- Физиология эндокринной системы

Содержание
- 2. Взаимосвязь между нервной и эндокринной системами
- 3. Различные формы гуморальных связей между клетками
- 4. Некоторые группы химических посредников и регуляторов
- 5. Химические посредники: клетки эндокринной системы секретируют гормоны, клетки иммунной системы - цитокины; различные клетки при иммунных
- 6. По химической структуре гормоны, подразделяют на: пептиды, стероиды, производные аминокислот, производные арахидоновой кислоты.
- 7. Пептидные гормоны - секретируются путем экзоцитоза, - относятся к полярным веществам, поэтому не могут проникать через
- 8. Стероидные гормоны - производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол). - относятся к неполярным, поэтому
- 9. Производные аминокислот: Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и дофамин. Молекулы тироксина (T4)
- 10. Производные арахидоновой кислоты (эйкозаноиды) - простагландины, простациклины, тромбоксаны, лейкотриены. - действуют как локальные биорегуляторы путем связывания
- 11. Основные механизмы нейроэндокринной регуляции клеток
- 12. I. Центральные регуляторные образования эндокринной системы: гипоталамус (нейросекреторные ядра); гипофиз (аденогипофиз и нейрогипофиз); эпифиз. II. Периферические
- 13. Транспорт гормонов осуществляется кровью в комплексе с белками. Часть этих белков - специфические транспортные протеины (например,
- 14. Образование связанной формы гормонов : 1. предохраняет организм от избыточного накопления в крови свободных гормонов (и,
- 15. Механизмы действия гормонов
- 16. Прямой механизм действия гормонов на клетку А. Жирорастворимый гормон проходит через мембрану и, присоединившись внутри клетки
- 17. Ядерные рецепторы Ядерные рецепторы - это ДНК-связывающие факторы транскрипции, локализованые в ядре. В отсутствие лиганда основная
- 18. Опосредованный механизм действия гормонов на клетку Б. Водорастворимый гормон присоединяется к рецептору на поверхности клетки и
- 19. Характерной особенностью действия этих гормонов является быстрый эффект – обусловленный активацией уже существующих ферментов и других
- 20. Вторичные мессенджеры
- 21. Требования предъявляемые вторичному мессенджеру: 1. небольшая молекулярная масса (чтобы с высокой скоростью диффундировать в цитоплазме), 2.
- 22. аденилатциклаза - цАМФ; гуанилатциклаза - цГМФ; фосфолипаза С - ИФ3; Са2+. Газовые посредники: NO, СО, H2S
- 23. Система аденилатциклаза - цАМФ - фосфорилирование белков меняет проницаемость мембран, активность и количество ферментов. - метилирование
- 24. Система гуанилатциклаза - цГМФ Активация гуанилатциклазы происходит опосредованно через Са2+ (ацетилхолин, натрийуретический гормон) и оксидантные системы
- 25. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны (сердце, гладкие мышцы кишечника) Ферментативный
- 26. Система фосфолипаза С - ИФ3
- 27. Система Са2+-кальмодулин
- 28. Система Са2+ -кальмодулин является пусковым стимулом для многочисленных физиологических процессов: сокращения мышц, секреции гормонов, выделения медиаторов,
- 29. NO как вторичный мессенджер образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего в нервной ткани, эндотелии
- 30. Механизм действия В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в активный центр гуанилатциклазы ионом
- 31. NO может либо усиливать интенсивность процессов, либо подавлять их. Примером подавления процесса при активации NOS является
- 33. Скачать презентацию
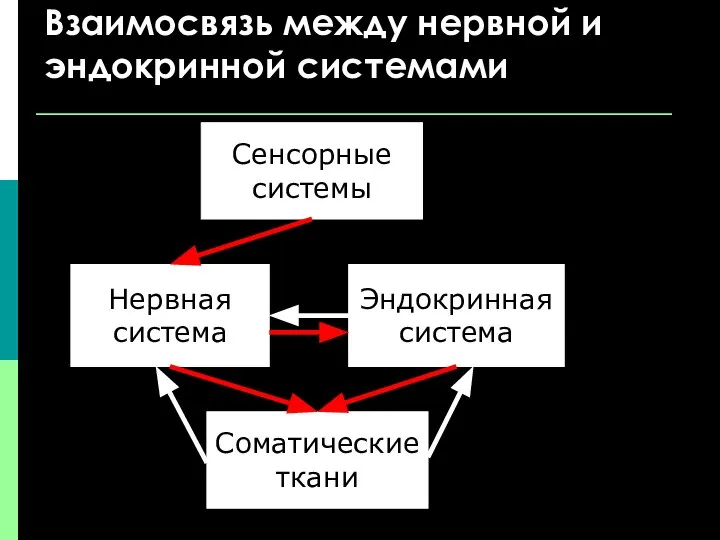
Взаимосвязь между нервной и эндокринной системами
Взаимосвязь между нервной и эндокринной системами
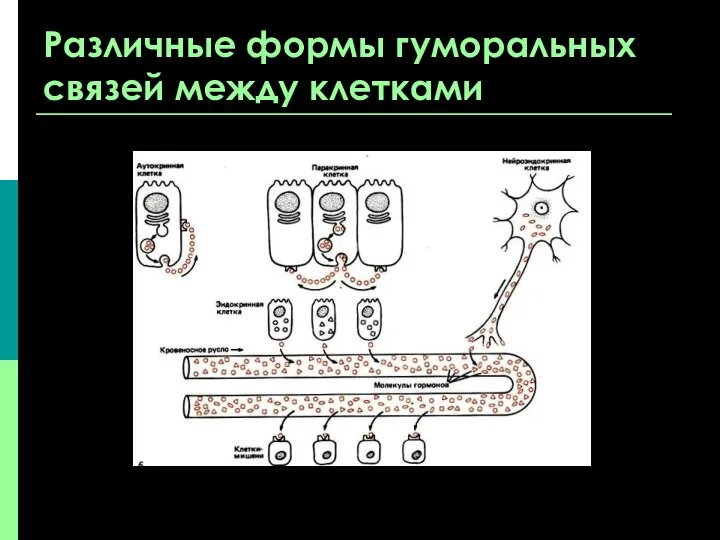
Различные формы гуморальных связей между клетками
Различные формы гуморальных связей между клетками

