- Репликация ДНК

Содержание
- 2. ПРИНЦИПЫ РЕПЛИКАЦИИ ДНК Комплемертарность Антипараллельность Полуконсервативность
- 3. Репликация ДНК – процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов и структурных белков Репликация ДНК
- 4. Три механизма роста дочерней нити ДНК, согласующиеся с полуконсервативной репликацией однонаправленный рост обеих нитей от двух
- 5. Принципы репликации ДНК Комплементарность Антипараллельность Полуконсервативность Двунаправленный рост двух дочерних цепей ДНК от одной специфической точки
- 6. Каждая эукариотическая хромосома - полирепликон Множественная инициация обеспечивает большую скорость репликации у эукариот Средняя хромосома (единица
- 7. Пять принципов репликации ДНК Комплементарность Антипараллельность Плуконсервативность Двунаправленность от одной специфической точки начала репликации (в большинстве
- 8. Положение каждого последующего нуклеотида в дочерней цепи ДНК однозначно определяется положением соответствующего нуклеотида матрицы
- 9. Общие свойства ДНК-полимера Не способны расплетать ДНК-дуплекс – нуждаются в однонитевой матрице. Могут только удлинять предсуществующую
- 10. Проблема репликации теломер – концов эукариотических хромосом Репликация конца запаздывающей цепи не может пройти полностью: после
- 11. Теломеры и телоомераза Функция теломер: - Защита от слияния концов линейных ДНК - Защита концов от
- 12. Длина теломерной ДНК как счетчик времени 1966 г. – гипотеза А. Оловникова (2-ой мед): Постепенное укорочение
- 13. Репликация теломер в гаметогенезе Укороченная запаздывающая цепь Удлинение ведущей цепи ДНК по матрице теломеразной РНК На
- 14. Повреждающиеся изменения в ДНК Точковые мутации Структурные нарушения
- 15. Система реперарации ДНК обеспечивают исправление 999 повреждений из 1000 Заболевания, обусловленные дефектами системы репарации: Пигментация ксеродерма
- 16. Репарация повреждений одной цепи ДНК Прямая реактивация повреждений Эксцизионная репарация (excizion – отсекание, вырезание): - Вырезание
- 17. Повреждения двух цепей ДНК Одноцепочные разрывы могут стать двухцепочечным во время репликации хромосом; Двухцепочечные разрывы могут
- 18. ГЕНЕТИЧЕСКИЙ КОД БИОСИНТЕЗ БЕЛКОВ (ТРАНСЛЯЦИЯ)
- 19. Биосинтез белков Наиболее сложный из генетических процессов У эукариот участвует > 300 макромолекул: >40 видов тРНК
- 20. Генетический код Генетический код – это способ записи генетической информации о структуре белков (полипептидов) посредством последовательности
- 22. Транскрипция и трансляция мРНК эукариот разобщены во времени и в пространстве
- 24. Скачать презентацию

ПРИНЦИПЫ РЕПЛИКАЦИИ ДНК
Комплемертарность
Антипараллельность
Полуконсервативность
ПРИНЦИПЫ РЕПЛИКАЦИИ ДНК
Комплемертарность
Антипараллельность
Полуконсервативность

Репликация ДНК – процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов
Репликация ДНК – процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов
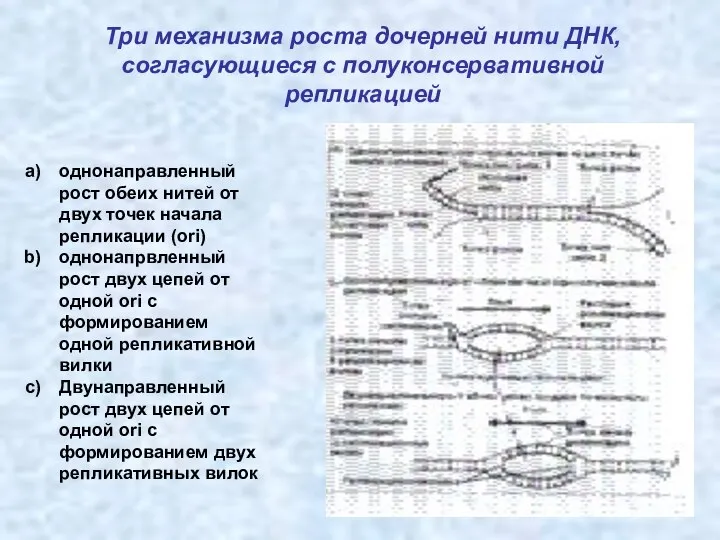
Три механизма роста дочерней нити ДНК, согласующиеся с полуконсервативной репликацией
однонаправленный рост
Три механизма роста дочерней нити ДНК, согласующиеся с полуконсервативной репликацией
однонаправленный рост

