- Генетика развития растений. Меристемы растений

Содержание
- 2. Меристемы растений Меристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные ткани, или меристематические ткани, —
- 3. Меристемы растений АМ соцветия Вегетативная АМ побега Меристема цветка Апикальные меристемы (АМ) Латеральные меристемым Камбий камбий
- 4. Первичные - закладываются в эмбриогенезе 3 - xylem pericycle lineage - phloem pericycle lineage Вторичные –
- 5. АМ корня SC – stem cells стволовые клетки Стволовые клетки в апикальных меристемах побега и корня
- 6. Стволовые клетки (stem cells): Делящиеся клетки, потомки которых либо остаются в том же состоянии, что и
- 7. Организация ниши стволовых клеток у животных и у растений Ниша стволовых клеток в яичнике Drosophila melanogaster
- 8. Общий принцип организации ниши стволовых клеток дифференцированные клетки ранние стадии дифференцировки недифференцированные клетки внутриклеточный сигнал, предотвращающий
- 9. мультипотентные плюрипотентные II III I олигопотентные Стволовые клетки: Обладают высокой потентностью 2. Способны к асимметричным делениям
- 10. Стволовая клетка Дифференцированная клетка Пополнение популяции столовых клеток Асимметричное деление стволовых клеток Асимметричное деление
- 11. Генетический контроль структуры и функции апикальной меристемы побега В отличие от животных, растения всю жизнь продуцируют
- 12. ПАМ лист узел междоузлие фитомер Clark, Current Opinion in Plant Biology 2001 Строение апикальной меристемы побега
- 13. Строение апикальной меристемы побега CZ (central zone, центральная зона) –стволовые клетки (в т.ч. организующий центр) и
- 14. «Симпластические поля» в апикальной меристеме побега
- 15. Скорость прохождения клеточного цикла в разных зонах ПАМ Besnard et al., 2011
- 16. Распределение гормонов в ПАМ
- 17. Взаимодействие факторов, регулирующих активность ПАМ
- 18. Изучение генетического контроля структуры и функции ПАМ методами классической («прямой») генетики: Выявлены два классов мутантов, имеющих
- 19. Мутанты с недоразвитием АМ WT wus stm
- 20. stm-5/+ x wus-1/+ 3 stm-5/+ 9 +/+ 3 wus-1/+ 1 stm-5/ wus-1 : : : По
- 21. Изучение взаимодействия генов STM и WUS: Анализ трансгенных растений с измененной экспрессией генов: ANT::STM (эктопическая экспрессия
- 22. Поддержание пула стволовых клеток в меристеме Пролиферация стволовых клеток в меристеме Роль генов STM и WUS
- 23. Мутанты по гену WUS (WUSCHEL) дикий тип wus Mayer et al., Cell 1998 в эмбриогенезе АМ
- 24. stm-1, stm-5 - аллели с высокой экспрессивностью мутантного признака («жесткие, суровые»): -АМ не закладывается в эмбриогенезе;
- 25. Мутанты с увеличенной АМ WT clv1
- 26. Мутанты по генам СLV1, CLV2, CLV3 (CLAVATA) многолистная розетка увеличение числа органов цветка фасциация стебля увеличение
- 27. Экспрессия генов STM, WUS, CLV1 и CLV3 в апикальной меристеме побега WUS – в ОЦ CLV1
- 28. Экспрессия генов STM и WUS в апикальной меристеме побега Bowman J., Yuval E. Trends in Plant
