- Регуляция дыхания

Содержание
- 2. План лекции 1. Регуляция дыхания: значение хемо и механо-рецепторов; дыхательный центр; рефлекторная и гуморальная регуляция. 2.
- 3. Дыхательная функция легких заключается в поддержании относительно постоянного уровня напряжения О2 и СО2 Различают : Нормоксию
- 4. Дыхательная система создает полное соответствие между количеством О2, поступающего через лёгкие в кровь и скоростью его
- 6. Регуляция дыхания Происхождение дыхательного ритма
- 7. Но кажется зачем контроль? О чём говорят следующие наблюдения? Не успел подумать, а уже сделал вдох.
- 8. – это совокупность взаимно связанных нейронов продолговатого мозга, обеспечивающих только периодичность дыхания(вдох-выдох) (эстафетная передача) М.Флуранс(1842)и Н.А.Миславский(1885
- 9. – это сложное структурно-функциональное объединение нервных клеток, расположенных на различных уровнях ЦНС (от спинного до коры
- 10. Система дыхания также как и сердце человека работают в определенном постоянном ритме Особенностью функции внешнего дыхания
- 11. Внешнее звено: Воздухоносные пути и легкие. Грудная клетка и мышцы (костно-мышечный каркас) Внутреннее звено 1. Кровь
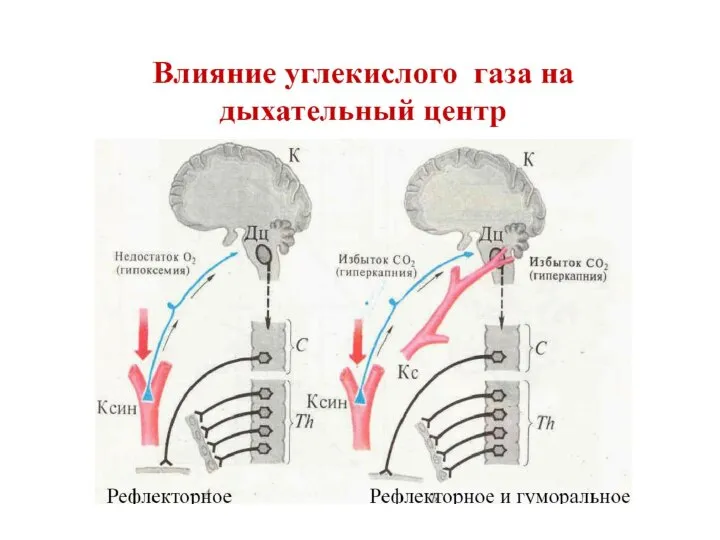
- 12. Структуры дыхательного центра подвержены нервной и гуморальной регуляции под влиянием метаболических изменений По отлонению (рО2, рСО2,
- 13. Система регуляции дыхания включает три основных элемента: 1) рецепторы, воспринимающие информацию и передающие ее в: 2)
- 14. Центральный регулятор Кора больших полушарий (ПРОИЗВОЛЬНЫЙ КОНТРОЛЬ) Лимбическая система Гипоталамус Варолиев мост-ПТЦ Продолговатый мозг Сегменты спинного
- 15. Опыт Фредерика с перекрестным кровообращением роль газового состава Перерезают сонные артерии и яремные вены и соединяют
- 16. Рецепторы, регулирующие дыхание Хеморецепторы (изменение О2, рН и СО2) 1.1. Периферические хеморецепторы (в каротидном и аортальных
- 17. Периферические хеморецепторы расположены в аортальных и каротидных тельцах Каротидное тельце Аортальные тельца Петля отрицательной обратной связи
- 18. Периферические хеморецепторы: Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их концентрация максимальна в синокаротидной
- 19. Рефлексогенные зоны реагируют на все три стимула; имеют высокую чувствительность к изменению pO2 артериальной крови (особенно
- 20. Рефлексогенные зоны от аортального тельца афферентная импульсация проводится по нерву Циона—Людвига (ветвь X нерва) и от
- 21. Рефлекторная активация центра вдоха
- 22. Центральные хеморецепторы расположены на вентральной поверхности продолговатого мозга Ростральная хеморецепторная область Каудальная хеморецепторная область
- 23. Центральные хеморецепторы (хеморецепторная зона дыхательного центра): Отличаются высокой чувствительностью к изменению рН (пороговые колебания рН примерно
- 24. Звенья системы регуляции дыхания М S L Центральные (медулярные) хеморецепторы ( рН )
- 27. р СО2 р О2
- 28. Современные представления о структуре дыхательного центра
- 29. НАПОМИНАЮ ! Дыхательным центром называют совокупность нейронов, расположенных в разных отделах центральной нервной системы, обеспечивающих соответствие
- 30. Большой вклад в изучение его работы внесли М.Флуранс (1822 г.), Н.А. Миславский (1885 г.), Р.Баумгартен (1956
- 31. С III-Y Th- I-X Иннервация органов дыхания
- 32. Происхождение дыхательного ритма Регуляция дыхания Спинальный контроль. Мотонейроны спинного мозга на уровне CIII-CV управляют диафрагмой, а
- 33. Отделы ДЦ расположены билатерально (правая – левая половины) в продолговато мозге в области дна 4-го желудочка.
- 34. Скопление инспираторных нейронов Скопление эксппир. нейронов их перекрытие Локализация дыхательного центра в продолговатом мозгу - а
- 35. ЯДЕРНЫЕ ГРУППЫ ПРОДОЛГОВАТОГО МОЗГА ВРГ ДРГ жкт NTS 4-й желудочек Баро Хемо П р о д
- 38. Продольный срез ствола ВРГ – вентральная респираторная группа нейронов ДРГ - дорсальная респираторная группа нейронов В
- 39. Генерация дыхательного ритма В течение дыхательного цикла выделяют три фазы активности дыхательных нейронов: инспираторную постинспираторную экспираторную
- 41. Инспираторная фаза(ИФ) Соответствует вдоху. Инспираторные нейроны, активные во время вдоха различаются по характеру разряда и времени
- 42. Дыхательные нейроны ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха (расположены в вентролатеральнои группе);
- 43. Постинспираторная фаза Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными нейронами, которые тормозят как
- 44. Экспираторная фаза Соответствует второй половине выдоха (активная экспирация) и обусловлена активацией экспираторных нейронов, иннервирующих мотонейроны мышц
- 45. Дыхательные нейроны постинспираторные нейроны — интернейроны, активные в первой половине выдоха и, тормозящие как инспираторные, так
- 46. Фазы активности дыхательного центра
- 47. Группы дыхательных нейронов Генераторы дыхательного ритма Формирующие дыхательный паттерн ранние инспираторные постинспираторные полные инспираторные поздние инспираторные
- 48. Почему возбуждаются ДРГ и ВРГ или природа дыхательного ритмогенеза Во сне, в бессознательном состоянии дыхание осуществляется
- 51. 4 2 1 3 5 6 Д М Network model
- 52. Принципиальная схема строения дыхательного центра Дыхательный центр образован ядрами продолговатого мозга и моста. В нем происходит
- 53. Дыхательные центры ствола головного мозга Продолговатого мозга (2) вентролатеральный (3) Дорсомедиальный (4) Варолиева моста (1) Пневмотаксический
- 54. Структура медуллярного центра ДДГ и ВДГ локализованы в Medulla двусторонне в виде 2 столбов около Obex
- 55. Вентральная дыхательная группа нейронов (ВДГ) расположена латеральнее обоюдного ядра продолговатого мозга, или ядра блуждающего нерва. ВДГ
- 56. Разделение обязанностей между структурами дыхательного центра
- 57. Схема нервных связей дыхательного центра: Ио — инспираторный отдел дыхательного центра продолговатого мозга; Эо — экспираторный
- 58. Дыхательные центры Варолиева моста
- 59. Пневмотаксический центр моста (Т.Лумсден, 1960). В клювовидно-латеральной (верхней) части варолиева моста располагается пневмотаксический центр [ Медиальное
- 60. Разделение обязанностей между структурами дыхательного центра
- 61. Дыхательные центры Варолиева моста
- 62. Нейроны варолиева моста - оригинальный выключатель вдоха Нейроны пневмотаксического центра реципрокно связаны с нейронами ДДГ и
- 63. Схема генерации дыхательного ритма ПТЦ Инс Экс II.
- 64. Влияние с механорецепторов на дыхательный центр
- 65. РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ В слизистой оболочке верхних носовых ходов между эпителиальными и опорными клетками расположены обонятельные рецепторы.
- 67. Рецепторы растяжения легких Расположены в гладких мышцах трахеи и бронхах, а также в подслизистом слое и
- 68. Классификация механорецепторов легких Статические Динамические Низкопороговые Высокопороговые
- 70. Рефлекторные реакции с рецепторов дыхательных путей Афферентные и эфферентные пути этих рефлексов проходят в стволах обонятельного,
- 71. Схема генерации дыхательного ритма Обратная связь от рецепторов легких Рефлекс Геринга-Брейера: растяжение легких прерывает вдох и
- 72. 2) Экспираторно-облегчающий проявляется в условиях, когда воздух поступает в дыхательные пути под давлением во время выдоха.
- 73. Ирритантные рецепторы Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов растяжения легких; Работают кратковременно. Могут
- 74. Парадоксальный рефлекс Хеда Ирритантные рецепторы принимают участие в возникновении своеобразного рефлекса, так называемого рефлекса "вздоха". В
- 75. Кашлевой рефлекс. вызывается раздражением ирритантных рецепторов глотки и нижележащих дыхательных путей, особенно области бифуркации трахеи характеризуется
- 76. Рефлекс чихания. Он возникает при раздражении рецепторов слизистой носа и носоглотки, в виде серии выдыхательных движений
- 77. Рецепторы легких и дыхательных путей (сигналы от них идут в продолговатый мозг по волокнам блуждающего нерва)
- 78. Юкстаальвеолярные рецепторы (J-рецепторы) Лёгочные J- рецепторы расположены в интерстиции альвеол в месте контакта их с капиллярами.
- 79. Проприорецепторы дыхательных мышц Проприорецепторы (в том числе мышечные веретена и γ- мотонейроны)в периферических суставах и мыщцах
- 80. Схема генерации дыхательного ритма ПТЦ Инс Экс II. III
- 81. Схема генерации дыхательного ритма ПТЦ Инс Экс II. I III
- 83. Схема генерации дыхательного ритма ПТЦ Инс Экс II. I III
- 87. Куссемауля- при снижении рН крови(метаболический ацидоз при сахарном диабете). Дыхание очень глубокое - компенсирует ацидоз.
- 89. Асфиксия (удушье) - прекращение поступления воздуха в дыхательные пути как в результате внешних причин , так
- 90. Влияние высших отделов ЦНС на дыхательный центр
- 91. Гипоталамус Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме. Регулирует дыхание для обеспечения поведенческих
- 92. Лимбическая система Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.
- 93. Кора больших полушарий По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно на спинальные моторные центры
- 94. Неспецифические регуляторы дыхания Возбуждают дыхательный центр Сильные холодовые или тепловые воздействия на кожу Физическая нагрузка Боль
- 95. Иннервация дыхательного центра Психические процессы Хеморецепторы Механорецепторы Терморецепторы Рецепторы верхних дыхательных путей Болевые воздействия Органы чувств
- 96. Контроль дыхания
- 97. Первый вдох новорожденного Стимулятором вдоха являются гуморальные факторы: Пережатие пуповины - аноксия Увеличение рСО2 - гиперкапния;
- 98. Гипоксия-типовой патологический процесс, возникающий в результате недостаточности биологического окисления и обусловленной ею энергетической необеспеченности жизненных процессов.
- 100. Основные типы гипоксии 1) экзогенный – гипобарический, гипербарический и нормобарический; 2) респираторный (дыхательный); 3) циркуляторный (сердечно-сосудистый);
- 101. Влияние высоты над уровнем моря на парциальное давление кислорода Высота Атмосфера Альвеола 0 760 149 2000
- 102. Denver, – 5280 футов (1609 м) над уровнем моря. Ратм – 630 мм рт ст
- 103. Дыхание в необычных условиях Пониженное атмосферное давление: Проявляется с высоты около 3,5 км, появляются симптомы горной
- 104. Влияние высоты на кривую диссоциации оксигемоглобина
- 106. В горах чайник кипит при более низкой температуре (давление паров насыщения = атмосферному давлению)
- 107. КРИТИЧЕСКИЕ ПОРОГИ ГИПОКСИИ Патологические эффекты кислородного голодания Нейтральная зона (индиферентнаая).- до 2км Зона полной компенсации -
- 108. Патологические эффекты кислородного голодания - 2 Зона неполной компенсации (зона опасности) . На высотах 4000-7000 м
- 110. Газовая эмболия. После погружения на глубину важно медленно возвращаться к нормальному давлению, т.к. азот не может
- 112. Мёртвое море – 1378 футов (420 м) ниже уровня моря. Ратм – 800 мм рт ст
- 115. Оксигенобаротерапия Лечение кислородом под повышенным давлением до 130-140 мм рт.ст (в барокамерах)при норме 100 мм рт.ст.
- 116. Допустимая продолжительность дыхания здорового человека при повышении рО2 0,1—2 ата не ограничена; 0,5 ата- до 133
- 120. Скачать презентацию