Принципы репликации ДНК
Комплементарность
Антипараллельность
Полуконсервативность
Двунаправленный рост двух дочерних цепей ДНК от одной специфической
Принципы репликации ДНК
Комплементарность
Антипараллельность
Полуконсервативность
Двунаправленный рост двух дочерних цепей ДНК от одной специфической

Каждая эукариотическая хромосома - полирепликон
Множественная инициация обеспечивает большую скорость репликации
Каждая эукариотическая хромосома - полирепликон
Множественная инициация обеспечивает большую скорость репликации

Пять принципов репликации ДНК
Комплементарность
Антипараллельность
Плуконсервативность
Двунаправленность от одной специфической точки начала репликации (в
Пять принципов репликации ДНК
Комплементарность
Антипараллельность
Плуконсервативность
Двунаправленность от одной специфической точки начала репликации (в
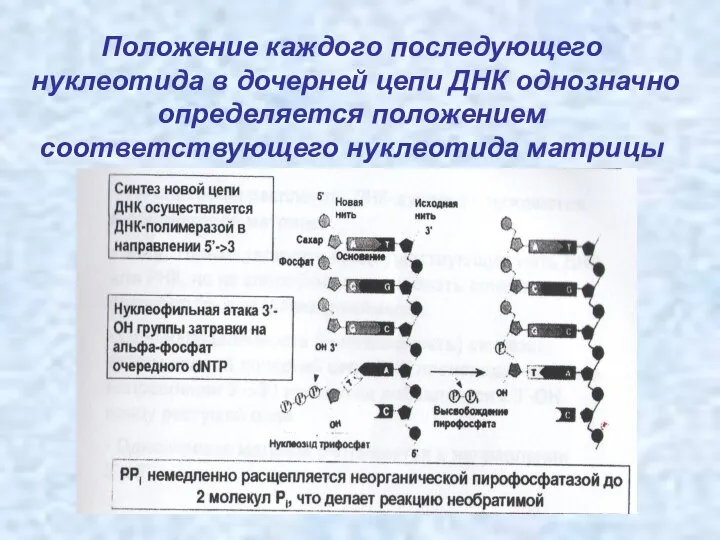
Положение каждого последующего нуклеотида в дочерней цепи ДНК однозначно определяется положением
Положение каждого последующего нуклеотида в дочерней цепи ДНК однозначно определяется положением

Общие свойства ДНК-полимера
Не способны расплетать ДНК-дуплекс – нуждаются в однонитевой
Общие свойства ДНК-полимера
Не способны расплетать ДНК-дуплекс – нуждаются в однонитевой

Проблема репликации теломер – концов эукариотических хромосом
Репликация конца запаздывающей
Проблема репликации теломер – концов эукариотических хромосом
Репликация конца запаздывающей

Теломеры и телоомераза
Функция теломер:
- Защита от слияния концов линейных
Теломеры и телоомераза
Функция теломер:
- Защита от слияния концов линейных

Длина теломерной ДНК как счетчик времени
1966 г. – гипотеза А.
Длина теломерной ДНК как счетчик времени
1966 г. – гипотеза А.

Репликация теломер в гаметогенезе
Укороченная запаздывающая цепь
Удлинение ведущей цепи ДНК по матрице
Репликация теломер в гаметогенезе
Укороченная запаздывающая цепь
Удлинение ведущей цепи ДНК по матрице

