- Транскрипция

Содержание
- 2. Центральная догма генетики. Генетическая информация закодирована в виде последовательности дезоксирибонуклеотидов на одной из двух цепей ДНК
- 3. Транскрипция ДНК → РНК Синтез РНК по матрице ДНК ферментом РНК-полимеразой Первый этап реализации генетической информации
- 4. Синтез РНК на ДНК-матрице – последовательность синтезированной мРНК комплементарна последовательности одной из цепей двойной спирали ДНК,
- 5. Подтверждение роли РНК как посредника между ДНК и белком: Как правило, ДНК в ядрах эукариотических клеток
- 6. Все это говорит о том, что генетическая информация, записанная в ДНК, передается на РНК-посредник (иРНК), с
- 7. Гены – транскрибируемые участки ДНК Транскрибируется не вся ДНК, а лишь отдельные ее участки – гены.
- 8. Строение гена Кодирующая часть АТГ STOP ДНК РНК-транскрипт Промотор Терминатор Точка начала транскрипции Окончание транскрипции 5'
- 9. Знаки начала и окончания матричных синтезов ДНК РНК белок транскрипция трансляция Знак начала Знак окончания Промотор
- 10. Принципы транскрипции Комплементарность Антипараллельность Униполярность Асимметричность
- 11. А Ц А Г Т Т Г А А Т Г Т Ц А А Ц
- 12. Общие параметры транскрипции Скорость – около 30 нуклеотидов / сек Частота ошибок – 1 на 104
- 13. Транскрипция генов в хромосоме Одна хромосома – одна молекула ДНК – около тысячи генов Матричной может
- 14. РНК-полимераза Чтобы доказать возможность синтеза РНК на ДНК-матрице нужно было обнаружить фермент, участвующий в таком синтезе.
- 15. Промоторы, связывание с ДНК-матрицей и σ-субъединица В результате транскрипции синтезируется одноцепочечная РНК, комплементарная фрагменту одной из
- 16. У прокариот, подобных E. coli, σ (sigma) субъединица RNA polymerase связывается с районом promoter на ДНК.
- 17. Промоторы разных генов слегка отличаются. Есть сильные и слабые промоторы. От промоторов зависит эффективность транскрипции. У
- 18. Важны два понятия 1) консенсусные последовательности ДНК – гомологичные различным генам одного организма или же одному
- 19. Инициация транскрипции и элонгация РНК РНК-полимераза катализирует введение молекул рибонуклеозид трифосфата в направлении 5 'к 3‘,
- 20. После инициации, σ (сигма) субъединица отделяется от холофермента, и продолжается удлинение цепи под руководством корового фермента,
- 21. После прохождения всего гена фермент встречает сигнал терминации. Терминальная последовательность длиной около 40 п.н. особенно важна
- 22. Гены, кодирующие близкородственные продукты, у бактерий часто сгруппированы на хромосоме. Часто эти прилегающие друг к другу
- 23. Транскрипция у эукариот Механизмы в основном сходны, но есть и различия. Основные: Транскрипция у эукариот происходит
- 24. Первичный РНК-транскрипт подвергается процессингу или созреванию – обычно к 5’ концу добавляется кэп, а к 3’
- 25. Инициация транскрипции у эукариот Эффективность начала транскрипции РНК-полимеразой определяется по меньшей мере тремя цис-активирующими элементами эукариотического
- 26. Один из таких цис-элементов – ТАТА-бокс или блок Голдберга-Хогнесса – был обнаружен в области промотора на
- 27. Второй цис-элемент – СААТ-бокс локализован в промоторах большинства генов, примерно на 80 нуклеотидов левее стартовой точки
- 28. Другой класс цис-активирующих элементов – энхансеры, которые локализуются на обоих концах (3’ и 5’) и внутри
- 29. Эукариотическая РНК-полимераза (RNP) существует в трех уникальных формах, каждая из которых транскрибирует различные типы генов. Каждый
- 30. Гетерогенные ядерные РНК и их процессинг – кэпы и хвосты Генетическая информация закодирована в матричной цепи
- 31. Первичная посттранскрипционная модификация эукариот РНК-транскриптов состоит в присоединении к 5’ концам молекул 7-метилгуанозина или (7mG)-кэпа. Кэп
- 32. Затем обнаружили, что гяРНК и иРНК имеют на 3’ конце последовательность из 250 А остатков. Эта
- 33. У эукариот, начальный транскрипт называется гетерогенной ядерной РНК (hnRNA), или пре-мРНК, содержащей некодирующие участки, называемые промежуточными
- 34. Поли-А "хвост" добавляется с разрезанному 3‘-концу. Некодирующие интроны удаляются. Кодирующие сегменты, называемые экзонами, соединеняются в процессе,
- 35. Интроны и прерывистые гены В 1977 г. обнаружено, что гены вирусов, инфицирующие животные клетки, содержат внутренние
- 36. Большинство эукариотических генов содержат интроны. Ген овальбумина кур в основном «молчащий", содержит семь интронов, которые вместе
- 37. Подавляющее большинство эукариотических генов содержит интроны, есть всего несколько исключений. Экзоны составляют 15% длины коллагенового гена,
- 38. Процессинг (созревание) пре-иРНК играет регуляторную роль в экспрессии генов. К примеру, некоторые интроны в молекуле пре-иРНК
- 39. Трансляция
- 40. Синтез белка по матрице и-РНК, осуществляемый на рибосомах Самый сложный из матричных синтезов – не просто
- 41. Нужны еще и молекулы-переводчики Матричный принцип в трансляции – генетический код
- 42. Участники трансляции Переводчики: 3. т-РНК с аминокислотой 4. Аминоацил-т-РНК-синтетазы (АРСазы) – ферменты, присоединяющие аминокислоту к т-РНК
- 43. Какие молекулы в клетке знают генетический код? Молекулы-переводчики
- 44. Гипотетический адаптор Адапторная гипотеза Крика аминокислота Участок связывания аминокислоты в адапторе Кодон данной аминокислоты в м-РНК
- 45. Транспортные РНК Молекула-адаптор. Один ее конец узнает кодон в м-РНК, а другой – несет аминокислоту. 3'
- 46. Сколько разных т-РНК ? Минимум – по числу аминокислот – 20 это реальное число пространственных форм
- 47. АРСазы аминоацил-тРНК-синтетазы Ферменты, присоединяющие аминокислоту к ее т-РНК Одна АРСаза узнает одну аминокислоту и все ее
- 48. Матричная РНК Кодирующая часть, транслируется 3' 5' Лидерная последовательность Шайна-Дальгарно БЕЛОК АУГ STOP 3‘ нетранслируемый район
- 49. Этапы трансляции Инициация (начало) Элонгация (удлинение) Терминация (окончание)
- 50. У бактерий в связывании участвует последовательность длиной до 6 рибонуклеотидов (АГГАГГ), которая предшествует стартовому кодону АУГ
- 51. м-РНК + малая субъединица рибосомы
- 52. Последовательность Шайна-Дальгарно Инициация
- 53. Элонгация
- 55. Терминация стоп
- 56. стоп
- 57. Параметры трансляции Самый медленный из матричных синтезов, максимальная скорость – 20 аминокислот в сек. Частота ошибок
- 58. Структура рибосом Структура рибосом хорошо изучена. В бактериальной клетке – около 10000 рибосом, в эукариотической -
- 59. Рибосомы прокариот состоит из большой и малой субъединиц. Обе субъединицы состоят из одной или более молекул
- 60. Эукариотические рибосомы структурно похожи на прокариотические, состоят из большой и малой субъединиц, каждая из которых состоит
- 61. Степеь избыточности генов, кодирующих рРНК. В геноме E coli 7 копий последовательности, кодирующей все 3 рРНК
- 62. Структура тРНК Наиболее хорошо изучены небольшие стабильные тРНК. Всего 75-90 нуклеотидов, молекулярная структура у про- и
- 63. 1965 г. – Р. Холли расшифровал последовательность тРНК аланина. Оказалось, что ряд нуклеотидов уникален только для
- 64. tRNA нуклеотиды могут содержать азотистые основания, модифицированные после транскрипции. Благодаря качанию, I (Inosine) и Im могут
- 65. Холли предложил двумерную модель молекулы тРНК в виде клеверного листа – несколько спаренных стеблей (за счет
- 66. Молекула тРНК содержит много модифицированных оснований и содержит ряд стеблей и петель, сложенных в 3-мерную конфигурацию.
- 67. Анализ структуры других тРНК выявил много общего. На 3’ конце у всех тРНК есть последовательность ССА-3.
- 68. Зарядка молекул тРНК Для успешной трансляции молекулы тРНК должны присоединить соответствующие аминокислоты. Такая зарядка тРНК происходит
- 69. «Зарядка» тРНК начинается, когда аминоацил-тРНК-синтетаза катализирует превращение аминокислоты в аминоациладениловую кислоту. Гидролизованная ATP отщепляет фосфат с
- 70. Инициация трансляции в E.coli затрагивает рибосомы, мРНК, энергоноситель GTP, несколько факторов инициации (IFs) и тРНК, которая
- 71. Инициация трансляции. Малая субъединица рибосомы связывается с несколькими факторами инициации (IF1, 2, 3); Этот комплекс связывается
- 72. Элонгация трансляции: шаг 1. Вторая заряженная tRNA вступает в A site, при содействии EF-Tu.
- 73. Элонгация трансляции: шаг 2. Пептидил трансфераза катализирует образование пептидной связи, которая связывает две аминокислоты. Незаряженная tRNA
- 74. Элонгация трансляции: шаг 3. Первая связь выполнена и первая незаряженная tRNA удаляется из E site, при
- 75. Элонгация трансляции: шаг 4. Третья заряженная tRNA вступает в A site, при содействии EF-Tu.
- 76. Элонгация трансляции: шаг 5. Вторая связь осуществлена, образовался трипептид который начинает проходить через туннель в большой
- 77. Элонгация трансляции: шаг 6. Целая полипептидная цепь синтезирована и освобождается от рибосомы, когда встречается один из
- 78. Терминация синтеза белка сигнализируется появлением стоп-кодона на A site: UAG, UAA, или UGA. Эти кодоны не
- 79. Полирибосомы После завершения элонгации часть молекулы мРНК проходит через рибосому и ассоциирует с малой субъединицей другой
- 80. иРНК Растущий полипетид старт Полисома
- 81. Трансляция у эукариот Основное различие между трансляцией у прокариот и эукариот состоит в том, что эукариотические
- 82. Инициация трансляции у эукариот имеет свои особенности. За счет появления на 5’ конце зрелой мРНК кэпа
- 84. Скачать презентацию
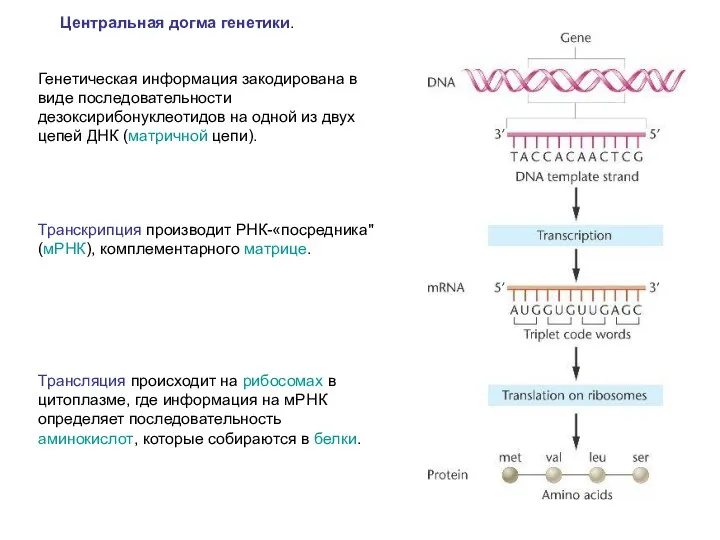
Центральная догма генетики.
Генетическая информация закодирована в виде последовательности дезоксирибонуклеотидов на
Центральная догма генетики.
Генетическая информация закодирована в виде последовательности дезоксирибонуклеотидов на