- 29. Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы Гены семейства KNOX арабидопсиса Гены
- 30. Гомеодомен-ДНК - комплекс вариабельный уч. гомеодомен N С Структура транскрипционного фактора с ДНК-связывающим гомеодоменом «Атипичный» гомеодомен
- 31. Экспрессия гена STM в АМ побега Moussian et al., The EMBO Journal 1998 Выявление экспрессии гена
- 32. Транспорт продуктов генов KNOX в апикальной меристеме побега
- 33. Нарушение транспорта белка STM вызывает дефекты развития ПАМ Plant Journal, 2017
- 34. ТФ BELL (тоже из семейства TALE) – партнеры ТФ KNOX (обеспечивают транспорт KNOX в ядро)
- 35. Мишени STM Экспрессия гена AS1 (Asymmetric leaves1), отвечающего за развитие листового примордия, у мутантов stm-1 stm-1
- 36. Модель эпигенетической регуляции генов KNOX в дифференцированных клетках листьев wt as1 hira Lodha et al., 2013
- 37. STM активирует экспрессию гена CUC1 Гены CUC экспрессируются на границе между листовым примордием и меристемой Spinelli
- 38. Мишени STM в ПАМ STM STM IPT (биосинтез цитокинина) GA2ox (дезактивация гибереллинов) Jasinski et al. 2005
- 39. Взаимодействие STM (генов KNOX) и генов, вовлеченных в метаболизм цитокинина и гиббереллина в апексе побега Мишени
- 40. Активация экспрессии KNOX важна для формирования сложных листьев
- 41. Транскрипционные факторы семейства WOX WUS-бокс (репрессия транскрипции) домен EAR (рекрутирование корепрессоров трнаскрипции) гомеодомен «древняя» клада «промежуточная»
- 42. Ядерная локализация гибридного белка WUS-GUS (в клетках эпидермиса лука) Экспрессия гена WUS в АМ побега Экспрессия
- 43. мхи зеленые водоросли WUS/WOX9 про-ортолог линия WUS линия WOX9 линия WOX13 WOX13 Плауны Семенные растения Папоротники
- 44. Геном Gnetum gnemon – содержит про-ортолог GgWUS/WOX5 (до дупликации генов WUS/WOX5 в ходе эволюции), экспрессирующийся как
- 45. Взаимодействие WUS и системы генов CLAVATA увеличение числа органов цветка фасциация стебля увеличение числа плодолистиков Плод
- 46. CLV1 и CLV2 кодируют рецепторные белки (c лейцин-богатым LRR–доменом), работающие в виде гетеродимера CLV1/CLV2 CLV3 –
- 47. Гены-антогонисты - частое явление в генетике развития. По-видимому, они позволяют осуществлять тонкую регуляцию процессов развития. Дикий
- 48. дикий тип мутант clv3 35S::CLV3 CLV3 WUS CLV3 WUS Увеличение домена экспрессии CLV3 в растениях ANT::WUS
- 49. Экспрессия генов CLV1, CLV3 и WUS в апикальной меристеме побега Экспрессия WUS в ОЦ CLV1 «защищает»
- 50. Рецепторные киназы растений c LRR–доменом (Leucine-rich repeat receptor kinases) Haffani et al., Can. J. Bot. 2004
- 51. Эксперименты по выявлению новых компонентов сигнального пути, опосредованного CLV3 Получение новых мутаций на фоне сверхэкспрессии CLV3
- 52. TDIF CLV3 – член семейства CLE-пептидов В А Тип:
- 53. Участие CLE-пептидов в развитии Arabidopsis Jun J et al. Plant Physiol. 2010;154:1721-1736 ©2010 by American Society
- 54. Экспрессия CLE-пептидов в ПАМ
- 55. Стволовые клетки WUS ? Каким образом WUS регулирует поддержание запаса стволовых клеток?
