- Эволюция филогенетических групп. (лекция 11)

Содержание
- 2. Филогенетика выясняет особенности эволюции крупных групп органического мира
- 4. Формы филогенеза первичные филетическая эволюция дивергенция вторичные параллелизм конвергенция
- 5. Филетическая эволюция изменения, происходящие в одном филогенетическом стволе без учета всегда возможных дивергентных ответвлений
- 6. Пример Развитие предков лошадей по прямой линии фенакодус — эогиппус — миогиппус — парагиппус — плиогиппус
- 9. Уровни Филетическая эволюция - в пределах любого вида любой вид развивается во времени вид в целом
- 10. Дивергенция первичная форма эволюции таксона в результате изменения направления отбора в разных условиях происходит дивергенция (расхождение)
- 13. Дивергенция – в результате изоляции, волн жизни, мутационного процесса и естественного отбора популяции и группы популяций
- 14. Дивергенции внутри вида (микроэволюционный уровень) и в группах более крупных (макроэволюционный уровень), - различие, на микроэволюционном
- 15. Конвергенция это процесс формирования сходного фенотипического облика особей двух или нескольких групп. примером конвергентного развития в
- 18. отличие конвергентное сходство от дивергентного родство зайцеобразных (Lagomorpha) с настоящими грызунами (Rodentia) основано на дивергенции групп
- 20. конвергенция не бывает глубокой (в отличие от сходства, основанного на филогенетическом родстве). конвергенция —сходство, обусловленное аналогичными,
- 21. форма конвергентного развития - параллелизм формирование сходного фенотипического облика первоначально дивергировавшими и генетически близкими группами. Классическим
- 22. Пример параллелизмов в эволюции: развитие саблезубости у крупных кошек: А—махайрод (Hoplophoneus), олигоцен; Б—лжесаблезубая настоящая кошка (Dinictis),
- 23. Macrauchenia patachonica
- 24. Пиротерий
- 25. Направления эволюции Аллогенез¹ — развитие группы внутри одной адаптивной зоны² с возникновением близких форм, различающихся адаптациями
- 26. При выделении аллогенеза как типа развития группы принципиален не его масштаб (который может быть и на
- 27. Степень специализации видов может быть различна: глубокая специализация затрагивает комплекс важных признаков, тогда как частичная —
- 28. Арогенез. Как свидетельствует палеонтологическая летопись, из одной природной зоны в другую обычно попадают лишь отдельные, сравнительно
- 29. В качестве примера арогенеза сравнительно небольшого масштаба можно рассматривать возникновение и расцвет класса птиц. Проникнуть в
- 30. В эволюции растений возникновение проводящей сосудистой системы, эпидермы, устьиц, а также семязачатков и пыльцевой трубки имело
- 31. Эволюция не всегда идет по пути усложнения. Иногда арогенез, ведущий к занятию новой адаптивной зоны, может
- 32. Следует отметить, что и ароморфозы (определяющие арогенез группы) и алломорфозы (определяющие аллогенез группы) всегда возникают как
- 33. Темпы эволюции групп Эволюция как грандиозный процесс изменения органического мира Земли характеризуется определенными временными параметрами. Скорость,
- 34. В эволюции большее значение имеет не абсолютное, астрономическое, время, а число поколений. Причины этого будут понятны,
- 35. Проблема выбора критериев скорости эволюции Теоретически скорость эволюции вида можно определить по темпу его изменения в
- 36. Более обычно в эволюции постепенное формообразование. Сейчас накопилось много данных, позволяющих сравнительно точно определять скорость образования
- 38. Скачать презентацию

Филогенетика
выясняет особенности эволюции крупных групп органического мира
Филогенетика
выясняет особенности эволюции крупных групп органического мира
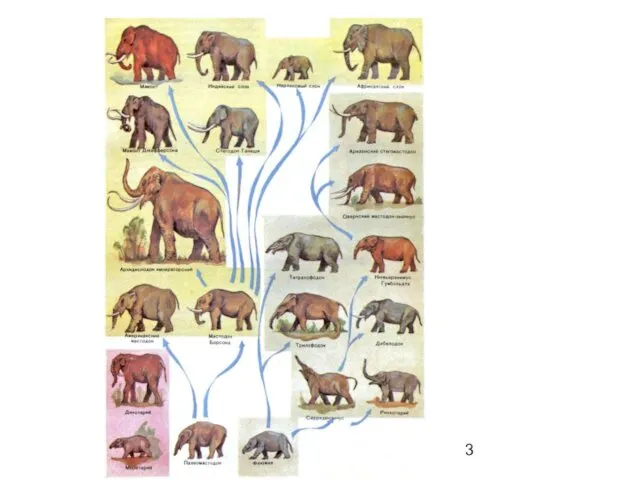

