- Фотосинтетические пигменты

Содержание
- 2. Вещества, поглощающие видимый свет и запускающие химические реакции фотосинтеза называются пигментами. Фотосинтетические пигменты составляют 10-15% сухой
- 4. Поглощение пигментом кванта света определяется характером распределения электронов в молекуле данного пигмента. Какие именно длины волн
- 5. В переносе электронов при фотосинтезе вовлекается только небольшая часть молекул хлорофилла. Прочие его молекулы выполняют роль
- 6. Растительные организмы содержат несколько видов пигментов. Обычно в пластидах высших растений и водорослей встречаются все три
- 7. ХЛОРОФИЛЛ Молекула хлорофилла состоит из порфириновой «головки» и фитольного «хвоста». При этом порфириновая часть молекулы находится
- 8. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются феофетины.
- 9. СИНТЕЗ ХЛОРОФИЛЛА
- 10. СВОЙСТВА ХЛОРОФИЛЛА Резко выраженные максимумы поглощения хлорофиллов находятся в сине-фиолетовой (430-460нм) и красной (650-700нм) частях спектра
- 11. Раствор хлорофилла отражает свет с измененной длиной волны, поэтому хлорофилл приобретает вишнево-красную окраску. Это явление носит
- 12. В основе механизмов флуоресценции и фосфоресценции лежит способность пигментов находится в электронновозбужденном состоянии.
- 13. ФУНКЦИИ ХЛОРОФИЛЛА Молекула хлорофилла благодаря своим структурным и физико-химическим особенностям способна выполнять три важнейшие функции: избирательно
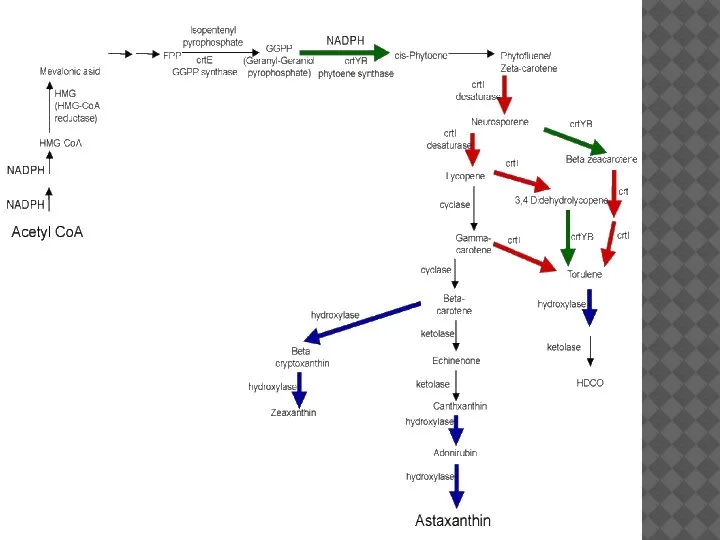
- 14. КАРОТИНОИДЫ Каротиноиды – это полиеновые соединения, имеющие систему сопряженных двойных связей. Они образуют 40-углеродную цепь, построенную
- 17. Спектры поглощения каротиноидов характеризуются двумя полосами в фиолетово-синей и синей области от 400 до 500 нм.
