- Обмен углеводов

Содержание
- 2. План лекции 6: Гипер- и гипогликемия: причины возникновения, механизмы компенсации. Метаболические и клинические последствия острых и
- 3. Гипер- и гипогликемия: причины возникновения, механизмы компенсации Концентрация глюкозы в артериальной крови в течение суток поддерживается
- 4. Рис. Динамика изменений концентрации глюкозы в течение суток: А-Б-В – период пищеварения (абсорбтивный период); В-Г –
- 5. Снижение концентрации глюкозы в крови ниже 80мг/дл или 3,3 мМоль/л называется гипоглюкоземией. И это более опасное
- 6. Заболевания вызывающие повышение концентрации глюкозы в крови Гипреглюкоземия - повышение сахара в крови, появление глюкозы в
- 7. Заболевания вызывающие понижение концентрации глюкозы в крови Гипогликемия – понижение содержания сахара в крови – нередко
- 8. Постоянное поступление глюкозы необходимо в качестве основного источника энергии для нервной ткани и эритроцитам. При понижении
- 10. Рис. Пути использования основных энергоносителей в абсорбтивном периоде 1- повышение синтеза гликогена в печени, 2- гликолиз,
- 11. Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное 1 – снижение инсулин-глюкагонового индекса,
- 12. Рис. Изменение метаболизма основных энергоносителей при голодании 1- снижение инсулин-глюкагонового индекса, 2- мобилизация гликогена, 3,4- транспорт
- 13. природа гормонов инсулина и глюкагона; механизм регуляции концентрации глюкозы гормонами. В зависимости от физиологического состояния концентрация
- 14. Регуляция синтез и распада гликогена Гликоген - депо глюкозы в организме
- 15. Метаболизм гликогена в печени, мышцах и др.клетках регулируется несколькими гормонами, одни из которых активируют синтез, а
- 16. Присоединение фосфатов к ферменту производят протеинкиназы, источником фосфора является АТФ: фосфорилаза гликогена активируется (+) после присоединения
- 17. Например: во время мышечной работы адреналин вызывает фосфорилирование внутримышечных ферментов обмена гликогена. В результате фосфорилаза гликогена
- 18. Способы активации синтазы гликогена Гликогенсинтаза при присоединении фосфата прекращает работу, т.е. она активна в дефосфорилированном виде.
- 19. Рис. Изменение активности ферментов обмена гликогена в зависимости от условий ферменты фосфорилаза гликогена и гликогенсинтаза реципрокны.
- 20. Инсулин Вырабатывается Бета клетками островков Лангерганса панкреатической (поджелудочной) железой. Состоит из 51 остатка АК мол Масса
- 21. СТРУКТУРА ИНСУЛИНА Инсулин - полипептид, состоит из 2-х цепей, включает 51 АК остаток. a-Цепь содержит 21
- 23. Глюкагон
- 24. адреналин
- 25. Изменение гормонального статуса и метаболизма при сахарном диабете Сахарный диабет (СД) возникает вследствие относительного или абсолютного
- 26. ИЗСД - СД I типа Является следствием разрушения бета-клеток островков Лангерганса в результате аутоиммунных реакций. ПРИЧИНЫ
- 27. ИНСД – СД II типа ПРИЧИНЫ Развивается вследствие нарушения: превращения проинсулина в инсулин, регуляции секреции инсулина,
- 28. Факторы увеличивающие риск развития ИНСД: ожирение, неправильный режим питания, гиподинамия, стрессы. Обычно после 40 лет. Симптомы
- 29. Изменения метаболизма при СД 1. соотношения инсулин-глюкагон (инсулин/глюкагон) снижено. При этом ослабевает стимуляция процессов депонирования гликогена
- 30. Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное 1 – снижение инсулин-глюкагонового индекса,
- 31. Рис. Изменение метаболизма основных энергоносителей при голодании 1- снижение инсулин-глюкагонового индекса, 2- мобилизация гликогена, 3,4- транспорт
- 32. Симптомы СД Гиперглюкоземия Кетонемия Гиперлипопротеинемия Азотемия Полиурия
- 33. Симптомы СД 1. Гиперглюкоземия Для всех форм диабета характерно повышение концентрации глюкозы в крови – гиперглюкоземия,
- 35. 2. Кетонемия характерный признак СД. При низком соотношении инсулин-глюкагонового индекса жиры не депонируются, ускоряется их катаболизм,
- 36. 3. Гиперлипопротеинемия Пищевые жиры не депонируются в жировой ткани в следствие ослабления процессов запасания и низкой
- 37. 5. Полиурия Для выведения большого количества глюкозы, кетоновых тел, мочевины требуется большой объем жидкости, в результате
- 38. Острые осложнения СД Механизм развития диабетической комы. Нарушения метаболизма углеводов, жиров и белков при СД могут
- 39. Коматозные состояния при СД кетоацидотическая кома, гиперосмолярная кома, лактоацидотическая кома. Для кетоацидотической формы характены выраженный дефицит
- 40. Рис. Изменения метаболизма при сахарном диабете и причины диабетической комы
- 41. Поздние осложнения СД Причина - длительная гиперглюкоземия. Следствие – инвалидизация. Из-за повреждения кровеносных сосудов и следовательно
- 42. Степень гликозилирования белков зависит от скорости их обновления. Также гликозилируются: - белки базальных мембран, - клисталлины
- 43. Для сохранения осмомолярности начинается компенсаторное перемещение жидкости из клеток и внеклеточного пространства в сосудистое русло. Это
- 44. Диабетические ангиопатии Макроангиопатии: снижение эластичности артерий, повреждение крупных и мелких сосудов мозга, сердца, нижних конечностей. Следствие
- 45. Благодарю за внимание
- 46. Метаболизм глюкозы
- 47. Ключевая роль глюкозо – 6-фосфата
- 48. Карта основных путей метаболизма
- 49. Обмен углеводов
- 50. 1. Обмен гликогена Гликогенез и гликогенолиз
- 51. Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид). У растений эту же функцию
- 52. В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение при распаде гликогена большого количества
- 53. Линейные участки молекулы гликогена образованы остатками глюкозы, связанными альфа-1,4-гликозидными связями, а в местах разветвления – альфа
- 54. Синтез и распад гликогена Синтез и распад гликогена гормонозависимый процессы и регулируются инсулином и глюкагоном (и
- 55. Синтез гликогена - гликогенез Синтез гликогена ускоряется в период пищеварения (абсорбтивный период – 1-2 часа после
- 56. Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент ветвления переносит фрагмент из 5-6 остатков
- 57. Синтез гликогена - гликогенез
- 58. Образование УДФ-глюкозы
- 59. Далее подробно расписаны реакции гликогенеза:
- 60. 1. Получение активной формы глюкозы реакцией фосфорилирования:
- 61. 2. Изомеризация активной формы глюкозы:
- 62. 3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
- 63. Строение УДФ-глюкозы Нуклеозид-фосфосахара используются как переносчики звеньев моноз в реакциях синтеза полимерных молекул
- 64. 4. Перенос остатка глюкозы с УДФ-глюкозы на затравочную цепь гликогена, которая содержит не менее 4-х звеньев
- 65. Ветвление гликогена 5. Синтез α-(1,6)-гликозидных связей происходит при участии гликоген-ветвящегося фермента – гликоген-трансферазы, которая переносит концевой
- 66. Подводим итог: Гликоген синтезируется в период пищеварения (в течение 1-2 часов после приема углеводной пищи). Гликогенез
- 67. Для подготовки и включения глюкозы в растущую полисахаридную цепь требуется энергия 1 моль АТФ и 1
- 68. Необходимость превращения глюкозы в гликоген связана с тем, что накопление значительного количества глюкозы в клетке привело
- 69. Распад гликогена - гликогенолиз или мобилизация гликогена – происходит в основном в период между приёмами пищи
- 70. Гликогенолиз - распад гликогена
- 71. 1. Освобождение глюкозы в виде глюкозо-1-фосфата (реакция 5) происходит в результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет
- 72. Рис. Схема гликогенолиза
- 73. Мобилизация гликогена в печени и мыщцах отличаются В печени есть фермент глюкозо-6-фосфорилаза, в мышцах нет. Этот
- 74. Постоянный уровень глюкозы в крови в постабсорбтивный период = 3,5-5,5 мМ/л Органы мозг и эритроциты потребляют
- 75. Функция мышечного гликогена заключается в высвобождении глюкозо-6фосфата, используемого в самой мышце для окисления и получения энергии.
- 76. Рис.Особенности мобилизации гликогена в печени и мышцах
- 77. Гормоны регулирующие обмен гликогена Переключение процессов синтеза и мобилизации гликогена в печени происходит при переходе состояния
- 78. Рис. Изменение активности гликогенсинтетазы и гликогенфосфорилазы
- 79. Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в Крови. Инсулин и глюкагон
- 80. В период пищеварения преобладающим является влияние инсулина, и инсулин/глюкагоновый индекс в этом случае повышается . Инсулин
- 81. При интенсивной мышечной работе и стрессе в кровь из мозгового вещества надпочечников секретируется гормон адреналин. Он
- 82. Болезни углеводного обмена Причины: нарушения переваривания и всасывания углеводов, синтеза и распада гликогена. - ферментопатии, -
- 83. Нарушения переваривания дисахаридов
- 84. Гликогеновые болезни их причины
- 85. 2. ГЛЮКОНЕОГЕНЕЗ
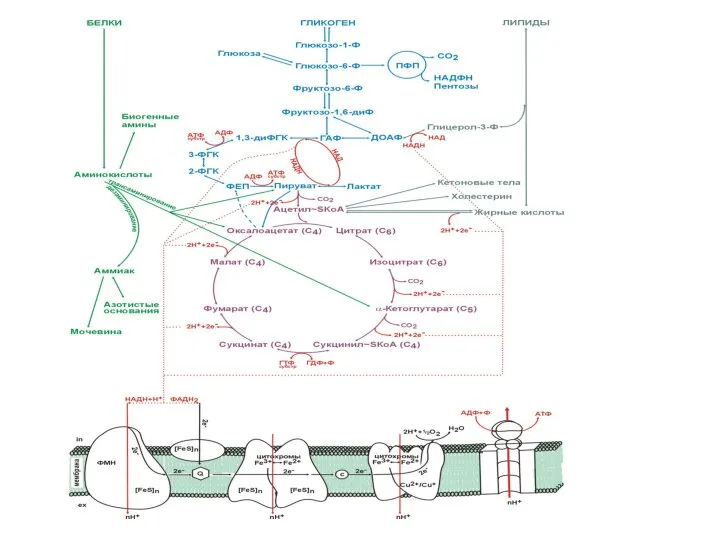
- 86. Карта основных путей метаболизма
- 87. Обмен углеводов
- 88. Глюконеогенез Глюконеогенез – процесс синтеза глюкозы из веществ неуглеводной природы. Субстраты глюконеогенеза: пируват, лактат, глицерол, аминокислоты.
- 89. Процесс протекает главным образом в печени и менее интенсивно в корковом веществе почек и слизистой кишечника.
- 90. Большинство реакций гликолиза и гликонеогенеза являются обратимыми и катализируются одними и теми же ферментами. Четыре реакции
- 91. Схема Гликолиза (слева) и глюконеогенеза (справа) I, II, III – необратимые циклы, катализируются разными ферментами.
- 92. В митохондриях под действием ферментов малатдегидроеназы и аминотрансферазы образуется малат и аспартат из оксалоацетата (ЩУК), которые
- 93. Превращение пирувата (ПВК) в оксалоацетат 1 - транспорт ПВК из цитозоля в митохондрю; 2 – превращение
- 94. Рис.Суммарное уравнение образования глюкозы из пирувата Видно, что синтез глюкозы энергозатратный процесс
- 95. Использование лактата в качестве субстрата в глюконеогенезе связано с транспортом его в печень и превращением в
- 96. Цикл Кори 1 – поступление лактата из сокращающейся мышцы и эритроцитов с током крови в печень;
- 97. Часть ПВК, образовавшегося из лактата, окисляется в печени до СО2 и Н2О. Энергия выделяющаяся при окислении,
- 98. Снижение использования лактата в качестве субстрата в синтезе глюкозы, вызванное дефектом ферментов глюконеогенеза, и может приводить
- 99. Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие: тканевой гипоксии, вызывающей активацию анаэробного
- 100. 3. Пентозофосфатный путь в метаболизме глюкозы (ПФП) Это альтернативный путь окиления глюкозы. К синтезу АТФ этот
- 101. Кроме того, пентозофосфатный путь поставляет клетке пентозофосфаты (пентозы) , необходимые для синтеза нуклеиновых кислот и коферментов
- 102. В пентозофосфатном пути превращения глюкозы можно выделить две части: А – окислительный этап и Б –
- 103. Пентозофосфатный путь превращений глюкозы
- 104. ПФП превращения глюкозы А – окислительный этап; Б – неокислительный этап; В – пентозофосфатний цикл
- 105. Пентозофосфатный путь является альтернативным путем окисления глюкозы. Он включает несколько циклов, в результате функционирования которых из
- 106. 3глюкозо-6-фосфат + 6 НАДФ+ → → 3СО2 + 2глюкозо-6-фосфат + глицеральдегид-3-фосфат + 6 НАДФ-Н2 Пентозофосфатный цикл
- 107. ПФП в эритроцитах ПФП в эритроцитах очень важен, так как восстановленный НАДФН используется для защиты клеток
- 108. Дефект первого фермента ПФП глюкозо-6-фосфат дегидрогеназы приводит к дефициту НАДФН и невозможности восстановления глутатиона. Это приводит
- 109. Рис. Восстановление глутатиона с участием НАДФН
- 110. Благодарю за внимание
- 113. Общие пути катаболизма Окисление продуктов окисления глюкозы гликолизом ПВК в общих путях катаболизма в митохондриях: Пируват
- 114. Карта катаболизма
- 115. Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:
- 116. Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в пируватдегидрогеназный комплекс (ПДК):
- 117. Этот комплекс ферментов работает подобно конвейеру, в котором продукт передается от фермента к ферменту. Такой принцип
- 118. Дигидролипоат-ацетилтрансфераза (2) - второй фермент комплекса. Катализирует перенос ацетильного остатка, соединенного с липоевой кислотой на второй
- 119. Дегидрогеназа дигидролипоевой кислоты (3й фермент) отщепляет водород от липоевой кислоты и переносит его на НАД+. Далее
- 120. Цикл лимонной кислоты Цитратный цикл (цикл Кребса, цикл трикарбоновых кислот (ЦТК) – это система реакций, приводящая
- 121. В ходе одного полного цитратного цикла происходит: 1. полное окисление ацетильного остатка до двух молекул СО2;
- 122. Цикл лимонной кислоты / цикл Кребса
- 123. Энергетика полного окисления глюкозы в клетке 1. Гликолиз (распад глю до 2х ПВК) – 8 АТФ
- 124. Ферменты цитратного цикла 1 – цитратсинтаза, 2 – аконитаза, 3 – изоцитратдегидрогеназа, 4 – a-кетоглутаратдегидрогеназный комплекс,
- 125. Сопряжение общих путей катаболизма с дыхательной цепью В общих путях катаболизма происходит пять реакций дегидрирования: (одна
- 126. Энергетика цитратного цикла и общих путей катаболизма За один оборот цитратного цикла синтезируется 12 молекул АТФ.
- 127. Кроме того, в цитратном цикле происходит одна реакция субстратного фосфорилирования, дающая 1 моль ГТФ ( аналог
- 128. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 129. задание на дом: записать все реакции ПДК и цикла Кребса Благодарю за внимание
- 130. Использование метаболитов ЦТК в синтезе различных соединений
- 131. Метаболизм глюкозы
- 132. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 133. Синтез и распад гликогена
- 134. Синтез гликогена гликогенез
- 136. Скачать презентацию