План лекции
1. Регуляция дыхания: значение хемо и механо-рецепторов; дыхательный центр;
План лекции
1. Регуляция дыхания: значение хемо и механо-рецепторов; дыхательный центр;

Дыхательная функция легких заключается в поддержании относительно постоянного уровня напряжения О2
Дыхательная функция легких заключается в поддержании относительно постоянного уровня напряжения О2

Дыхательная система создает полное соответствие между количеством О2, поступающего через лёгкие
Дыхательная система создает полное соответствие между количеством О2, поступающего через лёгкие


Регуляция дыхания
Происхождение дыхательного ритма
Регуляция дыхания
Происхождение дыхательного ритма

Но кажется зачем контроль?
О чём говорят следующие наблюдения?
Не успел подумать, а
Но кажется зачем контроль?
О чём говорят следующие наблюдения?
Не успел подумать, а

– это совокупность взаимно связанных нейронов продолговатого мозга, обеспечивающих только периодичность
– это совокупность взаимно связанных нейронов продолговатого мозга, обеспечивающих только периодичность

– это сложное структурно-функциональное объединение нервных клеток, расположенных на различных уровнях
– это сложное структурно-функциональное объединение нервных клеток, расположенных на различных уровнях

Система дыхания также как и сердце человека работают в определенном постоянном
Система дыхания также как и сердце человека работают в определенном постоянном

Внешнее звено:
Воздухоносные пути и легкие.
Грудная клетка и мышцы (костно-мышечный каркас)
Внешнее звено:
Воздухоносные пути и легкие.
Грудная клетка и мышцы (костно-мышечный каркас)
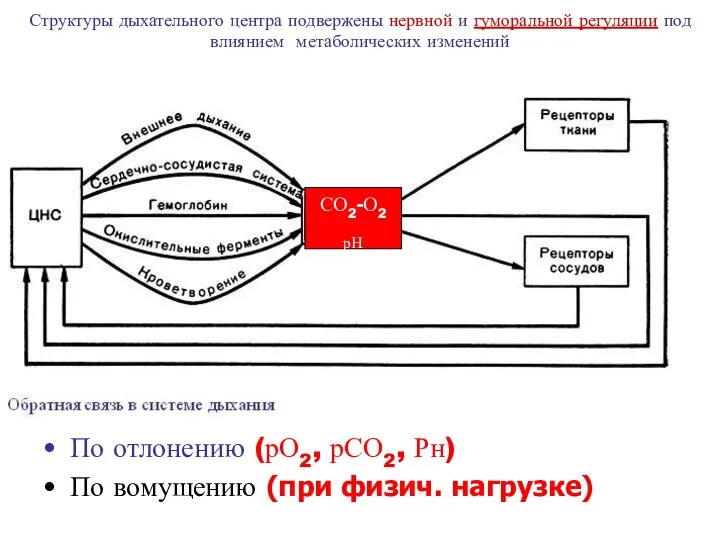
Структуры дыхательного центра подвержены нервной и гуморальной регуляции под влиянием метаболических
Структуры дыхательного центра подвержены нервной и гуморальной регуляции под влиянием метаболических

Система регуляции дыхания включает три основных элемента:
1) рецепторы, воспринимающие информацию и
Система регуляции дыхания включает три основных элемента:
1) рецепторы, воспринимающие информацию и
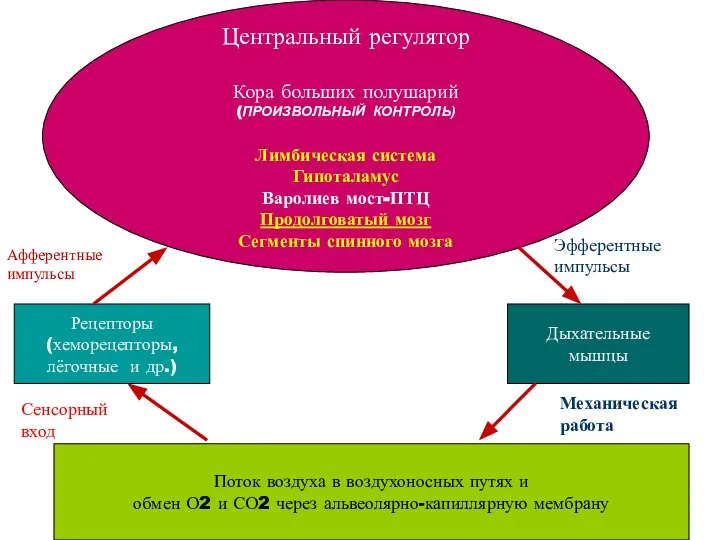
Центральный регулятор
Кора больших полушарий
(ПРОИЗВОЛЬНЫЙ КОНТРОЛЬ)
Лимбическая система
Гипоталамус
Варолиев мост-ПТЦ
Продолговатый мозг
Сегменты спинного мозга
Поток воздуха
Центральный регулятор
Кора больших полушарий
(ПРОИЗВОЛЬНЫЙ КОНТРОЛЬ)
Лимбическая система
Гипоталамус
Варолиев мост-ПТЦ
Продолговатый мозг
Сегменты спинного мозга
Поток воздуха
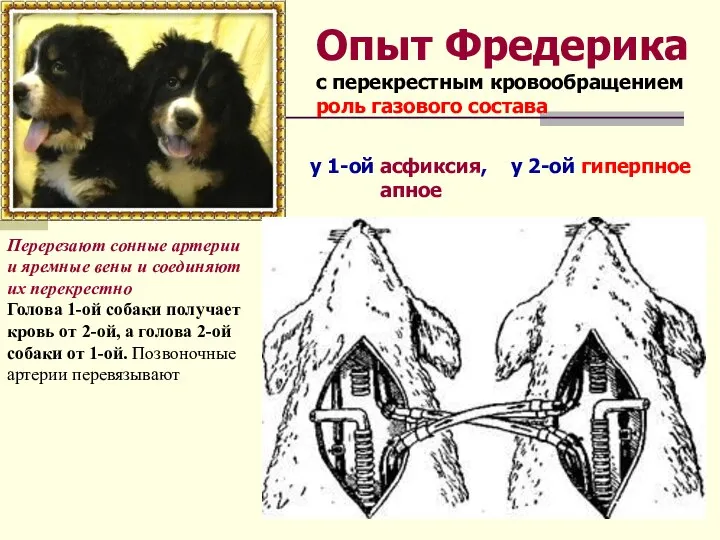
Опыт Фредерика
с перекрестным кровообращением
роль газового состава
Перерезают сонные артерии и яремные
Опыт Фредерика
с перекрестным кровообращением
роль газового состава
Перерезают сонные артерии и яремные

