- Водорастворимые Витамины

Содержание
- 3. Метаболизм. В желудочно-кишечном тракте различные формы витамина гидролизуются с образованием свободного тиамина. Большая часть тиамина всасывается
- 5. Существует мнение, что основной транспортной формой тиамина является ТМФ. Витамин В1 присутствует в различных органах и
- 6. Основной коферментной формой (60-80% от общего внутриклеточного содержания) является ТПФ. ТТФ играет важную роль в метаболизме
- 7. Биохимические функции В1 Участие ТПФ в реакции прямого декарбоксилирования пировиноградной кислоты (ПВК). При декарбоксилировании ПВК с
- 9. Окислительное декарбоксилирование ПВК является одной из ключевых реакций в обмене углеводов. В результате этой реакции ПВК,
- 10. Помимо окислительных превращений ПВК и α-кетоглутарата, ТПФ принимает участие в окислительном декарбоксилировании кетокислот с разветвлённым углеродным
- 11. ТПФ – кофермент транскетолазы. Транскетолаза – фермент пентозофосфатного пути окисления углеводов. Физиологическая роль этого пути заключается
- 12. Витамин В1 принимает участие в синтезе ацетилхолина, катализируя в пируватдегидрогеназной реакции образование ацетил-КоА – субстрата ацетилирования
- 13. Витамин В2 Метаболизм. В пище витамин В2 находится преимущественно в составе своих коферментных форм, связанных с
- 15. Биохимические функции. Основное значение витамина В2 состоит в том, что он входит в состав флавиновых коферментов
- 16. ФАД – кофермент пируват- и α-кетоглутаратдегидрогеназных комплексов (наряду с ТПФ и другими коферментами ФАД осуществляет окислительное
- 18. Пантотеновая кислота (пантоил-β-алан ин) – светло-жёлтая маслянистая жидкость, хорошо растворимая в воде и этаноле. Спиртовое производное
- 19. Коферментными формами витамина В3, образующимися в цитоплазме клеток, являются: 4′-фосфопантетеин, дефосфо-КоА и Ко А-SH. Фосфопантетеин является
- 21. Активирование ацетата (образование ацетил-КоА – СН3-СО~S-Ko A). Ацетил-КоА является субстратом для синтеза жирных кислот, холестерина и
- 22. Транспорт жирных кислот в митохондрии. Окислительное декарбоксилирование кетокислот – пировиноградной (при этом образуется ацетил-КоА) и α–кетоглутаровой
- 24. Витамин В6 Витамин В6 часто называют «королём обмена аминокислот»; вместе с тем его коферментные формы участвуют
- 25. 1. Аминотрансфераз аминокислот, катализирующих обратимый перенос NH2- группы от аминокислоты на α-кетокислоту. В этой реакции образуются
- 26. Изомераз аминокислот, с помощью которых организм разрушает D-аминокислоты (в состав тканевых белков млекопитающих входят L-аминокислоты).4. Аминотрансфераз
- 27. Таким образом, витамин В6 характеризуется исключительно широким спектра биологического действия. Он принимает участие в регуляции белкового,
- 30. Фолиевая кислота (Фолацин. Витамин В9. Витамин ВС). Витамин В9 , всасываясь в тонком кишечнике, восстанавливается в
- 31. В клетках организма фолиевая кислота восстанавливается в активную форму – ТГФК – с помощью NADPH.H+ -зависимых
- 32. Оба фермента нуждаются в антиоксидантной (препятствующей окислительному разрушению) защите. Эту функцию выполняет витамин С. Отсюда становится
- 33. Коферментная форма фолиевой кислоты – ТГФК – необходима для мобилизации и использования в реакциях метаболизма одноуглеродных
- 34. Важнейшими реакциями с участием одноуглеродных фрагментов, связанных с ТГФК, являются: 5,10-метенил-ТГФК и 10-формил ТГФК служат донорами
- 36. . Содержащийся в пище витамин В12 в желудочном соке связывается с вырабатываемым обкладочными клетками слизистой желудка
- 37. При пероральном назначении высоких доз цианкобаламина он может абсорбироваться в тонком кишечнике путём пассивной диффузии без
- 38. Транспорт оксикобаламина кровью осуществляется двумя специфическими белками: транскобаламином I (α-глобулин с молекулярной массой ̴120000) и транскобаламином
- 39. В печени и почках оксикобаламин превращается в свои коферментные формы: метилкобаламин (метил-В12) и дезоксиаденозинкобаламин (д-аденозин-В12). Коферменты
- 40. Биохимические функции. К настоящему времени известно ~ 15 различных В12-регулируемых реакций, но только две из них
- 41. CH3-B12 ТГФК 5-метил-ТГФК SH S- CH3 | В12 | CH2 CH2 | | CH2 CH2 |
- 42. При уменьшении содержания в диете витамина В12 синтез метионина метионинсинтазой снижается, но поскольку при полноценном питании
- 43. О О ║ ║ С~ S KoA С ~S KoA │ метилмалонил-КоА-мутаза │ CH2 – CH
- 47. Аскорбиновая кислота всасывается путём простой диффузии на всём протяжении желудочно-кишечного тракта, но преимущественно это происходит в
- 48. Биохимические функции. Витамин С занимает доминирующее положение во внеклеточной антиоксидантной защите, значительно превосходящее в этом отношении
- 49. Витамин С может включаться в работу дыхательной цепи митохондрий, являясь донором электронов для цитохрома С. Очень
- 50. Гидроксилирование триптофана в 5-гидрокситриптофан (в реакции синтеза серотонина). Реакции гидроксилирования при биосинтезе гормонов корковой и мозговой
- 51. Витамин С активно участвует в обезвреживании токсинов, антибиотиков и других чужеродных для организма соединений, осуществляемых оксигеназной
- 52. Усиление прооксидантного действия витамина С приводит к нежелательным последствиям, особенно в условиях «перегрузки» организма железом. В
- 53. Помимо белков (в плазме крови), эту роль может на себя брать мочевая кислота (в цереброспинальной жидкости)
- 54. Таким образом, аскорбиновая кислота стабилизирует витамин Е (который легко разрушается), а витамин Е усиливает антиоксидантное действие
- 56. Скачать презентацию
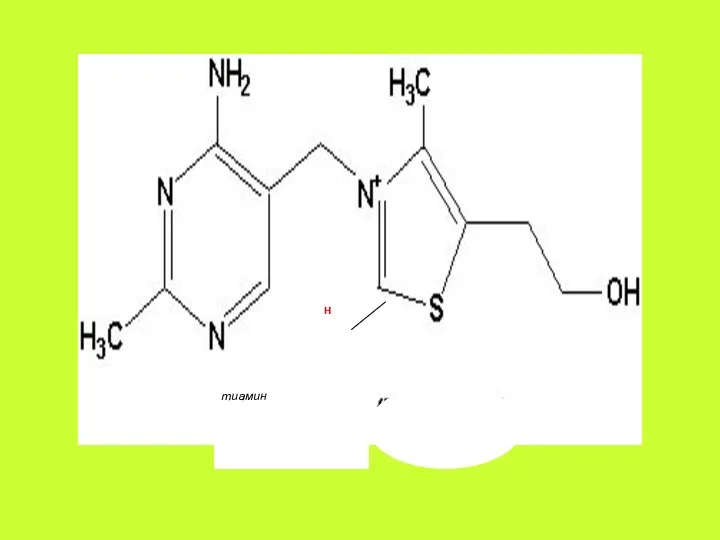