Некоторые группы химических посредников и регуляторов
Некоторые группы химических посредников и регуляторов

Химические посредники:
клетки эндокринной системы секретируют гормоны,
клетки иммунной системы - цитокины;
различные клетки
Химические посредники:
клетки эндокринной системы секретируют гормоны,
клетки иммунной системы - цитокины;
различные клетки

По химической структуре гормоны, подразделяют на:
пептиды,
стероиды,
производные аминокислот,
производные арахидоновой кислоты.
По химической структуре гормоны, подразделяют на:
пептиды,
стероиды,
производные аминокислот,
производные арахидоновой кислоты.

Пептидные гормоны
- секретируются путем экзоцитоза,
- относятся к полярным веществам, поэтому не
Пептидные гормоны
- секретируются путем экзоцитоза,
- относятся к полярным веществам, поэтому не

Стероидные гормоны
- производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол).
-
Стероидные гормоны
- производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол).
-

Производные аминокислот:
Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и
Производные аминокислот:
Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и

Производные арахидоновой кислоты (эйкозаноиды)
- простагландины, простациклины, тромбоксаны, лейкотриены.
- действуют как
Производные арахидоновой кислоты (эйкозаноиды)
- простагландины, простациклины, тромбоксаны, лейкотриены.
- действуют как
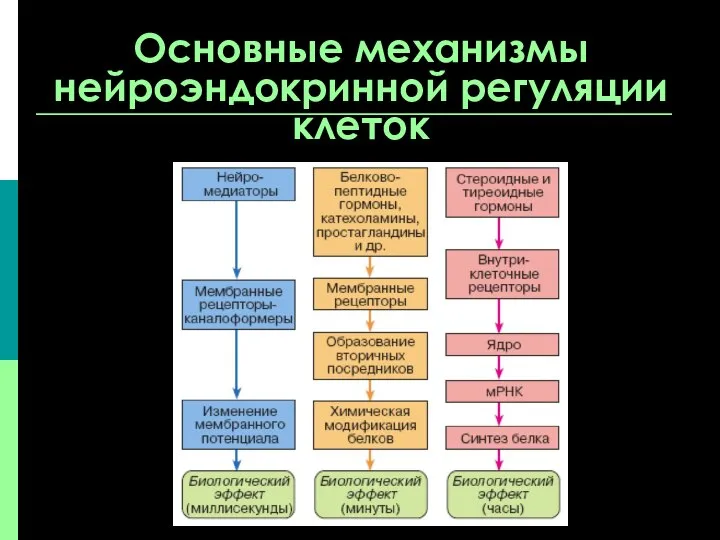
Основные механизмы нейроэндокринной регуляции клеток
Основные механизмы нейроэндокринной регуляции клеток