Транскрипция ДНК → РНК
Синтез РНК по матрице ДНК ферментом РНК-полимеразой
Первый этап
Транскрипция ДНК → РНК
Синтез РНК по матрице ДНК ферментом РНК-полимеразой
Первый этап

Синтез РНК на ДНК-матрице – последовательность синтезированной мРНК комплементарна последовательности одной
Синтез РНК на ДНК-матрице – последовательность синтезированной мРНК комплементарна последовательности одной

Подтверждение роли РНК как посредника между ДНК и белком:
Как правило, ДНК
Подтверждение роли РНК как посредника между ДНК и белком:
Как правило, ДНК

Все это говорит о том, что генетическая информация, записанная в ДНК,
Все это говорит о том, что генетическая информация, записанная в ДНК,
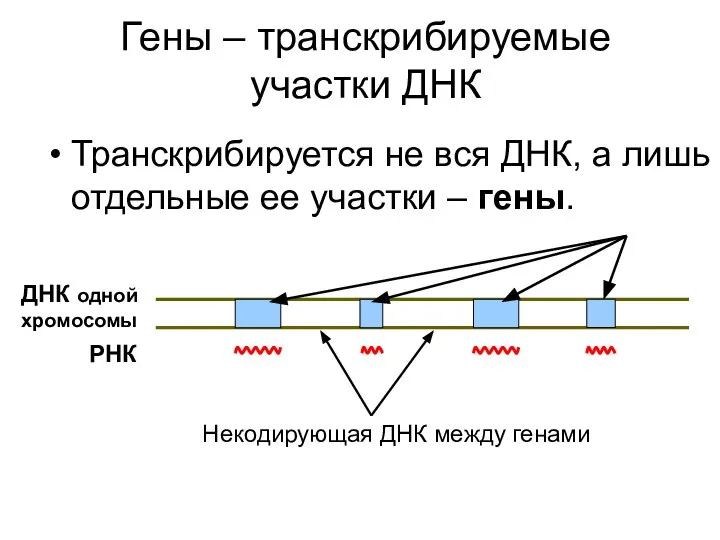
Гены – транскрибируемые участки ДНК
Транскрибируется не вся ДНК, а лишь отдельные
Гены – транскрибируемые участки ДНК
Транскрибируется не вся ДНК, а лишь отдельные
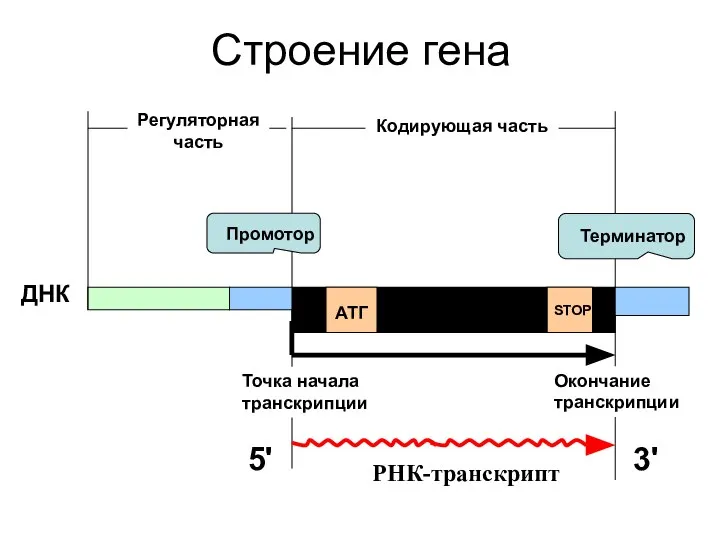
Строение гена
Кодирующая часть
АТГ
STOP
ДНК
РНК-транскрипт
Промотор
Терминатор
Точка начала транскрипции
Окончание транскрипции
5'
3'
Регуляторная часть
Строение гена
Кодирующая часть
АТГ
STOP
ДНК
РНК-транскрипт
Промотор
Терминатор
Точка начала транскрипции
Окончание транскрипции
5'
3'
Регуляторная часть
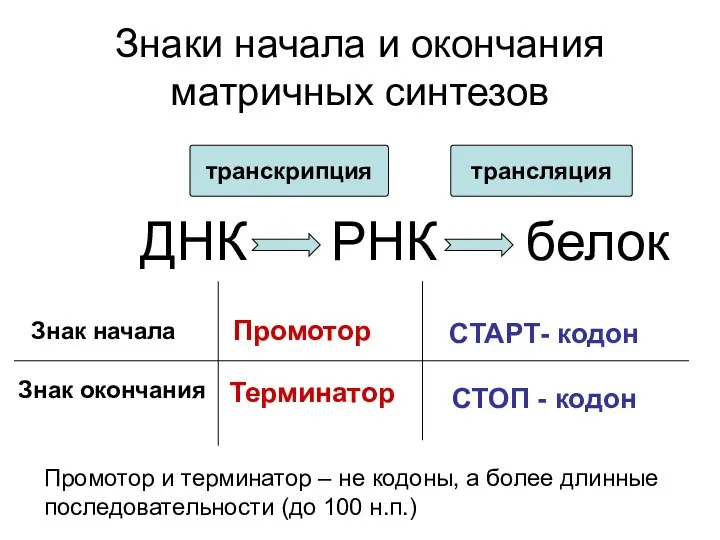
Знаки начала и окончания матричных синтезов
ДНК
РНК
белок
транскрипция
трансляция
Знак начала
Знак окончания
Промотор
Терминатор
СТАРТ- кодон
СТОП -
Знаки начала и окончания матричных синтезов
ДНК
РНК
белок
транскрипция
трансляция
Знак начала
Знак окончания
Промотор
Терминатор
СТАРТ- кодон
СТОП -

Принципы транскрипции
Комплементарность
Антипараллельность
Униполярность
Асимметричность
Принципы транскрипции
Комплементарность
Антипараллельность
Униполярность
Асимметричность
А
Ц
А
Г
Т
Т
Г
А
А
Т
Г
Т
Ц
А
А
Ц
Т
Т
У
Г
У
Ц
А
А
Ц
У
У
ДНК
ДНК
3'
5'
3'
5'
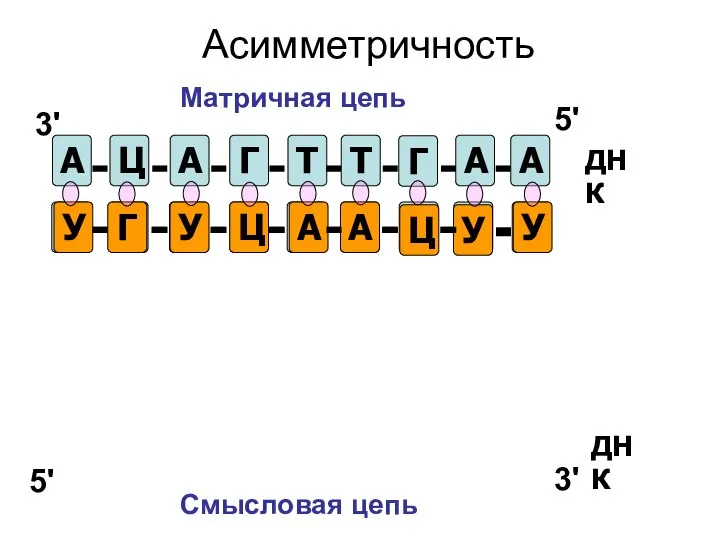
Асимметричность
Матричная цепь
Смысловая цепь
А
Ц
А
Г
Т
Т
Г
А
А
Т
Г
Т
Ц
А
А
Ц
Т
Т
У
Г
У
Ц
А
А
Ц
У
У
ДНК
ДНК
3'
5'
3'
5'
Асимметричность
Матричная цепь
Смысловая цепь