Формы филогенеза
первичные
филетическая эволюция
дивергенция
вторичные
параллелизм
конвергенция
Формы филогенеза
первичные
филетическая эволюция
дивергенция
вторичные
параллелизм
конвергенция

Филетическая эволюция
изменения, происходящие в одном филогенетическом стволе
без учета всегда возможных
Филетическая эволюция
изменения, происходящие в одном филогенетическом стволе
без учета всегда возможных

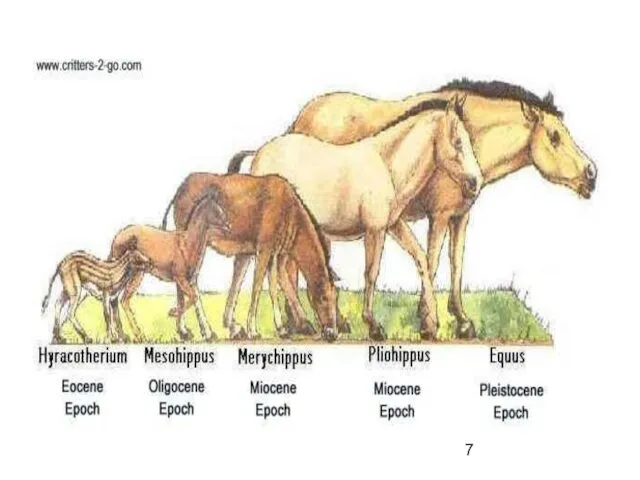
Пример
Развитие предков лошадей по прямой линии
фенакодус — эогиппус — миогиппус
Пример
Развитие предков лошадей по прямой линии
фенакодус — эогиппус — миогиппус


Уровни
Филетическая эволюция - в пределах любого вида
любой вид развивается во времени
Уровни
Филетическая эволюция - в пределах любого вида
любой вид развивается во времени

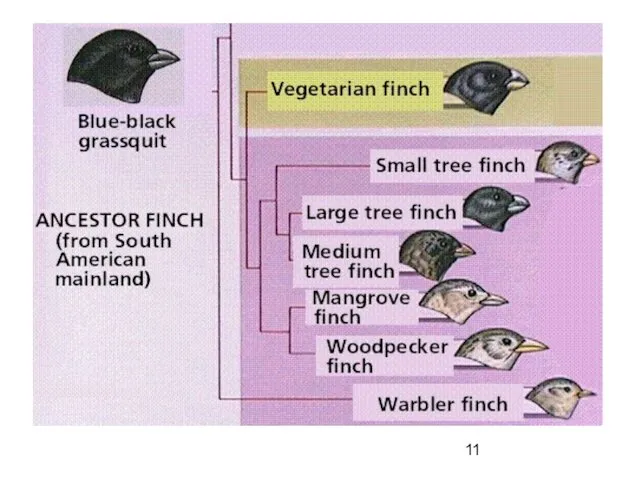
Дивергенция
первичная форма эволюции таксона
в результате изменения направления отбора в разных условиях
Дивергенция
первичная форма эволюции таксона
в результате изменения направления отбора в разных условиях
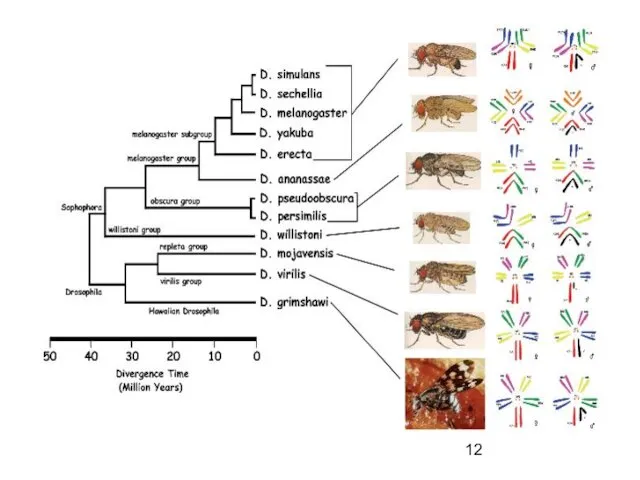

Дивергенция – в результате
изоляции, волн жизни, мутационного процесса и естественного
Дивергенция – в результате
изоляции, волн жизни, мутационного процесса и естественного

Дивергенции
внутри вида (микроэволюционный уровень) и в группах более крупных (макроэволюционный уровень),
Дивергенции
внутри вида (микроэволюционный уровень) и в группах более крупных (макроэволюционный уровень),

Конвергенция
это процесс формирования сходного фенотипического облика особей двух или нескольких групп.
Конвергенция
это процесс формирования сходного фенотипического облика особей двух или нескольких групп.