I. Центральные регуляторные образования эндокринной системы:
гипоталамус (нейросекреторные ядра);
гипофиз (аденогипофиз
I. Центральные регуляторные образования эндокринной системы:
гипоталамус (нейросекреторные ядра);
гипофиз (аденогипофиз

Транспорт гормонов осуществляется кровью в комплексе с белками.
Часть этих белков
Транспорт гормонов осуществляется кровью в комплексе с белками.
Часть этих белков

Образование связанной формы гормонов :
1. предохраняет организм от избыточного накопления в
Образование связанной формы гормонов :
1. предохраняет организм от избыточного накопления в

Механизмы действия гормонов
Механизмы действия гормонов
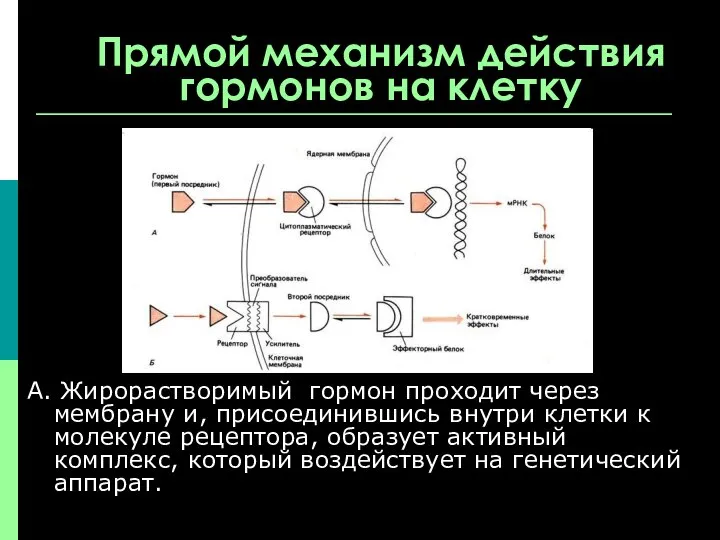
Прямой механизм действия гормонов на клетку
А. Жирорастворимый гормон проходит через мембрану
Прямой механизм действия гормонов на клетку
А. Жирорастворимый гормон проходит через мембрану

Ядерные рецепторы
Ядерные рецепторы - это ДНК-связывающие факторы транскрипции, локализованые в ядре.
В
Ядерные рецепторы
Ядерные рецепторы - это ДНК-связывающие факторы транскрипции, локализованые в ядре.
В
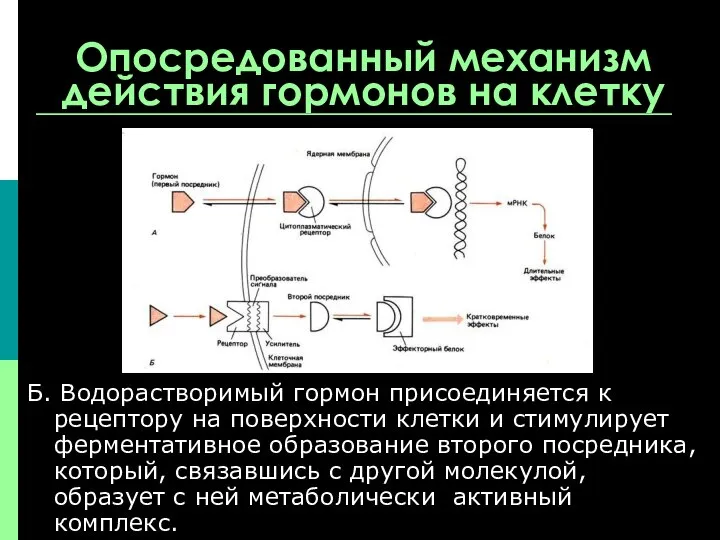
Опосредованный механизм действия гормонов на клетку
Б. Водорастворимый гормон присоединяется к рецептору
Опосредованный механизм действия гормонов на клетку
Б. Водорастворимый гормон присоединяется к рецептору