План лекции 6:
Гипер- и гипогликемия: причины возникновения, механизмы компенсации.
Метаболические и клинические
План лекции 6:
Гипер- и гипогликемия: причины возникновения, механизмы компенсации.
Метаболические и клинические

Гипер- и гипогликемия:
причины возникновения, механизмы компенсации
Концентрация глюкозы в артериальной крови
Гипер- и гипогликемия:
причины возникновения, механизмы компенсации
Концентрация глюкозы в артериальной крови
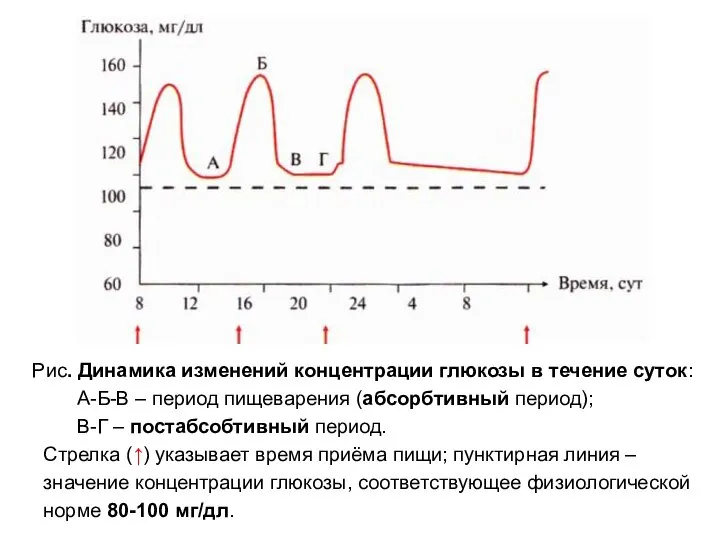
Рис. Динамика изменений концентрации глюкозы в течение суток:
А-Б-В – период
Рис. Динамика изменений концентрации глюкозы в течение суток:
А-Б-В – период

Снижение концентрации глюкозы в крови ниже
80мг/дл или 3,3 мМоль/л
называется гипоглюкоземией.
И
Снижение концентрации глюкозы в крови ниже
80мг/дл или 3,3 мМоль/л
называется гипоглюкоземией.
И

Заболевания вызывающие повышение
концентрации глюкозы в крови
Гипреглюкоземия - повышение сахара в
Заболевания вызывающие повышение
концентрации глюкозы в крови
Гипреглюкоземия - повышение сахара в

Заболевания вызывающие понижение
концентрации глюкозы в крови
Гипогликемия – понижение содержания сахара
Заболевания вызывающие понижение
концентрации глюкозы в крови
Гипогликемия – понижение содержания сахара

Постоянное поступление глюкозы необходимо в качестве
основного источника энергии для нервной ткани
Постоянное поступление глюкозы необходимо в качестве
основного источника энергии для нервной ткани
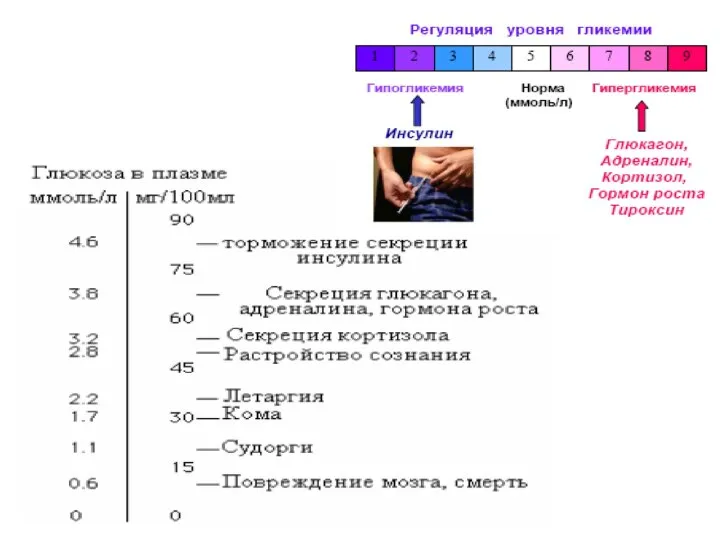
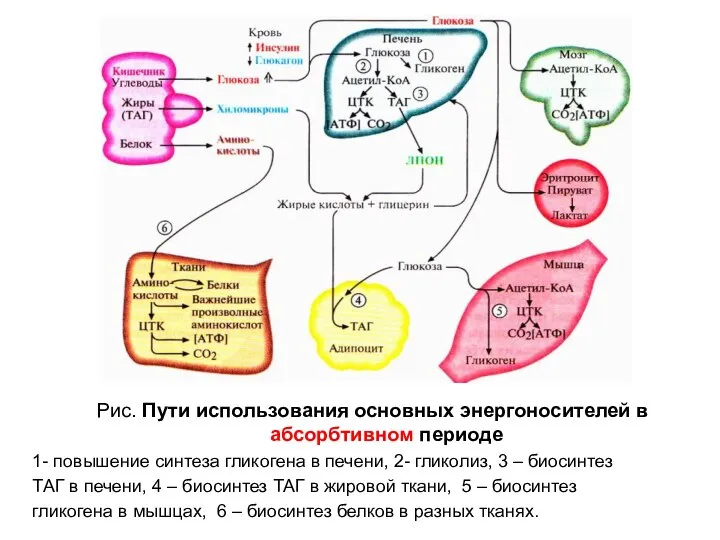
Рис. Пути использования основных энергоносителей в абсорбтивном периоде
1- повышение синтеза гликогена
Рис. Пути использования основных энергоносителей в абсорбтивном периоде
1- повышение синтеза гликогена
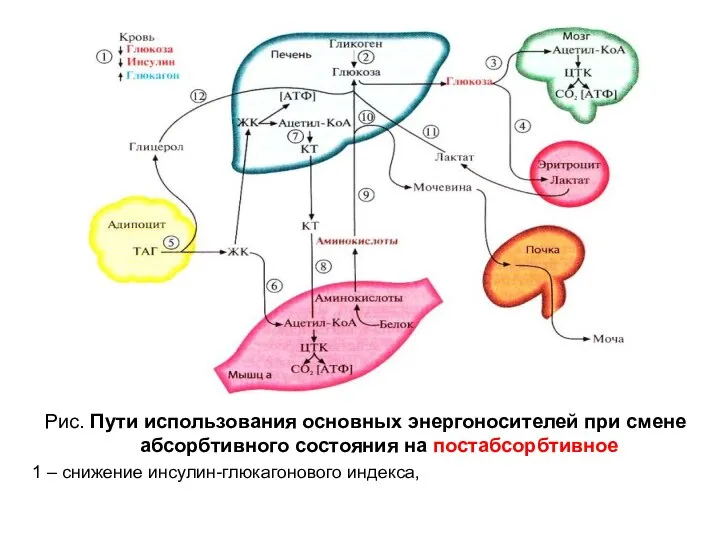
Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное
1
Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное
1
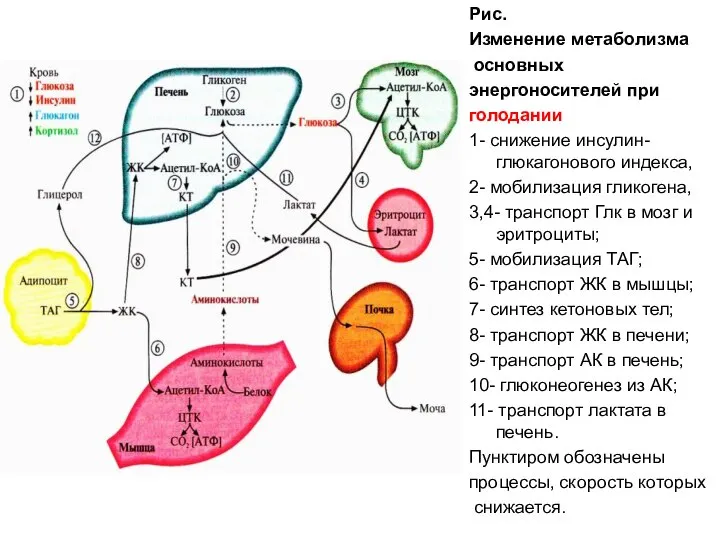
Рис.
Изменение метаболизма
основных
энергоносителей при
голодании
1- снижение инсулин-глюкагонового индекса,
2-
Рис.
Изменение метаболизма
основных
энергоносителей при
голодании
1- снижение инсулин-глюкагонового индекса,
2-