- 18. РОЛЬ КАРОТИНОИДОВ Они выполняют роль дополнительных пигментов, которые передают энергию поглощенных квантов хлорофиллу а для совершения
- 19. ФИКОБИЛИНЫ Фикобилины относятся к группе желчных пигментов – билинов. Это тетрапирролы с открытой цепью, имеющие систему
- 20. Фикобилины поглощают свет в желтой и зеленой областях спектра между двумя максимумами поглощения света хлорофиллов
- 21. Фикобилипротеины делятся на три основные группы: фикоэритрины – белки красного цвета с максимумом поглощения 498-568 нм;
- 22. РОЛЬ ФИКОБИЛИНОВ Фикобилипротеины агрегируют друг с другом, образуя специальные гранулы фикобилисомы. У цианобактерий, красных водорослей и
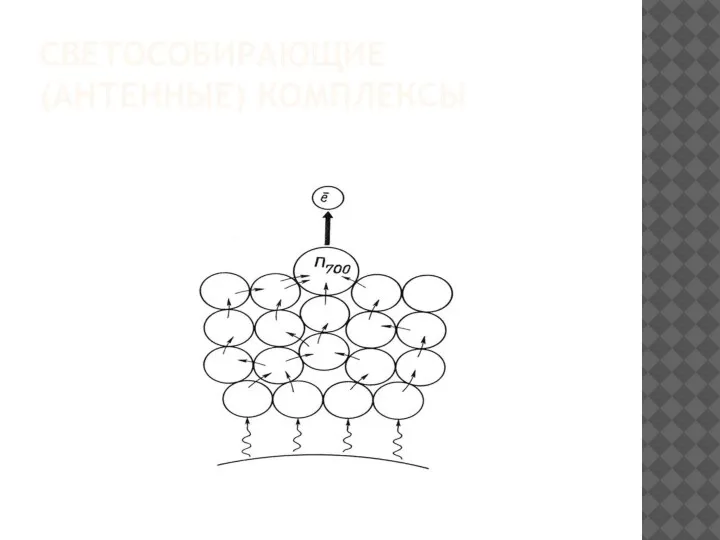
- 23. СВЕТОСОБИРАЮЩИЕ (АНТЕННЫЕ) КОМПЛЕКСЫ
- 25. Благодаря тому, что расстояние между пигментами в АК не превышает 10нм, передача энергии в реакционный центр
- 26. Необходимым условием для передачи энергии является перекрывание спектра флуоресценции молекулы-донора со спектром поглощения молекулы-акцептора. В АК
- 27. ССК ФСII содержит 40 молекул хлорофиллов а с максимумами поглощения 670-683 нм на один П680 и
- 29. РАЗДЕЛЕНИЕ ЗАРЯДОВ В ФОТОСИСТЕМАХ В реакционных центрах энергия возбуждения фотосинтетических пигментов используется для разделения зарядов. Разделение
- 30. В реакционном центре ФСI первичный донор электронов - Р700, а акцептор – мономерная форма хлорофилла а695
- 31. Процесс разделения зарядов, происходящий в реакционном центре фотосистемы, условно можно разделить на несколько этапов. На первом
- 32. На заключительном этапе (вторичное разделение зарядов), которое в среднем занимает около 200 пикосекунд, происходит процесс генерации
- 34. Скачать презентацию