Рецепторы, регулирующие дыхание
Хеморецепторы (изменение О2, рН и СО2)
1.1. Периферические
Рецепторы, регулирующие дыхание
Хеморецепторы (изменение О2, рН и СО2)
1.1. Периферические
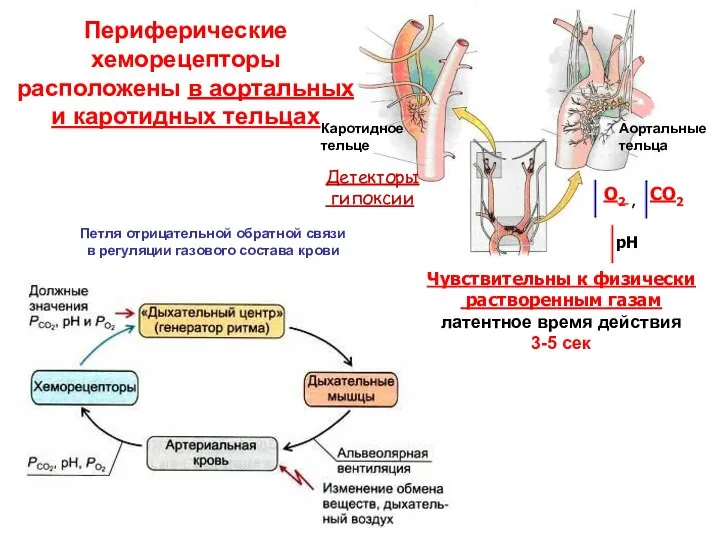
Периферические хеморецепторы расположены в аортальных
и каротидных тельцах
Каротидное тельце
Аортальные тельца
Петля отрицательной
Периферические хеморецепторы расположены в аортальных
и каротидных тельцах
Каротидное тельце
Аортальные тельца
Петля отрицательной
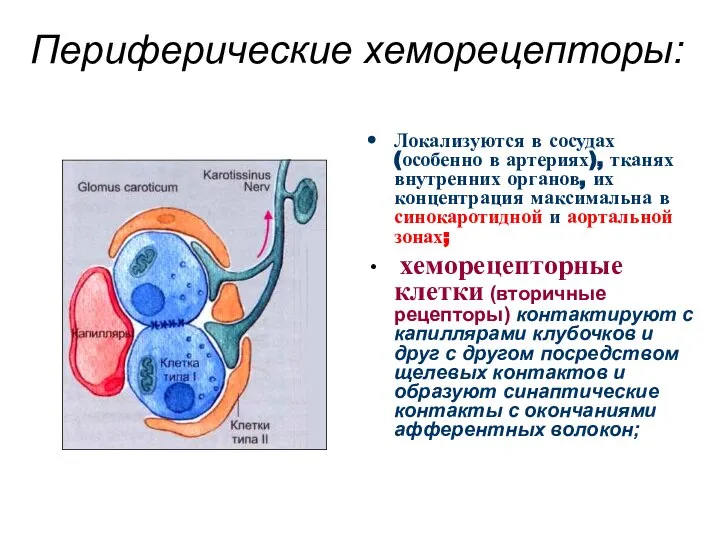
Периферические хеморецепторы:
Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их
Периферические хеморецепторы:
Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их
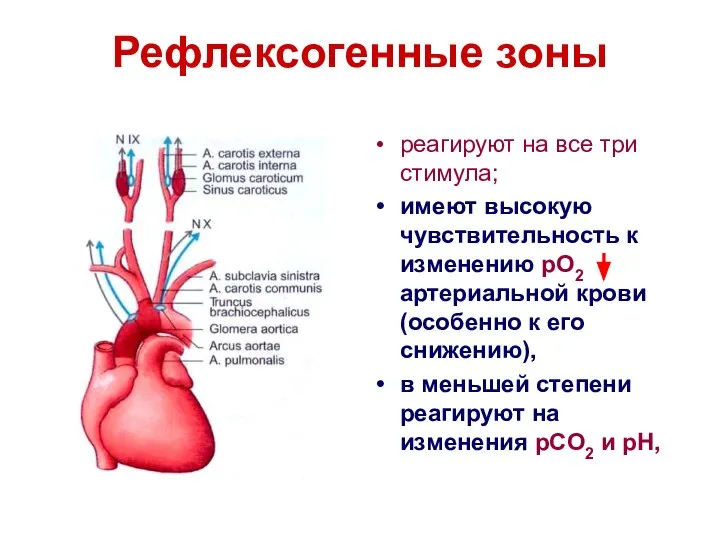
Рефлексогенные зоны
реагируют на все три стимула;
имеют высокую чувствительность к изменению pO2
Рефлексогенные зоны
реагируют на все три стимула;
имеют высокую чувствительность к изменению pO2
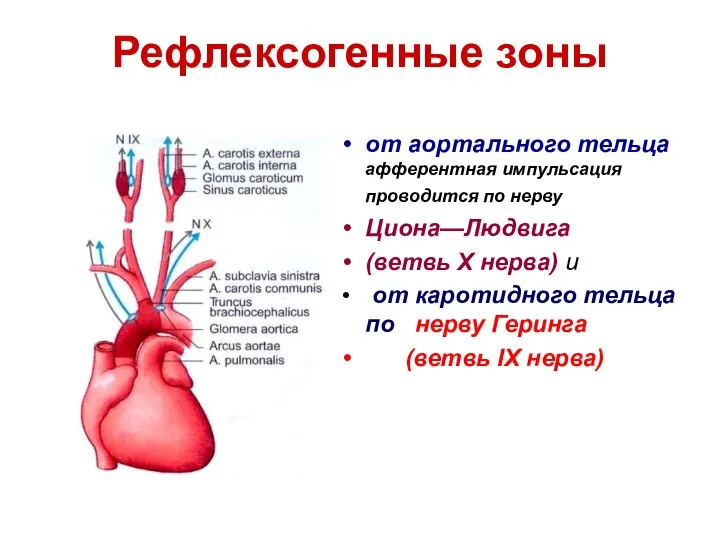
Рефлексогенные зоны
от аортального тельца афферентная импульсация проводится по нерву
Циона—Людвига
(ветвь
Рефлексогенные зоны
от аортального тельца афферентная импульсация проводится по нерву
Циона—Людвига
(ветвь
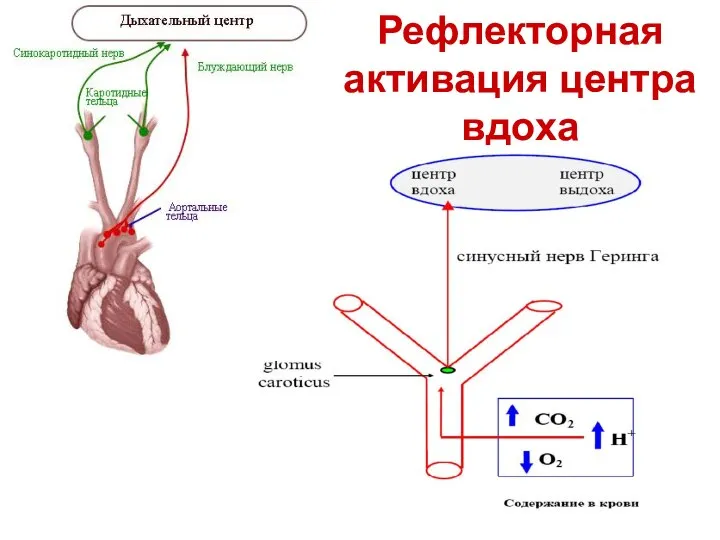
Рефлекторная активация центра вдоха
Рефлекторная активация центра вдоха
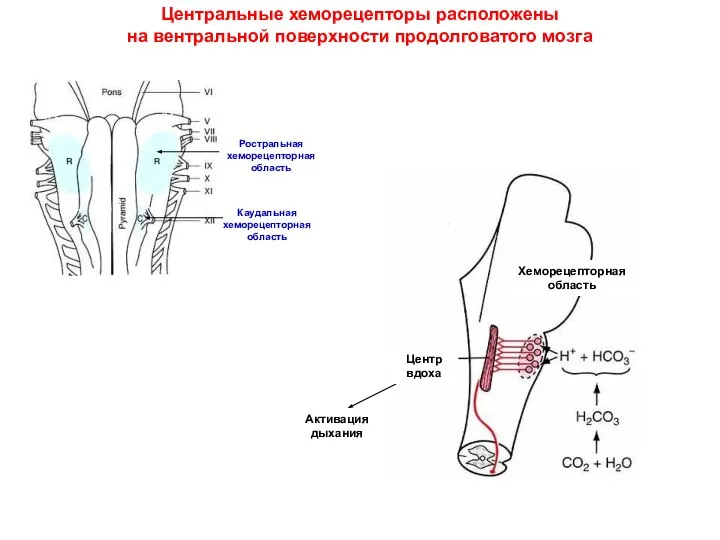
Центральные хеморецепторы расположены
на вентральной поверхности продолговатого мозга
Ростральная хеморецепторная область
Каудальная хеморецепторная
Центральные хеморецепторы расположены
на вентральной поверхности продолговатого мозга
Ростральная хеморецепторная область
Каудальная хеморецепторная

Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Отличаются высокой чувствительностью к изменению рН
Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Отличаются высокой чувствительностью к изменению рН
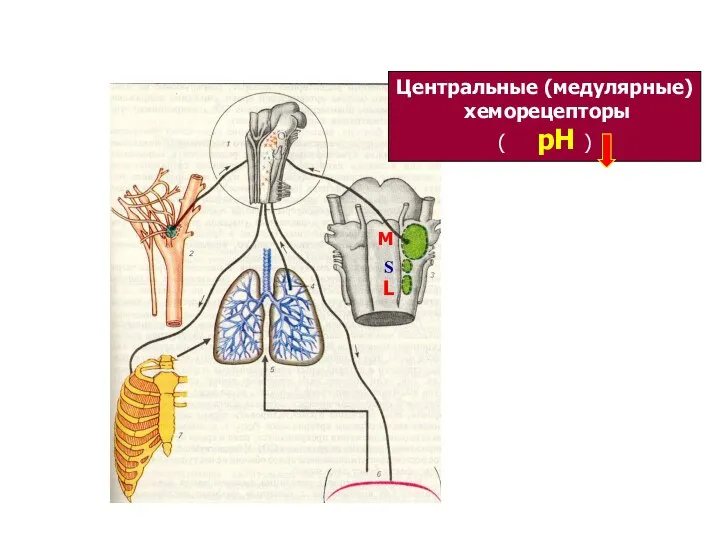
Звенья системы регуляции дыхания
М
S
L
Центральные (медулярные)
хеморецепторы
( рН )
Звенья системы регуляции дыхания
М
S
L
Центральные (медулярные)
хеморецепторы
( рН )
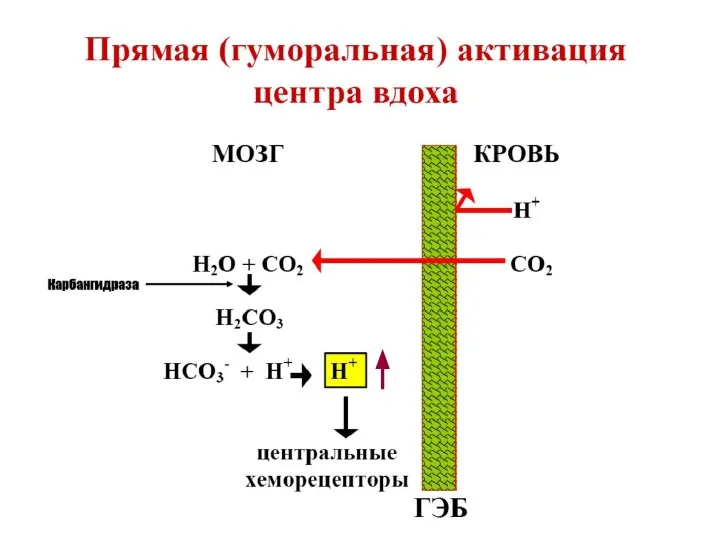
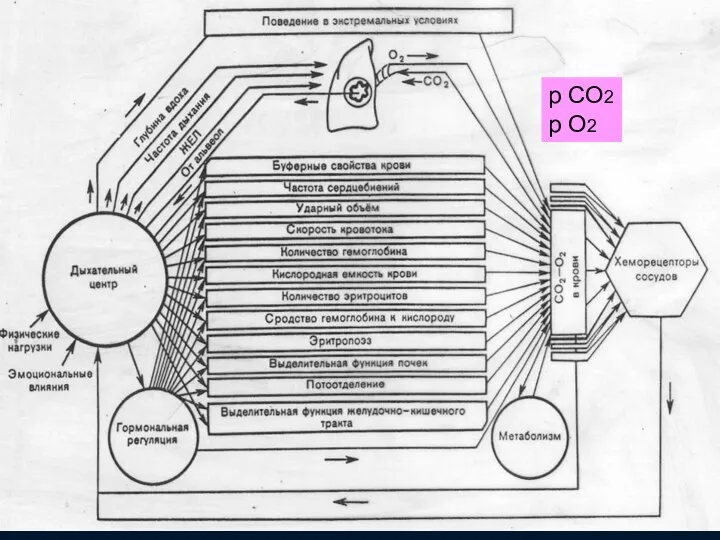
р СО2
р О2
р СО2
р О2