Общие параметры транскрипции
Скорость – около 30 нуклеотидов / сек
Частота ошибок –
Общие параметры транскрипции
Скорость – около 30 нуклеотидов / сек
Частота ошибок –
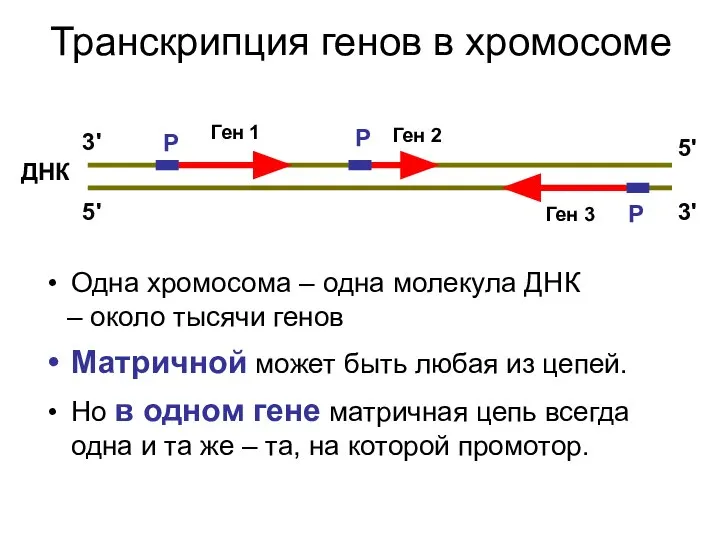
Транскрипция генов в хромосоме
Одна хромосома – одна молекула ДНК
– около
Транскрипция генов в хромосоме
Одна хромосома – одна молекула ДНК
– около

РНК-полимераза
Чтобы доказать возможность синтеза РНК на ДНК-матрице нужно было обнаружить фермент,
РНК-полимераза
Чтобы доказать возможность синтеза РНК на ДНК-матрице нужно было обнаружить фермент,

Промоторы, связывание с ДНК-матрицей и σ-субъединица
В результате транскрипции синтезируется одноцепочечная РНК,
Промоторы, связывание с ДНК-матрицей и σ-субъединица
В результате транскрипции синтезируется одноцепочечная РНК,
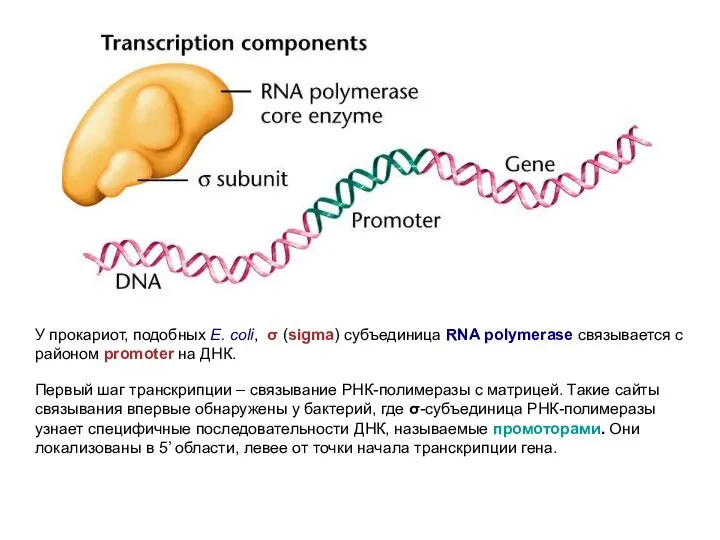
У прокариот, подобных E. coli, σ (sigma) субъединица RNA polymerase связывается
У прокариот, подобных E. coli, σ (sigma) субъединица RNA polymerase связывается

Промоторы разных генов слегка отличаются.
Есть сильные и слабые промоторы.
От промоторов зависит
Промоторы разных генов слегка отличаются.
Есть сильные и слабые промоторы.
От промоторов зависит

Важны два понятия
1) консенсусные последовательности ДНК – гомологичные различным генам
Важны два понятия
1) консенсусные последовательности ДНК – гомологичные различным генам
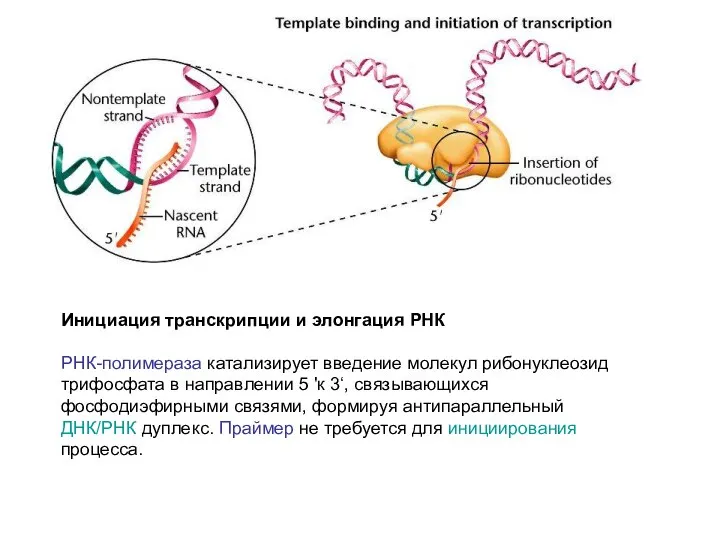
Инициация транскрипции и элонгация РНК
РНК-полимераза катализирует введение молекул рибонуклеозид трифосфата в
Инициация транскрипции и элонгация РНК
РНК-полимераза катализирует введение молекул рибонуклеозид трифосфата в
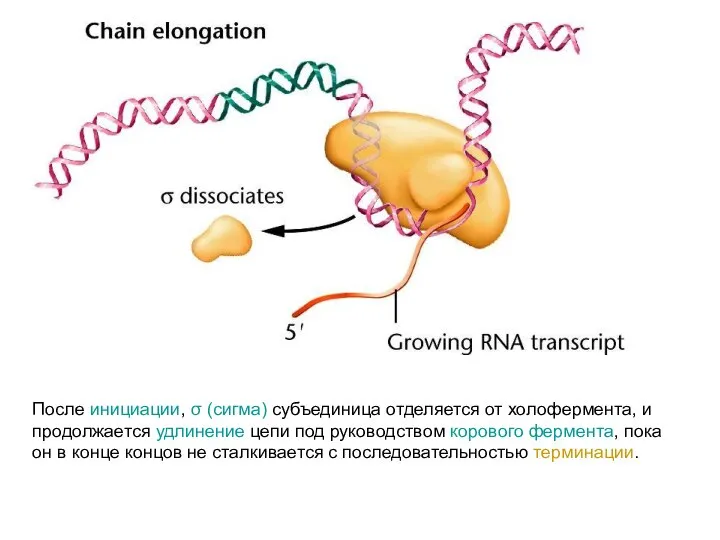
После инициации, σ (сигма) субъединица отделяется от холофермента, и продолжается удлинение
После инициации, σ (сигма) субъединица отделяется от холофермента, и продолжается удлинение

После прохождения всего гена фермент встречает сигнал терминации. Терминальная последовательность длиной
После прохождения всего гена фермент встречает сигнал терминации. Терминальная последовательность длиной

Гены, кодирующие близкородственные продукты, у бактерий часто сгруппированы на хромосоме. Часто
Гены, кодирующие близкородственные продукты, у бактерий часто сгруппированы на хромосоме. Часто

Транскрипция у эукариот
Механизмы в основном сходны, но есть и различия. Основные:
Транскрипция
Транскрипция у эукариот
Механизмы в основном сходны, но есть и различия. Основные:
Транскрипция

Первичный РНК-транскрипт подвергается процессингу или созреванию – обычно к 5’ концу
Первичный РНК-транскрипт подвергается процессингу или созреванию – обычно к 5’ концу

Инициация транскрипции у эукариот
Эффективность начала транскрипции РНК-полимеразой определяется по меньшей мере
Инициация транскрипции у эукариот
Эффективность начала транскрипции РНК-полимеразой определяется по меньшей мере

Один из таких цис-элементов – ТАТА-бокс или блок Голдберга-Хогнесса – был
Один из таких цис-элементов – ТАТА-бокс или блок Голдберга-Хогнесса – был

Второй цис-элемент – СААТ-бокс локализован в промоторах большинства генов, примерно на
Второй цис-элемент – СААТ-бокс локализован в промоторах большинства генов, примерно на

Другой класс цис-активирующих элементов – энхансеры, которые локализуются на обоих концах
Другой класс цис-активирующих элементов – энхансеры, которые локализуются на обоих концах

Эукариотическая РНК-полимераза (RNP) существует в трех уникальных формах, каждая из которых
Эукариотическая РНК-полимераза (RNP) существует в трех уникальных формах, каждая из которых

Гетерогенные ядерные РНК и их процессинг – кэпы и хвосты
Генетическая информация
Гетерогенные ядерные РНК и их процессинг – кэпы и хвосты
Генетическая информация

Первичная посттранскрипционная модификация эукариот РНК-транскриптов состоит в присоединении к 5’ концам
Первичная посттранскрипционная модификация эукариот РНК-транскриптов состоит в присоединении к 5’ концам