Вещества, поглощающие видимый свет и запускающие химические реакции фотосинтеза называются
Вещества, поглощающие видимый свет и запускающие химические реакции фотосинтеза называются

Поглощение пигментом кванта света определяется характером распределения электронов в молекуле данного
Поглощение пигментом кванта света определяется характером распределения электронов в молекуле данного
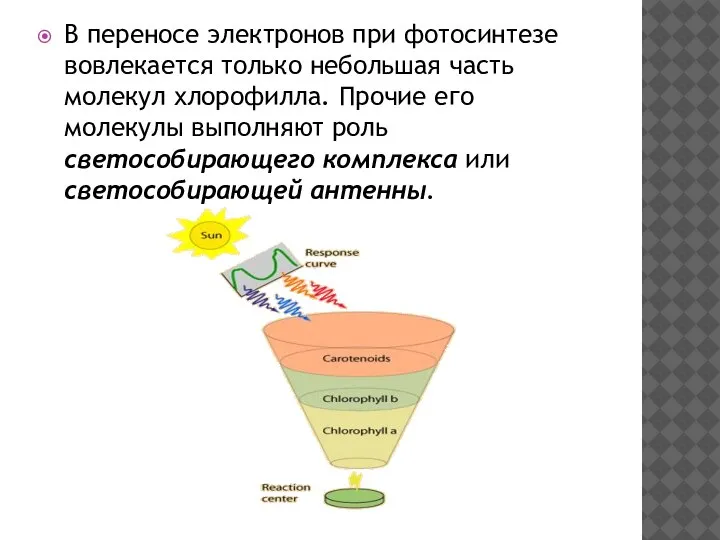
В переносе электронов при фотосинтезе вовлекается только небольшая часть молекул хлорофилла.
В переносе электронов при фотосинтезе вовлекается только небольшая часть молекул хлорофилла.
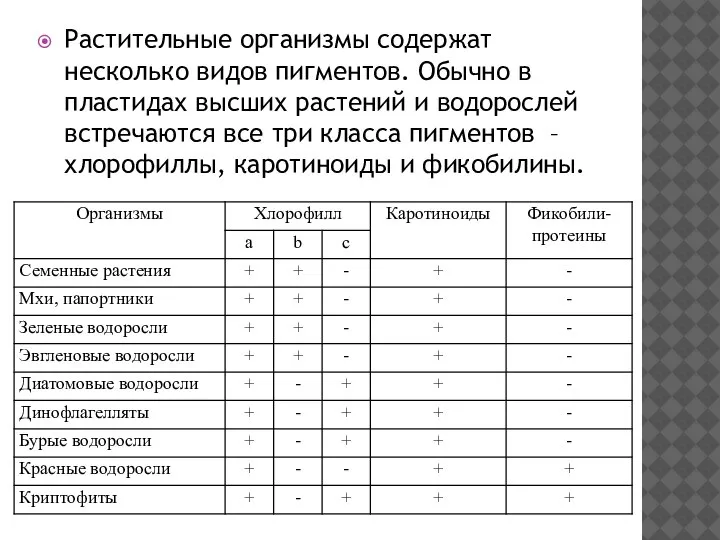
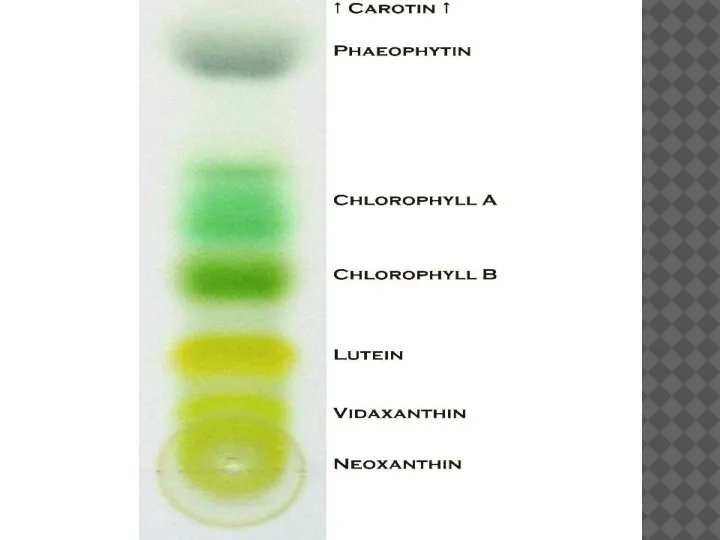
Растительные организмы содержат несколько видов пигментов. Обычно в пластидах высших растений
Растительные организмы содержат несколько видов пигментов. Обычно в пластидах высших растений