Современные представления
о структуре дыхательного центра
Современные представления
о структуре дыхательного центра

НАПОМИНАЮ !
Дыхательным центром называют совокупность нейронов, расположенных в разных отделах центральной
НАПОМИНАЮ !
Дыхательным центром называют совокупность нейронов, расположенных в разных отделах центральной

Большой вклад в изучение его работы внесли
М.Флуранс (1822 г.),
Н.А.
Большой вклад в изучение его работы внесли
М.Флуранс (1822 г.),
Н.А.
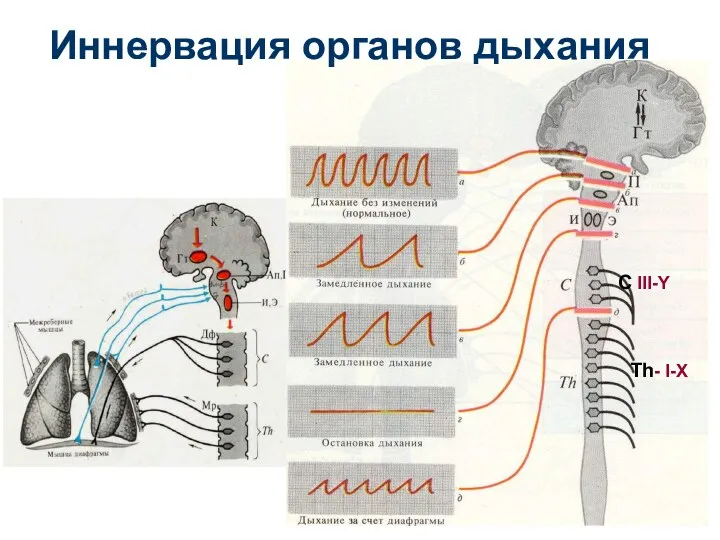
С III-Y
Th- I-X
Иннервация органов дыхания
С III-Y
Th- I-X
Иннервация органов дыхания
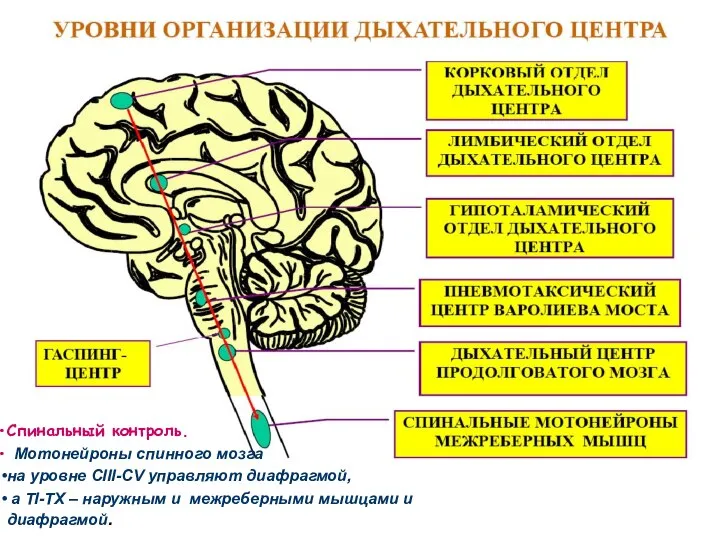
Происхождение дыхательного ритма
Регуляция дыхания
Спинальный контроль.
Мотонейроны спинного мозга
на уровне
Происхождение дыхательного ритма
Регуляция дыхания
Спинальный контроль.
Мотонейроны спинного мозга
на уровне

Отделы ДЦ расположены билатерально (правая – левая половины)
в продолговато мозге
Отделы ДЦ расположены билатерально (правая – левая половины)
в продолговато мозге
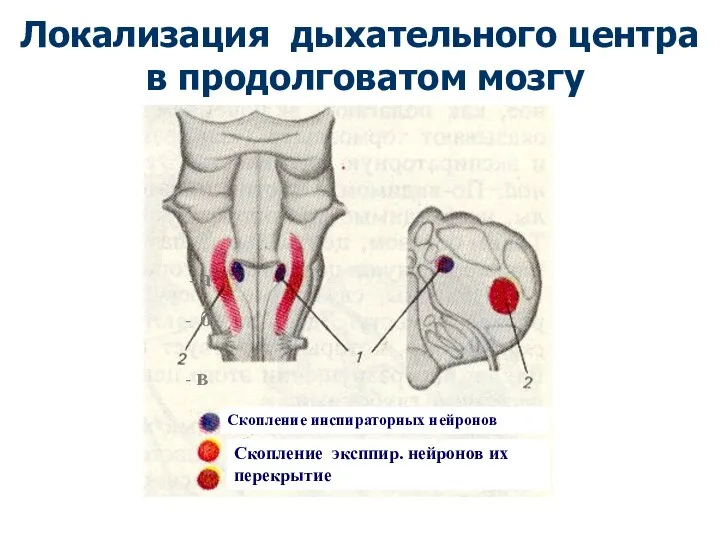
Скопление инспираторных нейронов
Скопление эксппир. нейронов их перекрытие
Локализация дыхательного центра
в продолговатом
Скопление инспираторных нейронов
Скопление эксппир. нейронов их перекрытие
Локализация дыхательного центра в продолговатом
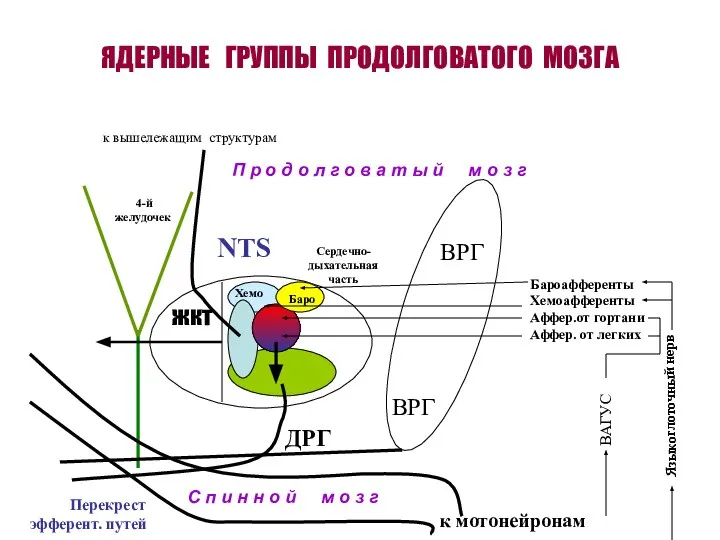
ЯДЕРНЫЕ ГРУППЫ ПРОДОЛГОВАТОГО МОЗГА
ВРГ
ДРГ
жкт
NTS
4-й желудочек
Баро
Хемо
П р о
ЯДЕРНЫЕ ГРУППЫ ПРОДОЛГОВАТОГО МОЗГА
ВРГ
ДРГ
жкт
NTS
4-й желудочек
Баро
Хемо
П р о


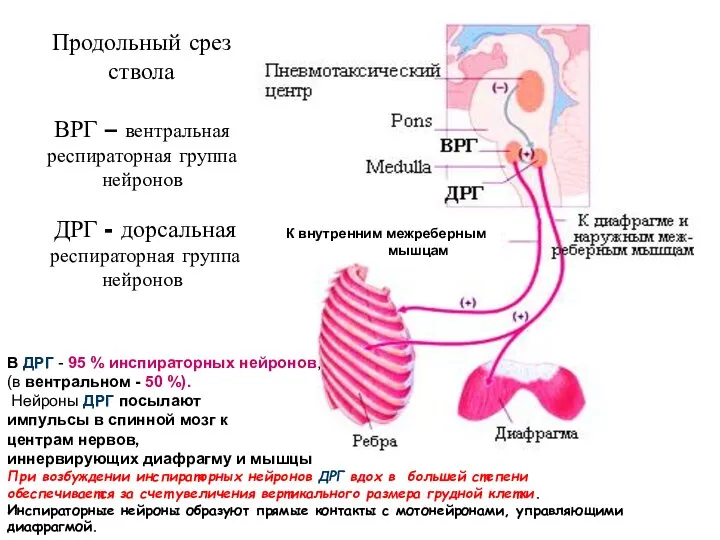
Продольный срез ствола
ВРГ – вентральная респираторная группа нейронов
ДРГ - дорсальная
Продольный срез ствола ВРГ – вентральная респираторная группа нейронов ДРГ - дорсальная

Генерация дыхательного ритма
В течение дыхательного цикла выделяют три фазы активности дыхательных
Генерация дыхательного ритма
В течение дыхательного цикла выделяют три фазы активности дыхательных


Инспираторная фаза(ИФ)
Соответствует вдоху.
Инспираторные нейроны, активные во время вдоха различаются по
Инспираторная фаза(ИФ)
Соответствует вдоху.
Инспираторные нейроны, активные во время вдоха различаются по

Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха
Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха

Постинспираторная фаза
Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными
Постинспираторная фаза
Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными

Экспираторная фаза
Соответствует второй половине выдоха (активная экспирация) и обусловлена активацией
Экспираторная фаза
Соответствует второй половине выдоха (активная экспирация) и обусловлена активацией

Дыхательные нейроны
постинспираторные нейроны — интернейроны, активные в первой половине выдоха и,
Дыхательные нейроны
постинспираторные нейроны — интернейроны, активные в первой половине выдоха и,
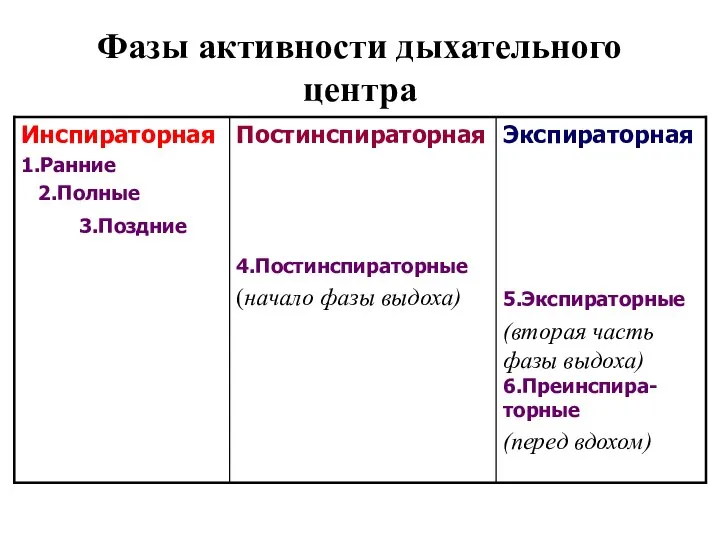
Фазы активности дыхательного центра
Фазы активности дыхательного центра

Группы дыхательных нейронов
Генераторы
дыхательного ритма
Формирующие
дыхательный паттерн
ранние инспираторные
постинспираторные
полные инспираторные
поздние
Группы дыхательных нейронов
Генераторы
дыхательного ритма
Формирующие
дыхательный паттерн
ранние инспираторные
постинспираторные
полные инспираторные
поздние

Почему возбуждаются ДРГ и ВРГ или природа дыхательного ритмогенеза
Во сне, в
Почему возбуждаются ДРГ и ВРГ или природа дыхательного ритмогенеза
Во сне, в


4
2
1
3
5
6
Д М
Network model
4
2
1
3
5
6
Д М
Network model
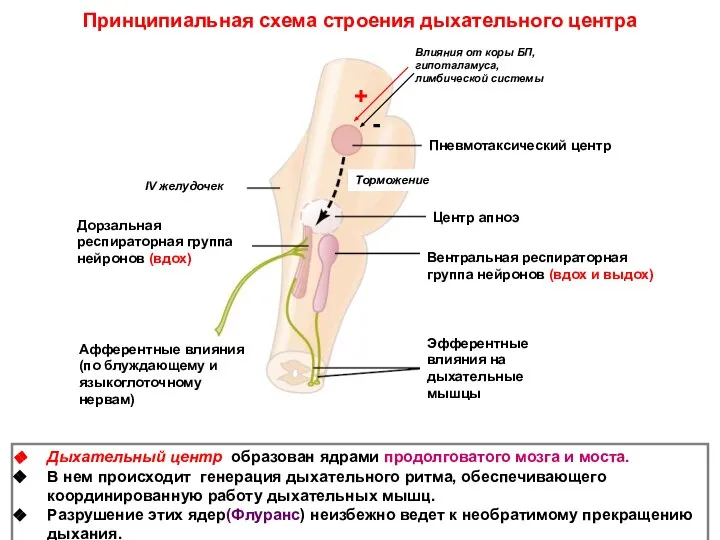
Принципиальная схема строения дыхательного центра
Дыхательный центр образован ядрами продолговатого мозга
Принципиальная схема строения дыхательного центра
Дыхательный центр образован ядрами продолговатого мозга
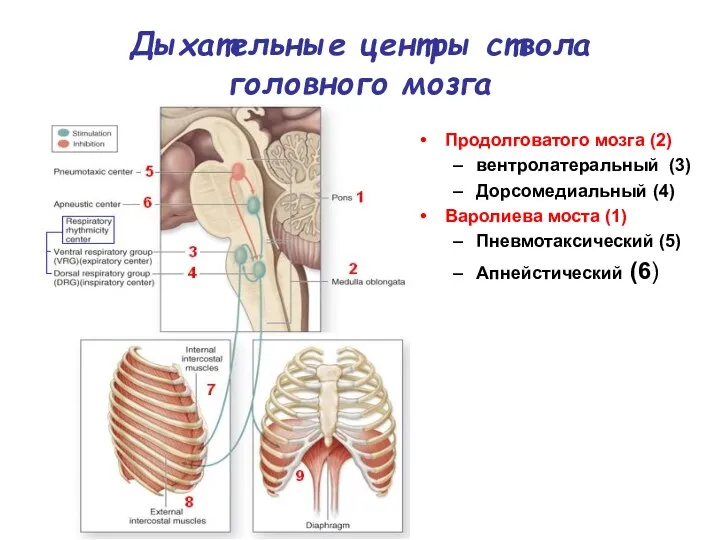
Дыхательные центры ствола головного мозга
Продолговатого мозга (2)
вентролатеральный (3)
Дорсомедиальный (4)
Варолиева моста
Дыхательные центры ствола головного мозга
Продолговатого мозга (2)
вентролатеральный (3)
Дорсомедиальный (4)
Варолиева моста

Структура медуллярного центра
ДДГ и ВДГ локализованы в Medulla двусторонне в виде
Структура медуллярного центра
ДДГ и ВДГ локализованы в Medulla двусторонне в виде

Вентральная дыхательная
группа нейронов (ВДГ)
расположена латеральнее обоюдного ядра продолговатого мозга, или
Вентральная дыхательная
группа нейронов (ВДГ)
расположена латеральнее обоюдного ядра продолговатого мозга, или
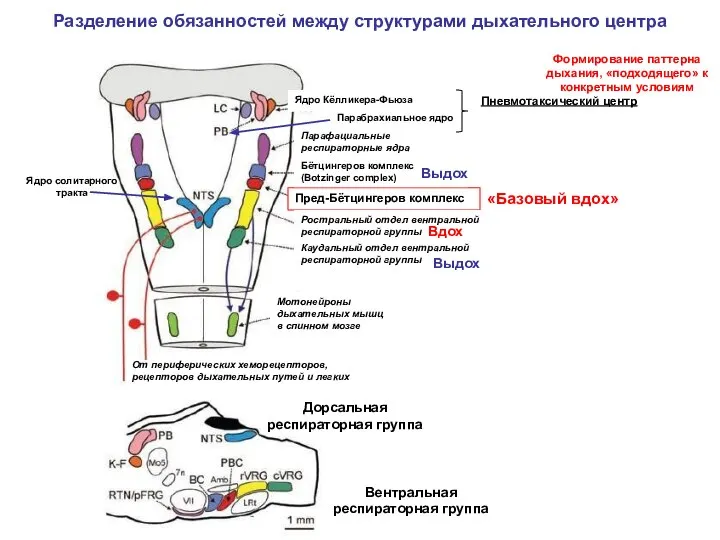
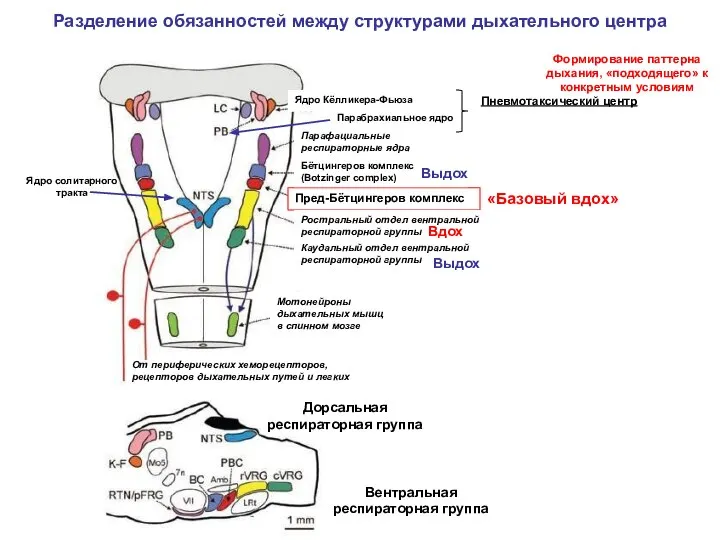
Разделение обязанностей между структурами дыхательного центра
Разделение обязанностей между структурами дыхательного центра
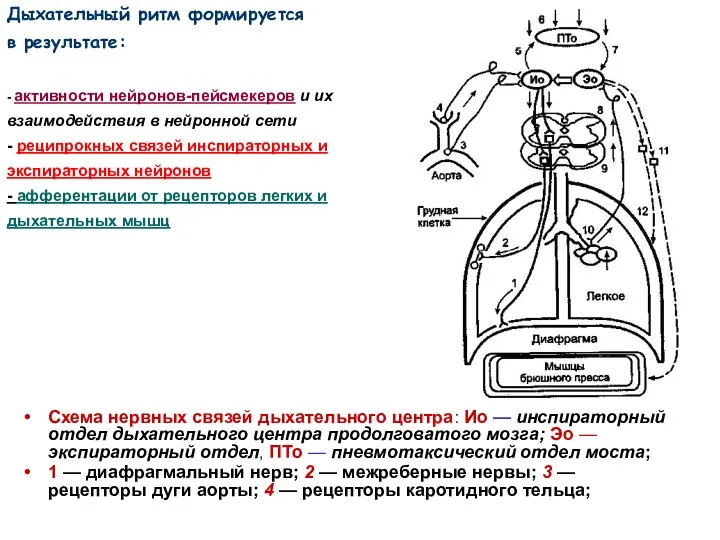
Схема нервных связей дыхательного центра: Ио — инспираторный отдел дыхательного центра
Схема нервных связей дыхательного центра: Ио — инспираторный отдел дыхательного центра