природа гормонов инсулина и глюкагона;
механизм регуляции концентрации глюкозы гормонами.
В зависимости от
природа гормонов инсулина и глюкагона;
механизм регуляции концентрации глюкозы гормонами.
В зависимости от
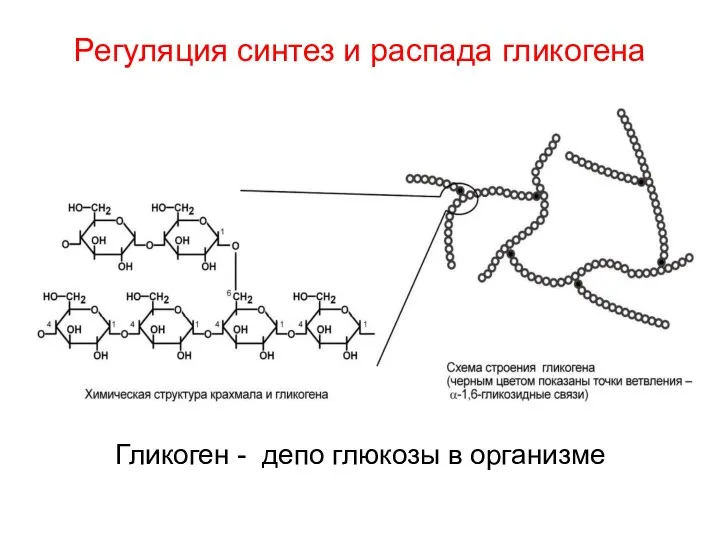
Регуляция синтез и распада гликогена
Гликоген - депо глюкозы в организме
Регуляция синтез и распада гликогена
Гликоген - депо глюкозы в организме

Метаболизм гликогена в печени, мышцах и др.клетках
регулируется несколькими гормонами, одни
Метаболизм гликогена в печени, мышцах и др.клетках
регулируется несколькими гормонами, одни

Присоединение фосфатов к ферменту производят
протеинкиназы, источником фосфора является АТФ:
фосфорилаза
Присоединение фосфатов к ферменту производят
протеинкиназы, источником фосфора является АТФ:
фосфорилаза

Например:
во время мышечной работы
адреналин вызывает фосфорилирование
внутримышечных ферментов обмена гликогена.
Например:
во время мышечной работы
адреналин вызывает фосфорилирование
внутримышечных ферментов обмена гликогена.

Способы активации синтазы гликогена
Гликогенсинтаза при присоединении фосфата прекращает
работу, т.е. она активна
Способы активации синтазы гликогена
Гликогенсинтаза при присоединении фосфата прекращает
работу, т.е. она активна
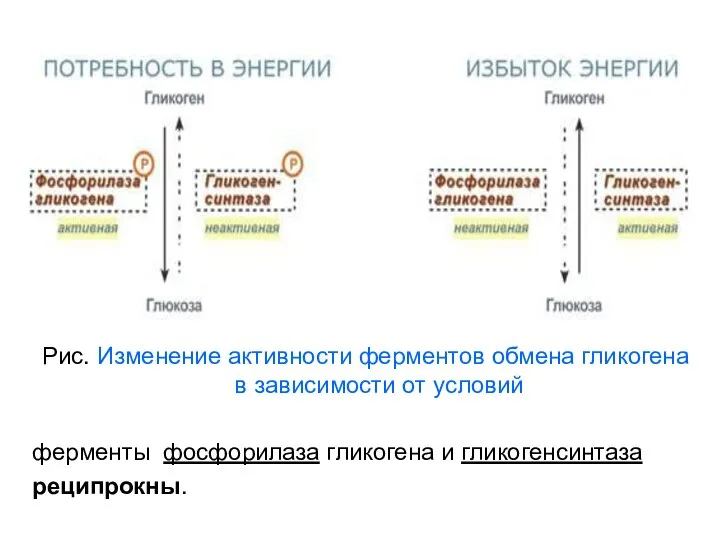
Рис. Изменение активности ферментов обмена гликогена в зависимости от условий
ферменты
Рис. Изменение активности ферментов обмена гликогена в зависимости от условий
ферменты

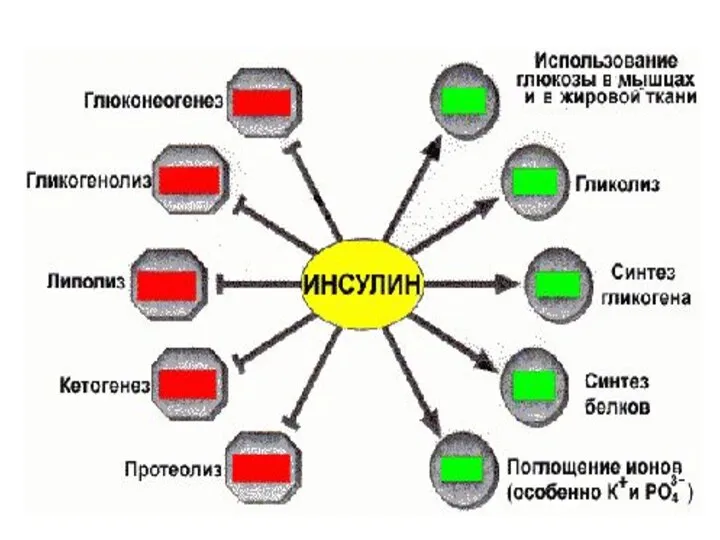
Инсулин
Вырабатывается Бета клетками островков Лангерганса
панкреатической (поджелудочной) железой.
Состоит из 51 остатка АК
мол
Инсулин
Вырабатывается Бета клетками островков Лангерганса
панкреатической (поджелудочной) железой.
Состоит из 51 остатка АК
мол
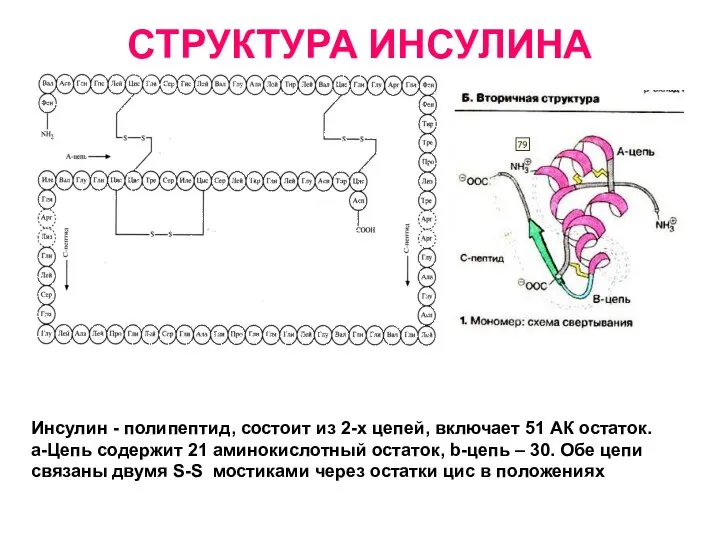
СТРУКТУРА ИНСУЛИНА
Инсулин - полипептид, состоит из 2-х цепей, включает 51 АК
СТРУКТУРА ИНСУЛИНА
Инсулин - полипептид, состоит из 2-х цепей, включает 51 АК
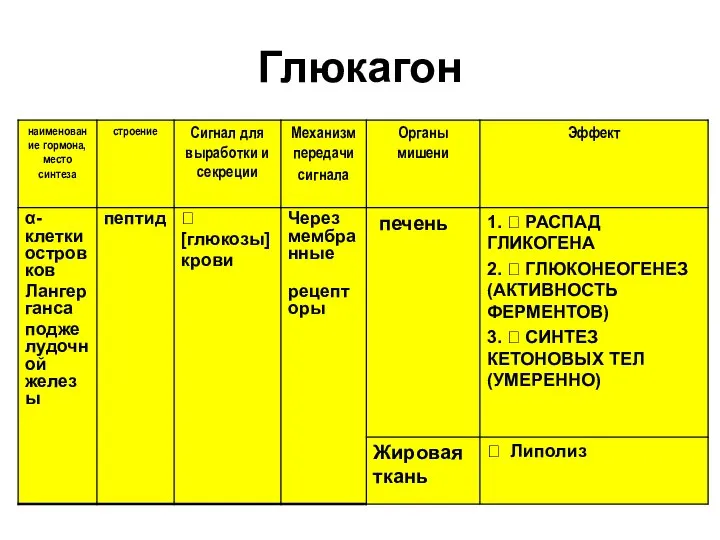
Глюкагон
Глюкагон

адреналин
адреналин

Изменение гормонального статуса и метаболизма при сахарном диабете
Сахарный диабет (СД) возникает
Изменение гормонального статуса и метаболизма при сахарном диабете
Сахарный диабет (СД) возникает

ИЗСД - СД I типа
Является следствием разрушения бета-клеток островков
Лангерганса в результате
ИЗСД - СД I типа
Является следствием разрушения бета-клеток островков
Лангерганса в результате

ИНСД – СД II типа
ПРИЧИНЫ
Развивается вследствие нарушения:
превращения проинсулина в инсулин,
регуляции секреции
ИНСД – СД II типа
ПРИЧИНЫ
Развивается вследствие нарушения:
превращения проинсулина в инсулин,
регуляции секреции

Факторы увеличивающие риск развития ИНСД:
ожирение,
неправильный режим питания,
гиподинамия,
стрессы.
Обычно после 40 лет.
Симптомы
Факторы увеличивающие риск развития ИНСД:
ожирение,
неправильный режим питания,
гиподинамия,
стрессы.
Обычно после 40 лет.
Симптомы

Изменения метаболизма при СД
1. соотношения инсулин-глюкагон (инсулин/глюкагон) снижено.
При этом ослабевает стимуляция
Изменения метаболизма при СД
1. соотношения инсулин-глюкагон (инсулин/глюкагон) снижено.
При этом ослабевает стимуляция
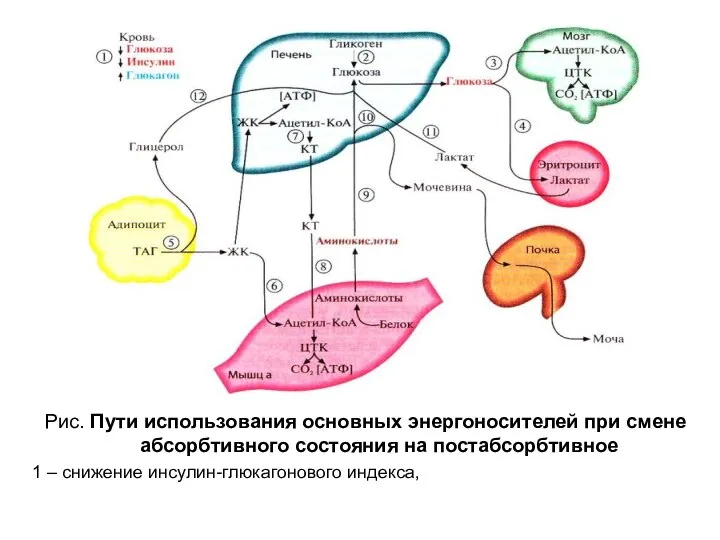
Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное
1
Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное
1
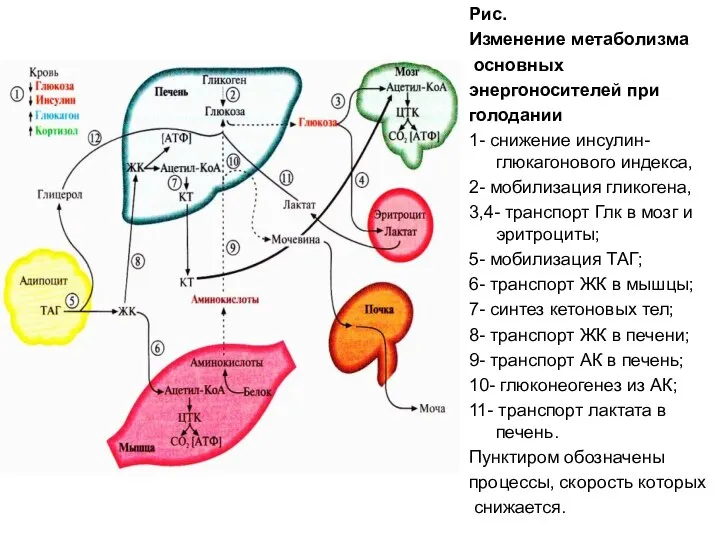
Рис.
Изменение метаболизма
основных
энергоносителей при
голодании
1- снижение инсулин-глюкагонового индекса,
2-
Рис.
Изменение метаболизма
основных
энергоносителей при
голодании
1- снижение инсулин-глюкагонового индекса,
2-

