- Синтез мочевины

Содержание
- 2. Пять путей обезвреживания аммиака Биосинтез мочевины в печени Восстановительное аминирование в тканях Образование амидов кислот в
- 3. Основной путь обезвреживания аммиака в организме – биосинтез мочевины в печени. В печень аммиак поступает в
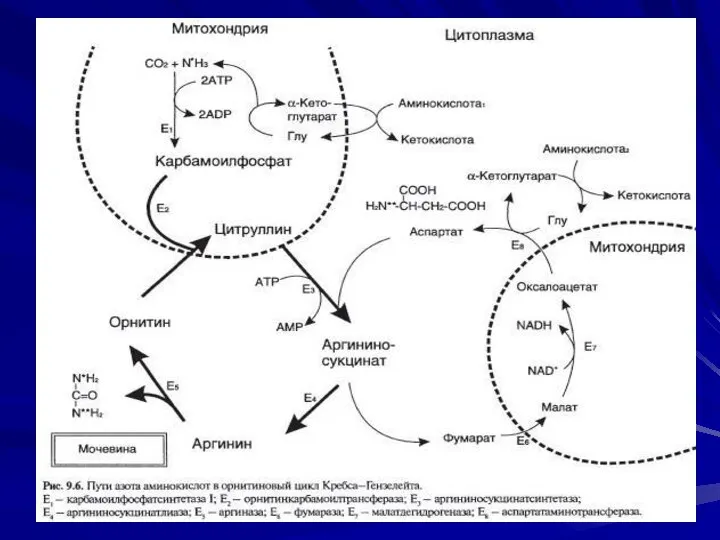
- 4. Орнитиновый цикл Суммарное уравнение синтезе мочевины печени: СО2+NH3+аспартат+3 АТФ+2 Н20 = Мочевина+фумарат+2АДФ+2H3P04+АМФ +Н4Р2О7 1 реакция:
- 5. 2-я реакция и 3 –я реакция
- 6. 4-я реакция
- 7. 5-я реакция
- 11. Энергетический баланс процесса В реакциях орнитинового цикла расходуются четыре макроэргических связи трёх молекул АТФ на каждый
- 12. Затраты энергии происходят также и при трансмембранном переносе веществ, связанном с синтезом и экскрецией мочевины. Первые
- 13. Ферменты орнитинового цикла распределены между митохондриями и цитозолем. Поэтому необходим трансмембранный перенос глутамата, цитруллина и орнитина
- 14. Биологическое значение орнитинового цикла 1. Обезвреживание аммиака в организме 2.Регуляция азотистого баланса в организме- при поступлении
- 15. Источники азота , углерода и кислорода в цикле мочевины 1-я амминогруппа - из свободного аммиака, образовавшегося
- 16. Гипераммониемия – повышение концентрации аммиака в крови. Причины: 1.Врожденная недостаточность хотя бы одного из ферментов биосинтеза
- 18. Скачать презентацию

Пять путей обезвреживания аммиака
Биосинтез мочевины в печени
Восстановительное аминирование в тканях
Образование
Пять путей обезвреживания аммиака
Биосинтез мочевины в печени
Восстановительное аминирование в тканях
Образование

Основной путь обезвреживания аммиака в организме – биосинтез мочевины в
Основной путь обезвреживания аммиака в организме – биосинтез мочевины в
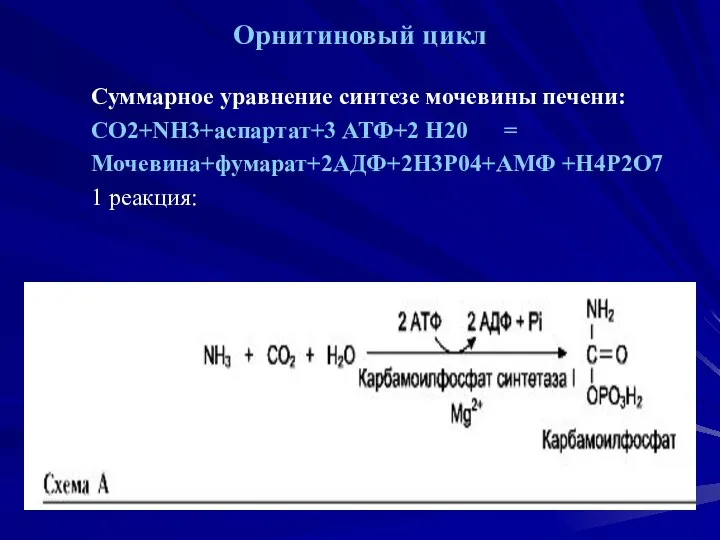
Орнитиновый цикл
Суммарное уравнение синтезе мочевины печени:
СО2+NH3+аспартат+3 АТФ+2 Н20 =
Мочевина+фумарат+2АДФ+2H3P04+АМФ +Н4Р2О7
1 реакция:
Орнитиновый цикл
Суммарное уравнение синтезе мочевины печени:
СО2+NH3+аспартат+3 АТФ+2 Н20 =
Мочевина+фумарат+2АДФ+2H3P04+АМФ +Н4Р2О7
1 реакция:

2-я реакция и 3 –я реакция
2-я реакция и 3 –я реакция

4-я реакция
4-я реакция
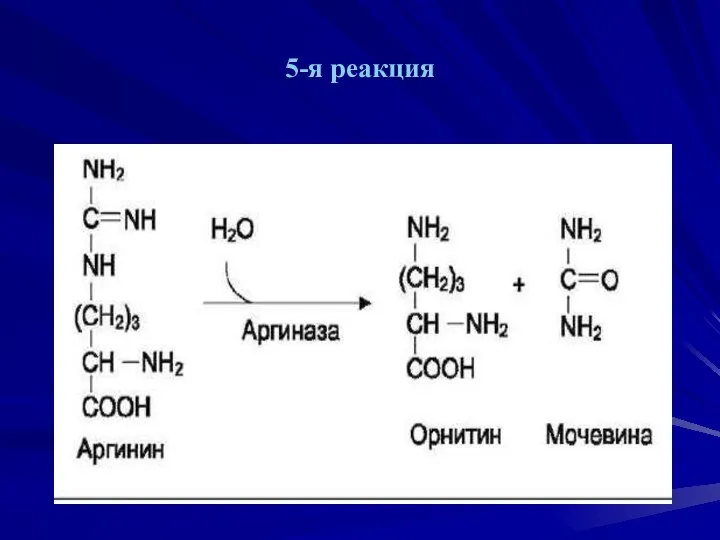
5-я реакция
5-я реакция
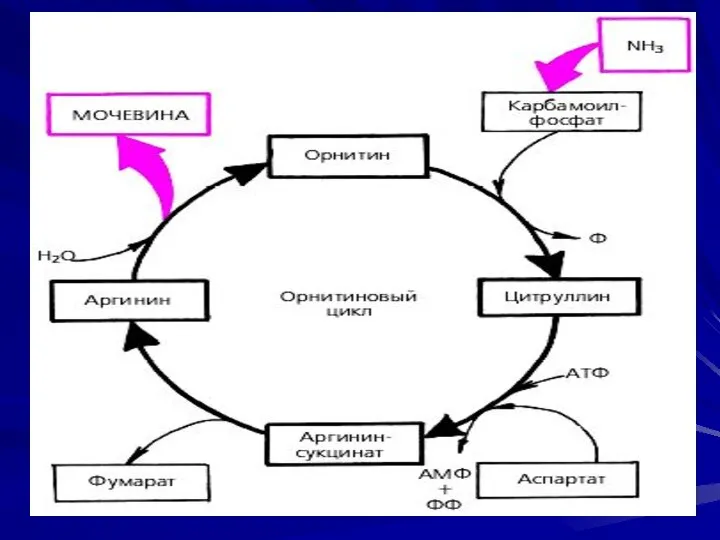

Энергетический баланс процесса
В реакциях орнитинового цикла расходуются четыре макроэргических связи
Энергетический баланс процесса
В реакциях орнитинового цикла расходуются четыре макроэргических связи

Затраты энергии происходят также и при трансмембранном переносе веществ, связанном с
Затраты энергии происходят также и при трансмембранном переносе веществ, связанном с