Метаболизм. В желудочно-кишечном тракте различные формы витамина гидролизуются с образованием
Метаболизм. В желудочно-кишечном тракте различные формы витамина гидролизуются с образованием
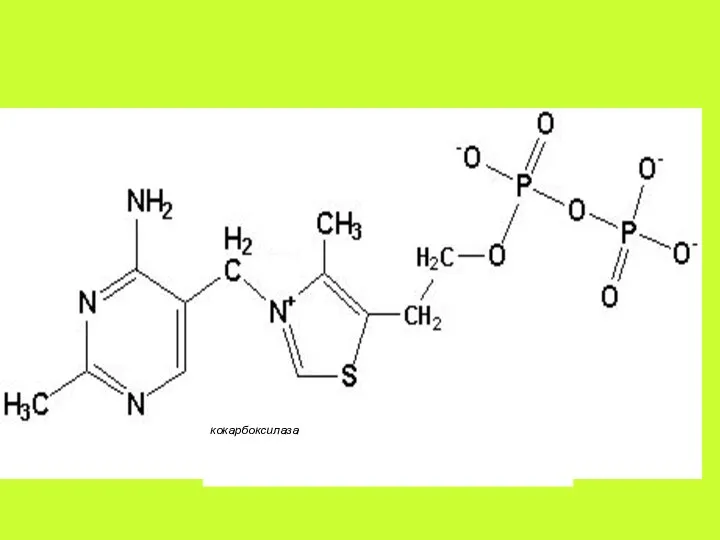

Существует мнение, что основной транспортной формой тиамина является ТМФ.
Витамин В1 присутствует
Существует мнение, что основной транспортной формой тиамина является ТМФ.
Витамин В1 присутствует

Основной коферментной формой (60-80% от общего внутриклеточного содержания) является ТПФ.
Основной коферментной формой (60-80% от общего внутриклеточного содержания) является ТПФ.