Симптомы СД
Гиперглюкоземия
Кетонемия
Гиперлипопротеинемия
Азотемия
Полиурия
Симптомы СД
Гиперглюкоземия
Кетонемия
Гиперлипопротеинемия
Азотемия
Полиурия

Симптомы СД
1. Гиперглюкоземия
Для всех форм диабета характерно повышение
концентрации глюкозы в
Симптомы СД
1. Гиперглюкоземия
Для всех форм диабета характерно повышение
концентрации глюкозы в
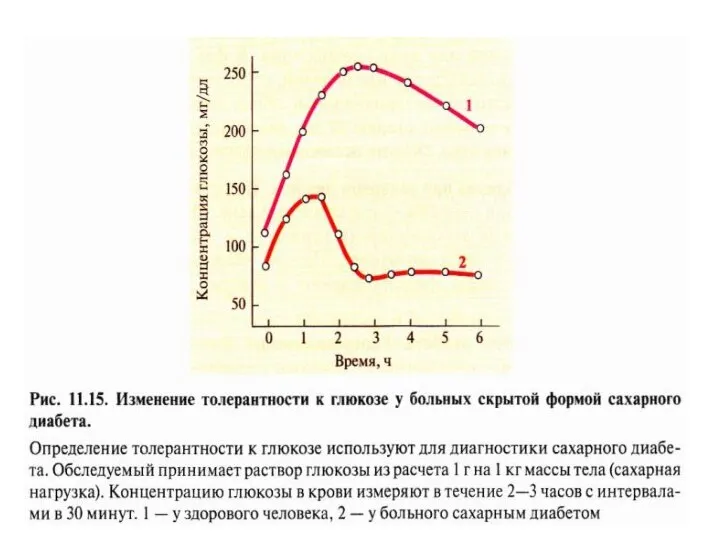

2. Кетонемия
характерный признак СД.
При низком соотношении инсулин-глюкагонового индекса
жиры не депонируются, ускоряется
2. Кетонемия
характерный признак СД.
При низком соотношении инсулин-глюкагонового индекса
жиры не депонируются, ускоряется

3. Гиперлипопротеинемия
Пищевые жиры не депонируются в жировой ткани в
следствие ослабления
3. Гиперлипопротеинемия
Пищевые жиры не депонируются в жировой ткани в
следствие ослабления

5. Полиурия
Для выведения большого количества глюкозы, кетоновых
тел, мочевины требуется большой объем
5. Полиурия
Для выведения большого количества глюкозы, кетоновых
тел, мочевины требуется большой объем

Острые осложнения СД
Механизм развития диабетической комы.
Нарушения метаболизма углеводов, жиров и белков
Острые осложнения СД
Механизм развития диабетической комы.
Нарушения метаболизма углеводов, жиров и белков

Коматозные состояния при СД
кетоацидотическая кома,
гиперосмолярная кома,
лактоацидотическая кома.
Для кетоацидотической формы характены выраженный
дефицит
Коматозные состояния при СД
кетоацидотическая кома,
гиперосмолярная кома,
лактоацидотическая кома.
Для кетоацидотической формы характены выраженный
дефицит
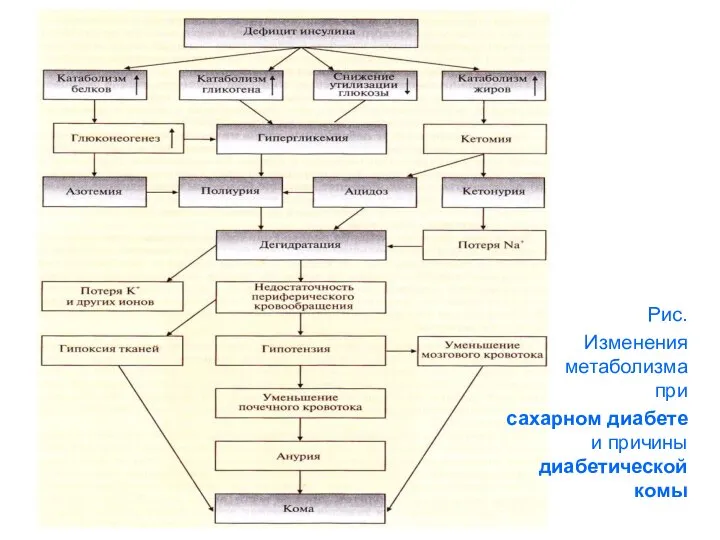
Рис.
Изменения метаболизма при
сахарном диабете и причины диабетической комы
Рис.
Изменения метаболизма при
сахарном диабете и причины диабетической комы

Поздние осложнения СД
Причина - длительная гиперглюкоземия.
Следствие – инвалидизация.
Из-за повреждения кровеносных
Поздние осложнения СД
Причина - длительная гиперглюкоземия.
Следствие – инвалидизация.
Из-за повреждения кровеносных

Степень гликозилирования белков зависит от скорости
их обновления.
Также гликозилируются:
- белки базальных
Степень гликозилирования белков зависит от скорости
их обновления.
Также гликозилируются:
- белки базальных

Для сохранения осмомолярности начинается
компенсаторное перемещение жидкости из клеток и
внеклеточного
Для сохранения осмомолярности начинается
компенсаторное перемещение жидкости из клеток и
внеклеточного

Диабетические ангиопатии
Макроангиопатии:
снижение эластичности артерий, повреждение крупных и
мелких сосудов мозга, сердца,
Макроангиопатии:
снижение эластичности артерий, повреждение крупных и
мелких сосудов мозга, сердца,

Благодарю за внимание
Благодарю за внимание
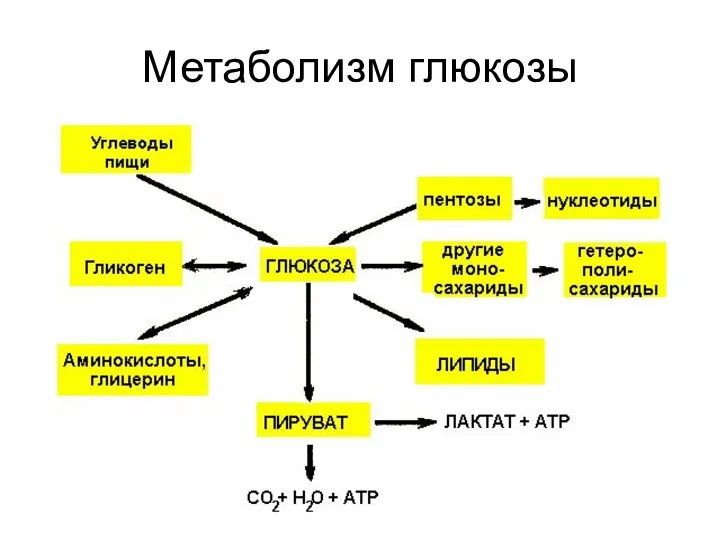
Метаболизм глюкозы
Метаболизм глюкозы
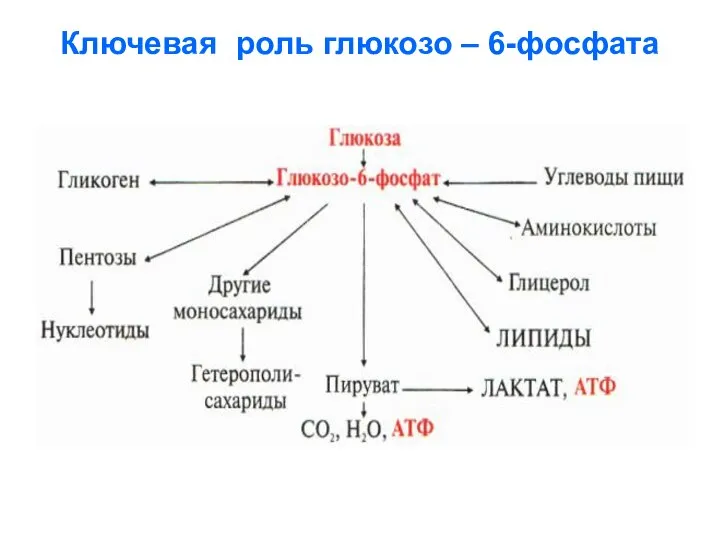
Ключевая роль глюкозо – 6-фосфата
Ключевая роль глюкозо – 6-фосфата
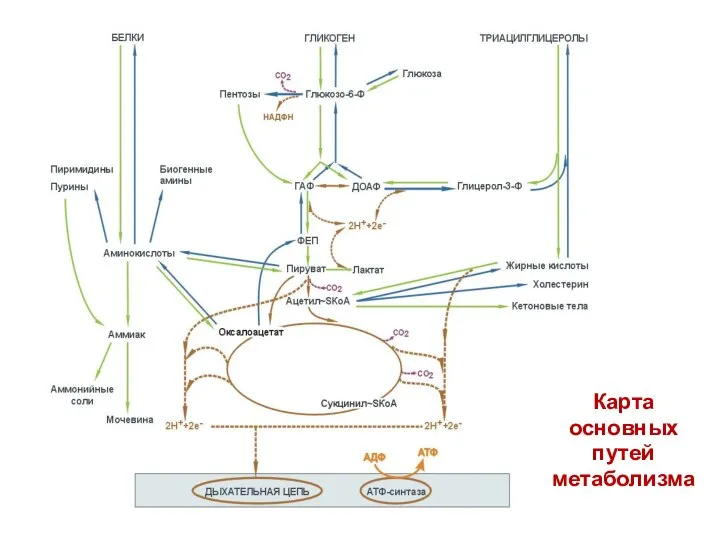
Карта
основных
путей
метаболизма
Карта
основных
путей
метаболизма
Обмен углеводов
Обмен углеводов

1. Обмен гликогена
Гликогенез и
гликогенолиз
1. Обмен гликогена
Гликогенез и
гликогенолиз

Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид).
Гликоген – основная форма депонирования глюкозы в клетках животных (резервный полисахарид).