Характерной особенностью действия этих гормонов является быстрый эффект – обусловленный активацией
Характерной особенностью действия этих гормонов является быстрый эффект – обусловленный активацией

Вторичные мессенджеры
Вторичные мессенджеры

Требования предъявляемые
вторичному мессенджеру:
1. небольшая молекулярная масса (чтобы с высокой скоростью диффундировать
Требования предъявляемые
вторичному мессенджеру:
1. небольшая молекулярная масса (чтобы с высокой скоростью диффундировать

аденилатциклаза - цАМФ;
гуанилатциклаза - цГМФ;
фосфолипаза С - ИФ3;
Са2+.
Газовые посредники:
NO, СО, H2S
Известны
аденилатциклаза - цАМФ;
гуанилатциклаза - цГМФ;
фосфолипаза С - ИФ3;
Са2+.
Газовые посредники:
NO, СО, H2S
Известны
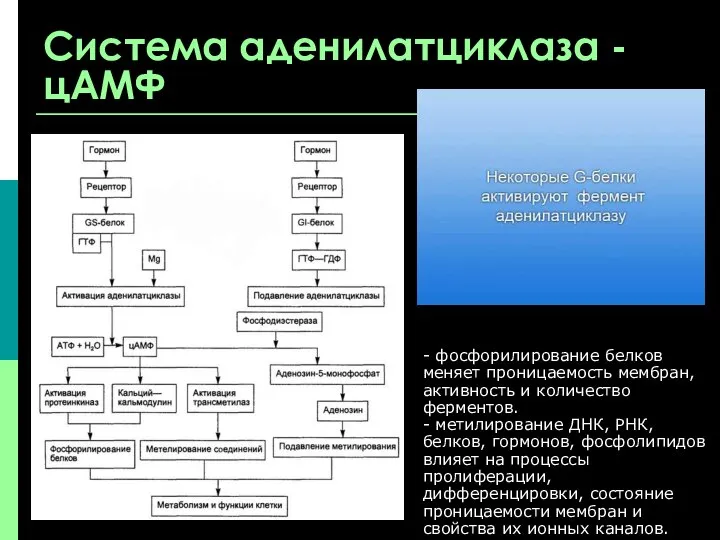
Система аденилатциклаза - цАМФ
- фосфорилирование белков меняет проницаемость мембран, активность
Система аденилатциклаза - цАМФ
- фосфорилирование белков меняет проницаемость мембран, активность
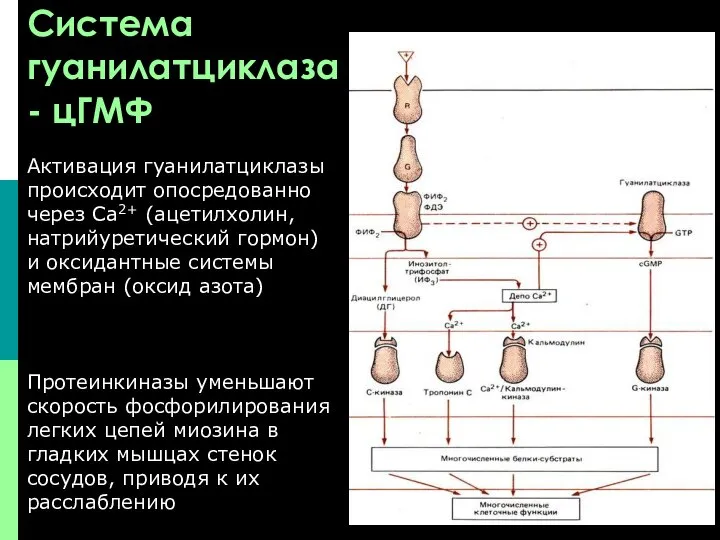
Система гуанилатциклаза - цГМФ
Активация гуанилатциклазы происходит опосредованно через Са2+ (ацетилхолин,
Система гуанилатциклаза - цГМФ
Активация гуанилатциклазы происходит опосредованно через Са2+ (ацетилхолин,

