфат + 6 НАДФ+ → → 3СО2 + 2глюкозо-6-фосфат + глицеральдегид-3-фосфат + 6 НАДФ-Н2 Пентозофосфатный цикл
- 65. ПФП в эритроцитах ПФП в эритроцитах очень важен, так как восстановленный НАДФН используется для защиты клеток
- 66. Дефект первого фермента ПФП глюкозо-6-фосфат дегидрогеназы приводит к дефициту НАДФН и невозможности восстановления глутатиона. Это приводит
- 67. Рис. Восстановление глутатиона с участием НАДФН
- 68. Благодарю за внимание
- 71. Общие пути катаболизма Окисление продуктов окисления глюкозы гликолизом ПВК в общих путях катаболизма в митохондриях: Пируват
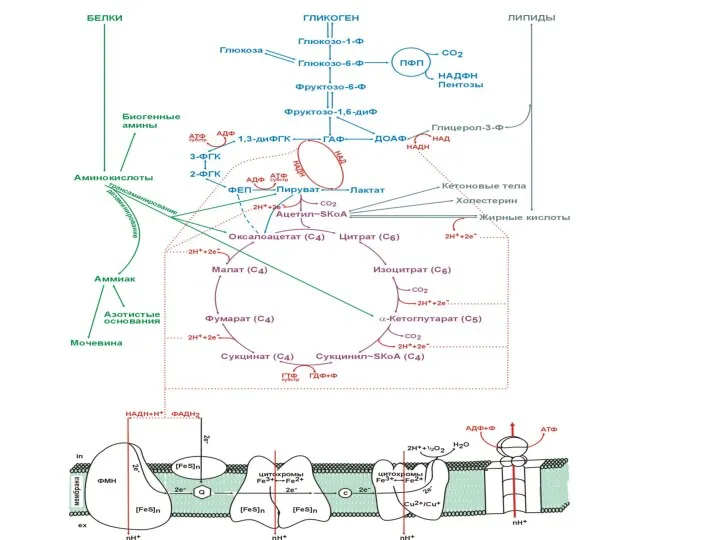
- 72. Карта катаболизма
- 73. Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:
- 74. Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в пируватдегидрогеназный комплекс (ПДК):
- 75. Этот комплекс ферментов работает подобно конвейеру, в котором продукт передается от фермента к ферменту. Такой принцип
- 76. Дигидролипоат-ацетилтрансфераза (2) - второй фермент комплекса. Катализирует перенос ацетильного остатка, соединенного с липоевой кислотой на второй
- 77. Дегидрогеназа дигидролипоевой кислоты (3й фермент) отщепляет водород от липоевой кислоты и переносит его на НАД+. Далее
- 78. Цикл лимонной кислоты Цитратный цикл (цикл Кребса, цикл трикарбоновых кислот (ЦТК) – это система реакций, приводящая
- 79. В ходе одного полного цитратного цикла происходит: 1. полное окисление ацетильного остатка до двух молекул СО2;
- 80. Цикл лимонной кислоты / цикл Кребса
- 81. Энергетика полного окисления глюкозы в клетке 1. Гликолиз (распад глю до 2х ПВК) – 8 АТФ
- 82. Ферменты цитратного цикла 1 – цитратсинтаза, 2 – аконитаза, 3 – изоцитратдегидрогеназа, 4 – a-кетоглутаратдегидрогеназный комплекс,
- 83. Сопряжение общих путей катаболизма с дыхательной цепью В общих путях катаболизма происходит пять реакций дегидрирования: (одна
- 84. Энергетика цитратного цикла и общих путей катаболизма За один оборот цитратного цикла синтезируется 12 молекул АТФ.
- 85. Кроме того, в цитратном цикле происходит одна реакция субстратного фосфорилирования, дающая 1 моль ГТФ ( аналог
- 86. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 87. задание на дом: записать все реакции ПДК и цикла Кребса Благодарю за внимание
- 88. Использование метаболитов ЦТК в синтезе различных соединений
- 89. Метаболизм глюкозы
- 90. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 91. Синтез и распад гликогена
- 92. Синтез гликогена гликогенез
- 94. Скачать презентацию