Повреждающиеся изменения в ДНК
Точковые мутации
Структурные нарушения
Повреждающиеся изменения в ДНК
Точковые мутации
Структурные нарушения
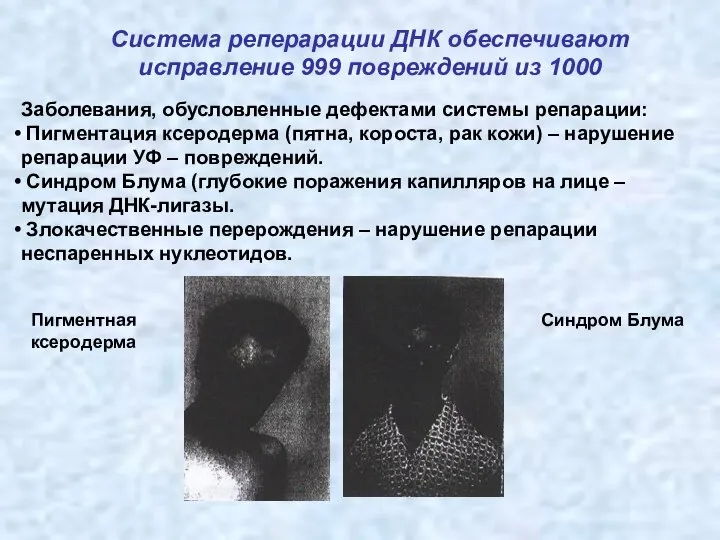
Система реперарации ДНК обеспечивают исправление 999 повреждений из 1000
Заболевания, обусловленные дефектами
Система реперарации ДНК обеспечивают исправление 999 повреждений из 1000
Заболевания, обусловленные дефектами

Репарация повреждений одной цепи ДНК
Прямая реактивация повреждений
Эксцизионная репарация (excizion
Репарация повреждений одной цепи ДНК
Прямая реактивация повреждений
Эксцизионная репарация (excizion
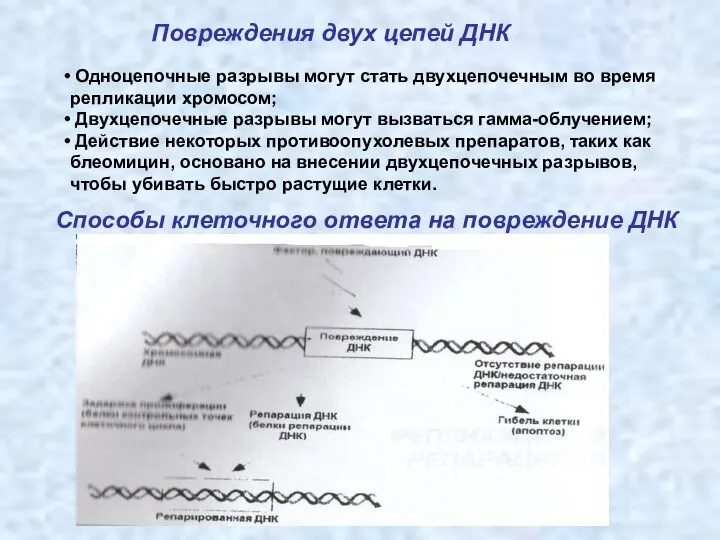
Повреждения двух цепей ДНК
Одноцепочные разрывы могут стать двухцепочечным во время
Повреждения двух цепей ДНК
Одноцепочные разрывы могут стать двухцепочечным во время

ГЕНЕТИЧЕСКИЙ КОД
БИОСИНТЕЗ БЕЛКОВ
(ТРАНСЛЯЦИЯ)
ГЕНЕТИЧЕСКИЙ КОД
БИОСИНТЕЗ БЕЛКОВ
(ТРАНСЛЯЦИЯ)

Биосинтез белков
Наиболее сложный из генетических процессов
У эукариот участвует >
Биосинтез белков
Наиболее сложный из генетических процессов
У эукариот участвует >