В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение
В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое освобождение

Линейные участки молекулы гликогена образованы
остатками глюкозы, связанными
альфа-1,4-гликозидными связями, а
Линейные участки молекулы гликогена образованы
остатками глюкозы, связанными
альфа-1,4-гликозидными связями, а

Синтез и распад гликогена
Синтез и распад гликогена гормонозависимый процессы и
Синтез и распад гликогена
Синтез и распад гликогена гормонозависимый процессы и

Синтез гликогена - гликогенез
Синтез гликогена ускоряется в период
пищеварения (абсорбтивный период –
Синтез гликогена - гликогенез
Синтез гликогена ускоряется в период
пищеварения (абсорбтивный период –

Когда длина синтезируемой цепи увеличивается на
11-12 остатков глюкозы, фермент ветвления
Когда длина синтезируемой цепи увеличивается на
11-12 остатков глюкозы, фермент ветвления
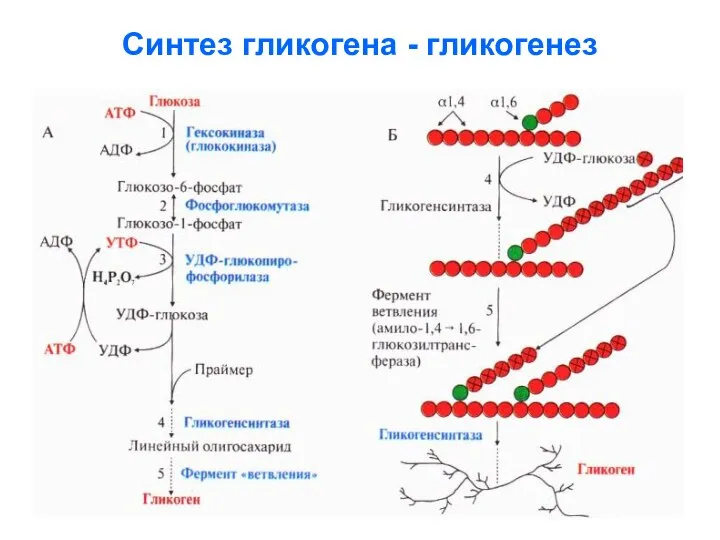
Синтез гликогена - гликогенез
Синтез гликогена - гликогенез
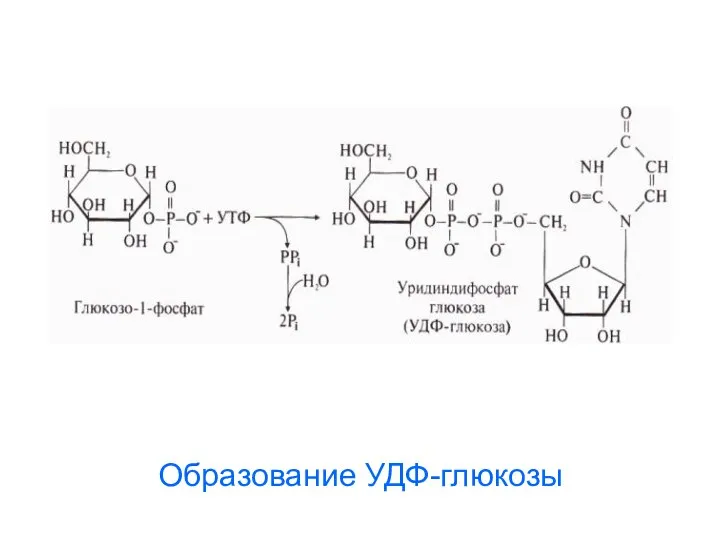
Образование УДФ-глюкозы
Образование УДФ-глюкозы

Далее подробно расписаны реакции гликогенеза:
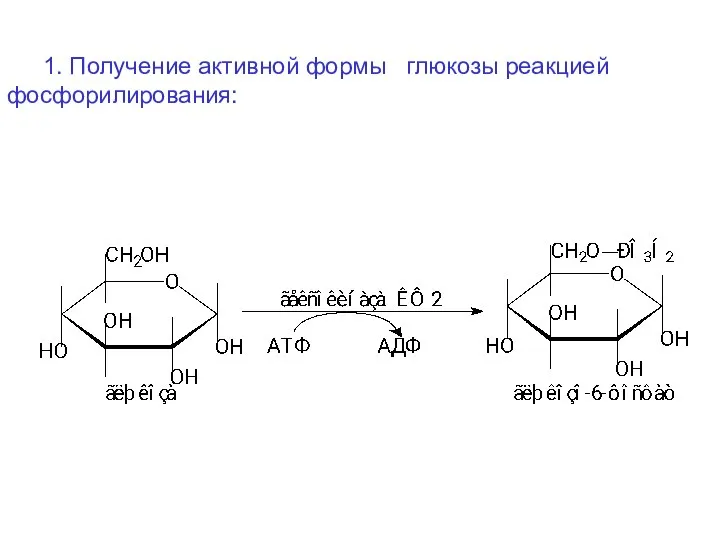
1. Получение активной формы глюкозы реакцией фосфорилирования:
1. Получение активной формы глюкозы реакцией фосфорилирования:
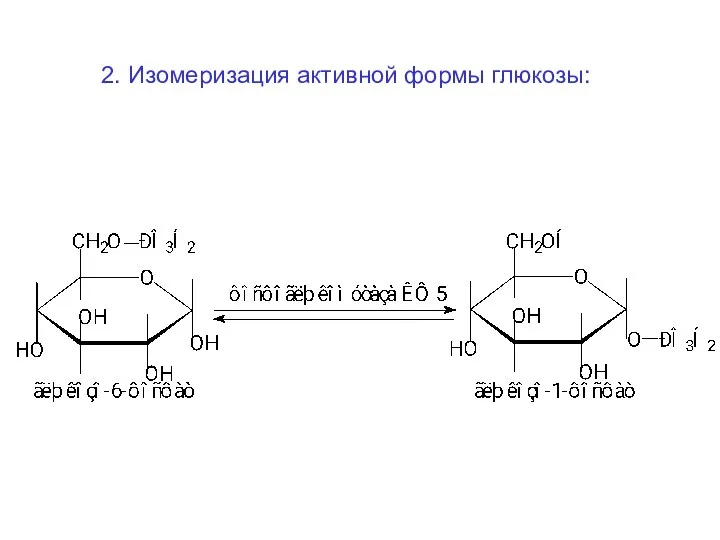
2. Изомеризация активной формы глюкозы:
2. Изомеризация активной формы глюкозы:
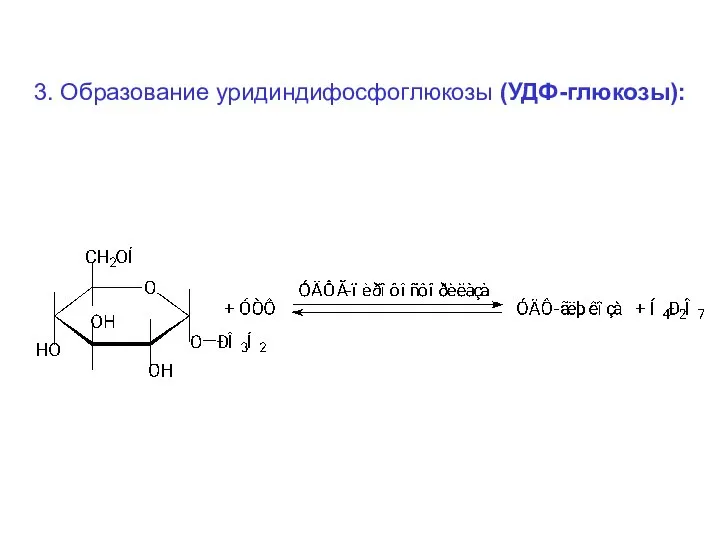
3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
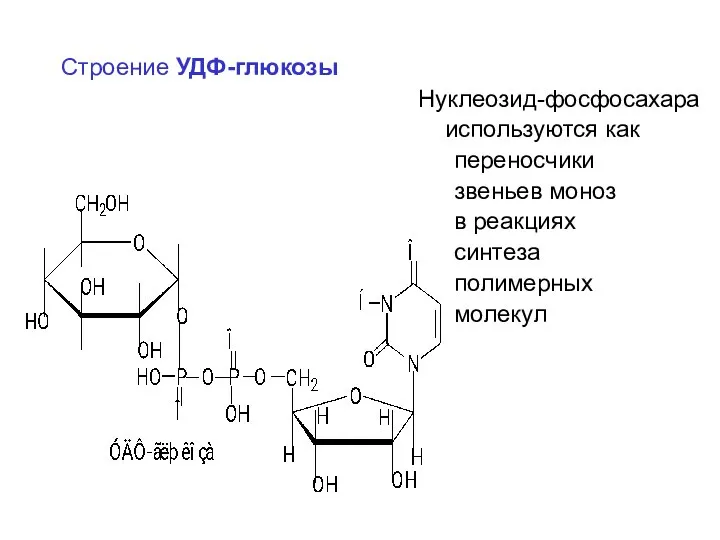
Строение УДФ-глюкозы
Нуклеозид-фосфосахара
используются как
переносчики
звеньев моноз
в реакциях
синтеза
полимерных
молекул
Строение УДФ-глюкозы
Нуклеозид-фосфосахара
используются как
переносчики
звеньев моноз
в реакциях
синтеза
полимерных
молекул

4. Перенос остатка глюкозы с УДФ-глюкозы на
затравочную цепь гликогена, которая
4. Перенос остатка глюкозы с УДФ-глюкозы на затравочную цепь гликогена, которая

Ветвление гликогена
5. Синтез α-(1,6)-гликозидных связей происходит при
участии гликоген-ветвящегося фермента –
гликоген-трансферазы, которая
Ветвление гликогена
5. Синтез α-(1,6)-гликозидных связей происходит при
участии гликоген-ветвящегося фермента –
гликоген-трансферазы, которая

Подводим итог:
Гликоген синтезируется в период пищеварения
(в течение 1-2 часов после
Подводим итог:
Гликоген синтезируется в период пищеварения
(в течение 1-2 часов после

Для подготовки и включения глюкозы в растущую
полисахаридную цепь требуется энергия
Для подготовки и включения глюкозы в растущую
полисахаридную цепь требуется энергия

Необходимость превращения глюкозы в гликоген
связана с тем, что накопление значительного
Необходимость превращения глюкозы в гликоген
связана с тем, что накопление значительного

Распад гликогена - гликогенолиз
или мобилизация гликогена – происходит в основном
в период
Распад гликогена - гликогенолиз
или мобилизация гликогена – происходит в основном
в период
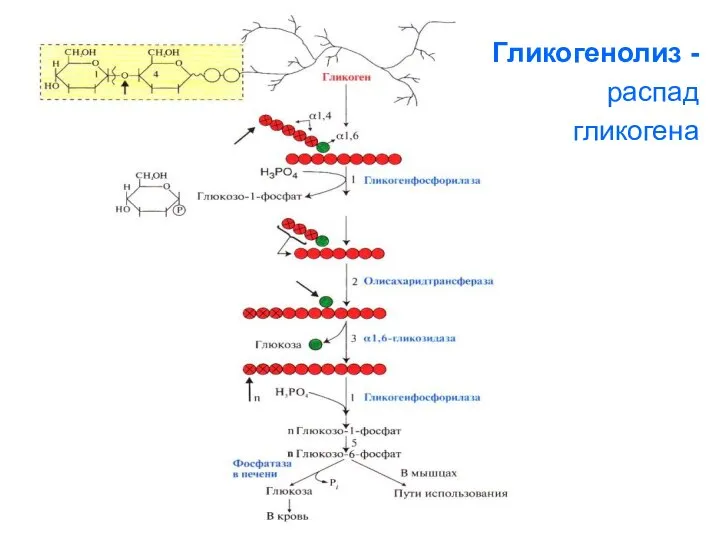
Гликогенолиз -
распад
гликогена
Гликогенолиз -
распад
гликогена