Затем обнаружили, что гяРНК и иРНК имеют на 3’ конце последовательность
Затем обнаружили, что гяРНК и иРНК имеют на 3’ конце последовательность
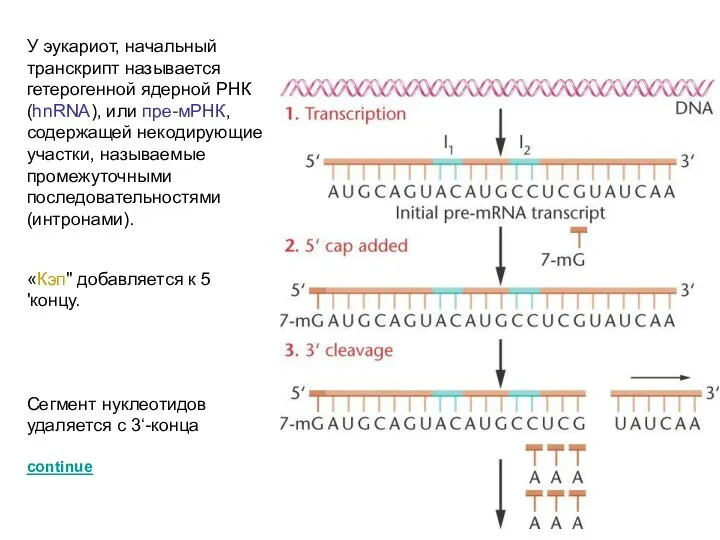
У эукариот, начальный транскрипт называется гетерогенной ядерной РНК (hnRNA), или пре-мРНК,
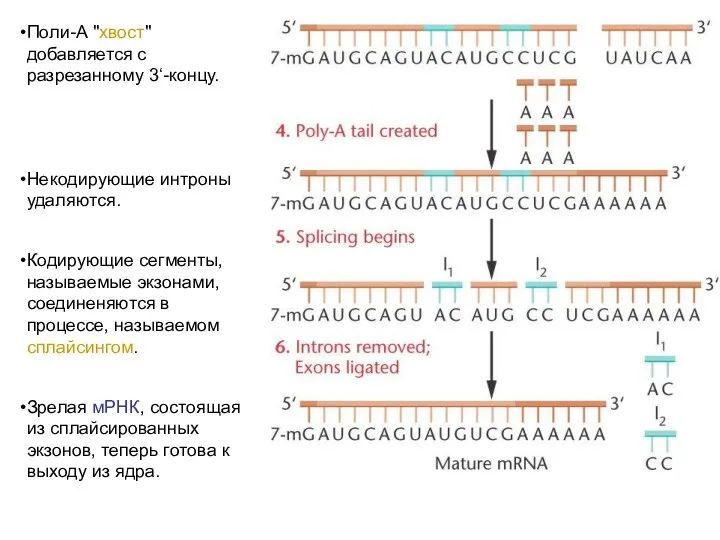
Поли-А "хвост" добавляется с разрезанному 3‘-концу.
Некодирующие интроны удаляются.
Кодирующие сегменты, называемые экзонами,
Поли-А "хвост" добавляется с разрезанному 3‘-концу.
Некодирующие интроны удаляются.
Кодирующие сегменты, называемые экзонами,

Интроны и прерывистые гены
В 1977 г. обнаружено, что гены вирусов, инфицирующие
Интроны и прерывистые гены
В 1977 г. обнаружено, что гены вирусов, инфицирующие
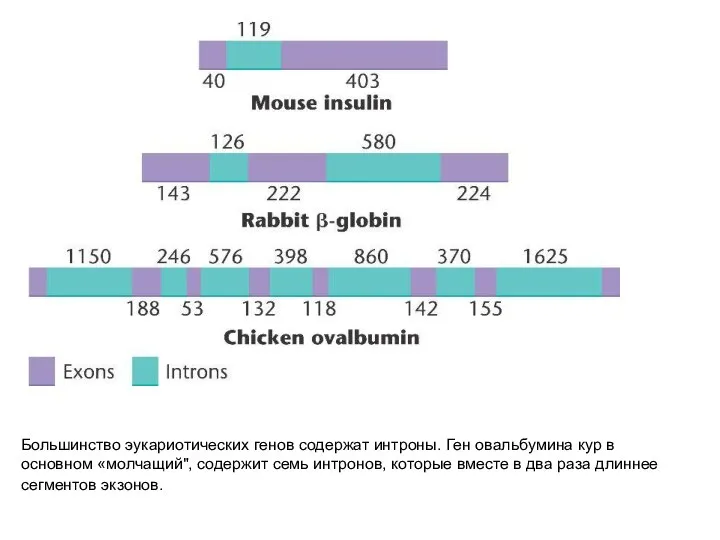
Большинство эукариотических генов содержат интроны. Ген овальбумина кур в основном «молчащий",
Большинство эукариотических генов содержат интроны. Ген овальбумина кур в основном «молчащий",
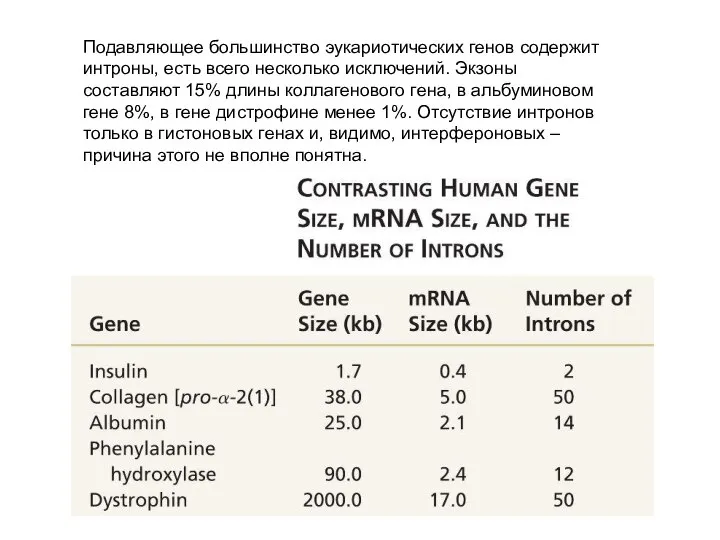
Подавляющее большинство эукариотических генов содержит интроны, есть всего несколько исключений. Экзоны
Подавляющее большинство эукариотических генов содержит интроны, есть всего несколько исключений. Экзоны

Процессинг (созревание) пре-иРНК играет регуляторную роль в экспрессии генов. К примеру,
Процессинг (созревание) пре-иРНК играет регуляторную роль в экспрессии генов. К примеру,

Трансляция
Трансляция

Синтез белка по матрице и-РНК, осуществляемый на рибосомах
Самый сложный из матричных
Синтез белка по матрице и-РНК, осуществляемый на рибосомах
Самый сложный из матричных

Нужны еще и
молекулы-переводчики
Матричный принцип в трансляции – генетический код
Нужны еще и
молекулы-переводчики
Матричный принцип в трансляции – генетический код

Участники трансляции Переводчики:
3. т-РНК с аминокислотой
4. Аминоацил-т-РНК-синтетазы (АРСазы) – ферменты, присоединяющие
Участники трансляции Переводчики:
3. т-РНК с аминокислотой
4. Аминоацил-т-РНК-синтетазы (АРСазы) – ферменты, присоединяющие

Какие молекулы в клетке знают генетический код?
Молекулы-переводчики
Какие молекулы в клетке знают генетический код?
Молекулы-переводчики
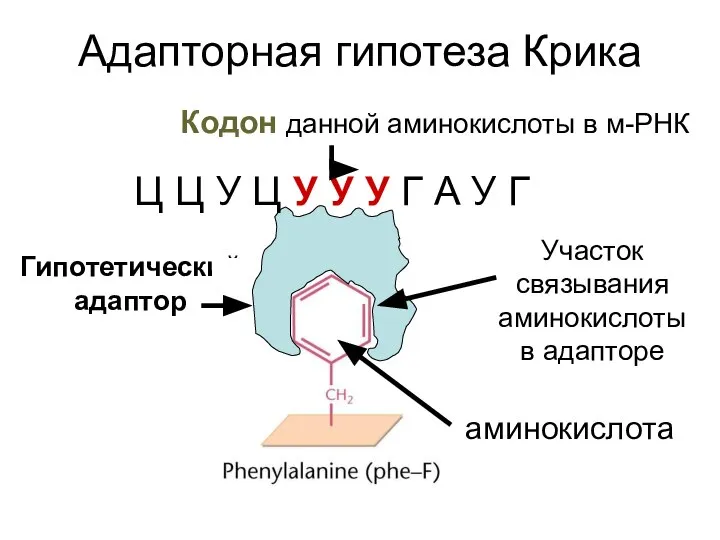
Гипотетический
адаптор
Адапторная гипотеза Крика
аминокислота
Участок
связывания
аминокислоты в адапторе
Кодон данной аминокислоты в м-РНК
Гипотетический
адаптор
Адапторная гипотеза Крика
аминокислота
Участок
связывания
аминокислоты в адапторе
Кодон данной аминокислоты в м-РНК
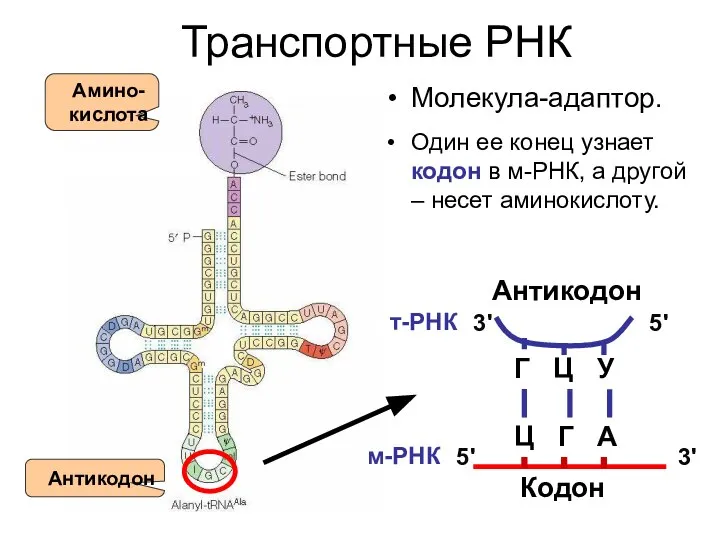
Транспортные РНК
Молекула-адаптор.
Один ее конец узнает кодон в м-РНК, а другой –
Транспортные РНК
Молекула-адаптор.
Один ее конец узнает кодон в м-РНК, а другой –