Биохимические функции В1
Участие ТПФ в реакции прямого декарбоксилирования пировиноградной кислоты
Биохимические функции В1
Участие ТПФ в реакции прямого декарбоксилирования пировиноградной кислоты
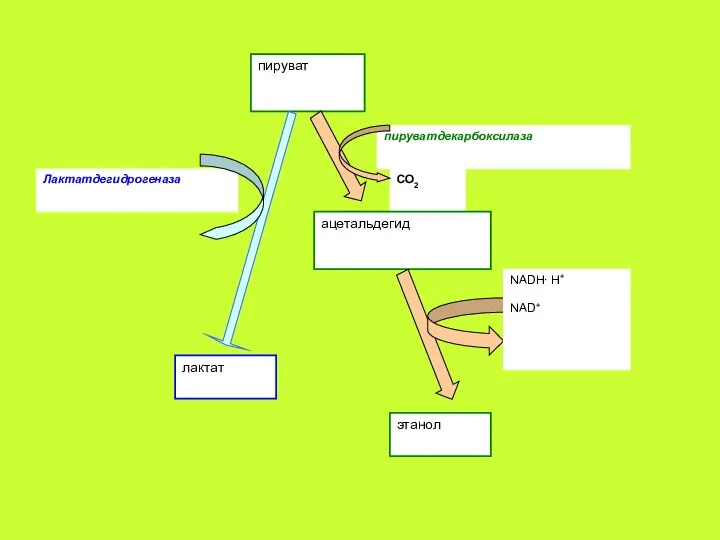

Окислительное декарбоксилирование ПВК является одной из ключевых реакций в обмене углеводов.
Окислительное декарбоксилирование ПВК является одной из ключевых реакций в обмене углеводов.

Помимо окислительных превращений ПВК и α-кетоглутарата, ТПФ принимает участие в
Помимо окислительных превращений ПВК и α-кетоглутарата, ТПФ принимает участие в

ТПФ – кофермент транскетолазы. Транскетолаза – фермент пентозофосфатного пути окисления
ТПФ – кофермент транскетолазы. Транскетолаза – фермент пентозофосфатного пути окисления

Витамин В1 принимает участие в синтезе ацетилхолина, катализируя в пируватдегидрогеназной
Витамин В1 принимает участие в синтезе ацетилхолина, катализируя в пируватдегидрогеназной

Витамин В2
Метаболизм. В пище витамин В2 находится преимущественно в составе своих
Витамин В2
Метаболизм. В пище витамин В2 находится преимущественно в составе своих
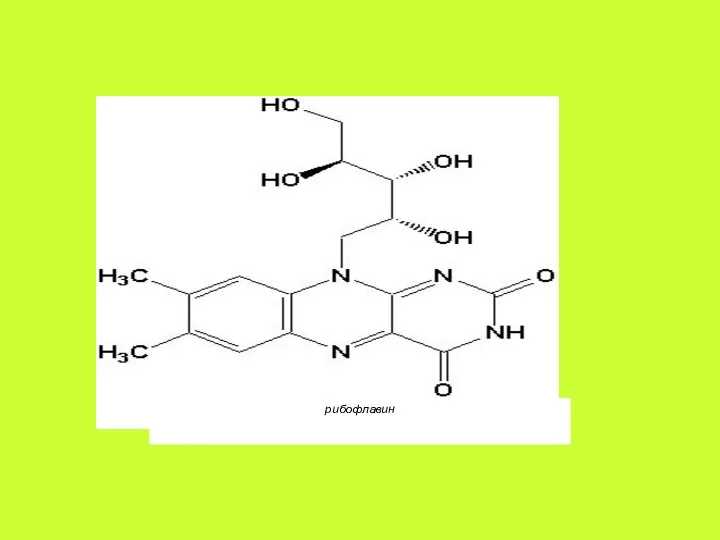

Биохимические функции. Основное значение витамина В2 состоит в том, что он
Биохимические функции. Основное значение витамина В2 состоит в том, что он