1. Освобождение глюкозы в виде глюкозо-1-фосфата
(реакция 5) происходит в результате
1. Освобождение глюкозы в виде глюкозо-1-фосфата
(реакция 5) происходит в результате
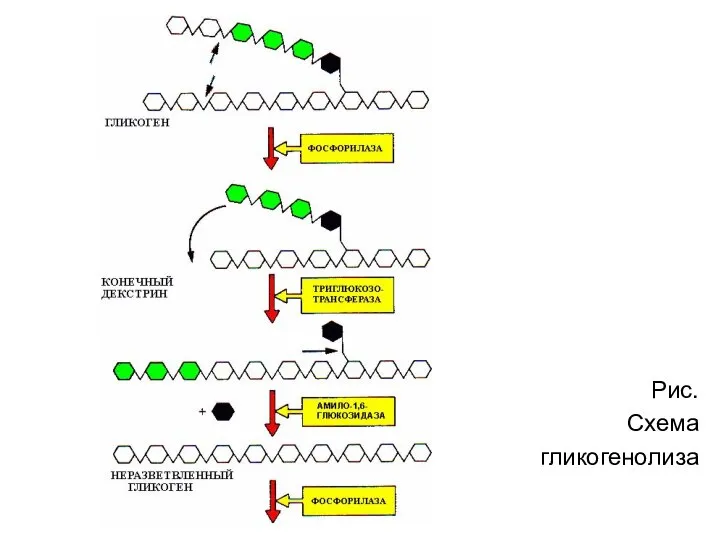
Рис.
Схема
гликогенолиза
Схема
гликогенолиза

Мобилизация гликогена в печени и мыщцах отличаются
В печени есть фермент
Мобилизация гликогена в печени и мыщцах отличаются
В печени есть фермент

Постоянный уровень глюкозы в крови в постабсорбтивный
период = 3,5-5,5 мМ/л
Органы
период = 3,5-5,5 мМ/л
Органы

Функция мышечного гликогена заключается в
высвобождении глюкозо-6фосфата, используемого в
самой мышце
Функция мышечного гликогена заключается в
высвобождении глюкозо-6фосфата, используемого в
самой мышце

Рис.Особенности мобилизации гликогена в печени и мышцах
Рис.Особенности мобилизации гликогена в печени и мышцах

Гормоны регулирующие обмен гликогена
Переключение процессов синтеза и мобилизации
гликогена в печени
Гормоны регулирующие обмен гликогена
Переключение процессов синтеза и мобилизации
гликогена в печени
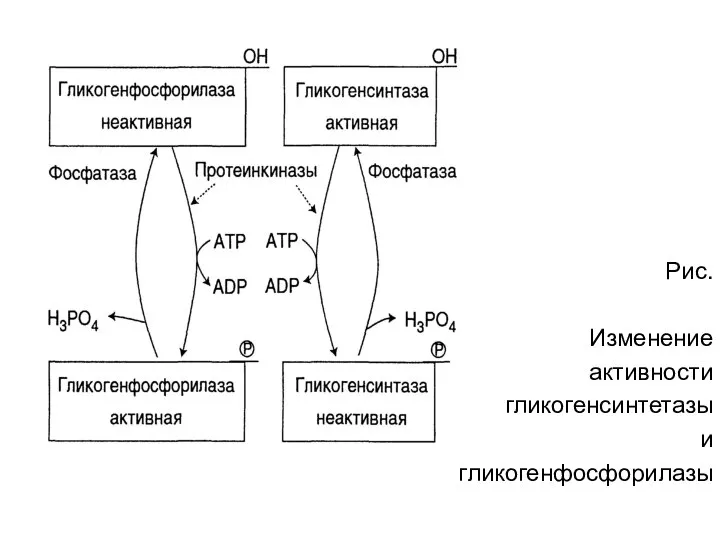
Рис.
Изменение
активности
гликогенсинтетазы
и
гликогенфосфорилазы
Рис.
Изменение
активности
гликогенсинтетазы
и
гликогенфосфорилазы

Первичным сигналом для синтеза инсулина и
глюкагона является изменение концентрации глюкозы
Первичным сигналом для синтеза инсулина и
глюкагона является изменение концентрации глюкозы

В период пищеварения преобладающим является
влияние инсулина, и инсулин/глюкагоновый индекс в
В период пищеварения преобладающим является
влияние инсулина, и инсулин/глюкагоновый индекс в

При интенсивной мышечной работе и стрессе в кровь
из мозгового вещества
При интенсивной мышечной работе и стрессе в кровь
из мозгового вещества

Болезни углеводного обмена
Причины:
нарушения переваривания и всасывания углеводов,
синтеза и распада гликогена.
- ферментопатии,
-
Болезни углеводного обмена
Причины:
нарушения переваривания и всасывания углеводов,
синтеза и распада гликогена.
- ферментопатии,
-
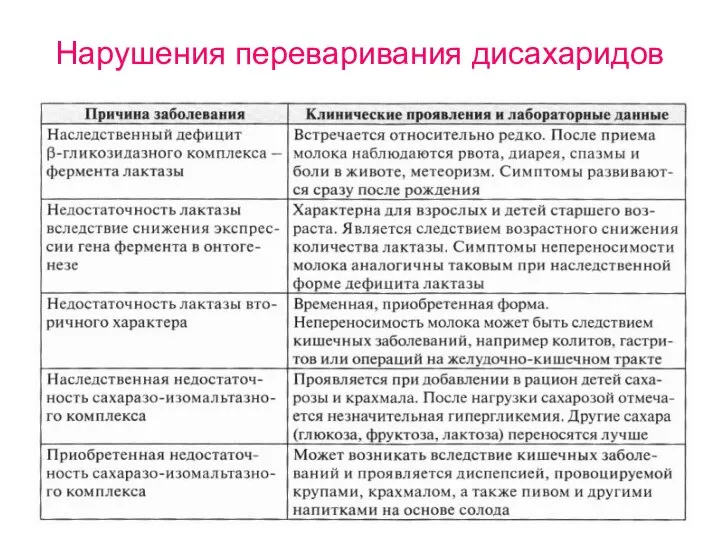
Нарушения переваривания дисахаридов
Нарушения переваривания дисахаридов
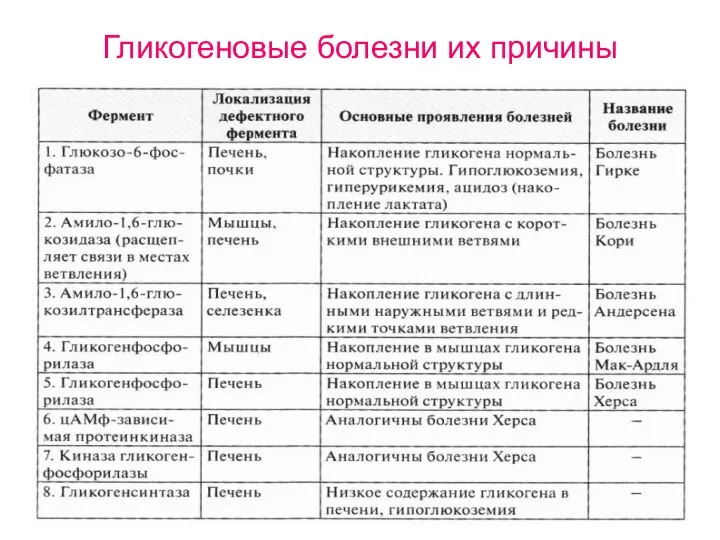
Гликогеновые болезни их причины
Гликогеновые болезни их причины

2. ГЛЮКОНЕОГЕНЕЗ
2. ГЛЮКОНЕОГЕНЕЗ
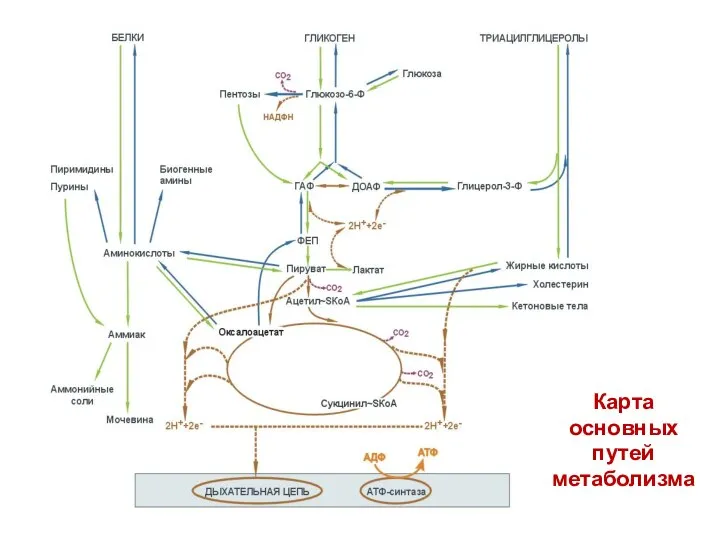
Карта
основных
путей
метаболизма
Карта
основных
путей
метаболизма
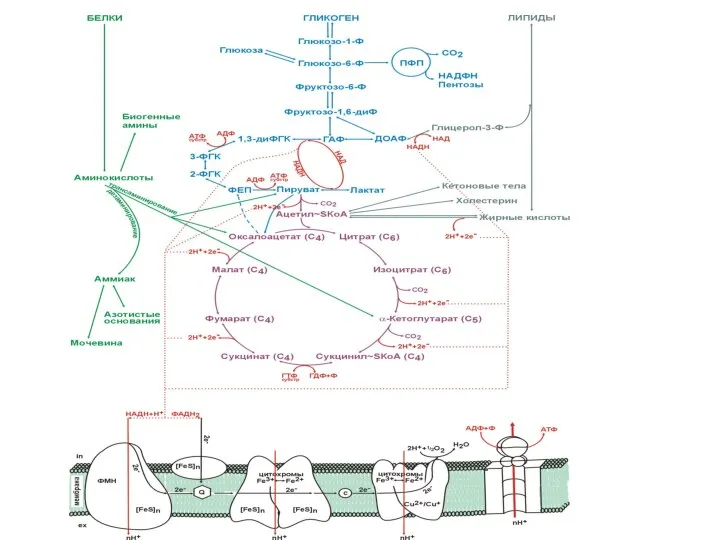
Обмен углеводов
Обмен углеводов

Глюконеогенез
Глюконеогенез – процесс синтеза глюкозы из веществ
неуглеводной природы.
Субстраты глюконеогенеза: пируват, лактат,
глицерол, аминокислоты.
Важнейшей
Глюконеогенез
Глюконеогенез – процесс синтеза глюкозы из веществ
неуглеводной природы.
Субстраты глюконеогенеза: пируват, лактат,
глицерол, аминокислоты.
Важнейшей

Процесс протекает главным образом в печени и менее
интенсивно в корковом веществе
Процесс протекает главным образом в печени и менее
интенсивно в корковом веществе

Большинство реакций гликолиза и гликонеогенеза
являются обратимыми и катализируются одними и
Большинство реакций гликолиза и гликонеогенеза
являются обратимыми и катализируются одними и
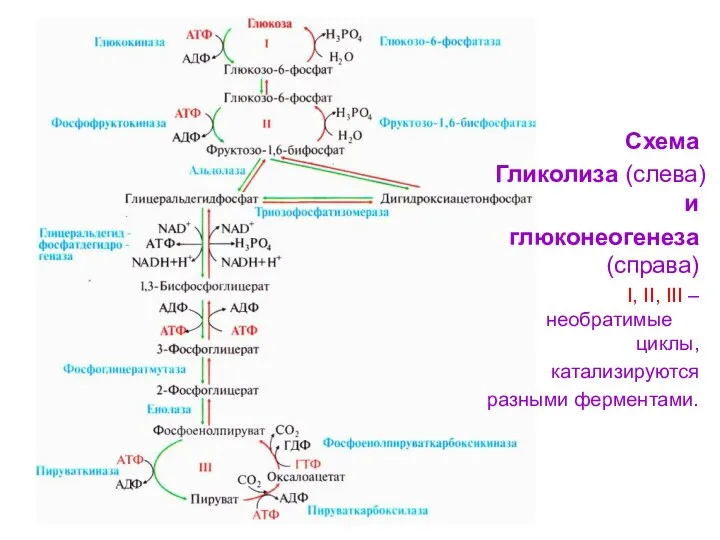
Схема
Гликолиза (слева) и
глюконеогенеза (справа)
I, II, III – необратимые циклы,
катализируются
разными
Схема
Гликолиза (слева) и
глюконеогенеза (справа)
I, II, III – необратимые циклы,
катализируются
разными