Сколько разных т-РНК ?
Минимум – по числу аминокислот – 20
Сколько разных т-РНК ?
Минимум – по числу аминокислот – 20

АРСазы
аминоацил-тРНК-синтетазы
Ферменты, присоединяющие аминокислоту к ее т-РНК
Одна АРСаза узнает одну аминокислоту
АРСазы
аминоацил-тРНК-синтетазы
Ферменты, присоединяющие аминокислоту к ее т-РНК
Одна АРСаза узнает одну аминокислоту
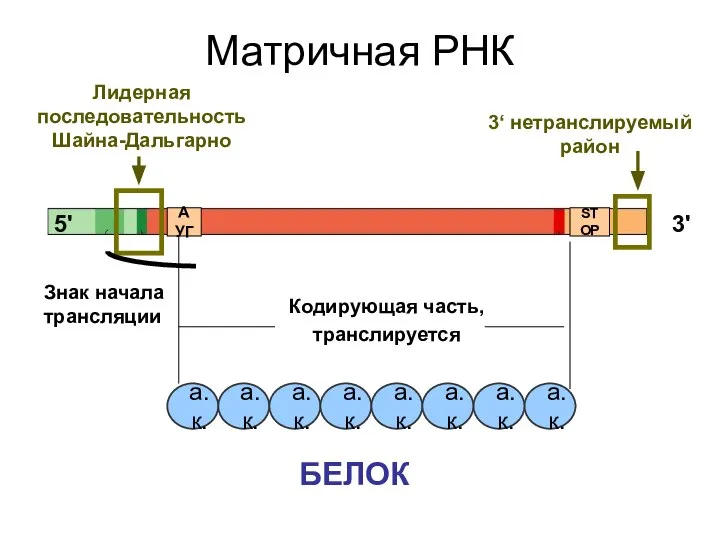
Матричная РНК
Кодирующая часть,
транслируется
3'
5'
Лидерная последовательность Шайна-Дальгарно
БЕЛОК
АУГ
STOP
3‘ нетранслируемый район
Знак начала трансляции
Матричная РНК
Кодирующая часть,
транслируется
3'
5'
Лидерная последовательность Шайна-Дальгарно
БЕЛОК
АУГ
STOP
3‘ нетранслируемый район
Знак начала трансляции

Этапы трансляции
Инициация (начало)
Элонгация (удлинение)
Терминация (окончание)
Этапы трансляции
Инициация (начало)
Элонгация (удлинение)
Терминация (окончание)

У бактерий в связывании участвует последовательность длиной до 6 рибонуклеотидов (АГГАГГ),
У бактерий в связывании участвует последовательность длиной до 6 рибонуклеотидов (АГГАГГ),
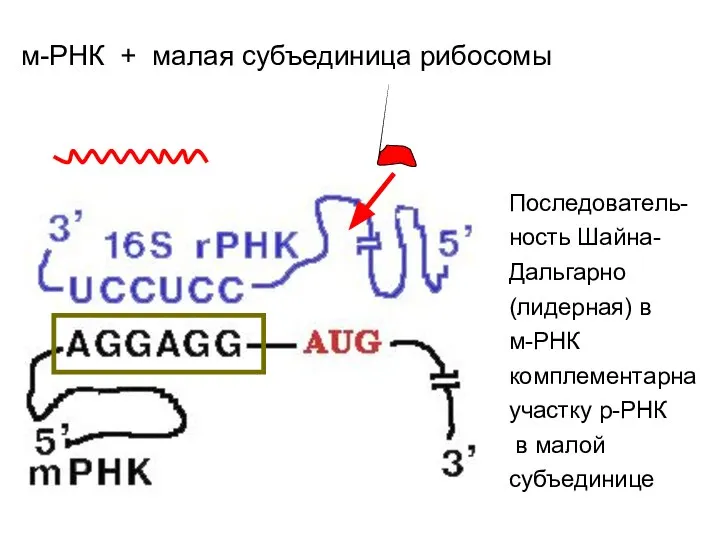
м-РНК + малая субъединица рибосомы
м-РНК + малая субъединица рибосомы
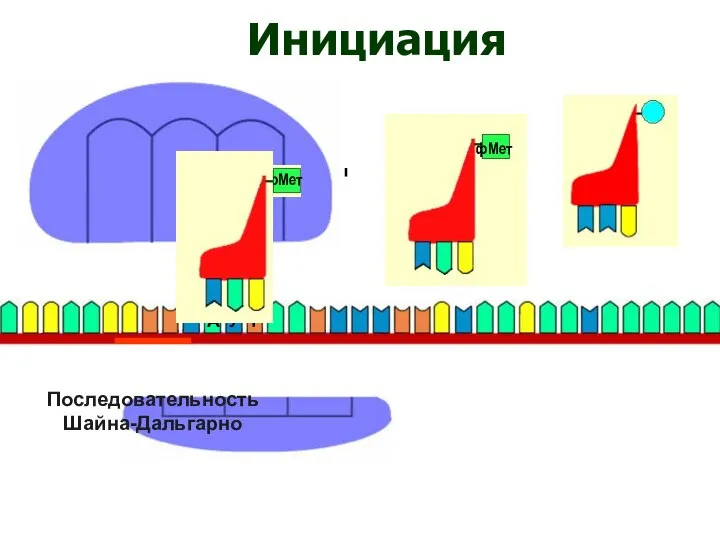
Последовательность
Шайна-Дальгарно
Инициация
Последовательность
Шайна-Дальгарно
Инициация
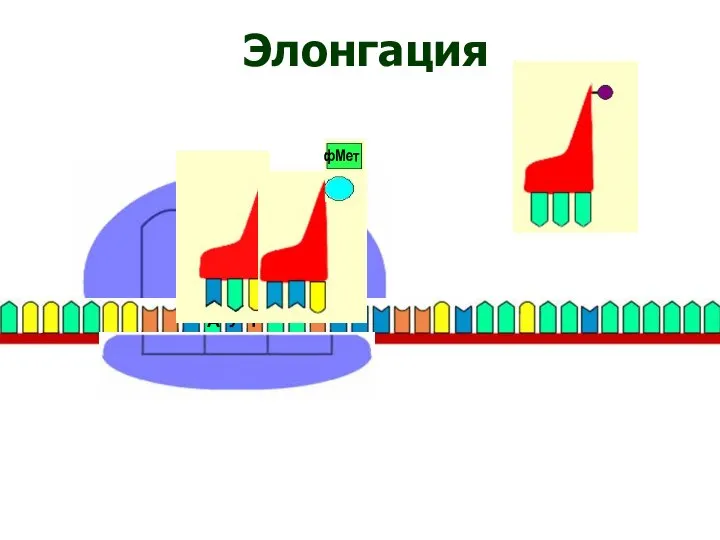
Элонгация
Элонгация
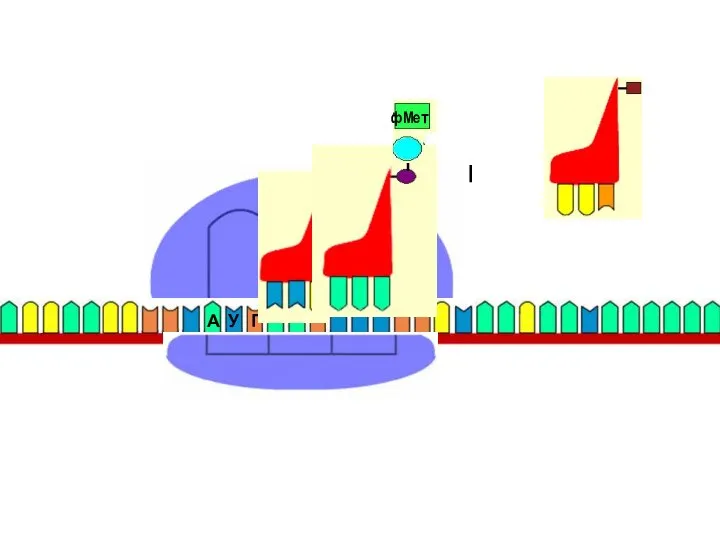
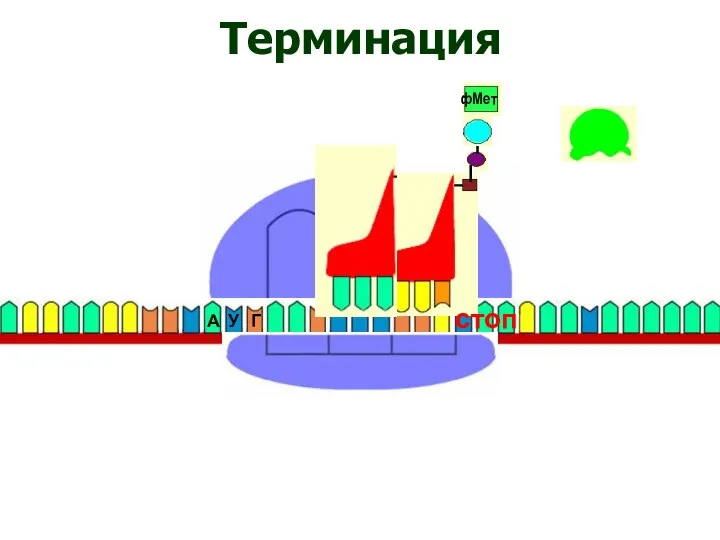
Терминация
стоп
Терминация
стоп
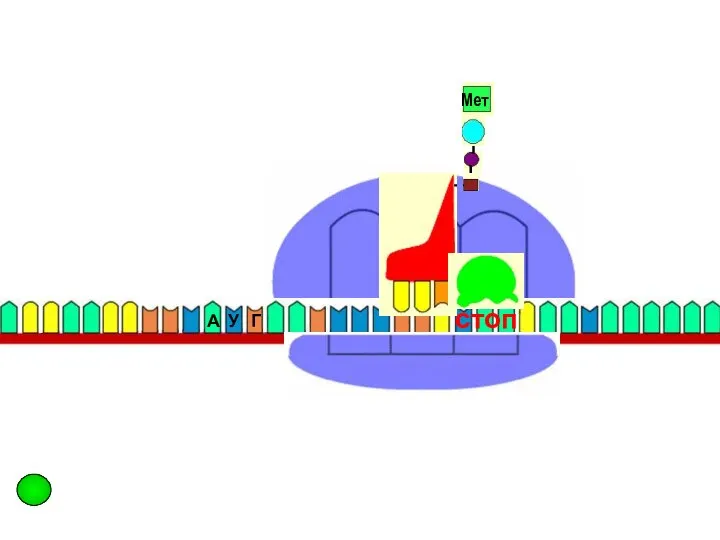
стоп
стоп