ХЛОРОФИЛЛ
Молекула хлорофилла состоит из порфириновой «головки» и фитольного «хвоста». При этом
ХЛОРОФИЛЛ
Молекула хлорофилла состоит из порфириновой «головки» и фитольного «хвоста». При этом
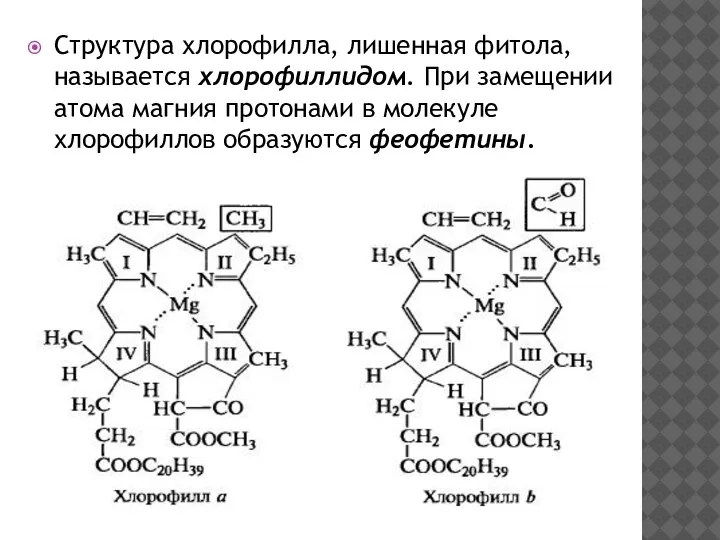
Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами
Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами
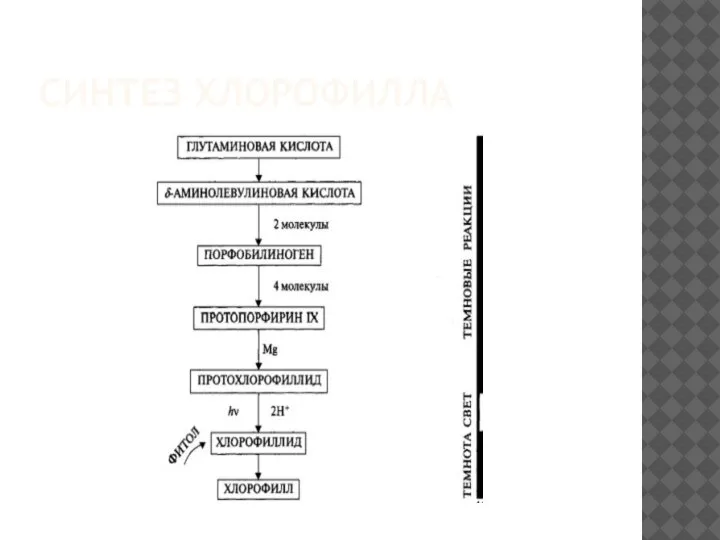
СИНТЕЗ ХЛОРОФИЛЛА
СИНТЕЗ ХЛОРОФИЛЛА
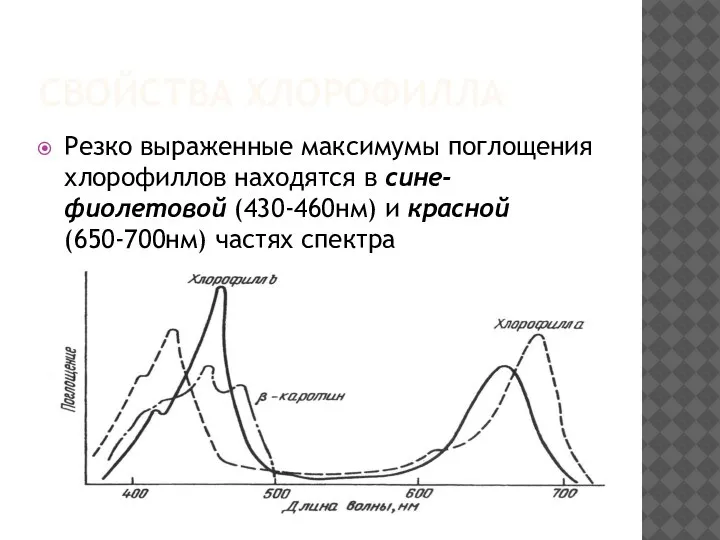
СВОЙСТВА ХЛОРОФИЛЛА
Резко выраженные максимумы поглощения хлорофиллов находятся в сине-фиолетовой (430-460нм) и