План лекции 5:
Гликоген. Особенности строения
Синтез и распад гликогена
Глюконеогенез
Пентозофосфатный путь
Дополнение к лекции
План лекции 5:
Гликоген. Особенности строения
Синтез и распад гликогена
Глюконеогенез
Пентозофосфатный путь
Дополнение к лекции
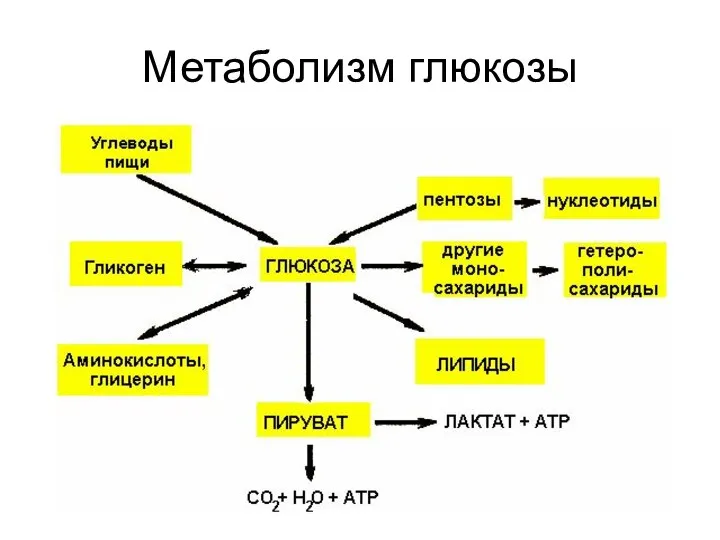
Метаболизм глюкозы
Метаболизм глюкозы
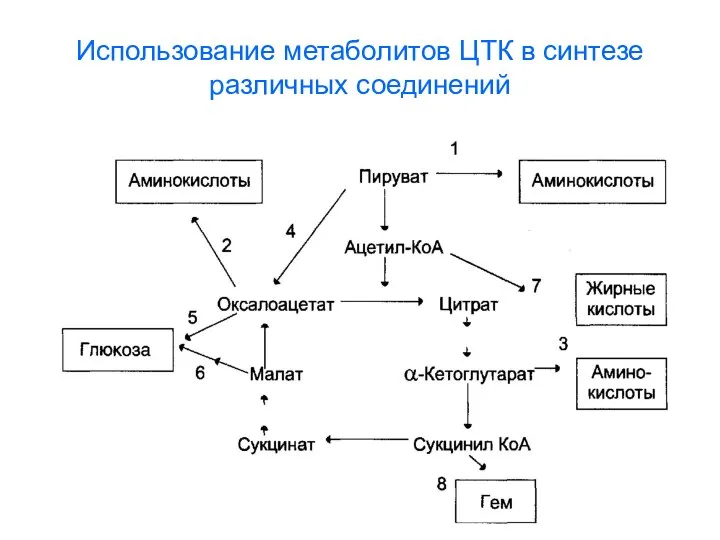
Использование метаболитов ЦТК в синтезе различных соединений
Использование метаболитов ЦТК в синтезе различных соединений
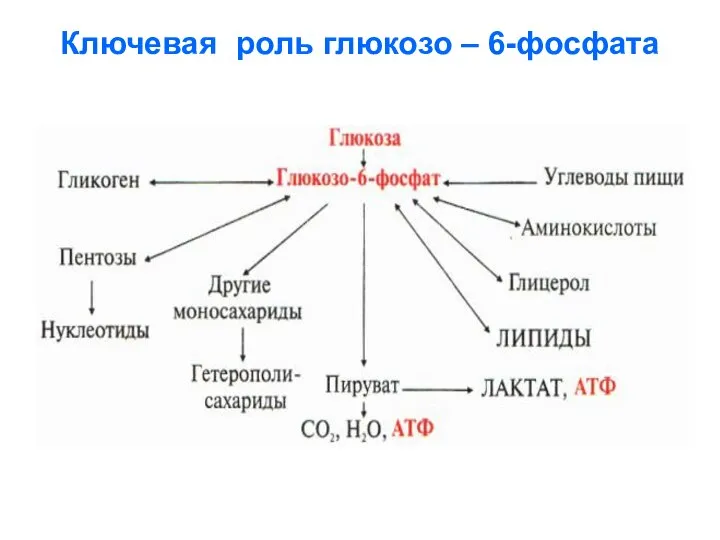
Ключевая роль глюкозо – 6-фосфата
Ключевая роль глюкозо – 6-фосфата
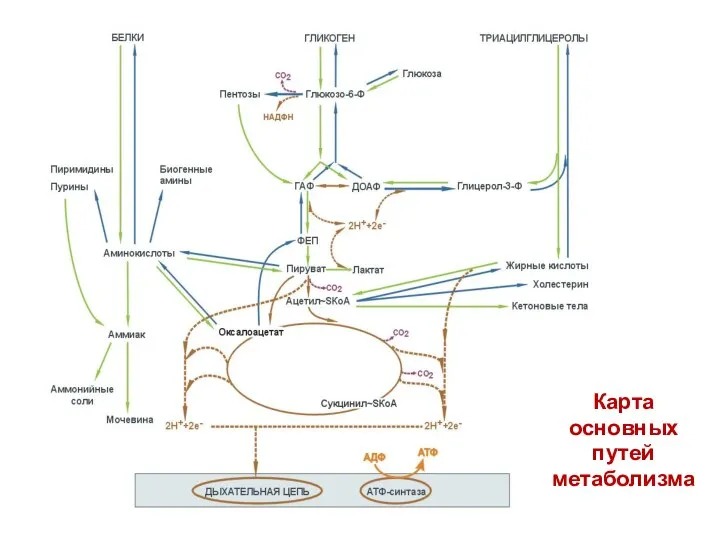
Карта
основных
путей
метаболизма
Карта
основных
путей
метаболизма
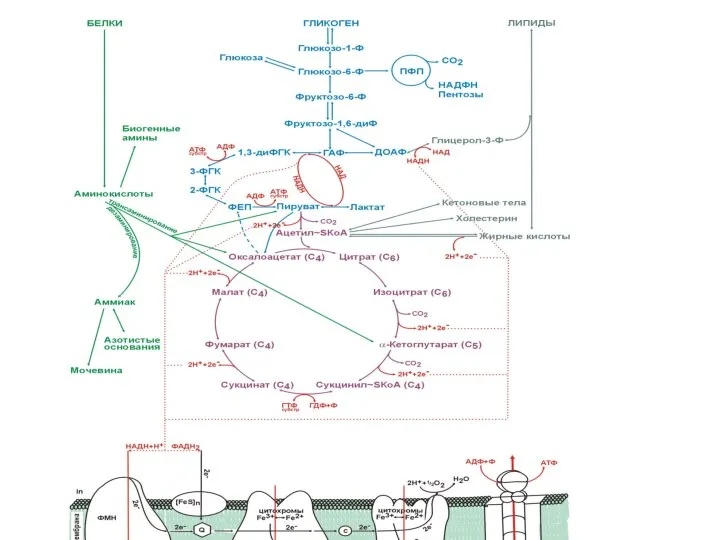
Обмен углеводов
Обмен углеводов

1. Обмен гликогена
Гликогенез и
гликогенолиз
1. Обмен гликогена
Гликогенез и
гликогенолиз

Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид).
Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид).

В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение
В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение

Линейные участки молекулы гликогена образованы
остатками глюкозы, связанными
альфа-1,4-гликозидными связями, а
Линейные участки молекулы гликогена образованы
остатками глюкозы, связанными
альфа-1,4-гликозидными связями, а

Синтез и распад гликогена
Синтез и распад гликогена гормонозависимый процессы и
Синтез и распад гликогена
Синтез и распад гликогена гормонозависимый процессы и

Синтез гликогена - гликогенез
Синтез гликогена ускоряется в период
пищеварения (абсорбтивный период –
Синтез гликогена - гликогенез
Синтез гликогена ускоряется в период
пищеварения (абсорбтивный период –

Когда длина синтезируемой цепи увеличивается на
11-12 остатков глюкозы, фермент ветвления
Когда длина синтезируемой цепи увеличивается на
11-12 остатков глюкозы, фермент ветвления
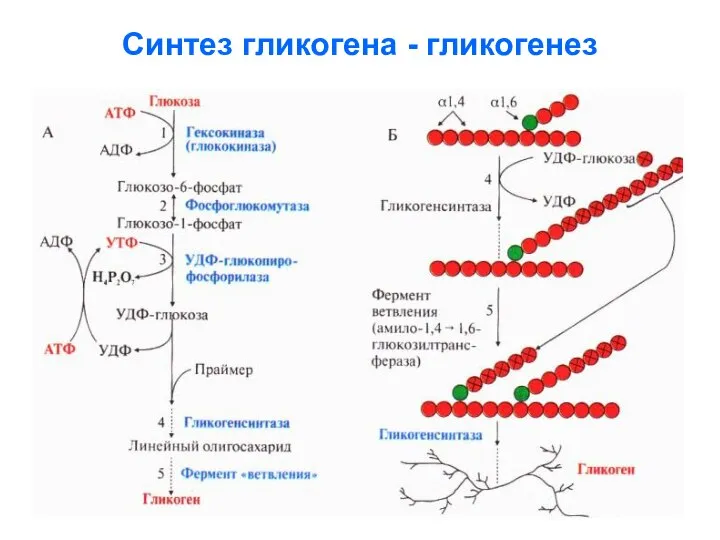
Синтез гликогена - гликогенез
Синтез гликогена - гликогенез
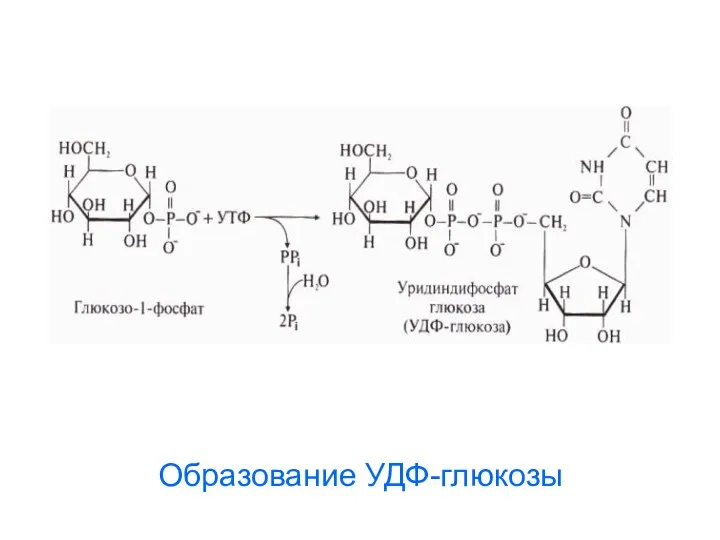
Образование УДФ-глюкозы
Образование УДФ-глюкозы