Параметры трансляции
Самый медленный из матричных синтезов, максимальная скорость –
20
Параметры трансляции
Самый медленный из матричных синтезов, максимальная скорость –
20

Структура рибосом
Структура рибосом хорошо изучена. В бактериальной клетке – около 10000
Структура рибосом
Структура рибосом хорошо изучена. В бактериальной клетке – около 10000
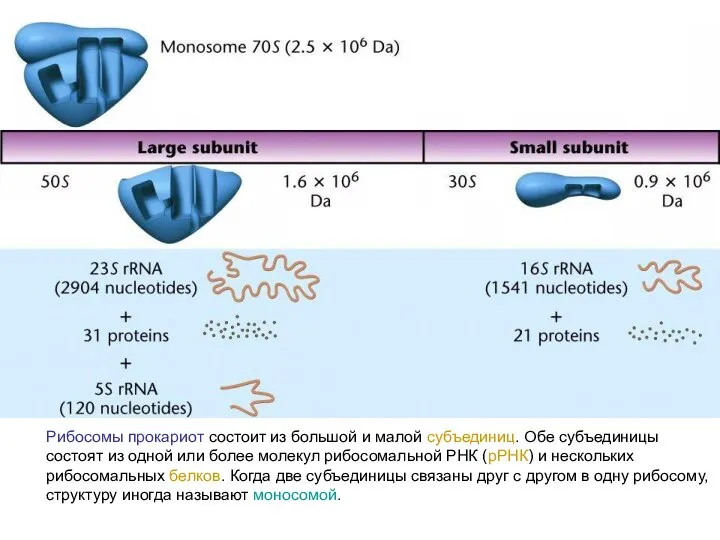
Рибосомы прокариот состоит из большой и малой субъединиц. Обе субъединицы состоят
Рибосомы прокариот состоит из большой и малой субъединиц. Обе субъединицы состоят
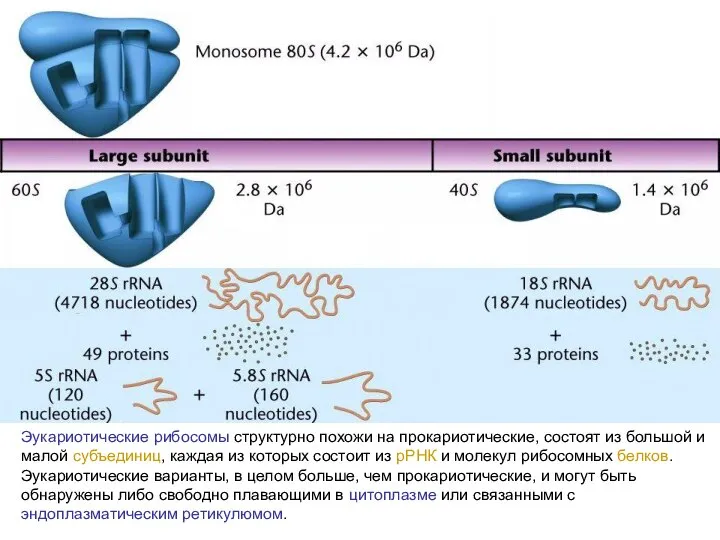
Эукариотические рибосомы структурно похожи на прокариотические, состоят из большой и малой
Эукариотические рибосомы структурно похожи на прокариотические, состоят из большой и малой

Степеь избыточности генов, кодирующих рРНК.
В геноме E coli 7
Степеь избыточности генов, кодирующих рРНК.
В геноме E coli 7

Структура тРНК
Наиболее хорошо изучены небольшие стабильные тРНК. Всего 75-90 нуклеотидов, молекулярная
Структура тРНК
Наиболее хорошо изучены небольшие стабильные тРНК. Всего 75-90 нуклеотидов, молекулярная

1965 г. – Р. Холли расшифровал последовательность тРНК аланина. Оказалось, что
1965 г. – Р. Холли расшифровал последовательность тРНК аланина. Оказалось, что
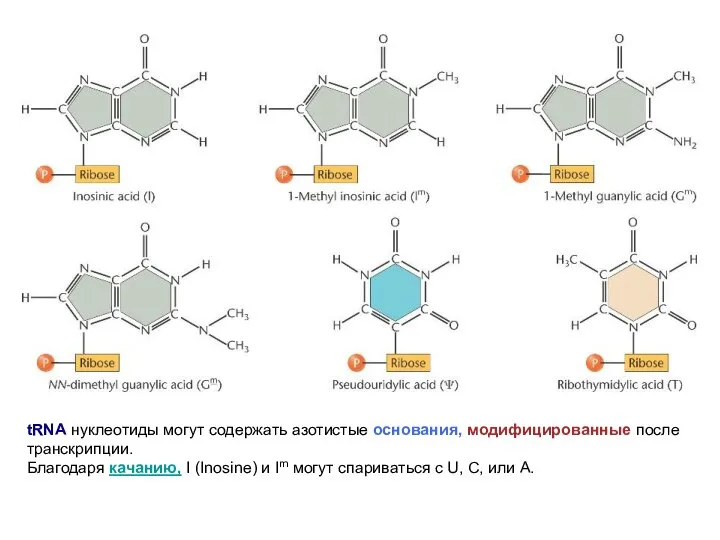
tRNA нуклеотиды могут содержать азотистые основания, модифицированные после транскрипции.
Благодаря качанию,
tRNA нуклеотиды могут содержать азотистые основания, модифицированные после транскрипции.
Благодаря качанию,

Холли предложил двумерную модель молекулы тРНК в виде клеверного листа –
Холли предложил двумерную модель молекулы тРНК в виде клеверного листа –
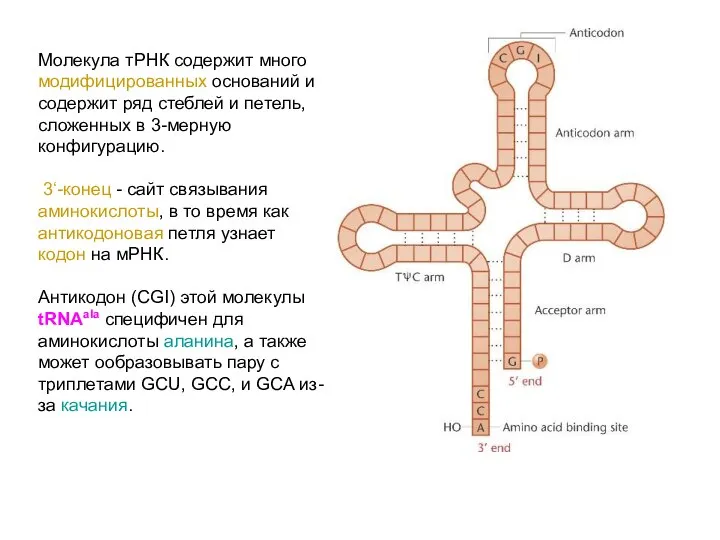
Молекула тРНК содержит много модифицированных оснований и содержит ряд стеблей и

Анализ структуры других тРНК выявил много общего.
На 3’ конце у
Анализ структуры других тРНК выявил много общего.
На 3’ конце у