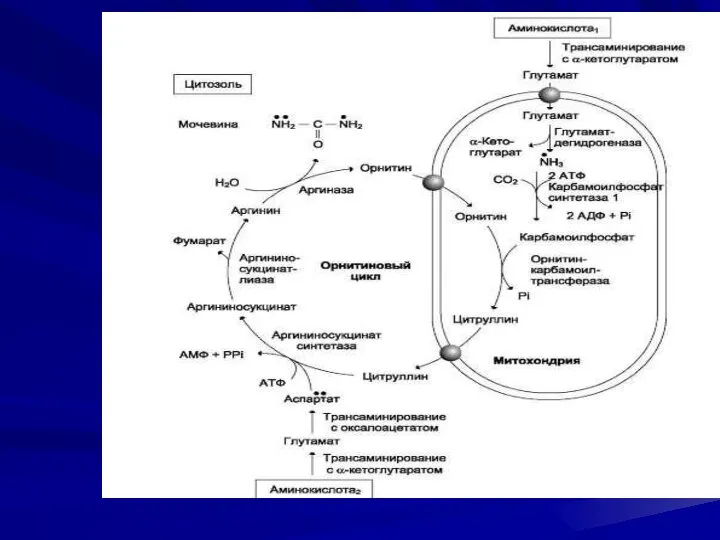
Ферменты орнитинового цикла распределены между митохондриями и цитозолем. Поэтому необходим трансмембранный
Ферменты орнитинового цикла распределены между митохондриями и цитозолем. Поэтому необходим трансмембранный

Биологическое значение орнитинового цикла
1. Обезвреживание аммиака в организме
2.Регуляция азотистого баланса в
Биологическое значение орнитинового цикла
1. Обезвреживание аммиака в организме
2.Регуляция азотистого баланса в

Источники азота , углерода и кислорода в цикле мочевины
1-я амминогруппа -
Источники азота , углерода и кислорода в цикле мочевины
1-я амминогруппа -

Гипераммониемия – повышение концентрации аммиака в крови.
Причины:
1.Врожденная недостаточность хотя бы
Гипераммониемия – повышение концентрации аммиака в крови.
Причины:
1.Врожденная недостаточность хотя бы
 ИНФОРМАЦИЯ И ЕЕ СВОЙСТВА Доцент кафедры информатики и информационных таможенных технологий Ю.И. Сомов
ИНФОРМАЦИЯ И ЕЕ СВОЙСТВА Доцент кафедры информатики и информационных таможенных технологий Ю.И. Сомов  История религий. Введение
История религий. Введение Фестиваль відкритих уроків
Фестиваль відкритих уроків Бетонные работы
Бетонные работы Схемы расположения земельных участков. Требования. Ошибки. Примеры
Схемы расположения земельных участков. Требования. Ошибки. Примеры Австралийский доллар
Австралийский доллар Ватикан
Ватикан Зиккурат в Уре Выполнил: …
Зиккурат в Уре Выполнил: …  Прийди з висот небесних
Прийди з висот небесних Indian English?
Indian English? Теория измерения культур Герта Хофстеда
Теория измерения культур Герта Хофстеда Елбасы туралы 1000 сұрақ
Елбасы туралы 1000 сұрақ Твердотельная электроника. Тиристоры
Твердотельная электроника. Тиристоры Компьютерная схемотехника
Компьютерная схемотехника Механизм воздействия реагентов на ПЗП
Механизм воздействия реагентов на ПЗП Предмет и научные основы системы государственного и муниципального управления. (Тема 1)
Предмет и научные основы системы государственного и муниципального управления. (Тема 1) Центр обеспечения развития образования 2010
Центр обеспечения развития образования 2010 Методические особенности физической подготовки футболистов
Методические особенности физической подготовки футболистов Профессионально-общественная аккредитация образовательных программ, основанных на профессиональных стандарта
Профессионально-общественная аккредитация образовательных программ, основанных на профессиональных стандарта Транспортная система страны
Транспортная система страны  Здравоохранение в системе рыночных отношений
Здравоохранение в системе рыночных отношений Безопасность детей в интернетe
Безопасность детей в интернетe Права и обязанности педагога ОУ НПО / СПО
Права и обязанности педагога ОУ НПО / СПО «Кто успевает в науках, «Кто успевает в науках, а отстает в добрых нравах, тот скорее отстает, чем успевает».
«Кто успевает в науках, «Кто успевает в науках, а отстает в добрых нравах, тот скорее отстает, чем успевает». «Монументальная скульптура. Выполнение рельефа (бумажная пластика) Учитель: Козлова В.В.
«Монументальная скульптура. Выполнение рельефа (бумажная пластика) Учитель: Козлова В.В. Актуальность. Применение ИКТ создает уникальную возможность для изучающих иностранный язык пользоваться аутентичными текстами,
Актуальность. Применение ИКТ создает уникальную возможность для изучающих иностранный язык пользоваться аутентичными текстами,  Шаблон по литературе
Шаблон по литературе Фиалковская Н.В. учитель начальных классов
Фиалковская Н.В. учитель начальных классов