ФАД – кофермент пируват- и α-кетоглутаратдегидрогеназных комплексов (наряду с ТПФ
ФАД – кофермент пируват- и α-кетоглутаратдегидрогеназных комплексов (наряду с ТПФ
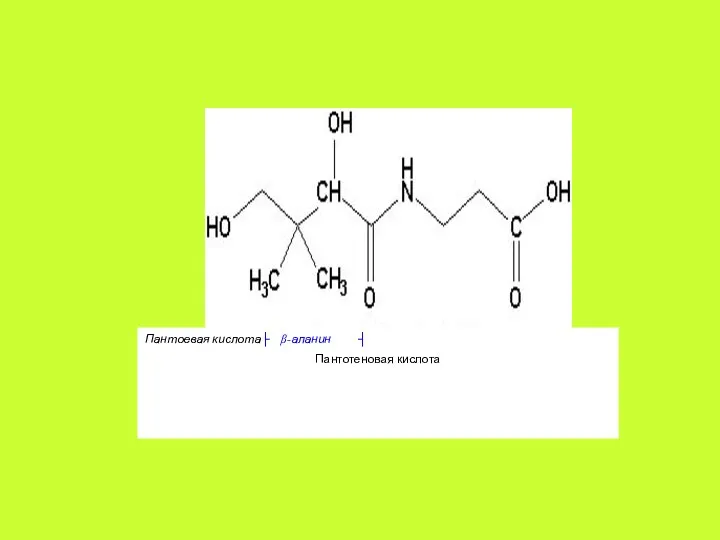

Пантотеновая кислота (пантоил-β-алан ин) – светло-жёлтая маслянистая жидкость, хорошо растворимая в
Пантотеновая кислота (пантоил-β-алан ин) – светло-жёлтая маслянистая жидкость, хорошо растворимая в

Коферментными формами витамина В3, образующимися в цитоплазме клеток, являются: 4′-фосфопантетеин,
Коферментными формами витамина В3, образующимися в цитоплазме клеток, являются: 4′-фосфопантетеин,
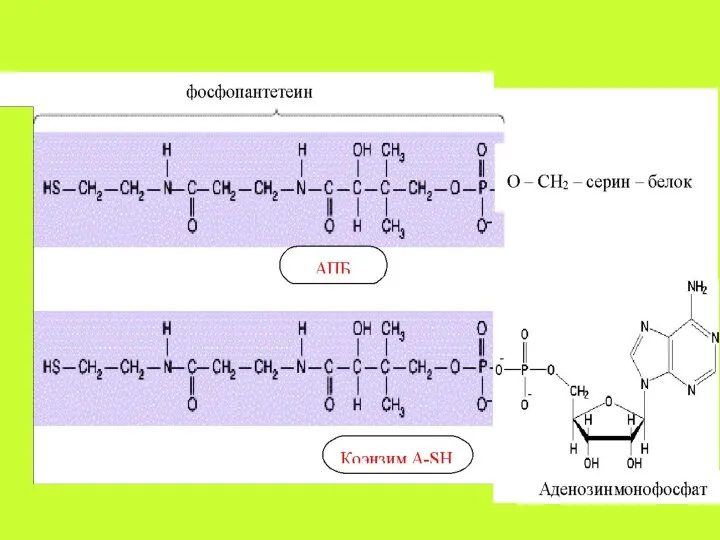

Активирование ацетата (образование ацетил-КоА – СН3-СО~S-Ko A). Ацетил-КоА является субстратом для
Активирование ацетата (образование ацетил-КоА – СН3-СО~S-Ko A). Ацетил-КоА является субстратом для

Транспорт жирных кислот в митохондрии.
Окислительное декарбоксилирование кетокислот – пировиноградной (при
Транспорт жирных кислот в митохондрии.
Окислительное декарбоксилирование кетокислот – пировиноградной (при
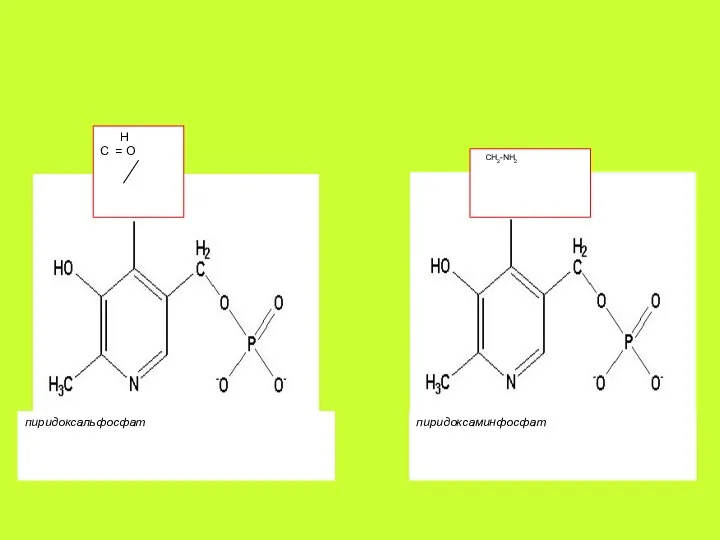