СВОЙСТВА ХЛОРОФИЛЛА
Резко выраженные максимумы поглощения хлорофиллов находятся в сине-фиолетовой (430-460нм) и
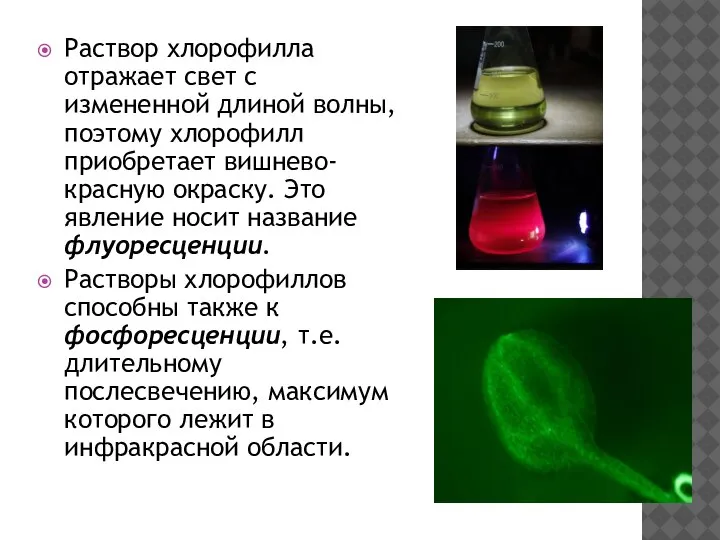
Раствор хлорофилла отражает свет с измененной длиной волны, поэтому хлорофилл приобретает
Раствор хлорофилла отражает свет с измененной длиной волны, поэтому хлорофилл приобретает
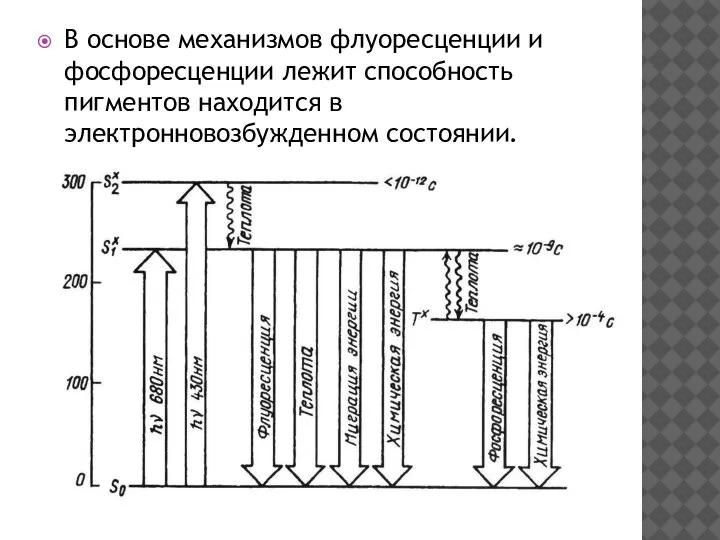
В основе механизмов флуоресценции и фосфоресценции лежит способность пигментов находится в
В основе механизмов флуоресценции и фосфоресценции лежит способность пигментов находится в