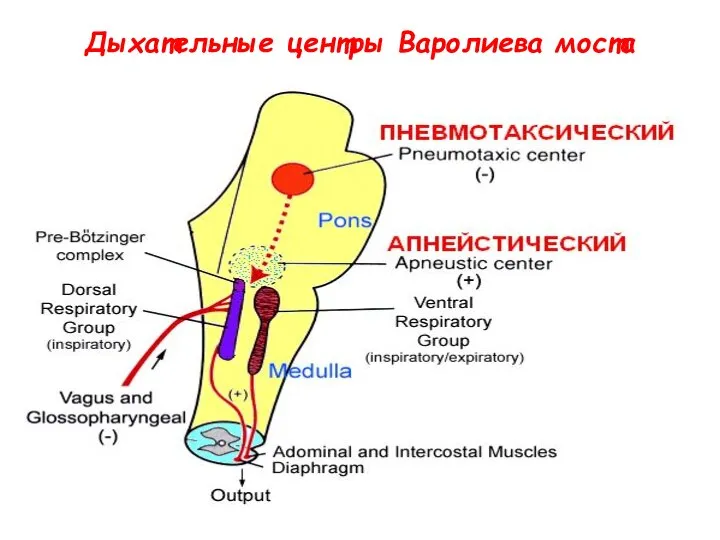
Дыхательные центры Варолиева моста
Дыхательные центры Варолиева моста

Пневмотаксический центр моста (Т.Лумсден, 1960).
В клювовидно-латеральной (верхней) части варолиева моста располагается
Пневмотаксический центр моста (Т.Лумсден, 1960).
В клювовидно-латеральной (верхней) части варолиева моста располагается
Разделение обязанностей между структурами дыхательного центра
Разделение обязанностей между структурами дыхательного центра
Дыхательные центры Варолиева моста
Дыхательные центры Варолиева моста

Нейроны варолиева моста - оригинальный выключатель вдоха
Нейроны пневмотаксического центра реципрокно связаны
Нейроны варолиева моста - оригинальный выключатель вдоха
Нейроны пневмотаксического центра реципрокно связаны
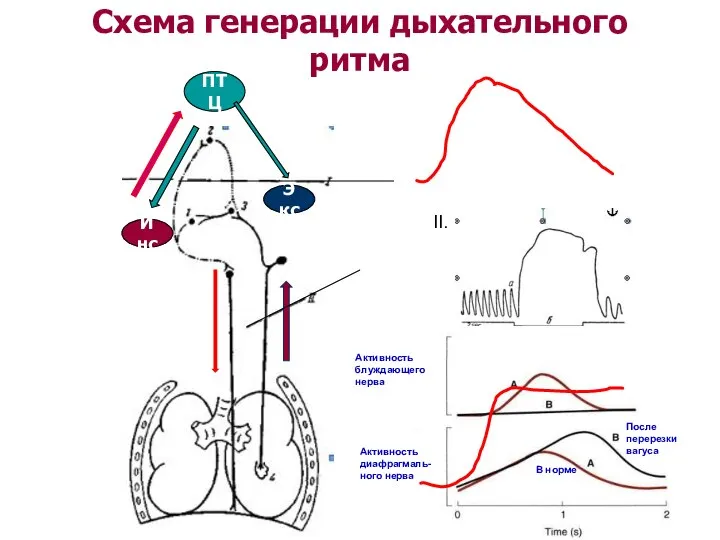
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.

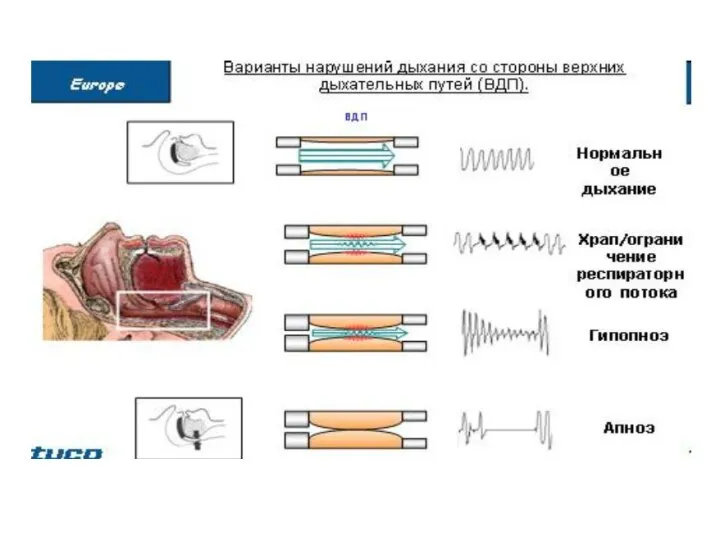
Влияние с механорецепторов на дыхательный центр
Влияние с механорецепторов на дыхательный центр

РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ
В слизистой оболочке верхних носовых ходов между эпителиальными и опорными
РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ
В слизистой оболочке верхних носовых ходов между эпителиальными и опорными

Рецепторы растяжения легких
Расположены в гладких мышцах трахеи и бронхах, а
Рецепторы растяжения легких
Расположены в гладких мышцах трахеи и бронхах, а

Классификация механорецепторов легких
Статические
Динамические
Низкопороговые
Высокопороговые
Классификация механорецепторов легких
Статические
Динамические
Низкопороговые
Высокопороговые


Рефлекторные реакции с рецепторов дыхательных путей
Афферентные и эфферентные пути этих рефлексов
Рефлекторные реакции с рецепторов дыхательных путей
Афферентные и эфферентные пути этих рефлексов
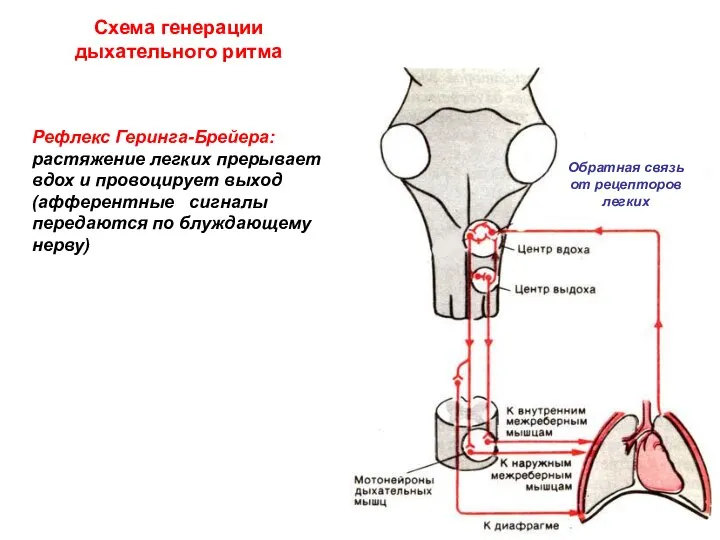
Схема генерации дыхательного ритма
Обратная связь от рецепторов легких
Рефлекс Геринга-Брейера:
растяжение легких
Схема генерации дыхательного ритма
Обратная связь от рецепторов легких
Рефлекс Геринга-Брейера:
растяжение легких

2) Экспираторно-облегчающий проявляется в условиях, когда воздух поступает в дыхательные пути
2) Экспираторно-облегчающий проявляется в условиях, когда воздух поступает в дыхательные пути

Ирритантные рецепторы
Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов
Ирритантные рецепторы
Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов

Парадоксальный рефлекс Хеда
Ирритантные рецепторы принимают участие в возникновении своеобразного рефлекса, так
Парадоксальный рефлекс Хеда
Ирритантные рецепторы принимают участие в возникновении своеобразного рефлекса, так

Кашлевой рефлекс.
вызывается раздражением ирритантных рецепторов глотки и нижележащих дыхательных
Кашлевой рефлекс.
вызывается раздражением ирритантных рецепторов глотки и нижележащих дыхательных

Рефлекс чихания.
Он возникает при раздражении рецепторов слизистой носа и носоглотки,
Рефлекс чихания. Он возникает при раздражении рецепторов слизистой носа и носоглотки,
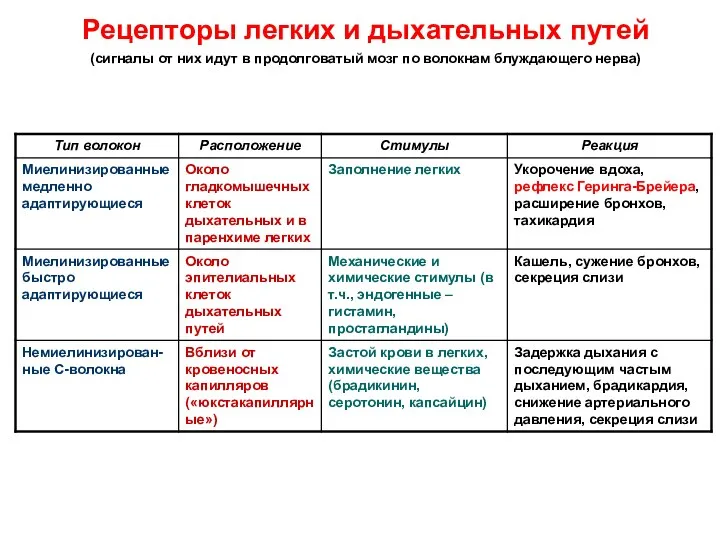
Рецепторы легких и дыхательных путей
(сигналы от них идут в продолговатый
Рецепторы легких и дыхательных путей (сигналы от них идут в продолговатый

Юкстаальвеолярные рецепторы (J-рецепторы)
Лёгочные J- рецепторы расположены в интерстиции альвеол в
Юкстаальвеолярные рецепторы (J-рецепторы)
Лёгочные J- рецепторы расположены в интерстиции альвеол в