- 56. Yadav et al., 2011 Белок WUS мигрирует в вышележащие слои центральной зоны меристемы, где расположены стволовые
- 57. WUS TAAT TAAT TAAT -1080 п.о. Активация транскрипции гена CLV3 Yadav et al., 2011 ТФ WUS
- 58. WUS STM LFY Мишени ТФ WUS Создание трансгенных растений с регулируемой (индуцибельной) экспрессией WUS (WUS под
- 59. Yadav et al., 2013 WUS негативно регулирует экспрессию генов многих ТФ, регулирующих дифференцировку клеток и развитие
- 60. Busch et al., 2010 Выявление мишеней ТФ WUS в транскриптомных исследованиях WUS CLV1 ауксиновый сигнальный путь
- 61. WUS негативно регулирует экспрессию генов ARR А-типа (гены первичного ответа на цитокинин): -1000 по ARR7 WUS
- 62. WUS негативно регулирует экспрессию генов LOG, кодирующих цитокинин-активирующие ферменты Активность пролиферации клеток ПАМ зависит от баланса
- 63. Биосинтез и сигнальный путь цитокининов: WUS WUS роль WUS в их регуляции
- 64. WUS STM AtIPT Цитокининовый ответ цитокинин Взаимодействие транскрипционных факторов STM и WUS и цитокининов Ауксиновый ответ
- 65. Роль гормонов в ПАМ
- 66. DR5:GFP Ауксины как регуляторы формирования листовых примордиев
- 67. Локальная экспрессия генов, кодирующих ТФ Локальное изменение концентрации гормона (или чувствительности клеток к действию гормона) в
- 68. Эпигенетическая регуляция экспрессии WUS с участием факторов ремоделирования хроматина
- 69. fas1-1 fas2-1 wt wt Нарушение организации ПАМ у мутантов по генам FASCIATA FAS1 и FAS2 кодируют
- 70. WUS CLV3 before just after 2 days after При выжигании зон экспрессии WUS или CLV3 они
- 71. Зоны экспрессии генов в «модельной» меристеме Centering the Organizing Center in the Arabidopsis thaliana Shoot Apical
- 72. Регуляции функции меристемы с участием сигналов от дифференцированных клеток дикий тип ham (hairy meristem) HAM HAM
- 73. Взаимодействие ТФ WOX и HAM в меристемах ТФ WOX и НАМ специфично взаимодействуют в меристемах: в
- 74. Филлотаксис Золотое сечение (1,61803…) Золотой угол (137,5°…) – «золотое сечение» окружности