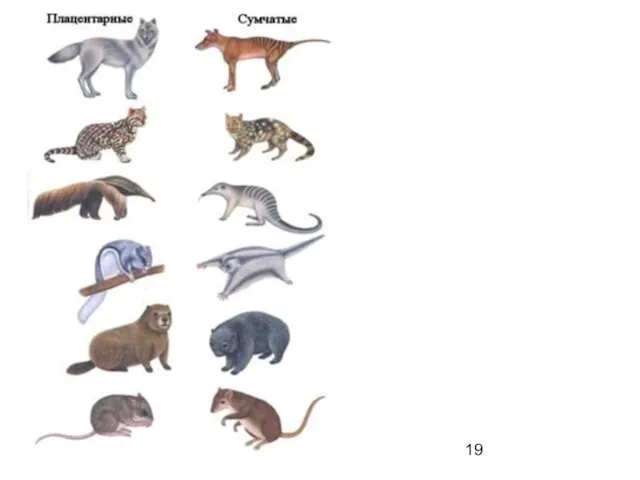
отличие конвергентное сходство от дивергентного
родство зайцеобразных (Lagomorpha) с настоящими грызунами (Rodentia)
отличие конвергентное сходство от дивергентного
родство зайцеобразных (Lagomorpha) с настоящими грызунами (Rodentia)

конвергенция
не бывает глубокой (в отличие от сходства, основанного на филогенетическом родстве).
конвергенция
не бывает глубокой (в отличие от сходства, основанного на филогенетическом родстве).

форма конвергентного развития - параллелизм
формирование сходного фенотипического облика первоначально дивергировавшими
форма конвергентного развития - параллелизм
формирование сходного фенотипического облика первоначально дивергировавшими
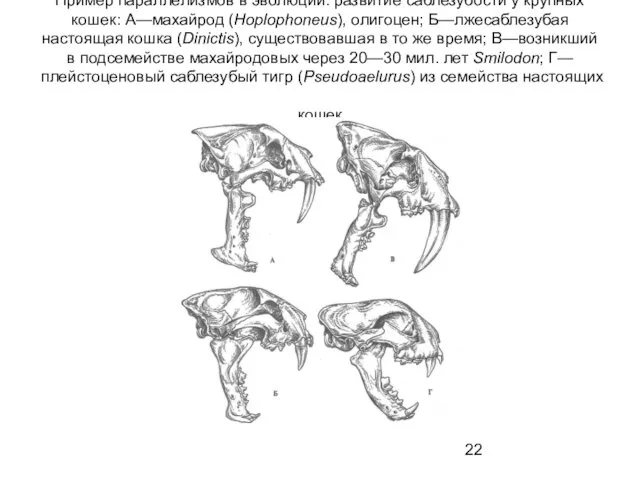
Пример параллелизмов в эволюции: развитие саблезубости у крупных кошек: А—махайрод (Hoplophoneus),
Пример параллелизмов в эволюции: развитие саблезубости у крупных кошек: А—махайрод (Hoplophoneus),

Macrauchenia patachonica
Macrauchenia patachonica

Пиротерий
Пиротерий

Направления эволюции
Аллогенез¹ — развитие группы внутри одной адаптивной зоны² с
Направления эволюции
Аллогенез¹ — развитие группы внутри одной адаптивной зоны² с

При выделении аллогенеза как типа развития группы принципиален не его масштаб
При выделении аллогенеза как типа развития группы принципиален не его масштаб

Степень специализации видов может быть различна: глубокая специализация затрагивает комплекс важных
Степень специализации видов может быть различна: глубокая специализация затрагивает комплекс важных

Арогенез. Как свидетельствует палеонтологическая летопись, из одной природной зоны в другую
Арогенез. Как свидетельствует палеонтологическая летопись, из одной природной зоны в другую

В качестве примера арогенеза сравнительно небольшого масштаба можно рассматривать возникновение и
В качестве примера арогенеза сравнительно небольшого масштаба можно рассматривать возникновение и

В эволюции растений возникновение проводящей сосудистой системы, эпидермы, устьиц, а также
В эволюции растений возникновение проводящей сосудистой системы, эпидермы, устьиц, а также

Эволюция не всегда идет по пути усложнения. Иногда арогенез, ведущий к
Эволюция не всегда идет по пути усложнения. Иногда арогенез, ведущий к