ФУНКЦИИ ХЛОРОФИЛЛА
Молекула хлорофилла благодаря своим структурным и физико-химическим особенностям способна выполнять
ФУНКЦИИ ХЛОРОФИЛЛА
Молекула хлорофилла благодаря своим структурным и физико-химическим особенностям способна выполнять

КАРОТИНОИДЫ
Каротиноиды – это полиеновые соединения, имеющие систему сопряженных двойных связей. Они
КАРОТИНОИДЫ
Каротиноиды – это полиеновые соединения, имеющие систему сопряженных двойных связей. Они

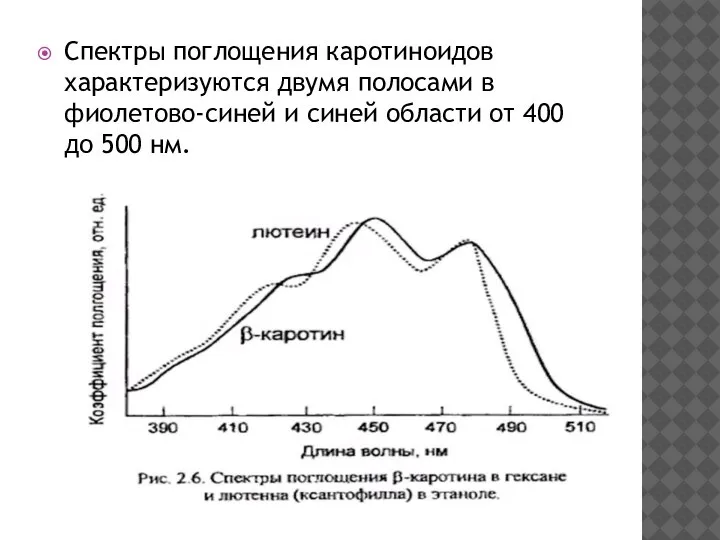
Спектры поглощения каротиноидов характеризуются двумя полосами в фиолетово-синей и синей области
Спектры поглощения каротиноидов характеризуются двумя полосами в фиолетово-синей и синей области

РОЛЬ КАРОТИНОИДОВ
Они выполняют роль дополнительных пигментов, которые передают энергию поглощенных квантов
РОЛЬ КАРОТИНОИДОВ
Они выполняют роль дополнительных пигментов, которые передают энергию поглощенных квантов

ФИКОБИЛИНЫ
Фикобилины относятся к группе желчных пигментов – билинов. Это тетрапирролы с
ФИКОБИЛИНЫ
Фикобилины относятся к группе желчных пигментов – билинов. Это тетрапирролы с
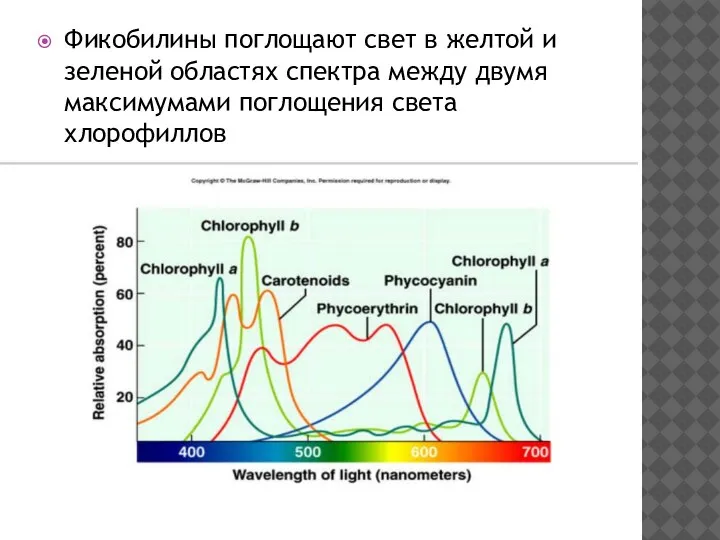
Фикобилины поглощают свет в желтой и зеленой областях спектра между двумя
Фикобилины поглощают свет в желтой и зеленой областях спектра между двумя

Фикобилипротеины делятся на три основные группы:
фикоэритрины – белки красного цвета
Фикобилипротеины делятся на три основные группы:
фикоэритрины – белки красного цвета

РОЛЬ ФИКОБИЛИНОВ
Фикобилипротеины агрегируют друг с другом, образуя специальные гранулы фикобилисомы. У
РОЛЬ ФИКОБИЛИНОВ
Фикобилипротеины агрегируют друг с другом, образуя специальные гранулы фикобилисомы. У
СВЕТОСОБИРАЮЩИЕ (АНТЕННЫЕ) КОМПЛЕКСЫ
СВЕТОСОБИРАЮЩИЕ (АНТЕННЫЕ) КОМПЛЕКСЫ


Благодаря тому, что расстояние между пигментами в АК не превышает 10нм,
Благодаря тому, что расстояние между пигментами в АК не превышает 10нм,

Необходимым условием для передачи энергии является перекрывание спектра флуоресценции молекулы-донора со
Необходимым условием для передачи энергии является перекрывание спектра флуоресценции молекулы-донора со