Далее подробно расписаны реакции гликогенеза:
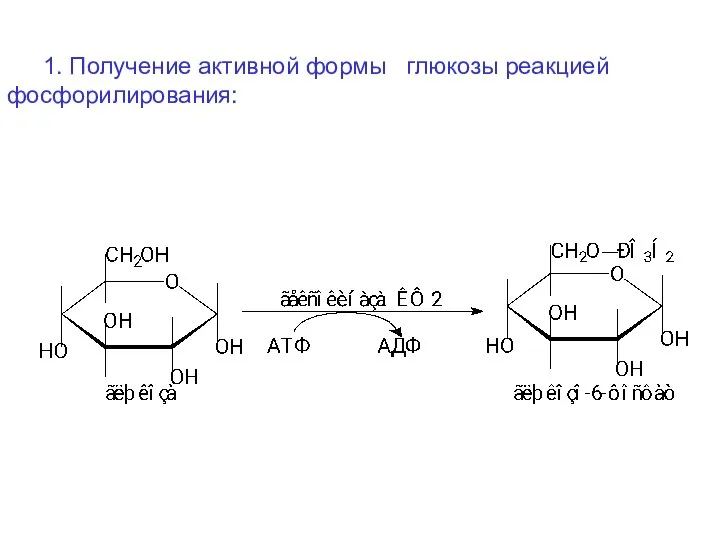
1. Получение активной формы глюкозы реакцией фосфорилирования:
1. Получение активной формы глюкозы реакцией фосфорилирования:
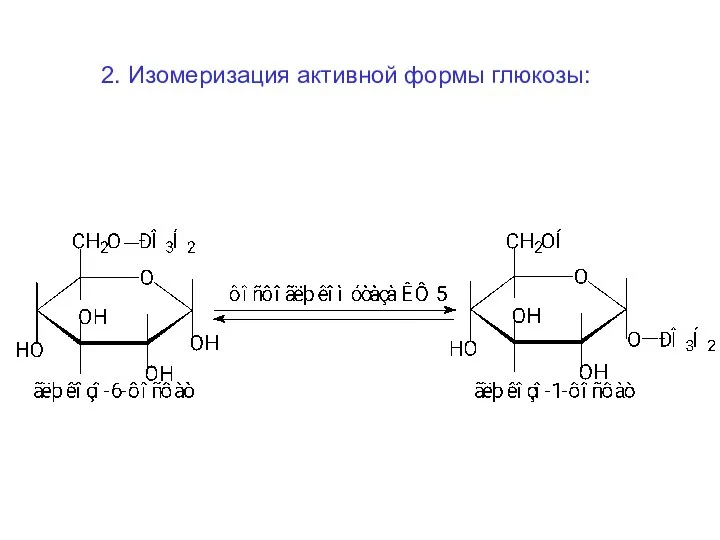
2. Изомеризация активной формы глюкозы:
2. Изомеризация активной формы глюкозы:
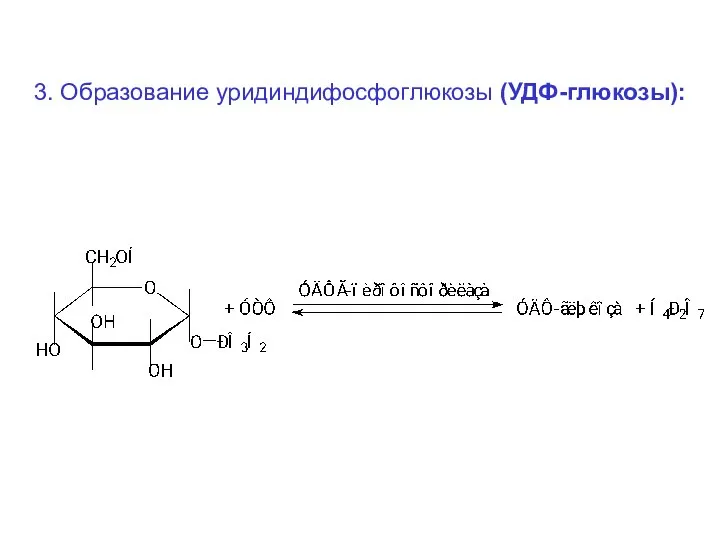
3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
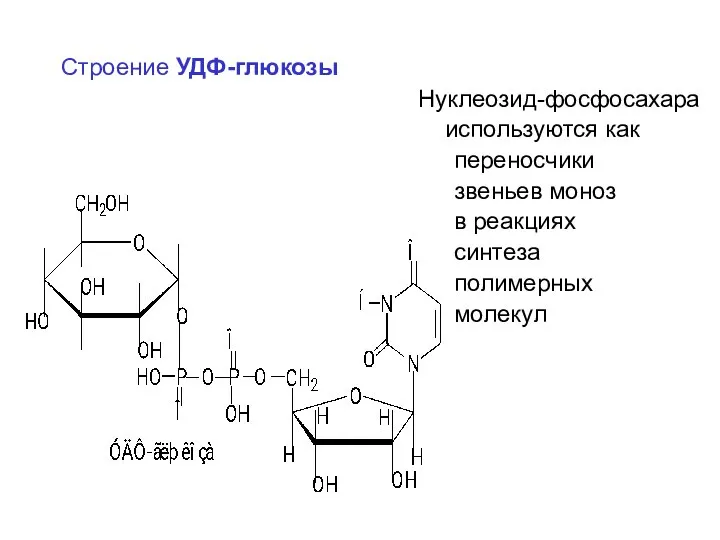
Строение УДФ-глюкозы
Нуклеозид-фосфосахара
используются как
переносчики
звеньев моноз
в реакциях
синтеза
полимерных
молекул
Строение УДФ-глюкозы
Нуклеозид-фосфосахара
используются как
переносчики
звеньев моноз
в реакциях
синтеза
полимерных
молекул

4. Перенос остатка глюкозы с УДФ-глюкозы на
затравочную цепь гликогена, которая
4. Перенос остатка глюкозы с УДФ-глюкозы на затравочную цепь гликогена, которая

Ветвление гликогена
5. Синтез α-(1,6)-гликозидных связей происходит при
участии гликоген-ветвящегося фермента –
гликоген-трансферазы, которая
Ветвление гликогена
5. Синтез α-(1,6)-гликозидных связей происходит при
участии гликоген-ветвящегося фермента –
гликоген-трансферазы, которая

Подводим итог:
Гликоген синтезируется в период пищеварения
(в течение 1-2 часов после
Подводим итог:
Гликоген синтезируется в период пищеварения
(в течение 1-2 часов после

Для подготовки и включения глюкозы в растущую
полисахаридную цепь требуется энергия
Для подготовки и включения глюкозы в растущую
полисахаридную цепь требуется энергия

Необходимость превращения глюкозы в гликоген
связана с тем, что накопление значительного
Необходимость превращения глюкозы в гликоген
связана с тем, что накопление значительного