Проприорецепторы дыхательных мышц
Проприорецепторы (в том числе мышечные веретена и γ- мотонейроны)в
Проприорецепторы дыхательных мышц
Проприорецепторы (в том числе мышечные веретена и γ- мотонейроны)в
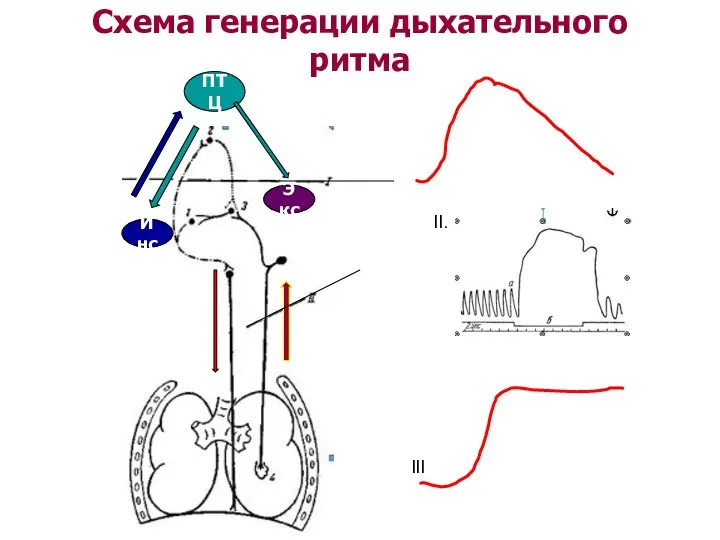
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
III
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
III
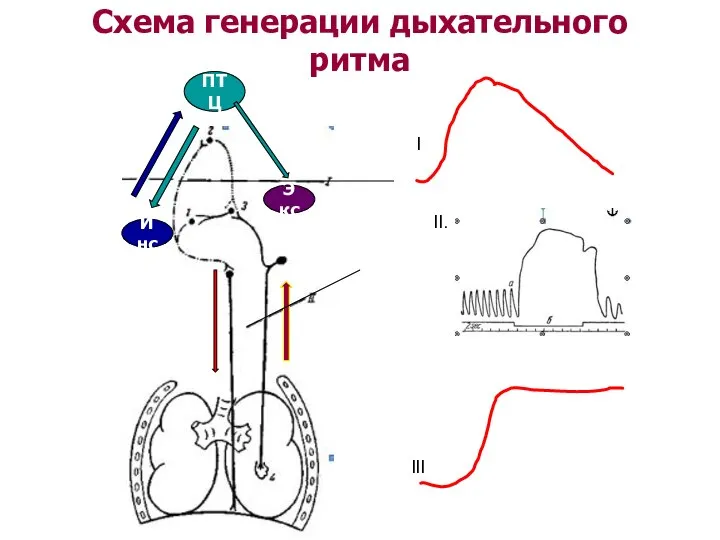
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
I
III
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
I
III

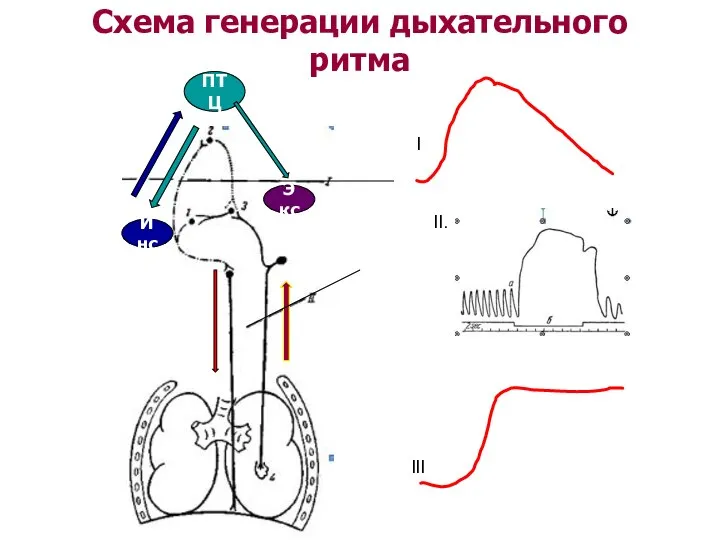
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
I
III
Схема генерации дыхательного ритма
ПТЦ
Инс
Экс
II.
I
III



Куссемауля- при снижении рН крови(метаболический ацидоз при сахарном диабете). Дыхание очень
Куссемауля- при снижении рН крови(метаболический ацидоз при сахарном диабете). Дыхание очень
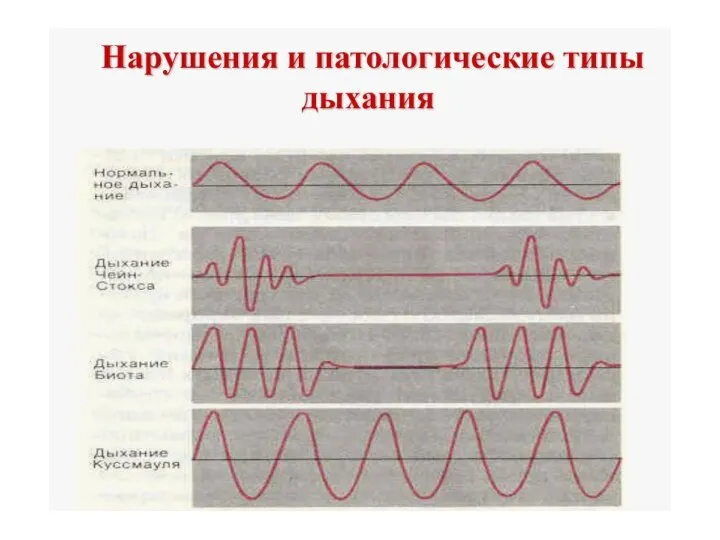

Асфиксия (удушье) - прекращение поступления воздуха в дыхательные пути как в
Асфиксия (удушье) - прекращение поступления воздуха в дыхательные пути как в

Влияние высших отделов ЦНС на дыхательный центр
Влияние высших отделов ЦНС на дыхательный центр

Гипоталамус
Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме.
Регулирует дыхание
Гипоталамус
Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме.
Регулирует дыхание

Лимбическая система
Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.
Лимбическая система
Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.

Кора больших полушарий
По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно
Кора больших полушарий
По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно

Неспецифические регуляторы дыхания
Возбуждают дыхательный центр
Сильные холодовые или тепловые воздействия на
Неспецифические регуляторы дыхания
Возбуждают дыхательный центр
Сильные холодовые или тепловые воздействия на
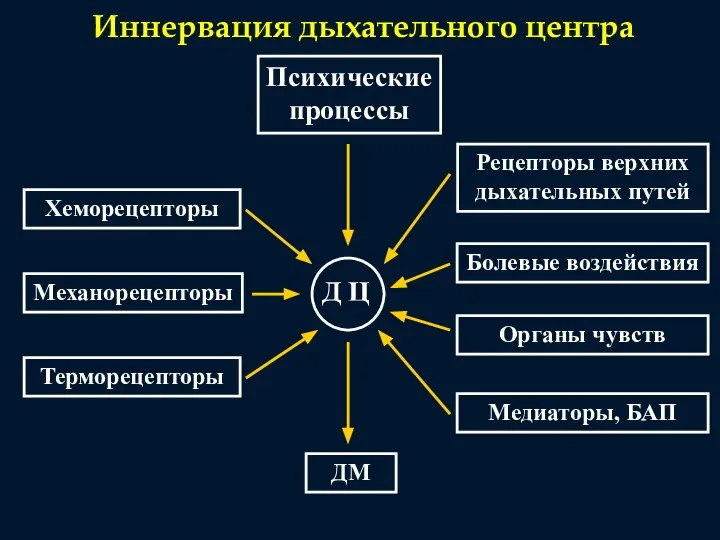
Иннервация дыхательного центра
Психические
процессы
Хеморецепторы
Механорецепторы
Терморецепторы
Рецепторы верхних
дыхательных путей
Болевые воздействия
Органы чувств
Медиаторы, БАП
ДМ
Д Ц
Иннервация дыхательного центра
Психические
процессы
Хеморецепторы
Механорецепторы
Терморецепторы
Рецепторы верхних
дыхательных путей
Болевые воздействия
Органы чувств
Медиаторы, БАП
ДМ
Д Ц
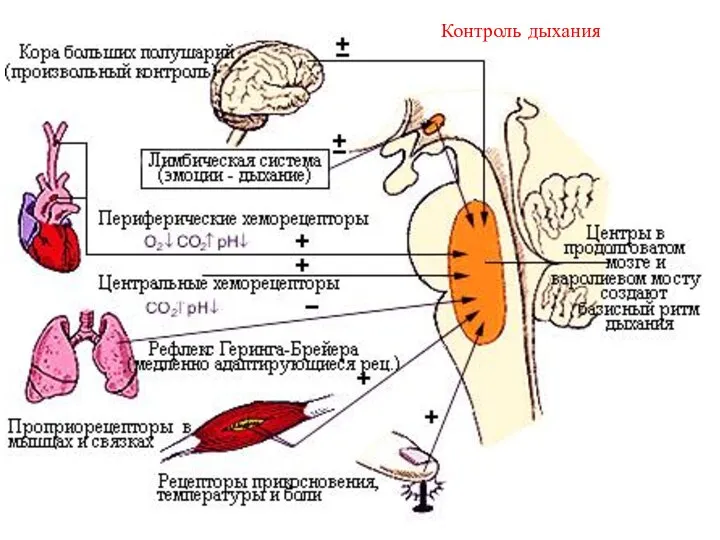
Контроль дыхания
Контроль дыхания

Первый вдох новорожденного
Стимулятором вдоха являются гуморальные факторы:
Пережатие пуповины - аноксия
Увеличение рСО2
Первый вдох новорожденного
Стимулятором вдоха являются гуморальные факторы:
Пережатие пуповины - аноксия
Увеличение рСО2

Гипоксия-типовой патологический процесс, возникающий в результате недостаточности биологического окисления и обусловленной
Гипоксия-типовой патологический процесс, возникающий в результате недостаточности биологического окисления и обусловленной
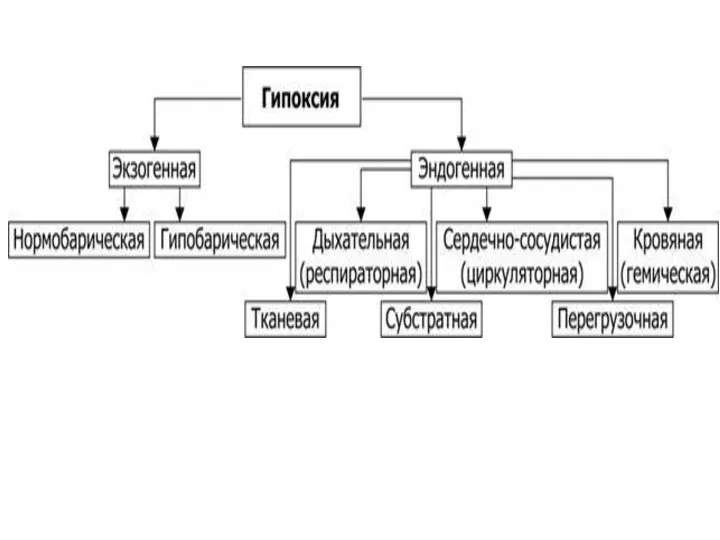