Зарядка молекул тРНК
Для успешной трансляции молекулы тРНК должны присоединить соответствующие аминокислоты.
Зарядка молекул тРНК
Для успешной трансляции молекулы тРНК должны присоединить соответствующие аминокислоты.
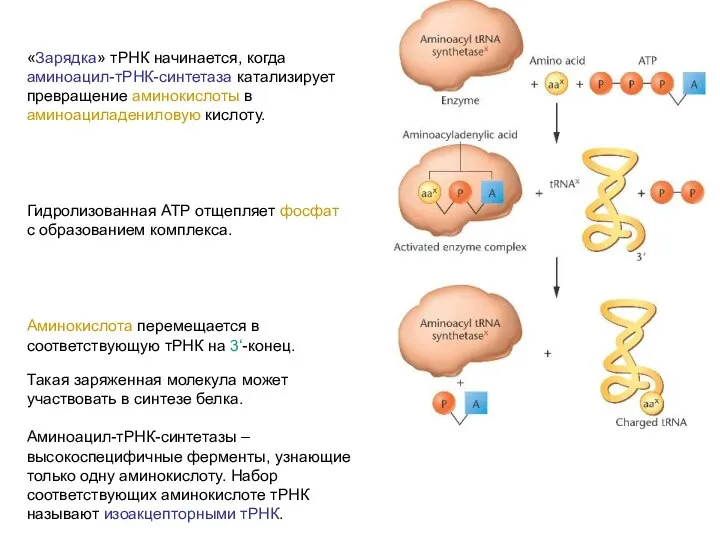
«Зарядка» тРНК начинается, когда аминоацил-тРНК-синтетаза катализирует превращение аминокислоты в аминоациладениловую кислоту.
«Зарядка» тРНК начинается, когда аминоацил-тРНК-синтетаза катализирует превращение аминокислоты в аминоациладениловую кислоту.
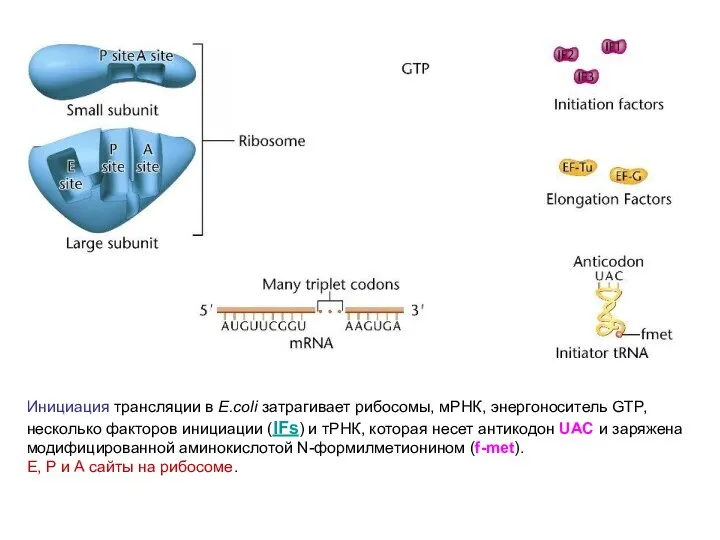
Инициация трансляции в E.coli затрагивает рибосомы, мРНК, энергоноситель GTP, несколько факторов
Инициация трансляции в E.coli затрагивает рибосомы, мРНК, энергоноситель GTP, несколько факторов
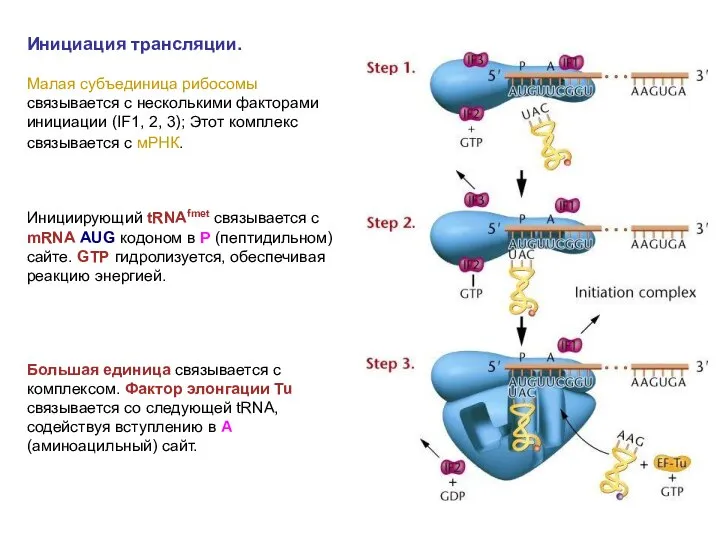
Инициация трансляции.
Малая субъединица рибосомы связывается с несколькими факторами инициации (IF1, 2,
Инициация трансляции. Малая субъединица рибосомы связывается с несколькими факторами инициации (IF1, 2,
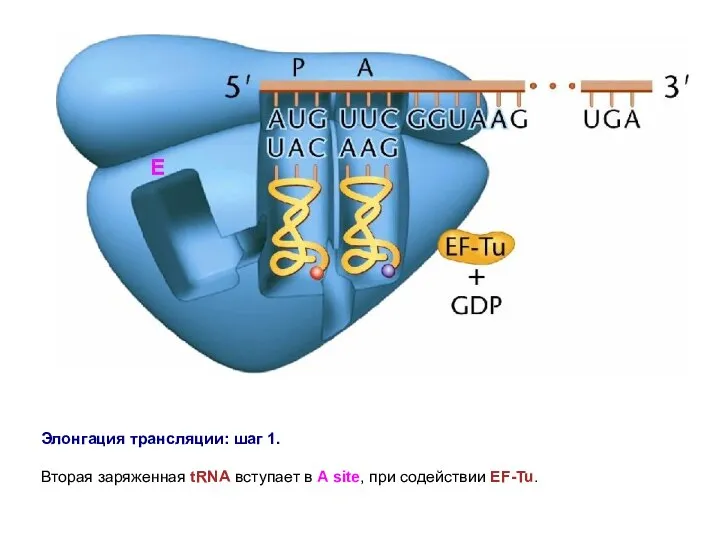
Элонгация трансляции: шаг 1.
Вторая заряженная tRNA вступает в A site,
Элонгация трансляции: шаг 1.
Вторая заряженная tRNA вступает в A site,
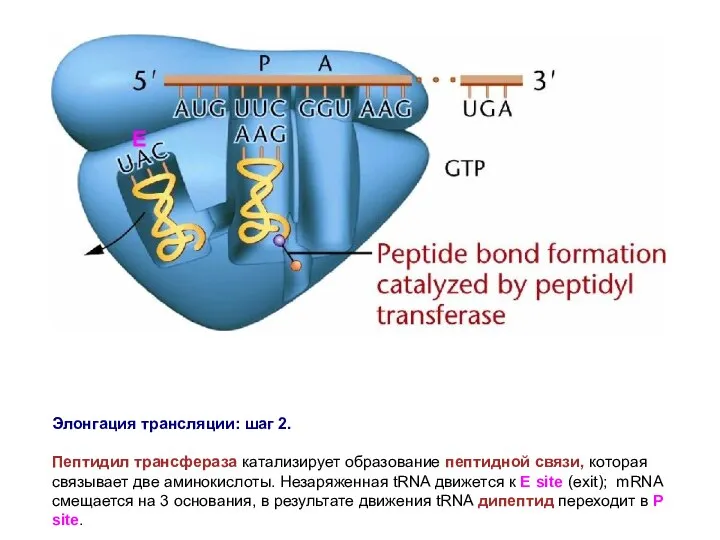
Элонгация трансляции: шаг 2.
Пептидил трансфераза катализирует образование пептидной связи, которая
Элонгация трансляции: шаг 2.
Пептидил трансфераза катализирует образование пептидной связи, которая
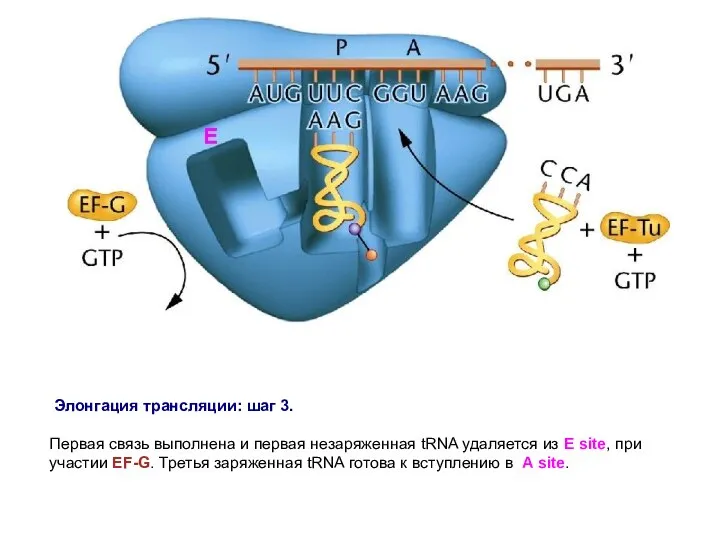
Элонгация трансляции: шаг 3.
Первая связь выполнена и первая незаряженная
Элонгация трансляции: шаг 3.
Первая связь выполнена и первая незаряженная
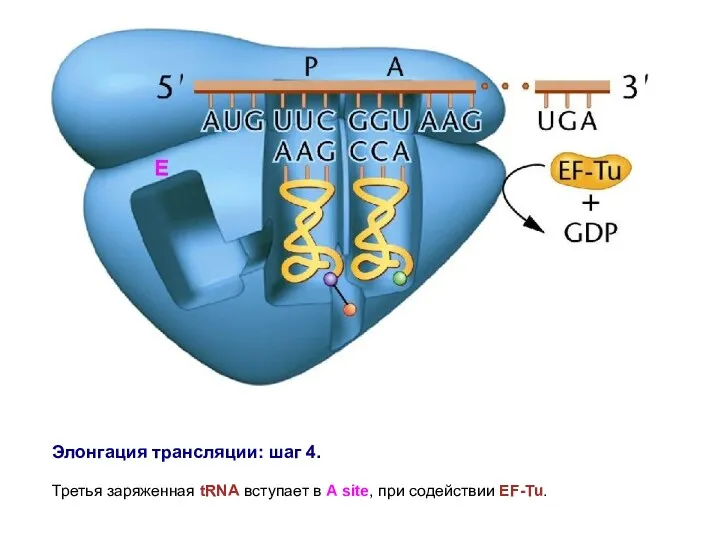
Элонгация трансляции: шаг 4.
Третья заряженная tRNA вступает в A site,
Элонгация трансляции: шаг 4.
Третья заряженная tRNA вступает в A site,
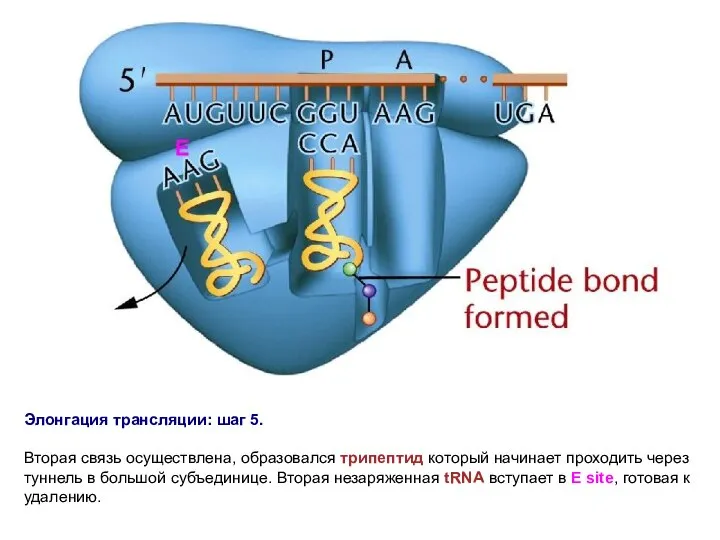
Элонгация трансляции: шаг 5.
Вторая связь осуществлена, образовался трипептид который начинает
Элонгация трансляции: шаг 5.
Вторая связь осуществлена, образовался трипептид который начинает
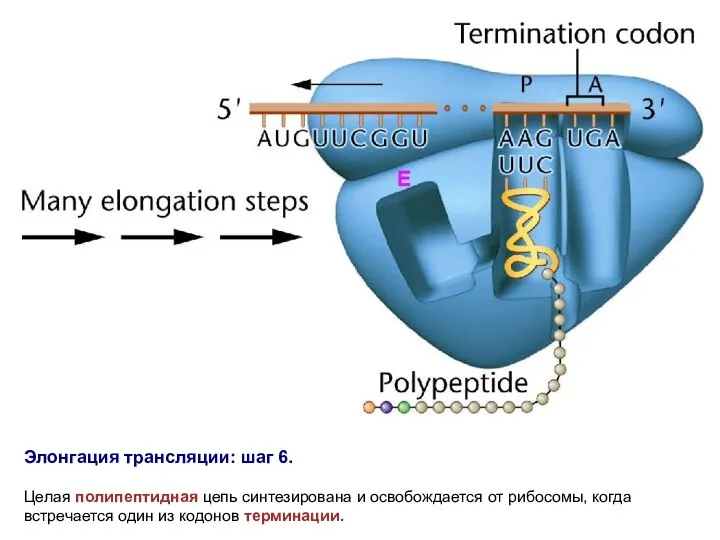
Элонгация трансляции: шаг 6.
Целая полипептидная цепь синтезирована и освобождается от
Элонгация трансляции: шаг 6.
Целая полипептидная цепь синтезирована и освобождается от
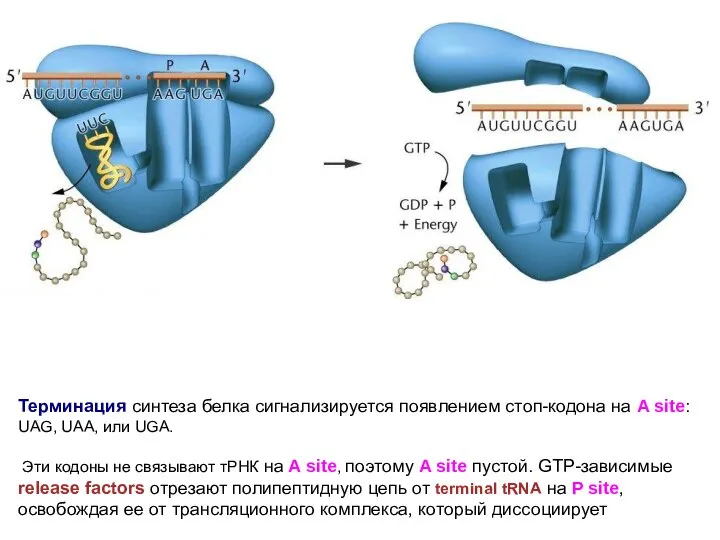
Терминация синтеза белка сигнализируется появлением стоп-кодона на A site: UAG, UAA,
Терминация синтеза белка сигнализируется появлением стоп-кодона на A site: UAG, UAA,