Распад гликогена - гликогенолиз
или мобилизация гликогена – происходит в основном
в период
Распад гликогена - гликогенолиз
или мобилизация гликогена – происходит в основном
в период
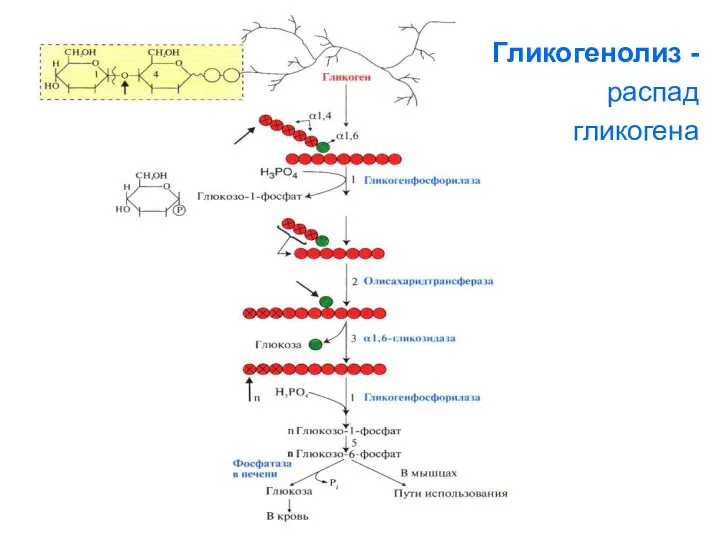
Гликогенолиз -
распад
гликогена
Гликогенолиз -
распад
гликогена

1. Освобождение глюкозы в виде глюкозо-1-фосфата
(реакция 5) происходит в результате
1. Освобождение глюкозы в виде глюкозо-1-фосфата
(реакция 5) происходит в результате
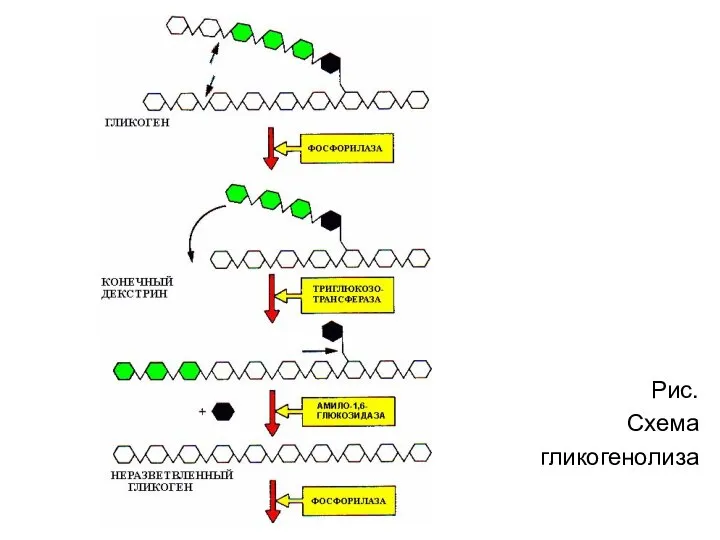
Рис.
Схема
гликогенолиза
Схема
гликогенолиза

Мобилизация гликогена в печени и мыщцах отличаются
В печени есть фермент
Мобилизация гликогена в печени и мыщцах отличаются
В печени есть фермент

Постоянный уровень глюкозы в крови в постабсорбтивный
период = 3,5-5,5 мМ/л
Органы
период = 3,5-5,5 мМ/л
Органы

Функция мышечного гликогена заключается в
высвобождении глюкозо-6фосфата, используемого в
самой мышце
Функция мышечного гликогена заключается в
высвобождении глюкозо-6фосфата, используемого в
самой мышце

Рис.Особенности мобилизации гликогена в печени и мышцах
Рис.Особенности мобилизации гликогена в печени и мышцах

Гормоны регулирующие обмен гликогена
Переключение процессов синтеза и мобилизации
гликогена в печени
Гормоны регулирующие обмен гликогена
Переключение процессов синтеза и мобилизации
гликогена в печени
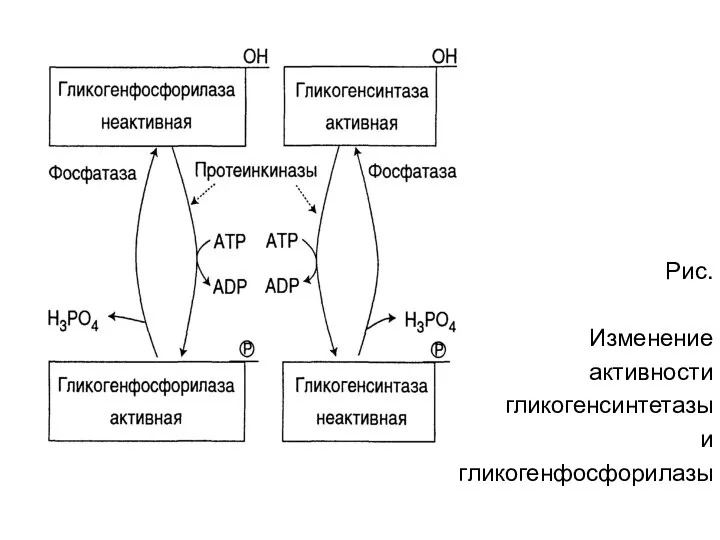
Рис.
Изменение
активности
гликогенсинтетазы
и
гликогенфосфорилазы
Рис.
Изменение
активности
гликогенсинтетазы
и
гликогенфосфорилазы

Первичным сигналом для синтеза инсулина и
глюкагона является изменение концентрации глюкозы
Первичным сигналом для синтеза инсулина и
глюкагона является изменение концентрации глюкозы

В период пищеварения преобладающим является
влияние инсулина, и инсулин/глюкагоновый индекс в
В период пищеварения преобладающим является
влияние инсулина, и инсулин/глюкагоновый индекс в

При интенсивной мышечной работе и стрессе в кровь
из мозгового вещества
При интенсивной мышечной работе и стрессе в кровь
из мозгового вещества

Болезни углеводного обмена
Причины:
нарушения переваривания и всасывания углеводов,
синтеза и распада гликогена.
- ферментопатии,
-
Болезни углеводного обмена
Причины:
нарушения переваривания и всасывания углеводов,
синтеза и распада гликогена.
- ферментопатии,
-
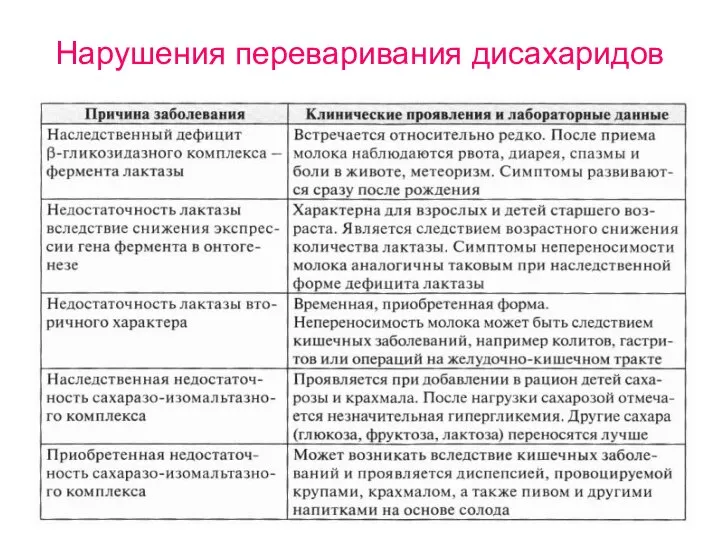
Нарушения переваривания дисахаридов
Нарушения переваривания дисахаридов
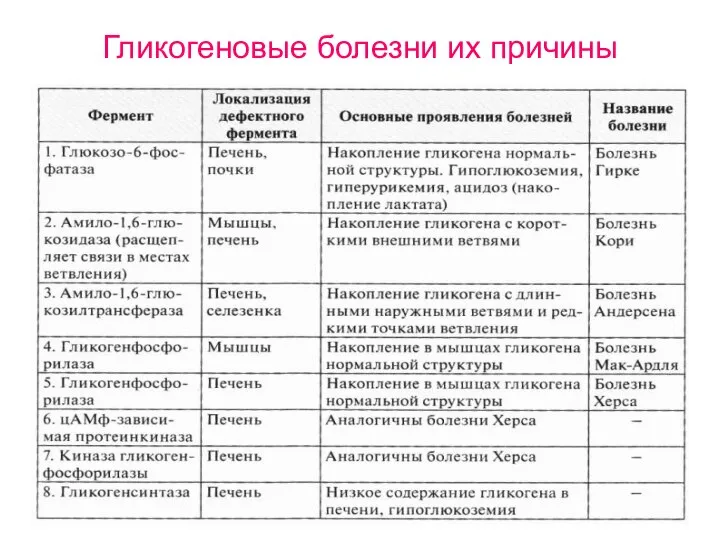
Гликогеновые болезни их причины
Гликогеновые болезни их причины