Следует отметить, что и ароморфозы (определяющие арогенез группы) и алломорфозы (определяющие
Следует отметить, что и ароморфозы (определяющие арогенез группы) и алломорфозы (определяющие

Темпы эволюции групп
Эволюция как грандиозный процесс изменения органического мира Земли
Темпы эволюции групп
Эволюция как грандиозный процесс изменения органического мира Земли

В эволюции большее значение имеет не абсолютное, астрономическое, время, а число
В эволюции большее значение имеет не абсолютное, астрономическое, время, а число

Проблема выбора критериев скорости эволюции
Теоретически скорость эволюции вида можно определить
Проблема выбора критериев скорости эволюции
Теоретически скорость эволюции вида можно определить

Более обычно в эволюции постепенное формообразование. Сейчас накопилось много данных, позволяющих
Более обычно в эволюции постепенное формообразование. Сейчас накопилось много данных, позволяющих
 Види кровотеч
Види кровотеч  Черепаха Гарриета животное, признанное одним из старейших в мире.
Черепаха Гарриета животное, признанное одним из старейших в мире.  ПЧЕЛЫ И МУРАВЬИ -ОБЩЕСТВЕННЫЕ НАСЕКОМЫЕ УЧИТЕЛЬ БИОЛОГИИ: ГАЛКОВСКАЯ И.И. 7 КЛАСС
ПЧЕЛЫ И МУРАВЬИ -ОБЩЕСТВЕННЫЕ НАСЕКОМЫЕ УЧИТЕЛЬ БИОЛОГИИ: ГАЛКОВСКАЯ И.И. 7 КЛАСС Вегетативное размножение растений
Вегетативное размножение растений Органы дыхания человека Учитель биологии МОУ « Езвинская СОШ» Касьянова Светлана Валентиновна ( урок с элементами моделирования)
Органы дыхания человека Учитель биологии МОУ « Езвинская СОШ» Касьянова Светлана Валентиновна ( урок с элементами моделирования) Функции врожденного иммунитета
Функции врожденного иммунитета Продолговатый, задний, ромбовидный, средний, промежуточный мозг
Продолговатый, задний, ромбовидный, средний, промежуточный мозг Физиология бактерий. Рост и размножение бактерий
Физиология бактерий. Рост и размножение бактерий Республиканский центр по генетическому маркированию и паспортизации растений, животных, микроорганизмов и человека
Республиканский центр по генетическому маркированию и паспортизации растений, животных, микроорганизмов и человека Аттестационная работа. Основы физико-химических явлений в биологии и биотехнологии
Аттестационная работа. Основы физико-химических явлений в биологии и биотехнологии Апельсин, или Citrus sinensis
Апельсин, или Citrus sinensis ГМО: мифы и реальность
ГМО: мифы и реальность Презентация на тему "Опорно – двигательная система" - скачать презентации по Биологии
Презентация на тему "Опорно – двигательная система" - скачать презентации по Биологии Происхождение вещества. Биологическая эволюция в развитии биосферы
Происхождение вещества. Биологическая эволюция в развитии биосферы Наука о растениях – ботаника Мир растений
Наука о растениях – ботаника Мир растений Клещевой энцефалит
Клещевой энцефалит  Пищеварительная система. Остальные системы жизнедеятельности человека
Пищеварительная система. Остальные системы жизнедеятельности человека Белка обыкновенная Работа ученицы 2 «А» класса МОУ «ООШ № 57» Серебряковой Станиславы Руководитель: Воробьёва М.А. Учитель
Белка обыкновенная Работа ученицы 2 «А» класса МОУ «ООШ № 57» Серебряковой Станиславы Руководитель: Воробьёва М.А. Учитель  Cчастливый случай Покрытосеменные
Cчастливый случай Покрытосеменные Мочеполовая система
Мочеполовая система Quorum sensing
Quorum sensing Возбудимые ткани. Происхождение потенциала действия, условия возникновения возбуждения
Возбудимые ткани. Происхождение потенциала действия, условия возникновения возбуждения Обитатели подводного мира
Обитатели подводного мира Презентация на тему "Лист и видоизменения листа" - скачать презентации по Биологии
Презентация на тему "Лист и видоизменения листа" - скачать презентации по Биологии Растения. Подготовка к ЕГЭ по биологии
Растения. Подготовка к ЕГЭ по биологии Презентация на тему "Отряды насекомых Стрекозы, Вши, Жуки," - скачать бесплатно презентации по Биологии
Презентация на тему "Отряды насекомых Стрекозы, Вши, Жуки," - скачать бесплатно презентации по Биологии 03 К УРОКУ 5 Происхождение современного человека
03 К УРОКУ 5 Происхождение современного человека Как живут животные
Как живут животные