В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны
В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны
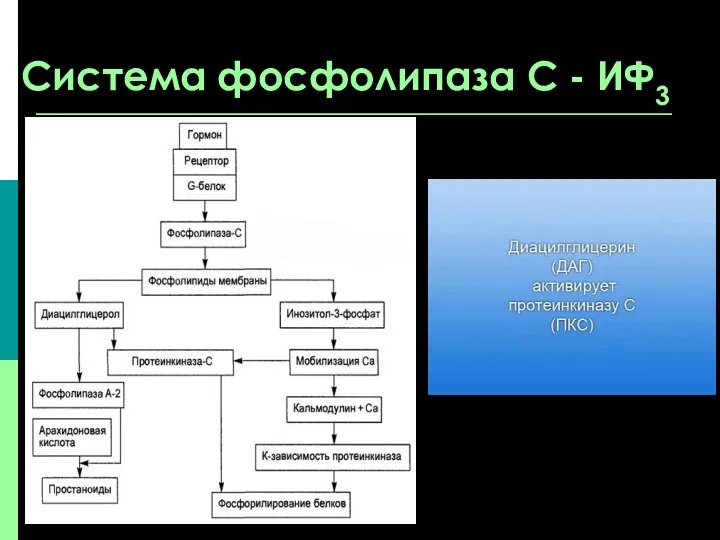
Система фосфолипаза С - ИФ3
Система фосфолипаза С - ИФ3
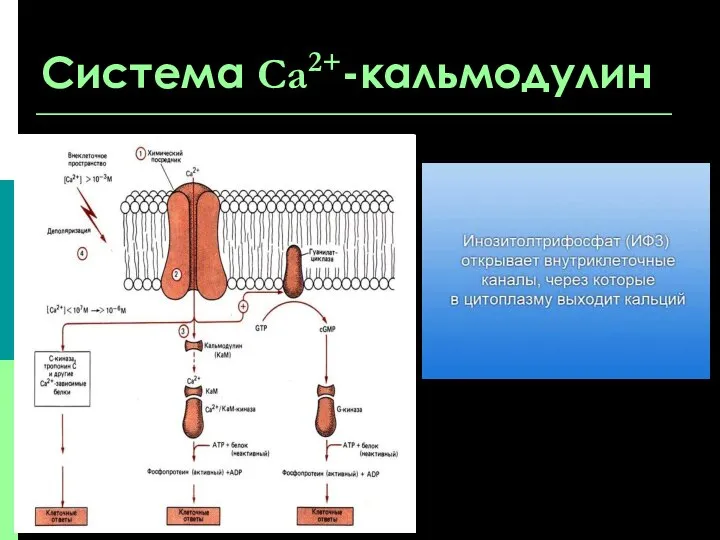
Система Са2+-кальмодулин
Система Са2+-кальмодулин
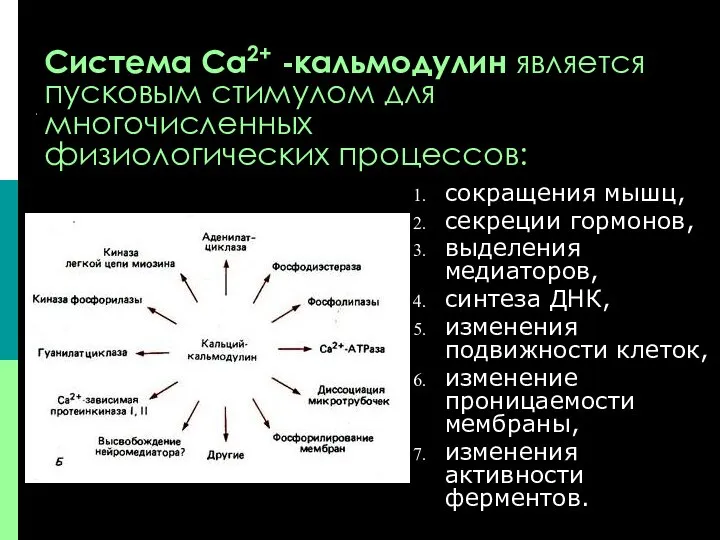
Система Са2+ -кальмодулин является
пусковым стимулом для
многочисленных
физиологических процессов:
сокращения мышц,
Система Са2+ -кальмодулин является
пусковым стимулом для
многочисленных
физиологических процессов:
сокращения мышц,

NO
как вторичный мессенджер
образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего
NO
как вторичный мессенджер
образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего

Механизм действия
В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в
Механизм действия
В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в

NO может либо усиливать интенсивность процессов, либо подавлять их.
Примером подавления процесса
NO может либо усиливать интенсивность процессов, либо подавлять их.
Примером подавления процесса
 Высшая нервная деятельность. Высшие мозговые функции
Высшая нервная деятельность. Высшие мозговые функции Дыхание животных
Дыхание животных Фрукты-гибриды
Фрукты-гибриды Виды декоративных деревьев и кустарников пришкольного учебно-опытного участка
Виды декоративных деревьев и кустарников пришкольного учебно-опытного участка Основы термодинамики живых организмов. (Лекция 11)
Основы термодинамики живых организмов. (Лекция 11) Роль биосферы в природе Презентация по биологии
Роль биосферы в природе Презентация по биологии  Обитатели Тайги
Обитатели Тайги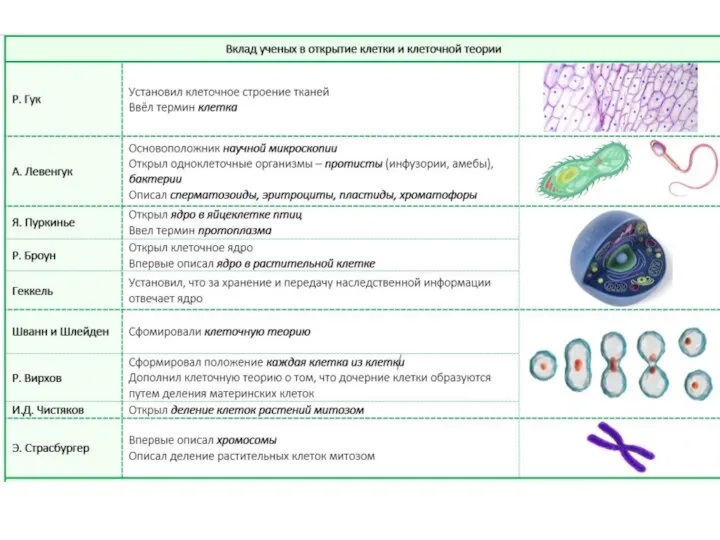 Клеточная теория. Основные положения современной клеточной теории
Клеточная теория. Основные положения современной клеточной теории Царство животных
Царство животных Антропосоциогенез 1.Понятие антропосоциогенеза. Современная наука об основных факторах, этапах и закономерностях антропосоциоге
Антропосоциогенез 1.Понятие антропосоциогенеза. Современная наука об основных факторах, этапах и закономерностях антропосоциоге Видоизмененные подземные побеги урок - исследование
Видоизмененные подземные побеги урок - исследование Ленивцы
Ленивцы Генетика. Грегор Иоганн Мендель
Генетика. Грегор Иоганн Мендель Основы молекулярной генетики. Генетические механизмы. Биосинтез белков и нуклеиновых кислот. (Лекция 3)
Основы молекулярной генетики. Генетические механизмы. Биосинтез белков и нуклеиновых кислот. (Лекция 3) Презентация на тему "Мир комнатных растений" - скачать презентации по Биологии
Презентация на тему "Мир комнатных растений" - скачать презентации по Биологии Сравнение процессов деления и биологическое значение митоза и мейоза
Сравнение процессов деления и биологическое значение митоза и мейоза Сердечно-сосудистая система
Сердечно-сосудистая система Организм человека
Организм человека Презентация на тему "Пион тонколистый" - скачать бесплатно презентации по Биологии
Презентация на тему "Пион тонколистый" - скачать бесплатно презентации по Биологии Анатомия, физиология и возрастные особенности пищеварения
Анатомия, физиология и возрастные особенности пищеварения Органы выделения насекомых
Органы выделения насекомых Эволюция систем регуляции
Эволюция систем регуляции Виды регуляции. Гуморальная регуляция
Виды регуляции. Гуморальная регуляция Генетика пола
Генетика пола Презентация на тему "Дихання" - скачать бесплатно презентации по Биологии
Презентация на тему "Дихання" - скачать бесплатно презентации по Биологии Широколиственные леса
Широколиственные леса Рациональное питание
Рациональное питание Разнообразие, распространение и значение растений
Разнообразие, распространение и значение растений