Витамин В6
Витамин В6 часто называют «королём обмена аминокислот»; вместе с тем
Витамин В6
Витамин В6 часто называют «королём обмена аминокислот»; вместе с тем

1. Аминотрансфераз аминокислот, катализирующих обратимый перенос NH2-
группы от аминокислоты на
1. Аминотрансфераз аминокислот, катализирующих обратимый перенос NH2-
группы от аминокислоты на

Изомераз аминокислот, с помощью которых организм разрушает D-аминокислоты (в состав тканевых
Изомераз аминокислот, с помощью которых организм разрушает D-аминокислоты (в состав тканевых

Таким образом, витамин В6 характеризуется исключительно широким спектра биологического действия. Он
Таким образом, витамин В6 характеризуется исключительно широким спектра биологического действия. Он
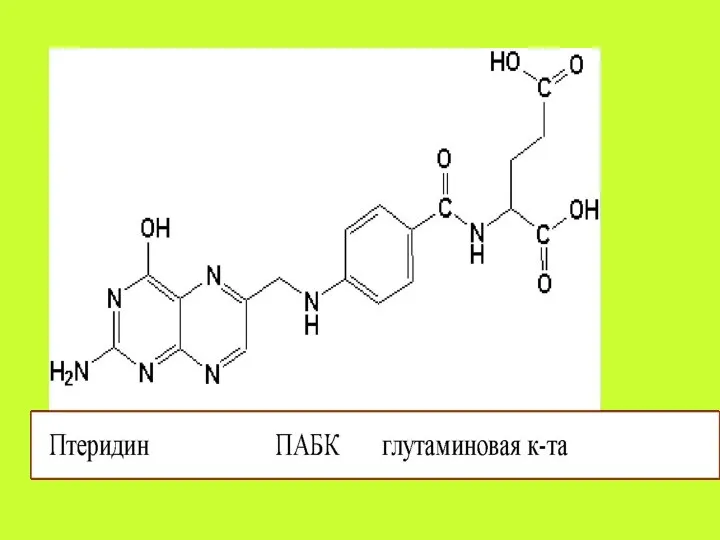
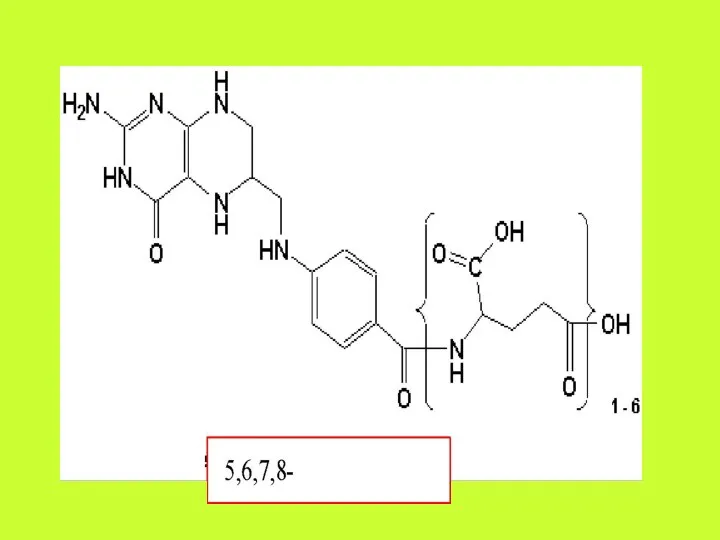

Фолиевая кислота (Фолацин. Витамин В9. Витамин ВС).
Витамин В9 , всасываясь в
Фолиевая кислота (Фолацин. Витамин В9. Витамин ВС).
Витамин В9 , всасываясь в

В клетках организма фолиевая кислота восстанавливается в активную форму – ТГФК
В клетках организма фолиевая кислота восстанавливается в активную форму – ТГФК

Оба фермента нуждаются в антиоксидантной (препятствующей окислительному разрушению) защите. Эту функцию
Оба фермента нуждаются в антиоксидантной (препятствующей окислительному разрушению) защите. Эту функцию

Коферментная форма фолиевой кислоты – ТГФК – необходима для мобилизации и
Коферментная форма фолиевой кислоты – ТГФК – необходима для мобилизации и