2. ГЛЮКОНЕОГЕНЕЗ
2. ГЛЮКОНЕОГЕНЕЗ
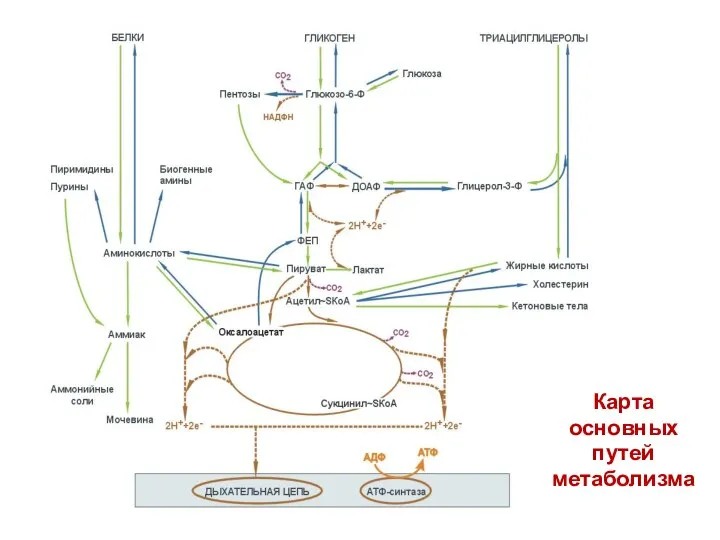
Карта
основных
путей
метаболизма
Карта
основных
путей
метаболизма
Обмен углеводов
Обмен углеводов

Глюконеогенез
Глюконеогенез – процесс синтеза глюкозы из веществ
неуглеводной природы.
Субстраты глюконеогенеза: пируват, лактат,
глицерол, аминокислоты.
Важнейшей
Глюконеогенез
Глюконеогенез – процесс синтеза глюкозы из веществ
неуглеводной природы.
Субстраты глюконеогенеза: пируват, лактат,
глицерол, аминокислоты.
Важнейшей

Процесс протекает главным образом в печени и менее
интенсивно в корковом веществе
Процесс протекает главным образом в печени и менее
интенсивно в корковом веществе

Большинство реакций гликолиза и гликонеогенеза
являются обратимыми и катализируются одними и
Большинство реакций гликолиза и гликонеогенеза
являются обратимыми и катализируются одними и
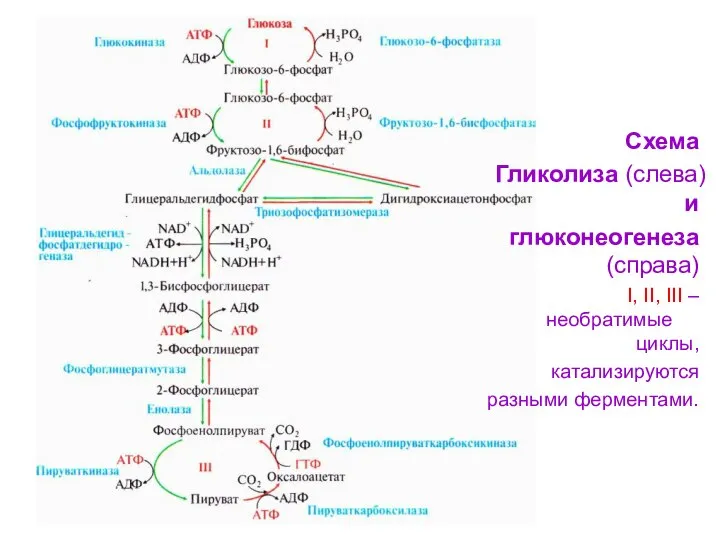
Схема
Гликолиза (слева) и
глюконеогенеза (справа)
I, II, III – необратимые циклы,
катализируются
разными
Схема
Гликолиза (слева) и
глюконеогенеза (справа)
I, II, III – необратимые циклы,
катализируются
разными

В митохондриях под действием ферментов
малатдегидроеназы и аминотрансферазы образуется
малат и
В митохондриях под действием ферментов
малатдегидроеназы и аминотрансферазы образуется
малат и
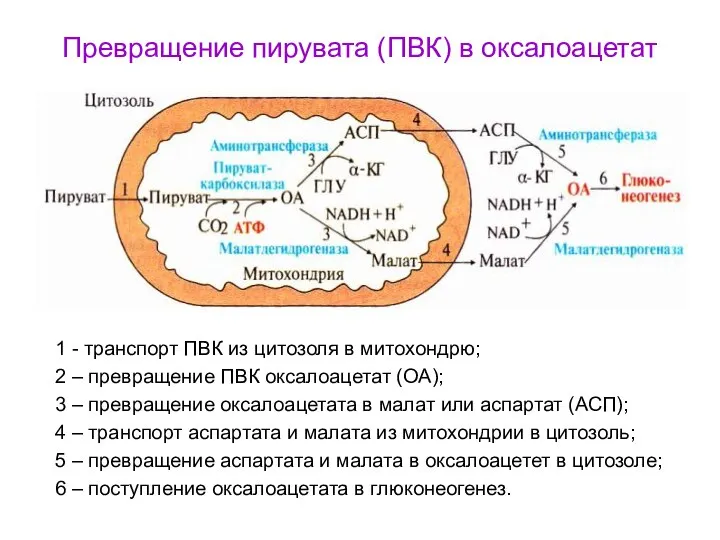
Превращение пирувата (ПВК) в оксалоацетат
1 - транспорт ПВК из цитозоля в
Превращение пирувата (ПВК) в оксалоацетат
1 - транспорт ПВК из цитозоля в

Рис.Суммарное уравнение образования глюкозы из пирувата
Видно, что синтез глюкозы энергозатратный процесс
Рис.Суммарное уравнение образования глюкозы из пирувата
Видно, что синтез глюкозы энергозатратный процесс