Полирибосомы
После завершения элонгации часть молекулы мРНК проходит через рибосому и ассоциирует
Полирибосомы
После завершения элонгации часть молекулы мРНК проходит через рибосому и ассоциирует
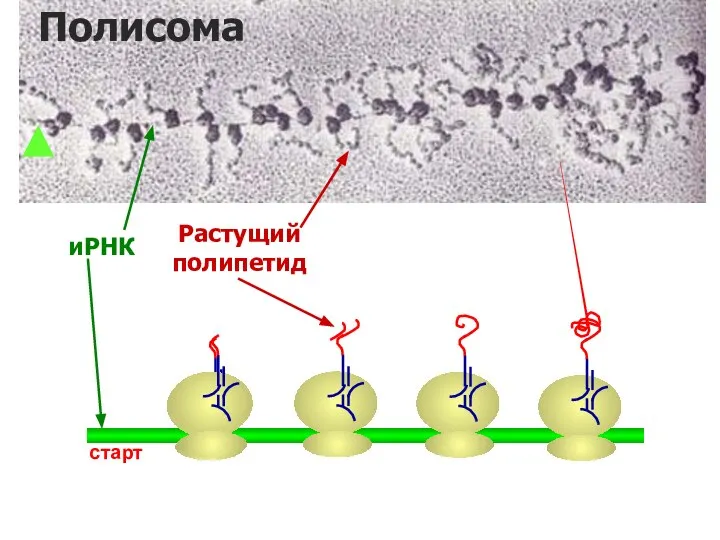
иРНК
Растущий
полипетид
старт
Полисома
иРНК
Растущий
полипетид
старт
Полисома

Трансляция у эукариот
Основное различие между трансляцией у прокариот и эукариот состоит
Трансляция у эукариот
Основное различие между трансляцией у прокариот и эукариот состоит

Инициация трансляции у эукариот имеет свои особенности. За счет появления на
Инициация трансляции у эукариот имеет свои особенности. За счет появления на
 Презентация "Оборудование" - скачать презентации по Экономике
Презентация "Оборудование" - скачать презентации по Экономике Финансы и кредит Тема 6. Внебюджетные фонды
Финансы и кредит Тема 6. Внебюджетные фонды Укажите функцию для получения общего объема памяти на диске
Укажите функцию для получения общего объема памяти на диске Законотворческий процесс в РФ
Законотворческий процесс в РФ Субъекты коллективных трудовых отношений
Субъекты коллективных трудовых отношений Архитектура процессора i8086
Архитектура процессора i8086 Поверхности. Лекция №4
Поверхности. Лекция №4 Презентация "Теория маркетинга" - скачать презентации по Экономике
Презентация "Теория маркетинга" - скачать презентации по Экономике Обучающий семинар для тьюторов проекта «Школьный стандарт профилактики» Ведущие: Екатерина Львовна Лашевская, начальник инфор
Обучающий семинар для тьюторов проекта «Школьный стандарт профилактики» Ведущие: Екатерина Львовна Лашевская, начальник инфор Я співаю, як спаситель
Я співаю, як спаситель характеристика й обробка ГРИБІВ
характеристика й обробка ГРИБІВ Презентация Политическая мысль России
Презентация Политическая мысль России Тәуелсіздік стратегиясы
Тәуелсіздік стратегиясы Банковская система,ее структура и функции Презентацию подготовила: Чудакова Кристина 11 «А» класс
Банковская система,ее структура и функции Презентацию подготовила: Чудакова Кристина 11 «А» класс  Қазақстан Республикасының Конституциясы, 45. 46, 47, 48 - баптар
Қазақстан Республикасының Конституциясы, 45. 46, 47, 48 - баптар Технологии регистрации заряженных частиц, сбор и анализ данных детекторов в ФВЭ. Введение. (Лекция 1)
Технологии регистрации заряженных частиц, сбор и анализ данных детекторов в ФВЭ. Введение. (Лекция 1) Тема 5. Доходы предприятия и финансовые результаты его деятельности.
Тема 5. Доходы предприятия и финансовые результаты его деятельности.  Организация производственной инфраструктуры предприятий сервиса
Организация производственной инфраструктуры предприятий сервиса Генеалогия - презентация для начальной школы_
Генеалогия - презентация для начальной школы_ Тема "Методы психологии".
Тема "Методы психологии".  Культура России XVIII века
Культура России XVIII века Мораль и политика
Мораль и политика Сервисный тренинг EXD06 DVD-навигация Phaeton
Сервисный тренинг EXD06 DVD-навигация Phaeton Проект «Начальная школа русской культуры»
Проект «Начальная школа русской культуры» Культура мови
Культура мови Новогодние алкогольные фанты
Новогодние алкогольные фанты Метрические задачи. Преобразования комплексного чертежа
Метрические задачи. Преобразования комплексного чертежа Роль и значение математических методов в экономике
Роль и значение математических методов в экономике