Важнейшими реакциями с участием одноуглеродных фрагментов, связанных с ТГФК, являются:
Важнейшими реакциями с участием одноуглеродных фрагментов, связанных с ТГФК, являются:
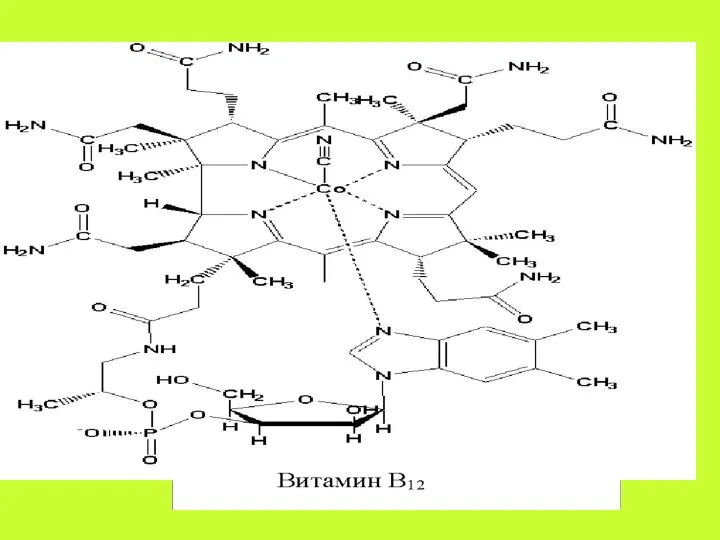

. Содержащийся в пище витамин В12 в желудочном соке связывается с
. Содержащийся в пище витамин В12 в желудочном соке связывается с

При пероральном назначении высоких доз цианкобаламина он может абсорбироваться в тонком
При пероральном назначении высоких доз цианкобаламина он может абсорбироваться в тонком

Транспорт оксикобаламина кровью осуществляется двумя специфическими белками: транскобаламином I (α-глобулин с
Транспорт оксикобаламина кровью осуществляется двумя специфическими белками: транскобаламином I (α-глобулин с

В печени и почках оксикобаламин превращается в свои коферментные формы: метилкобаламин
В печени и почках оксикобаламин превращается в свои коферментные формы: метилкобаламин

Биохимические функции. К настоящему времени известно ~ 15 различных В12-регулируемых реакций,
Биохимические функции. К настоящему времени известно ~ 15 различных В12-регулируемых реакций,
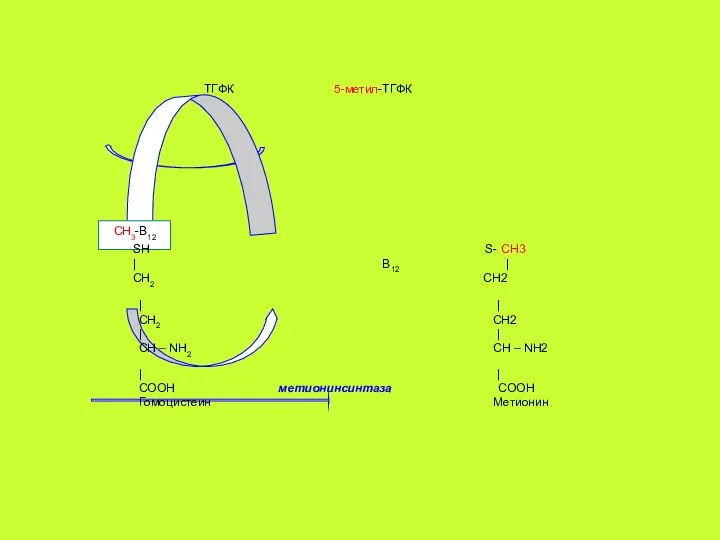
CH3-B12
ТГФК 5-метил-ТГФК
SH S- CH3
| В12 |
CH2 CH2
| |
CH2 CH2
|
CH3-B12
ТГФК 5-метил-ТГФК
SH S- CH3
| В12 |
CH2 CH2
| |
CH2 CH2
|