Использование лактата в качестве субстрата в
глюконеогенезе связано с транспортом его в
Использование лактата в качестве субстрата в
глюконеогенезе связано с транспортом его в
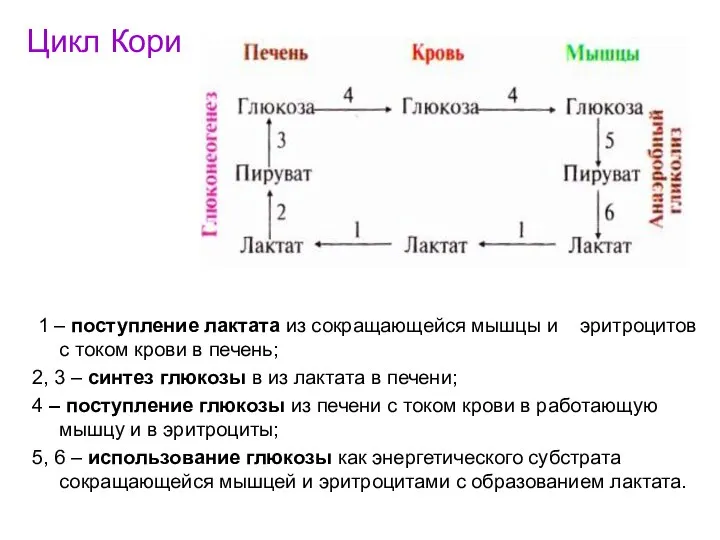
Цикл Кори
1 – поступление лактата из сокращающейся мышцы и эритроцитов
Цикл Кори
1 – поступление лактата из сокращающейся мышцы и эритроцитов

Часть ПВК, образовавшегося из лактата, окисляется в
печени до СО2 и
Часть ПВК, образовавшегося из лактата, окисляется в
печени до СО2 и

Снижение использования лактата в качестве субстрата
в синтезе глюкозы, вызванное дефектом
Снижение использования лактата в качестве субстрата
в синтезе глюкозы, вызванное дефектом

Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие:
тканевой
Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие:
тканевой

3. Пентозофосфатный путь
в метаболизме глюкозы (ПФП)
Это альтернативный путь окиления глюкозы.
3. Пентозофосфатный путь
в метаболизме глюкозы (ПФП)
Это альтернативный путь окиления глюкозы.

Кроме того, пентозофосфатный путь поставляет клетке
пентозофосфаты (пентозы) ,
необходимые для
Кроме того, пентозофосфатный путь поставляет клетке
пентозофосфаты (пентозы) ,
необходимые для

В пентозофосфатном пути превращения глюкозы можно
выделить две части:
А –
В пентозофосфатном пути превращения глюкозы можно
выделить две части:
А –
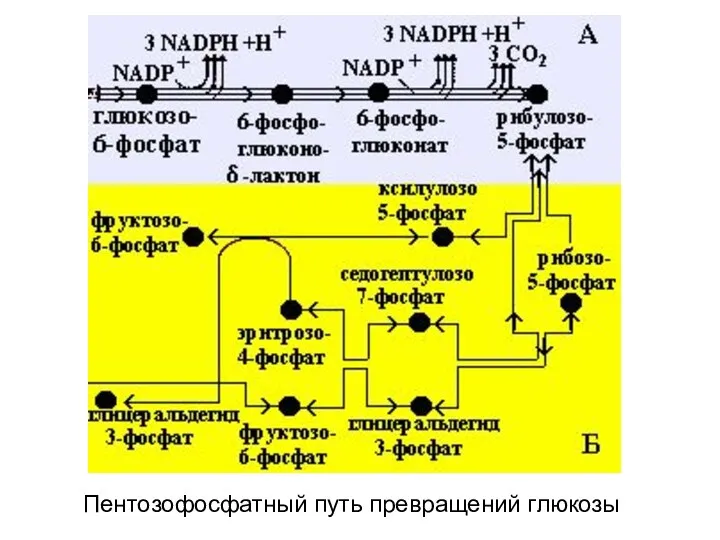
Пентозофосфатный путь превращений глюкозы
Пентозофосфатный путь превращений глюкозы
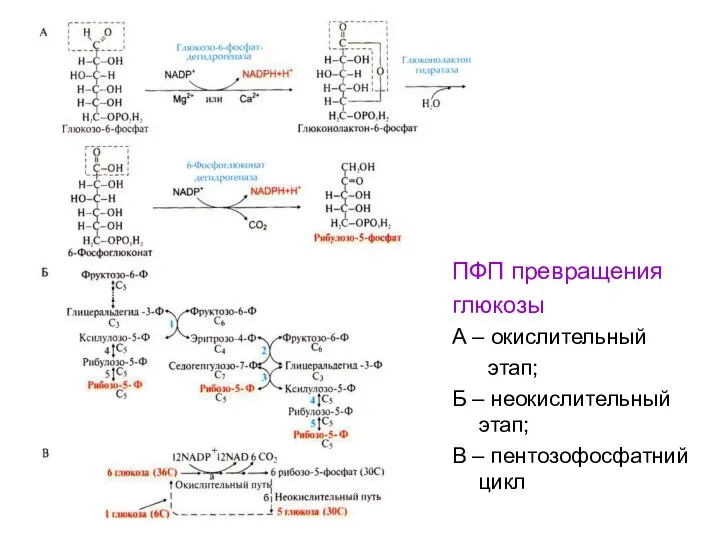
ПФП превращения
глюкозы
А – окислительный
этап;
Б – неокислительный этап;
В – пентозофосфатний цикл
ПФП превращения
глюкозы
А – окислительный
этап;
Б – неокислительный этап;
В – пентозофосфатний цикл

Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает несколько
Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает несколько

3глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат +
3глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат +

ПФП в эритроцитах
ПФП в эритроцитах очень важен, так как
восстановленный НАДФН
ПФП в эритроцитах
ПФП в эритроцитах очень важен, так как
восстановленный НАДФН

Дефект первого фермента ПФП
глюкозо-6-фосфат дегидрогеназы приводит к
дефициту НАДФН и невозможности
Дефект первого фермента ПФП
глюкозо-6-фосфат дегидрогеназы приводит к
дефициту НАДФН и невозможности
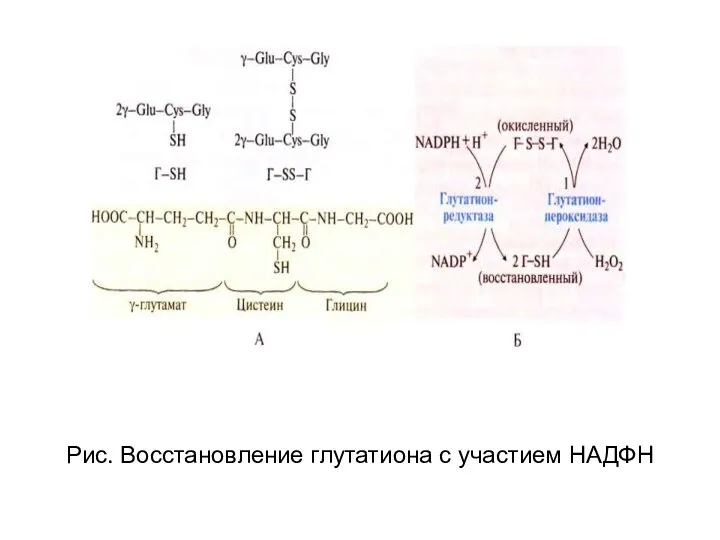
Рис. Восстановление глутатиона с участием НАДФН
Рис. Восстановление глутатиона с участием НАДФН