В митохондриях под действием ферментов
малатдегидроеназы и аминотрансферазы образуется
малат и
В митохондриях под действием ферментов
малатдегидроеназы и аминотрансферазы образуется
малат и
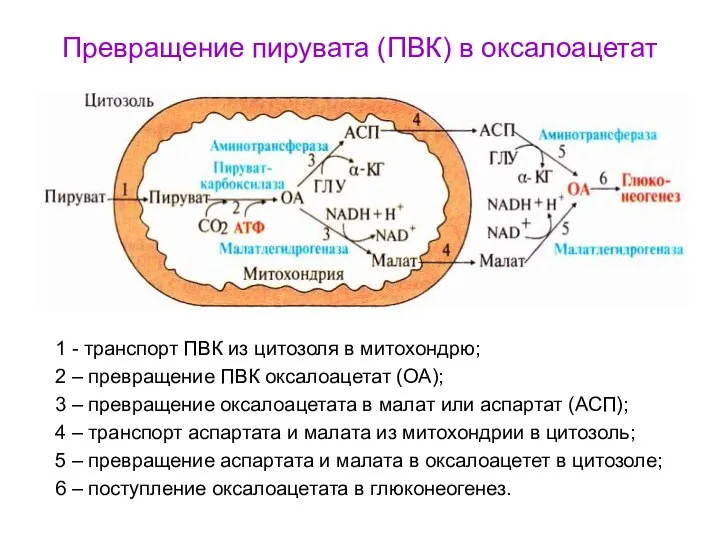
Превращение пирувата (ПВК) в оксалоацетат
1 - транспорт ПВК из цитозоля в
Превращение пирувата (ПВК) в оксалоацетат
1 - транспорт ПВК из цитозоля в

Рис.Суммарное уравнение образования глюкозы из пирувата
Видно, что синтез глюкозы энергозатратный процесс
Рис.Суммарное уравнение образования глюкозы из пирувата
Видно, что синтез глюкозы энергозатратный процесс

Использование лактата в качестве субстрата в
глюконеогенезе связано с транспортом его в
Использование лактата в качестве субстрата в
глюконеогенезе связано с транспортом его в
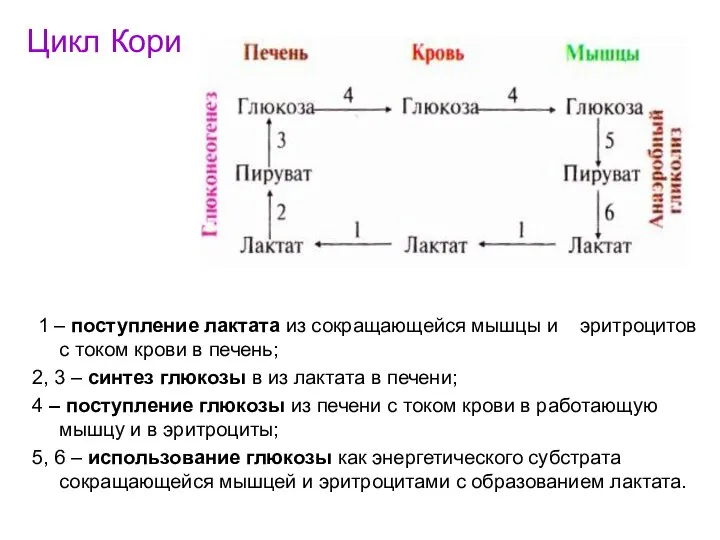
Цикл Кори
1 – поступление лактата из сокращающейся мышцы и эритроцитов
Цикл Кори
1 – поступление лактата из сокращающейся мышцы и эритроцитов

Часть ПВК, образовавшегося из лактата, окисляется в
печени до СО2 и
Часть ПВК, образовавшегося из лактата, окисляется в
печени до СО2 и

Снижение использования лактата в качестве субстрата
в синтезе глюкозы, вызванное дефектом
Снижение использования лактата в качестве субстрата
в синтезе глюкозы, вызванное дефектом

Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие:
тканевой
Причиной повышения лактата в крови может быть нарушение метаболизма пирувата вследствие:
тканевой

3. Пентозофосфатный путь
в метаболизме глюкозы (ПФП)
Это альтернативный путь окиления глюкозы.
3. Пентозофосфатный путь
в метаболизме глюкозы (ПФП)
Это альтернативный путь окиления глюкозы.

Кроме того, пентозофосфатный путь поставляет клетке
пентозофосфаты (пентозы) ,
необходимые для
Кроме того, пентозофосфатный путь поставляет клетке
пентозофосфаты (пентозы) ,
необходимые для

В пентозофосфатном пути превращения глюкозы можно
выделить две части:
А –
В пентозофосфатном пути превращения глюкозы можно
выделить две части:
А –
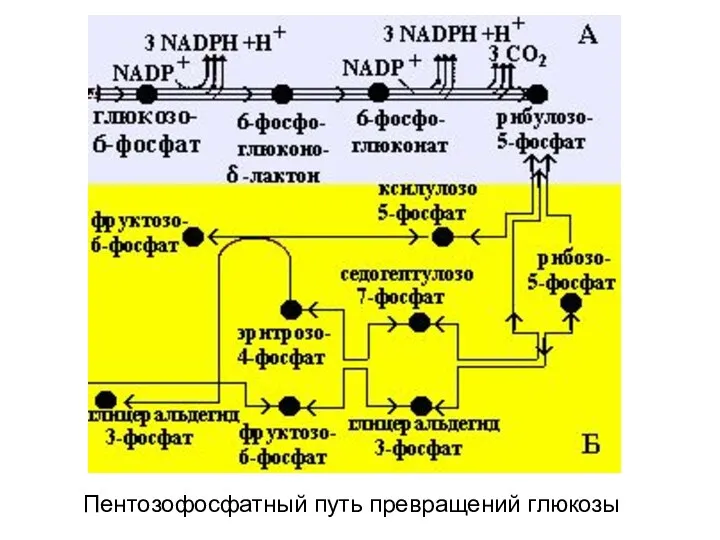
Пентозофосфатный путь превращений глюкозы
Пентозофосфатный путь превращений глюкозы
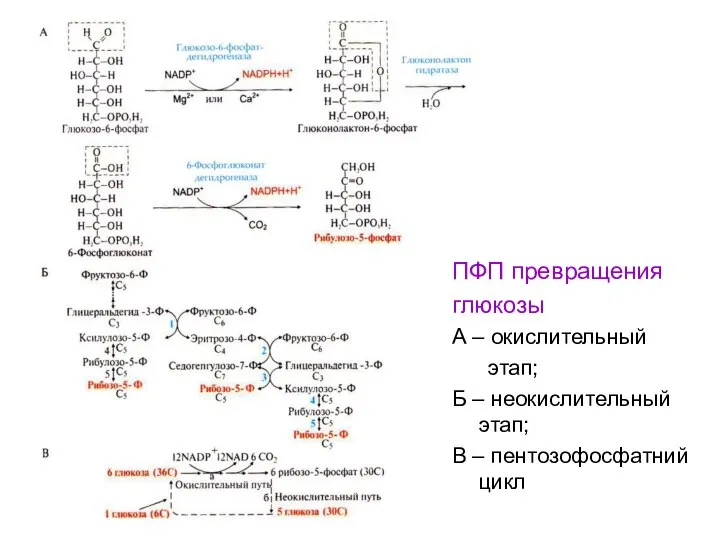
ПФП превращения
глюкозы
А – окислительный
этап;
Б – неокислительный этап;
В – пентозофосфатний цикл
ПФП превращения
глюкозы
А – окислительный
этап;
Б – неокислительный этап;
В – пентозофосфатний цикл

Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает несколько
Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает несколько

3глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат +
3глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат +

ПФП в эритроцитах
ПФП в эритроцитах очень важен, так как
восстановленный НАДФН
ПФП в эритроцитах
ПФП в эритроцитах очень важен, так как
восстановленный НАДФН

Дефект первого фермента ПФП
глюкозо-6-фосфат дегидрогеназы приводит к
дефициту НАДФН и невозможности
Дефект первого фермента ПФП
глюкозо-6-фосфат дегидрогеназы приводит к
дефициту НАДФН и невозможности
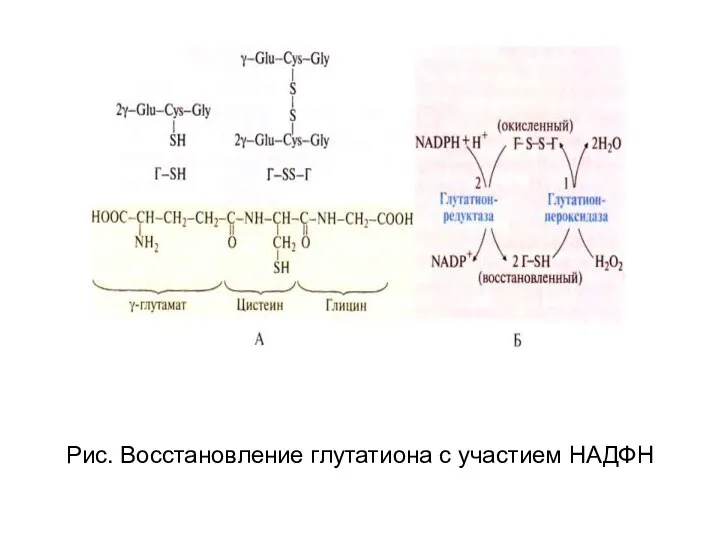
Рис. Восстановление глутатиона с участием НАДФН
Рис. Восстановление глутатиона с участием НАДФН

Благодарю за внимание



Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих путях
Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих путях
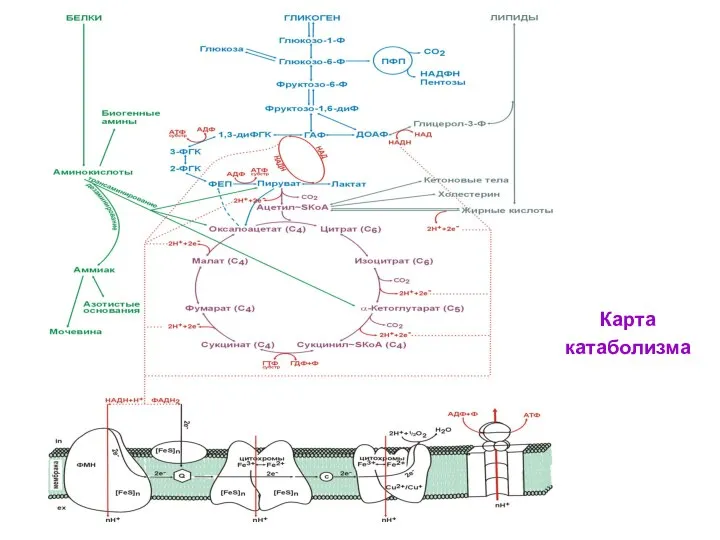
Карта
катаболизма
Карта
катаболизма

Окислительное декарбоксилирование пировиноградной кислоты
Суммарный результат многостадийной
реакции выглядит следующим образом:
Окислительное декарбоксилирование пировиноградной кислоты
Суммарный результат многостадийной
реакции выглядит следующим образом:
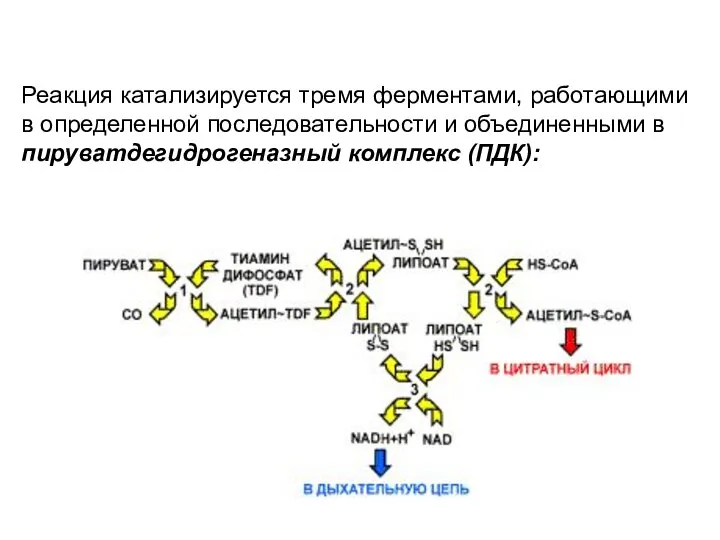
Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в
Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в

Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается от
Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается от

Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,
Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,

Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты и
Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты и

Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК) –
это
Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК) –
это

В ходе одного полного цитратного цикла происходит:
1. полное окисление ацетильного
В ходе одного полного цитратного цикла происходит:
1. полное окисление ацетильного
Цикл лимонной кислоты / цикл Кребса
Цикл лимонной кислоты / цикл Кребса

Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до 2х
Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до 2х

Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4 –
Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4 –

Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма происходит
Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма происходит

Энергетика цитратного цикла и общих путей катаболизма
За один оборот цитратного
Энергетика цитратного цикла и общих путей катаболизма
За один оборот цитратного

Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая
Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая
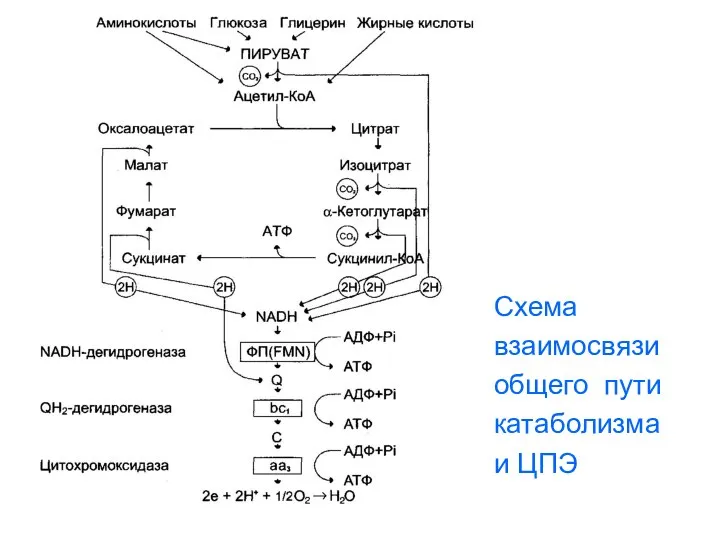
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ

задание на дом:
записать все реакции ПДК и цикла Кребса
Благодарю за внимание
задание на дом:
записать все реакции ПДК и цикла Кребса
Благодарю за внимание
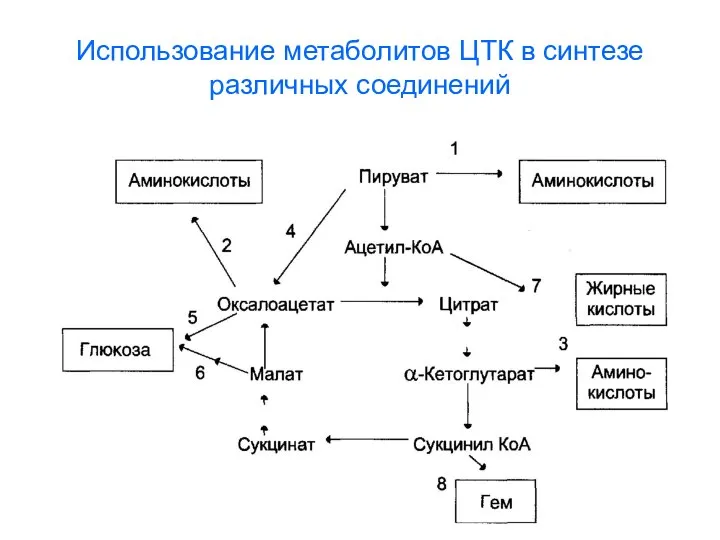
Использование метаболитов ЦТК в синтезе различных соединений
Использование метаболитов ЦТК в синтезе различных соединений
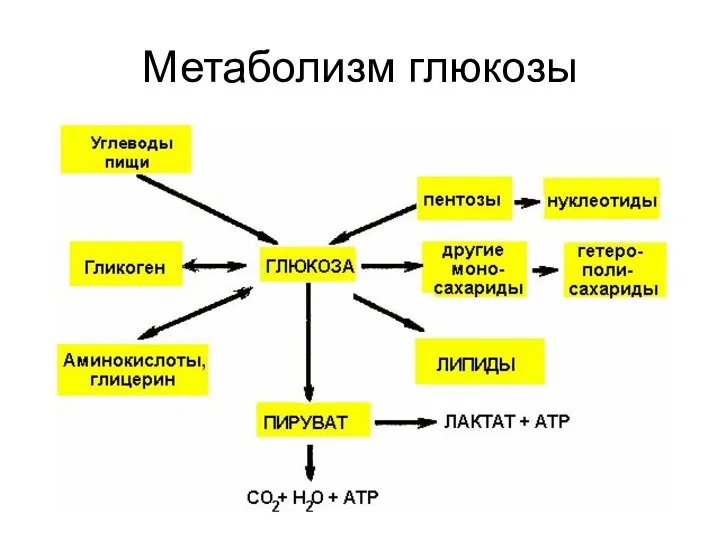
Метаболизм глюкозы
Метаболизм глюкозы
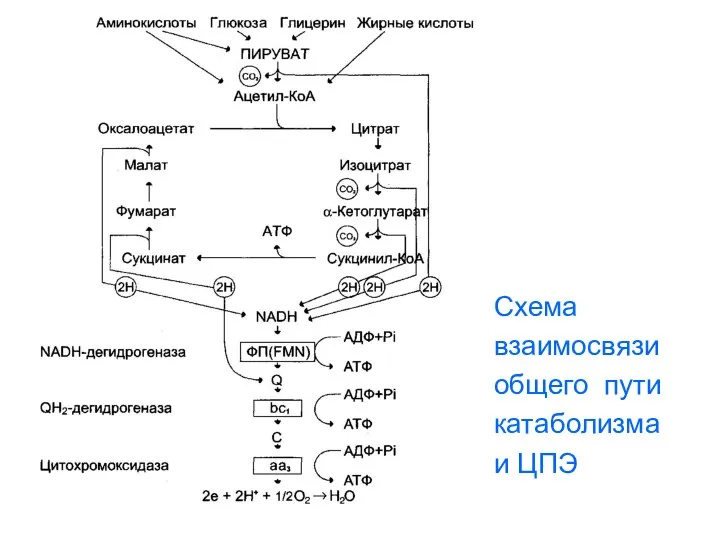
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
Схема
взаимосвязи
общего пути
катаболизма
и ЦПЭ
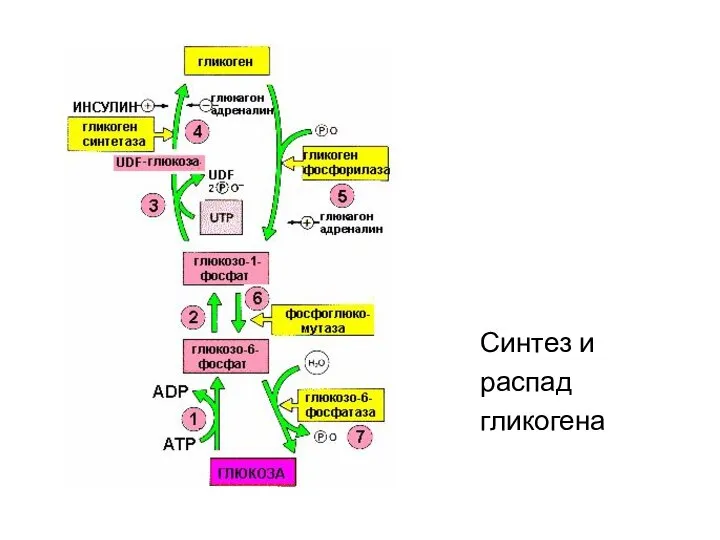
Синтез и
распад
гликогена
Синтез и
распад
гликогена
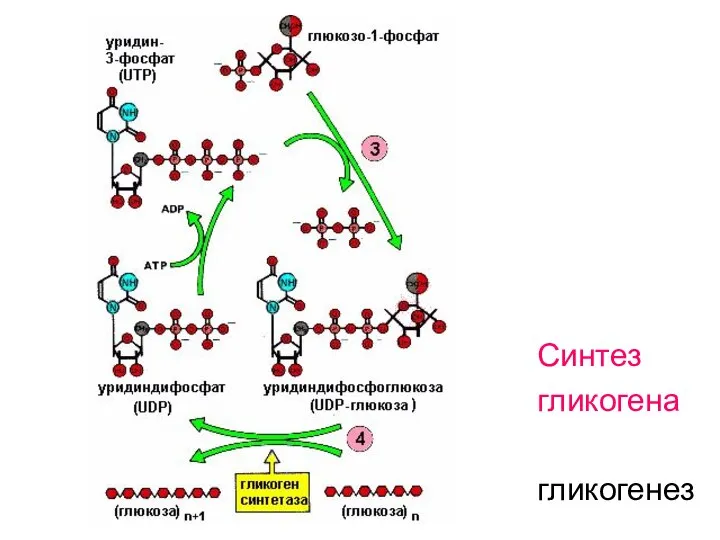
Синтез
гликогена
гликогенез
Синтез
гликогена
гликогенез
 Международная торговля Учитель географии и экономики МОУ «Кингисеппская гимназия» Демянчук Наталья Николаевна
Международная торговля Учитель географии и экономики МОУ «Кингисеппская гимназия» Демянчук Наталья Николаевна  Презентация Тендер на строительство детской площадки
Презентация Тендер на строительство детской площадки Система зажигания
Система зажигания Хоккей в Латвии, сборная, лига
Хоккей в Латвии, сборная, лига . Историко-культурный стандарт
. Историко-культурный стандарт Культура Мадагаскара
Культура Мадагаскара Технология работ по переработке вагонов на сортировочных станциях
Технология работ по переработке вагонов на сортировочных станциях  Принципы решения неструктурированных проблем
Принципы решения неструктурированных проблем Мультивибратор
Мультивибратор Механизм таможенно-тарифного регулирования в ЕАЭС. (Лекция 5)
Механизм таможенно-тарифного регулирования в ЕАЭС. (Лекция 5) Презентация____
Презентация____ Драматический театр 19 век
Драматический театр 19 век Коррозия металлов
Коррозия металлов  Введение в программирование (часть 2). Переменные и константы
Введение в программирование (часть 2). Переменные и константы Технология изготовления сварной двутавровой балки
Технология изготовления сварной двутавровой балки Состояние и перспективы развития сферы культуры Прионежского муниципального района
Состояние и перспективы развития сферы культуры Прионежского муниципального района Концепция наружного рекламно-информационного оформления сооружения
Концепция наружного рекламно-информационного оформления сооружения ВКР: «Проектирование ремонтной прорези и грузового причала на Сеньковском перекате реки Оки»
ВКР: «Проектирование ремонтной прорези и грузового причала на Сеньковском перекате реки Оки» Кунгурская городская управа
Кунгурская городская управа Қазақстан республикасы президенті, Нұрсұлтан Назарбаевтың халыққа жолдауы - нақты істер қазығы
Қазақстан республикасы президенті, Нұрсұлтан Назарбаевтың халыққа жолдауы - нақты істер қазығы Методы планирования потребности в персонале
Методы планирования потребности в персонале Инновационная экономика и технологическое предпринимательство
Инновационная экономика и технологическое предпринимательство Образование оболочек сердца
Образование оболочек сердца PHONETICS and other sciences
PHONETICS and other sciences  Эвристические процедуры в ПР
Эвристические процедуры в ПР Презентация Фрахт
Презентация Фрахт Презентация на тему "Программа начального образования" - скачать презентации по Педагогике
Презентация на тему "Программа начального образования" - скачать презентации по Педагогике الخطأ السياقي
الخطأ السياقي