При уменьшении содержания в диете витамина В12 синтез метионина метионинсинтазой снижается,
При уменьшении содержания в диете витамина В12 синтез метионина метионинсинтазой снижается,
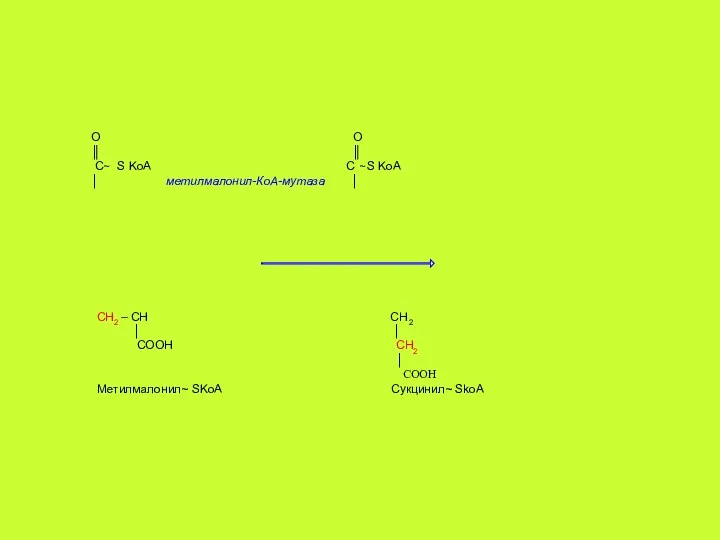
О О
║ ║
С~ S KoA С ~S KoA
О О
║ ║
С~ S KoA С ~S KoA
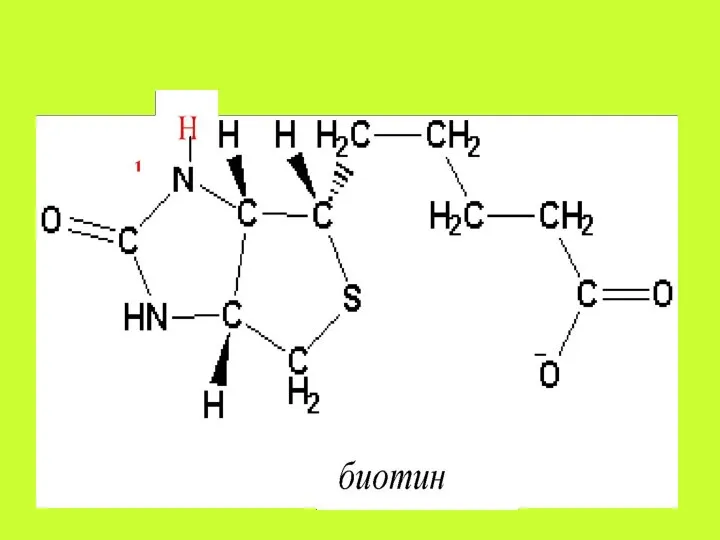
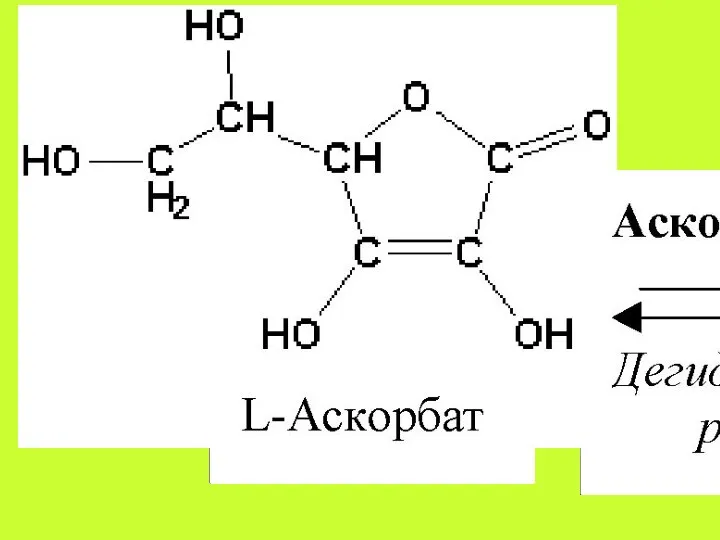
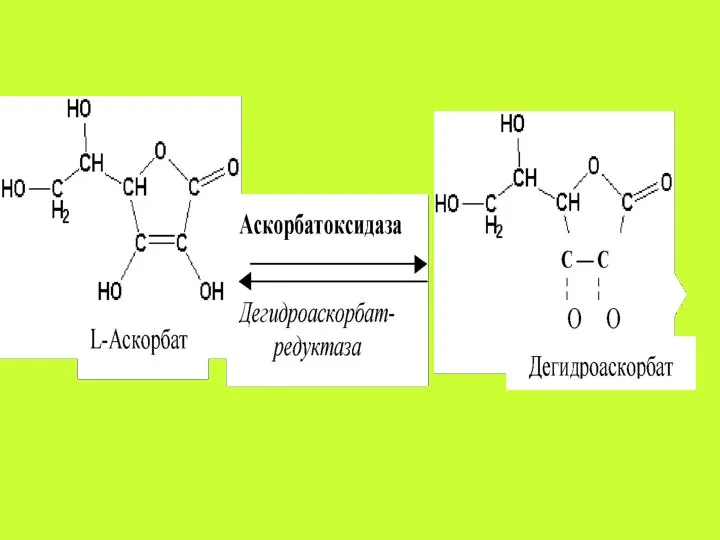