Благодарю за внимание



Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих путях
Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих путях
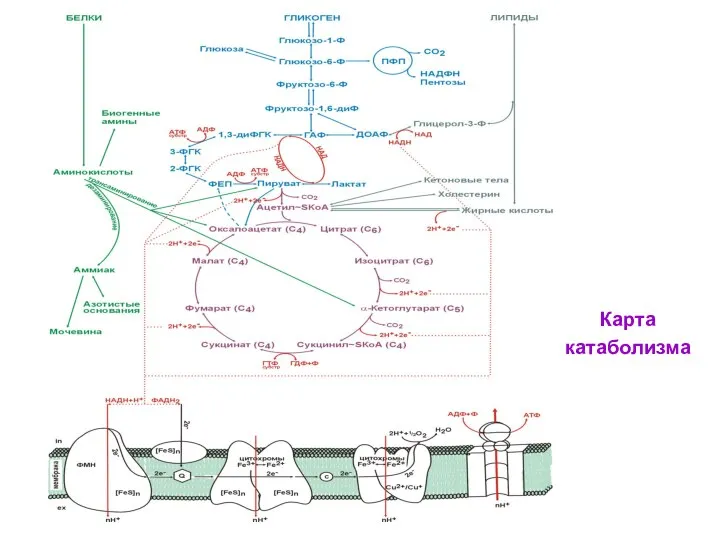
Карта
катаболизма
Карта
катаболизма

Окислительное декарбоксилирование пировиноградной кислоты
Суммарный результат многостадийной
реакции выглядит следующим образом:
Окислительное декарбоксилирование пировиноградной кислоты
Суммарный результат многостадийной
реакции выглядит следующим образом:
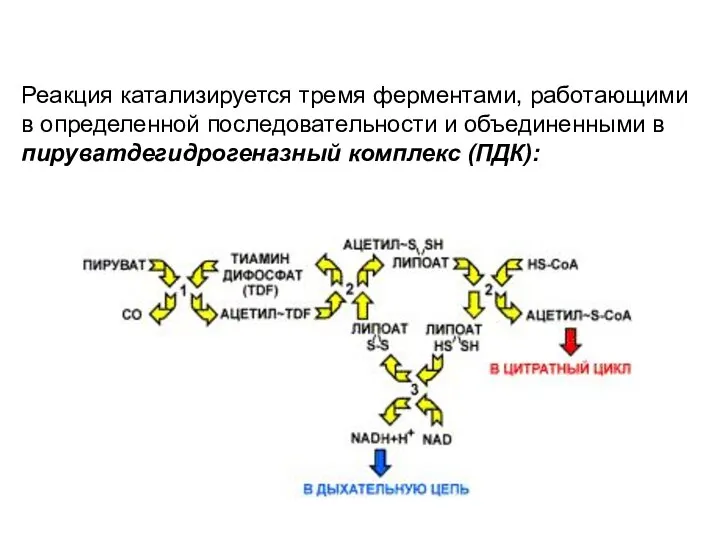
Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в
Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в

Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается от
Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается от

Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,
Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,

Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты и
Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты и

Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК) –
это
Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК) –
это

В ходе одного полного цитратного цикла происходит:
1. полное окисление ацетильного
В ходе одного полного цитратного цикла происходит:
1. полное окисление ацетильного
Цикл лимонной кислоты / цикл Кребса
Цикл лимонной кислоты / цикл Кребса

Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до 2х
Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до 2х

Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4 –
Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4 –

Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма происходит
Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма происходит

Энергетика цитратного цикла и общих путей катаболизма
За один оборот цитратного
Энергетика цитратного цикла и общих путей катаболизма
За один оборот цитратного

Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая
Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая
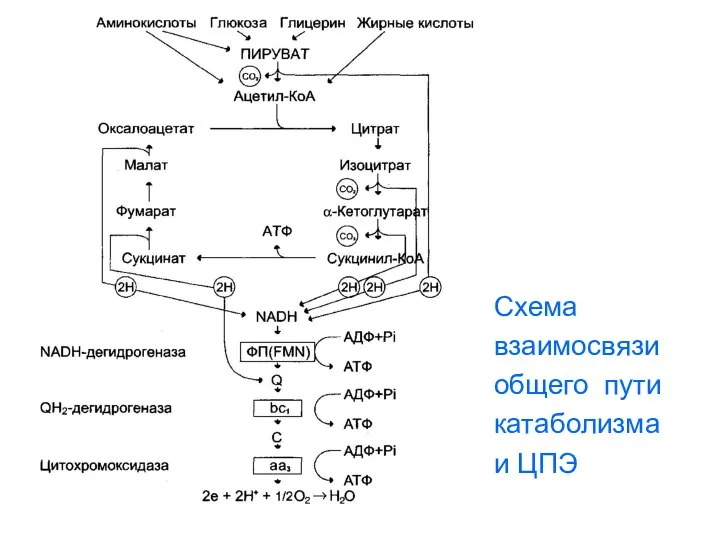
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ

задание на дом:
записать все реакции ПДК и цикла Кребса
Благодарю за внимание
задание на дом:
записать все реакции ПДК и цикла Кребса
Благодарю за внимание
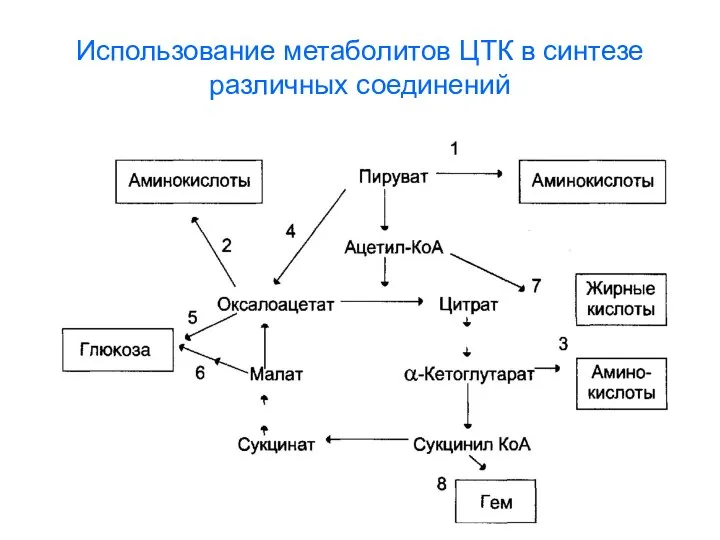
Использование метаболитов ЦТК в синтезе различных соединений
Использование метаболитов ЦТК в синтезе различных соединений
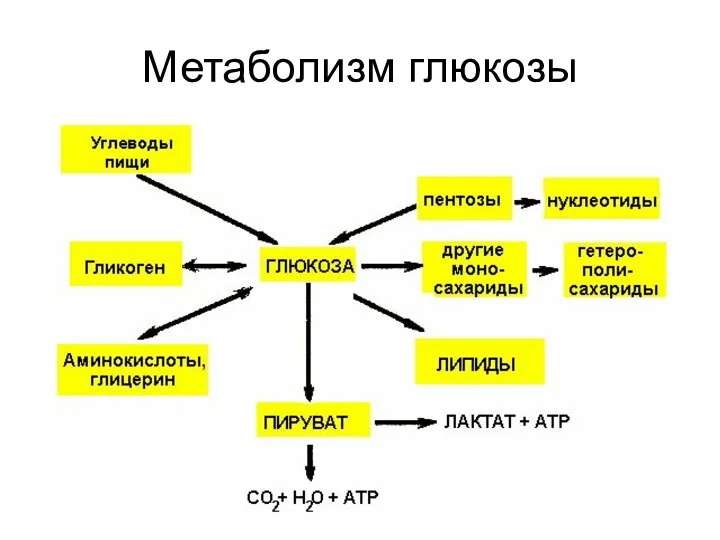
Метаболизм глюкозы
Метаболизм глюкозы
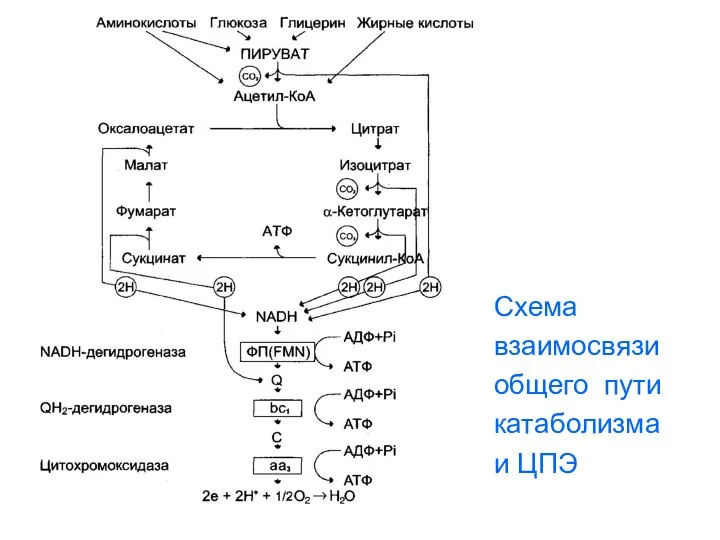
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
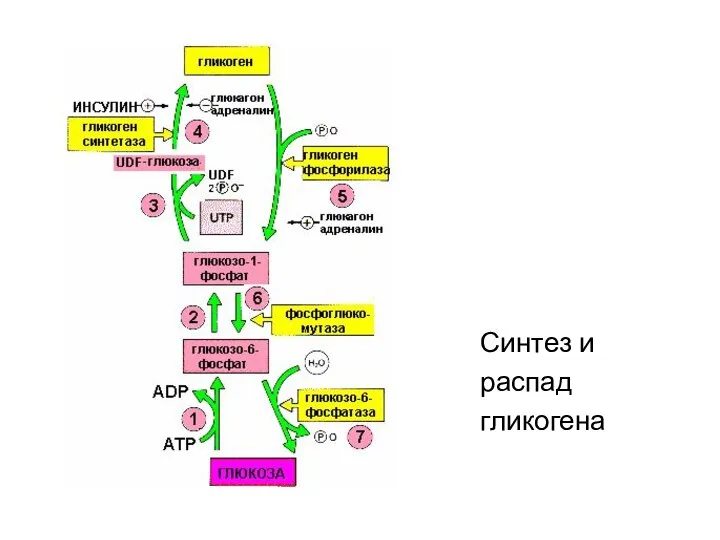
Синтез и
распад
гликогена
Синтез и
распад
гликогена
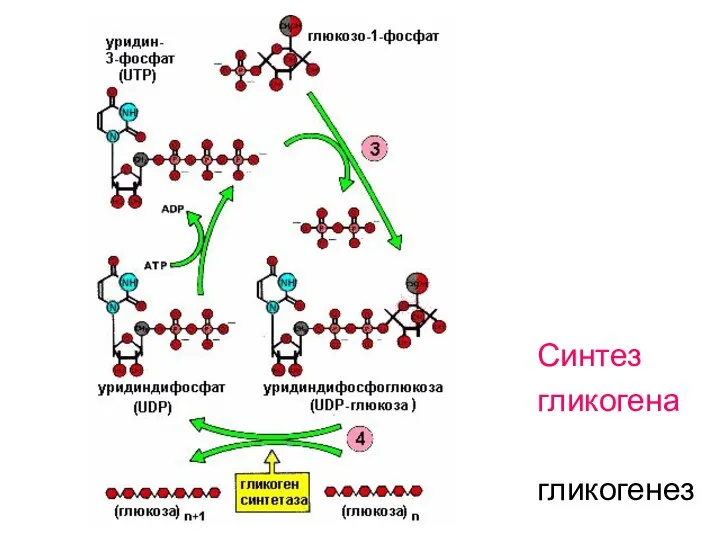
Синтез
гликогена
гликогенез
Синтез
гликогена
гликогенез
 Формирование сведений о ДО в зависимости от условий государственного контракта (договора )
Формирование сведений о ДО в зависимости от условий государственного контракта (договора ) Арома масла в спорте
Арома масла в спорте Основные понятия математической статистики: оценки параметров распределения, проверка гипотез, системы случайных величин: коррел
Основные понятия математической статистики: оценки параметров распределения, проверка гипотез, системы случайных величин: коррел Русская прялка
Русская прялка Разработка управленческих решений Введение в курс Проф. Дульзон А.А.
Разработка управленческих решений Введение в курс Проф. Дульзон А.А.  История создания “Каролины”
История создания “Каролины”  Презентация Состав основных фондов
Презентация Состав основных фондов  Поиск информации в интернете
Поиск информации в интернете Қазақ халқының қолданбалы-сәндік өнері
Қазақ халқының қолданбалы-сәндік өнері Спорт и допинг
Спорт и допинг Нормализация Этапы проектирования
Нормализация Этапы проектирования ВКР: Проблемы стимулирования деятельности государственных служащих
ВКР: Проблемы стимулирования деятельности государственных служащих Интерфейс Делфи 7
Интерфейс Делфи 7 ДРАГОЦЕННЫЕ МЕТАЛЛЫ И КАМНИ
ДРАГОЦЕННЫЕ МЕТАЛЛЫ И КАМНИ Романтический стиль одежды
Романтический стиль одежды HTML - Язык разметки гипертекста
HTML - Язык разметки гипертекста Тренинг Игоря Зорина «Управленческие поединки» 26-27 февраля 2011 года Центр развития персонала «Бизнес-Инсайт»
Тренинг Игоря Зорина «Управленческие поединки» 26-27 февраля 2011 года Центр развития персонала «Бизнес-Инсайт»  Исторические личности XVIII века
Исторические личности XVIII века Брагар С. Диденко А. Трофимова Е.
Брагар С. Диденко А. Трофимова Е. Информационная безопасность на предприятии
Информационная безопасность на предприятии  Daň z příjmů fyzických osob
Daň z příjmů fyzických osob Живопись-это немая поэзия , а поэзия – говорящая живопись (Вольтер .)
Живопись-это немая поэзия , а поэзия – говорящая живопись (Вольтер .) Святитель Стефан Пермский
Святитель Стефан Пермский Политическая наука на Кубе
Политическая наука на Кубе Признаки предметов по цвету и размеру
Признаки предметов по цвету и размеру Особливості усної і писемної наукової мови
Особливості усної і писемної наукової мови Идея и схема опыта Майкельсона-Морли
Идея и схема опыта Майкельсона-Морли  Бухгалтерские и юридические услуги, кадровый учет, услуги эколога для ИП, ООО (ОДО), ЧУП
Бухгалтерские и юридические услуги, кадровый учет, услуги эколога для ИП, ООО (ОДО), ЧУП