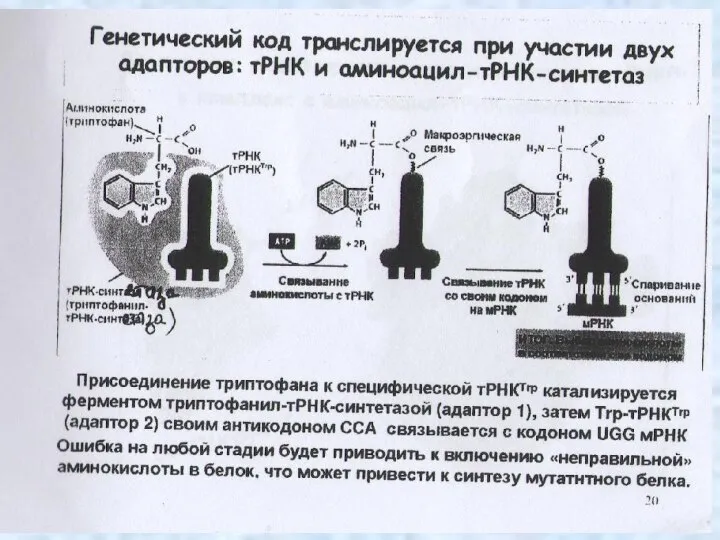
Генетический код
Генетический код – это способ записи генетической информации о структуре
Генетический код
Генетический код – это способ записи генетической информации о структуре
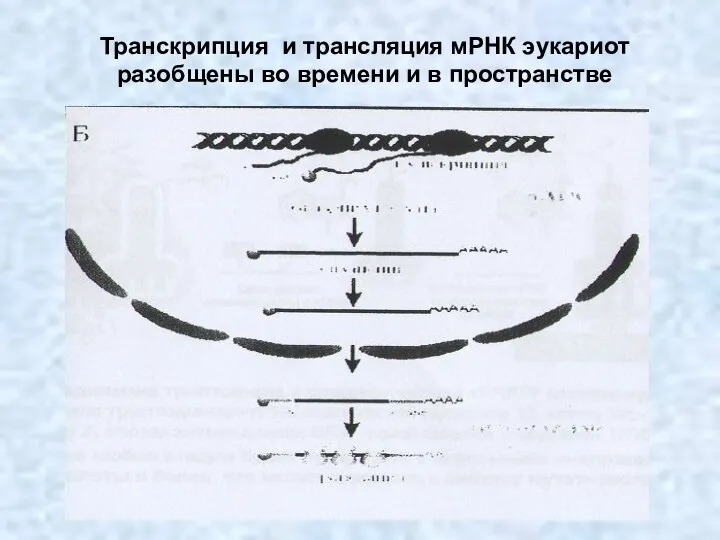
Транскрипция и трансляция мРНК эукариот разобщены во времени и в пространстве
Транскрипция и трансляция мРНК эукариот разобщены во времени и в пространстве
 Уильям Шекспир. Английский поэт и драматург
Уильям Шекспир. Английский поэт и драматург Психологическое сопровождение в спорте
Психологическое сопровождение в спорте Клинический разбор на тему: Особенности анестезиологического обеспечения лапароскопических операций у гинекологических больны
Клинический разбор на тему: Особенности анестезиологического обеспечения лапароскопических операций у гинекологических больны Символьные переменные в языке программирования Бейсик
Символьные переменные в языке программирования Бейсик Экология питания
Экология питания Презентация на тему "Развивающие приемы на уроках ИЗО" - скачать презентации по Педагогике
Презентация на тему "Развивающие приемы на уроках ИЗО" - скачать презентации по Педагогике Приложение в программе DELPHI в виде теста
Приложение в программе DELPHI в виде теста Генезис культуры в первобытную эпоху
Генезис культуры в первобытную эпоху Инфекционный процесс
Инфекционный процесс «CSS » Каскадные таблицы стилей.
«CSS » Каскадные таблицы стилей. Рабочая тетрадь
Рабочая тетрадь Электроника
Электроника  Презентация Тест по философии
Презентация Тест по философии Вставка таблиц в HTML-документ
Вставка таблиц в HTML-документ Уроки корейского языка
Уроки корейского языка Презентация Демографические проблемы России_
Презентация Демографические проблемы России_ Управление в таможенной деятельности
Управление в таможенной деятельности  Масленица Кустодиев Борис Михайлович (1878-1927) (автопортрет) 5 класс Учитель: Перевозникова Е.В.
Масленица Кустодиев Борис Михайлович (1878-1927) (автопортрет) 5 класс Учитель: Перевозникова Е.В. Аномалии развития кишечника
Аномалии развития кишечника  Эпоха возраждения и гуманизма
Эпоха возраждения и гуманизма Край любимый! Моя Родина, мой мир
Край любимый! Моя Родина, мой мир  Положительные стороны занятий в секции спортивного ориентирования
Положительные стороны занятий в секции спортивного ориентирования Место чувств в психике человека
Место чувств в психике человека Анимированный кроссворд «НАСЕКОМЫЕ» Составила: Травнева Ольга Юрьевна учитель начальных классов Республика Казахстан Кара
Анимированный кроссворд «НАСЕКОМЫЕ» Составила: Травнева Ольга Юрьевна учитель начальных классов Республика Казахстан Кара Scanner and printer
Scanner and printer Площадка для отдыха населения “Сказочный городок”
Площадка для отдыха населения “Сказочный городок” Заявка на разработку проекта «Производительность труда и поддержка занятости»
Заявка на разработку проекта «Производительность труда и поддержка занятости» Основы нравственной культуры народов России
Основы нравственной культуры народов России