Аскорбиновая кислота всасывается путём простой диффузии на всём протяжении желудочно-кишечного тракта,
Аскорбиновая кислота всасывается путём простой диффузии на всём протяжении желудочно-кишечного тракта,

Биохимические функции. Витамин С занимает доминирующее положение во внеклеточной антиоксидантной защите,
Биохимические функции. Витамин С занимает доминирующее положение во внеклеточной антиоксидантной защите,

Витамин С может включаться в работу дыхательной цепи митохондрий, являясь донором
Витамин С может включаться в работу дыхательной цепи митохондрий, являясь донором

Гидроксилирование триптофана в 5-гидрокситриптофан (в реакции синтеза серотонина).
Реакции гидроксилирования при биосинтезе
Гидроксилирование триптофана в 5-гидрокситриптофан (в реакции синтеза серотонина).
Реакции гидроксилирования при биосинтезе

Витамин С активно участвует в обезвреживании токсинов, антибиотиков и других чужеродных
Витамин С активно участвует в обезвреживании токсинов, антибиотиков и других чужеродных

Усиление прооксидантного действия витамина С приводит к нежелательным последствиям, особенно в
Усиление прооксидантного действия витамина С приводит к нежелательным последствиям, особенно в

Помимо белков (в плазме крови), эту роль может на себя брать
Помимо белков (в плазме крови), эту роль может на себя брать

Таким образом, аскорбиновая кислота стабилизирует витамин Е (который легко разрушается), а
Таким образом, аскорбиновая кислота стабилизирует витамин Е (который легко разрушается), а
 Хлор в организме человека
Хлор в организме человека Ключевые события
Ключевые события PHONE STAGE
PHONE STAGE Как разрешать кризисные душевные состояния и не попадать в них
Как разрешать кризисные душевные состояния и не попадать в них Технологический подход к организации образовательного процесса В.А. Степихова, доцент кафедры социально-педагогического процес
Технологический подход к организации образовательного процесса В.А. Степихова, доцент кафедры социально-педагогического процес Операционные усилители
Операционные усилители  Летний лагерь личностного самоопределения и роста для детей и подростков
Летний лагерь личностного самоопределения и роста для детей и подростков Динамика, типология, функции культуры
Динамика, типология, функции культуры Автоматические межпланетные станции(АМС)
Автоматические межпланетные станции(АМС) Производственная практика в компании ООО «Спецмонтаж-Ц1»
Производственная практика в компании ООО «Спецмонтаж-Ц1» Политическая и социогуманитарная составляющие идеологии белорусского государства
Политическая и социогуманитарная составляющие идеологии белорусского государства Расчет на сейсмические воздействия по методике СП 14.13330.2014
Расчет на сейсмические воздействия по методике СП 14.13330.2014 Kurzgutachten Zentrifugalmotor für die Fa. Etorque GmbH
Kurzgutachten Zentrifugalmotor für die Fa. Etorque GmbH Улучшение системы видеонаблюдения на железнодорожном вокзале
Улучшение системы видеонаблюдения на железнодорожном вокзале Мастера Гжели
Мастера Гжели Презентация «Философия» Лекция 2. Философия древнего мира.
Презентация «Философия» Лекция 2. Философия древнего мира. Kinds of sports
Kinds of sports Историко-правовой опыт Российской империи и возможности современной правовой политики на Северном Кавказе
Историко-правовой опыт Российской империи и возможности современной правовой политики на Северном Кавказе Теориялық механика
Теориялық механика Цифровое зарядное устройство для сотовых телефонов
Цифровое зарядное устройство для сотовых телефонов Пришло Рождество – начинай торжество Бинарный урок 1 класс Подготовили учитель ИЗО Ли Елена Александровна и учитель музыки Пе
Пришло Рождество – начинай торжество Бинарный урок 1 класс Подготовили учитель ИЗО Ли Елена Александровна и учитель музыки Пе Как правильно утеплить крышу дома
Как правильно утеплить крышу дома Центр лыжного и горнолыжного спорта "Малиновка"
Центр лыжного и горнолыжного спорта "Малиновка" Жыл басы - Наурыз
Жыл басы - Наурыз Микропроцессорный анализатор дисперсности мад-4
Микропроцессорный анализатор дисперсности мад-4 Преступления против несовершеннолетних и их семей
Преступления против несовершеннолетних и их семей  Язык и культура как две взаимообусловленные сущности
Язык и культура как две взаимообусловленные сущности Достоинства леммы Маркова и неравенства Чебышева при оценивании риска
Достоинства леммы Маркова и неравенства Чебышева при оценивании риска