Основные типы гипоксии
1) экзогенный – гипобарический, гипербарический и нормобарический;
2) респираторный
Основные типы гипоксии
1) экзогенный – гипобарический, гипербарический и нормобарический;
2) респираторный
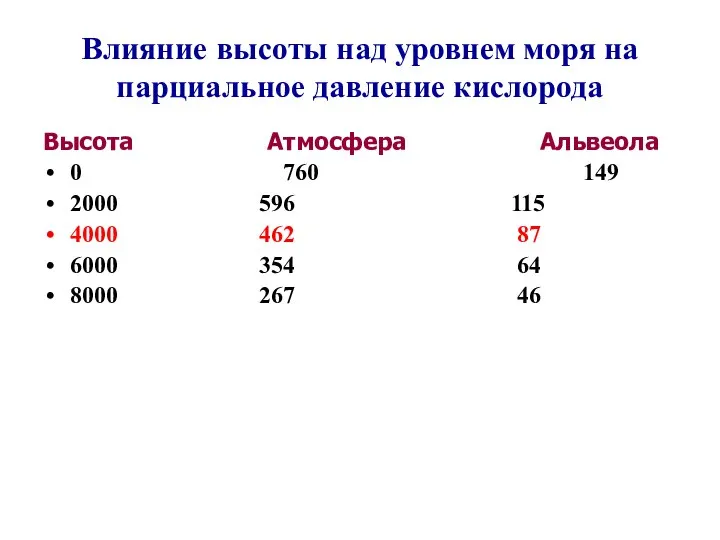
Влияние высоты над уровнем моря на парциальное давление кислорода
Высота Атмосфера Альвеола
Влияние высоты над уровнем моря на парциальное давление кислорода
Высота Атмосфера Альвеола

Denver, – 5280 футов (1609 м) над уровнем моря. Ратм –
Denver, – 5280 футов (1609 м) над уровнем моря. Ратм –

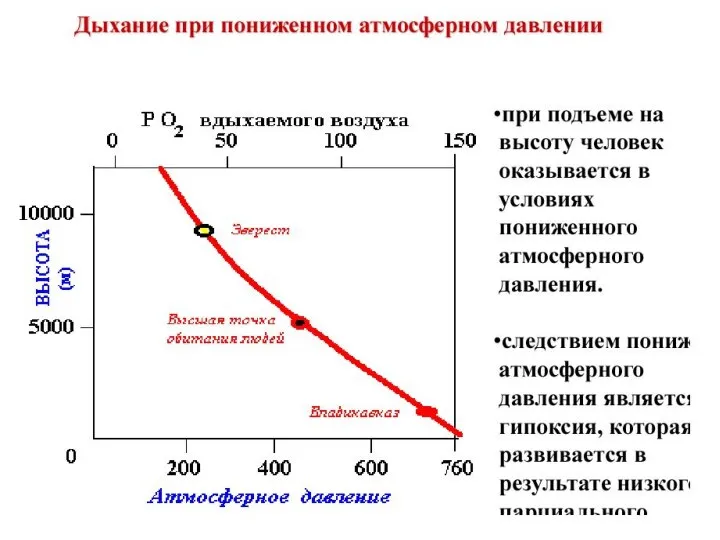
Дыхание в необычных условиях
Пониженное атмосферное давление:
Проявляется с высоты около 3,5
Дыхание в необычных условиях
Пониженное атмосферное давление:
Проявляется с высоты около 3,5
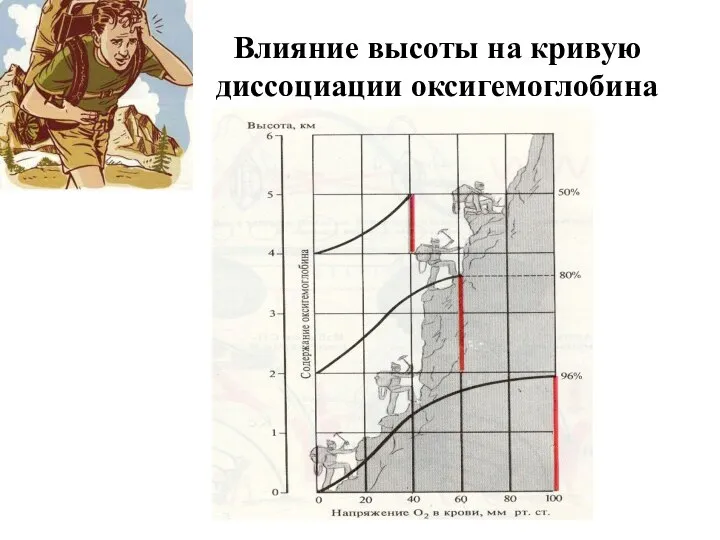
Влияние высоты на кривую диссоциации оксигемоглобина
Влияние высоты на кривую диссоциации оксигемоглобина

В горах чайник кипит при более низкой температуре
(давление паров насыщения
В горах чайник кипит при более низкой температуре (давление паров насыщения

КРИТИЧЕСКИЕ ПОРОГИ ГИПОКСИИ
Патологические эффекты кислородного голодания
Нейтральная зона (индиферентнаая).- до 2км
Зона
КРИТИЧЕСКИЕ ПОРОГИ ГИПОКСИИ
Патологические эффекты кислородного голодания
Нейтральная зона (индиферентнаая).- до 2км
Зона

Патологические эффекты кислородного голодания - 2
Зона неполной компенсации (зона опасности)
. На
Патологические эффекты кислородного голодания - 2
Зона неполной компенсации (зона опасности)
. На

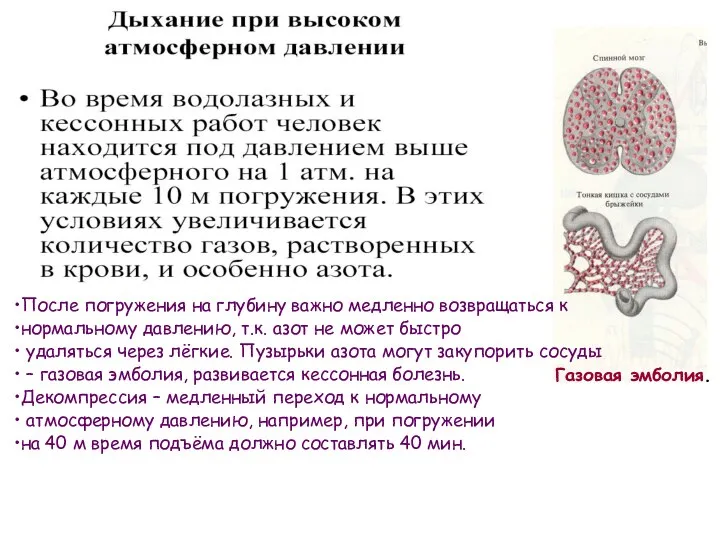
Газовая эмболия.
После погружения на глубину важно медленно возвращаться к
нормальному давлению,
Газовая эмболия.
После погружения на глубину важно медленно возвращаться к
нормальному давлению,


Мёртвое море – 1378 футов (420 м) ниже уровня моря. Ратм
Мёртвое море – 1378 футов (420 м) ниже уровня моря. Ратм



Оксигенобаротерапия
Лечение кислородом под повышенным давлением до 130-140 мм рт.ст (в барокамерах)при
Оксигенобаротерапия
Лечение кислородом под повышенным давлением до 130-140 мм рт.ст (в барокамерах)при

Допустимая продолжительность дыхания
здорового человека при повышении рО2
0,1—2 ата не ограничена;
0,5
Допустимая продолжительность дыхания
здорового человека при повышении рО2
0,1—2 ата не ограничена;
0,5

 Объединенная Партия России
Объединенная Партия России Закономерности распределения напряжений в грунте фазы деформаций грунта под фундаментом
Закономерности распределения напряжений в грунте фазы деформаций грунта под фундаментом УЧЕНИЕ О СМЕРТИ СУДЕБНО-МЕДИЦИНСКАЯ ТАНАТОЛОГИЯ
УЧЕНИЕ О СМЕРТИ СУДЕБНО-МЕДИЦИНСКАЯ ТАНАТОЛОГИЯ Примеры использования OpenMP. Вычисление определенного интеграла
Примеры использования OpenMP. Вычисление определенного интеграла Алфёров Жорес Иванович
Алфёров Жорес Иванович Роль образовательного учреждения по организации системы профилактики жестокого обращения с детьми
Роль образовательного учреждения по организации системы профилактики жестокого обращения с детьми  ВКР: Повышение производительности зерноуборочного комбайна «ACROS-595Plus»
ВКР: Повышение производительности зерноуборочного комбайна «ACROS-595Plus» Физическая нагрузка и адаптация в спорте. Работоспособность и восстановление в ИВС
Физическая нагрузка и адаптация в спорте. Работоспособность и восстановление в ИВС Глава 1 Принципы экономики 6. Из истории экономики и экономической науки
Глава 1 Принципы экономики 6. Из истории экономики и экономической науки  Л. Фейербах 1804-1872
Л. Фейербах 1804-1872 Всеволод Мейерхольд
Всеволод Мейерхольд Электронная подпись
Электронная подпись Культура этрусков
Культура этрусков  Загальні основи і закони ринкової економіки
Загальні основи і закони ринкової економіки Виды и структуры знаний, отражаемые в языке
Виды и структуры знаний, отражаемые в языке Camnoopy profile of smart home IP camera
Camnoopy profile of smart home IP camera Система технического обслуживания и ремонта автомобильной
Система технического обслуживания и ремонта автомобильной Особенности развития речи у детей с умственной отсталостью
Особенности развития речи у детей с умственной отсталостью Выбор конфигурации сетей Ethernet и Fast Ethernet
Выбор конфигурации сетей Ethernet и Fast Ethernet Эл пособие ч 2
Эл пособие ч 2 Предмет изучения дисциплины «Культурология и религиоведение»
Предмет изучения дисциплины «Культурология и религиоведение» Косач Лариса Петрівна - перлина та гордість Звягельщини
Косач Лариса Петрівна - перлина та гордість Звягельщини Гербы и геральдика
Гербы и геральдика Мәдениет құрылымы, функциясы. Мәдениет теориясы. Әлемдік мәдениеттер мен өркениеттер
Мәдениет құрылымы, функциясы. Мәдениет теориясы. Әлемдік мәдениеттер мен өркениеттер Нормативно-правовые документы, регламентирующие деятельность тренера
Нормативно-правовые документы, регламентирующие деятельность тренера Анализ процесса становления и развития института рецидива
Анализ процесса становления и развития института рецидива Эгейское искусство Материалы к уроку МХК В 10 классе
Эгейское искусство Материалы к уроку МХК В 10 классе Тема: «Историческое становление возрастной психологии»
Тема: «Историческое становление возрастной психологии»