ССК ФСII содержит 40 молекул хлорофиллов а с максимумами поглощения 670-683
ССК ФСII содержит 40 молекул хлорофиллов а с максимумами поглощения 670-683


РАЗДЕЛЕНИЕ ЗАРЯДОВ В ФОТОСИСТЕМАХ
В реакционных центрах энергия возбуждения фотосинтетических пигментов используется
РАЗДЕЛЕНИЕ ЗАРЯДОВ В ФОТОСИСТЕМАХ
В реакционных центрах энергия возбуждения фотосинтетических пигментов используется

В реакционном центре ФСI первичный донор электронов - Р700, а акцептор
В реакционном центре ФСI первичный донор электронов - Р700, а акцептор

Процесс разделения зарядов, происходящий в реакционном центре фотосистемы, условно можно разделить
Процесс разделения зарядов, происходящий в реакционном центре фотосистемы, условно можно разделить

На заключительном этапе (вторичное разделение зарядов), которое в среднем занимает около
На заключительном этапе (вторичное разделение зарядов), которое в среднем занимает около
 Введение в медицинскую генетику
Введение в медицинскую генетику Презентация по биологии Отдел моховидные
Презентация по биологии Отдел моховидные  Развитие осевого скелета. Тема 3
Развитие осевого скелета. Тема 3 Цікаве про птахів Цікаве про птахів Птахи “жують” шлунком. Більшість птахів не мають зубів. Зазвичай вони ковтають гальку або гравій, які енергійно переміщуються в шлунку і сприяють перемелюванню їжі. Пташеняті може знадобитися два дні, щоб вибра
Цікаве про птахів Цікаве про птахів Птахи “жують” шлунком. Більшість птахів не мають зубів. Зазвичай вони ковтають гальку або гравій, які енергійно переміщуються в шлунку і сприяють перемелюванню їжі. Пташеняті може знадобитися два дні, щоб вибра Значение грибов и грибоподобных организмов в природе
Значение грибов и грибоподобных организмов в природе Теории возникновения жизни на Земле
Теории возникновения жизни на Земле Адаптация организмов к условиям обитания как результат действия естественного отбора
Адаптация организмов к условиям обитания как результат действия естественного отбора Презентация на тему Развитие половых клеток
Презентация на тему Развитие половых клеток Мейоз
Мейоз Введение в фитотоксикологию
Введение в фитотоксикологию Голосеменные растения
Голосеменные растения Кожа. Дерматология как наука
Кожа. Дерматология как наука Презентация на тему Экологические ситуации
Презентация на тему Экологические ситуации  Опорно-двигательный аппарат. Мышечная система. Осанка
Опорно-двигательный аппарат. Мышечная система. Осанка «Разнообразие птиц и их значение» Автор: Зиновьева Ольга, Максимук Марина ученицы 10 «В» класса
«Разнообразие птиц и их значение» Автор: Зиновьева Ольга, Максимук Марина ученицы 10 «В» класса Микроскопия. Строение микроскопа
Микроскопия. Строение микроскопа Photosynthesis
Photosynthesis Неживая и живая природа (2 класс)
Неживая и живая природа (2 класс) Знания, полученные во время исследований, помогают сохранять серых китов
Знания, полученные во время исследований, помогают сохранять серых китов Появление первичных живых организмов. Возникновение биосферы
Появление первичных живых организмов. Возникновение биосферы Белки, их строение,свойства, биологические функции
Белки, их строение,свойства, биологические функции Основы медицинской экологии
Основы медицинской экологии Движение крови по сосудам 8 класс - Презентация_
Движение крови по сосудам 8 класс - Презентация_ Общая характеристика подцарства Низшие растения (Thallobionta)
Общая характеристика подцарства Низшие растения (Thallobionta) Борьба за существование Формы борьбы за существование
Борьба за существование Формы борьбы за существование  Flora of the river Vodla
Flora of the river Vodla Эндокринная система
Эндокринная система Типы питания и дыхания бактерий
Типы питания и дыхания бактерий