- 75. Модели филлотаксиса «Ауксиновая» модель «Механическая» модель MAP4 (Microtubule Associated Protein4):GFP
- 76. Giacomo et al., 2013 Взаимодействие факторов, регулирующих активность ПАМ
- 78. Скачать презентацию

Меристемы растений
Меристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные ткани, или меристематические ткани, — обобщающее название для тканей растений, состоящих
Меристемы растений
Меристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные ткани, или меристематические ткани, — обобщающее название для тканей растений, состоящих
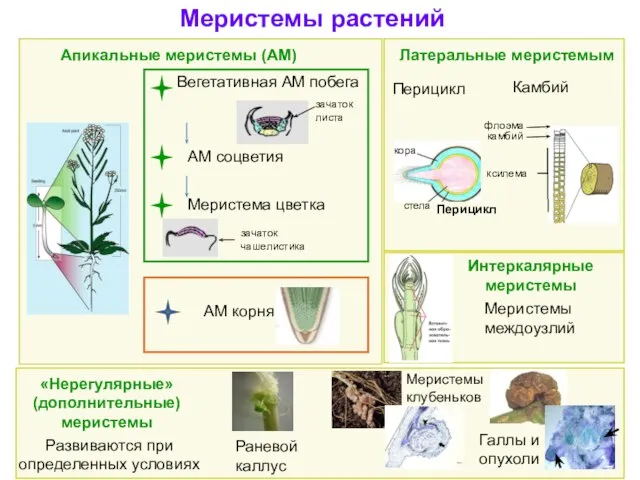
Меристемы растений
АМ соцветия
Вегетативная АМ побега
Меристема цветка
Апикальные меристемы (АМ)
Латеральные меристемым
Камбий
камбий
ксилема
флоэма
АМ корня
зачаток
чашелистика
зачаток листа
Интеркалярные
Меристемы растений
АМ соцветия
Вегетативная АМ побега
Меристема цветка
Апикальные меристемы (АМ)
Латеральные меристемым
Камбий
камбий
ксилема
флоэма
АМ корня
зачаток
чашелистика
зачаток листа
Интеркалярные
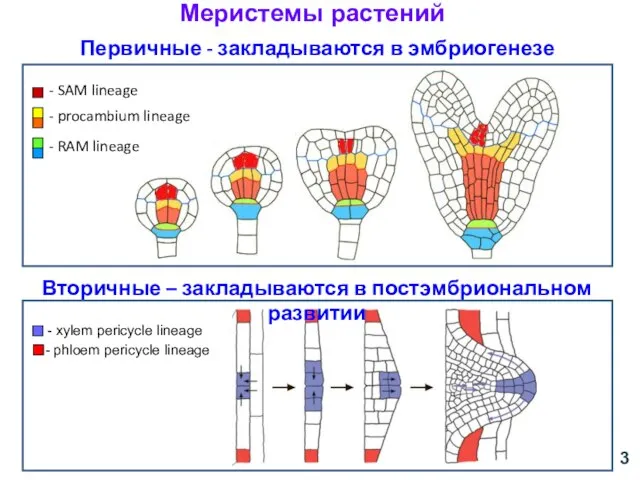
Первичные - закладываются в эмбриогенезе
3
- xylem pericycle lineage
- phloem pericycle
Первичные - закладываются в эмбриогенезе
3
- xylem pericycle lineage
- phloem pericycle
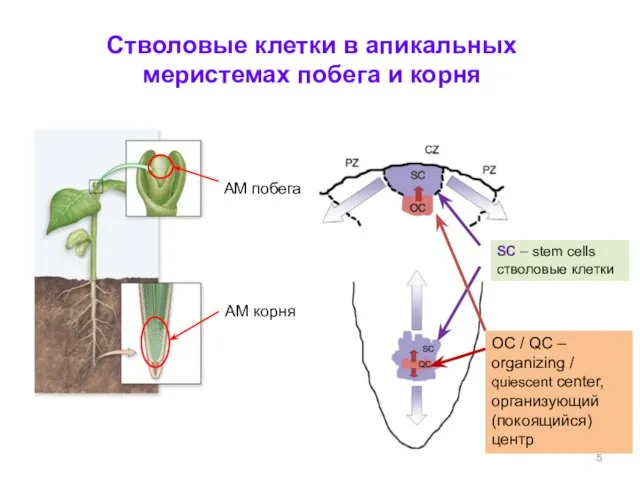
АМ корня
SC – stem cells
стволовые клетки
Стволовые клетки в апикальных меристемах побега
АМ корня
SC – stem cells
стволовые клетки
Стволовые клетки в апикальных меристемах побега

Стволовые клетки (stem cells):
Делящиеся клетки, потомки которых либо остаются в
Стволовые клетки (stem cells):
Делящиеся клетки, потомки которых либо остаются в

Организация ниши стволовых клеток
у животных и у растений
Ниша стволовых клеток
в
Организация ниши стволовых клеток
у животных и у растений
Ниша стволовых клеток
в
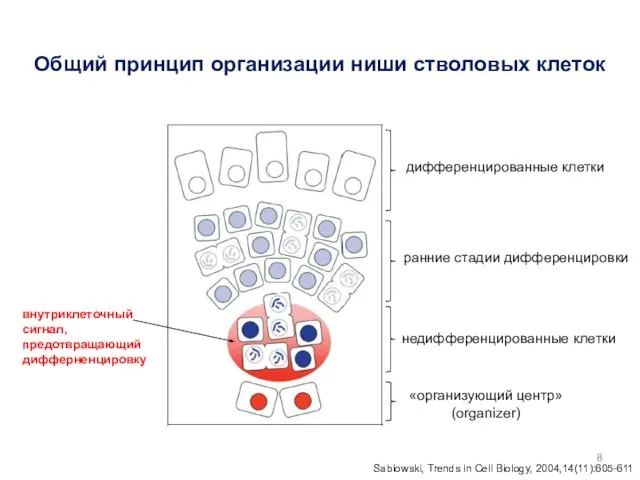
Общий принцип организации ниши стволовых клеток
дифференцированные клетки
ранние стадии дифференцировки
недифференцированные клетки
внутриклеточный
сигнал,
Общий принцип организации ниши стволовых клеток
дифференцированные клетки
ранние стадии дифференцировки
недифференцированные клетки
внутриклеточный
сигнал,
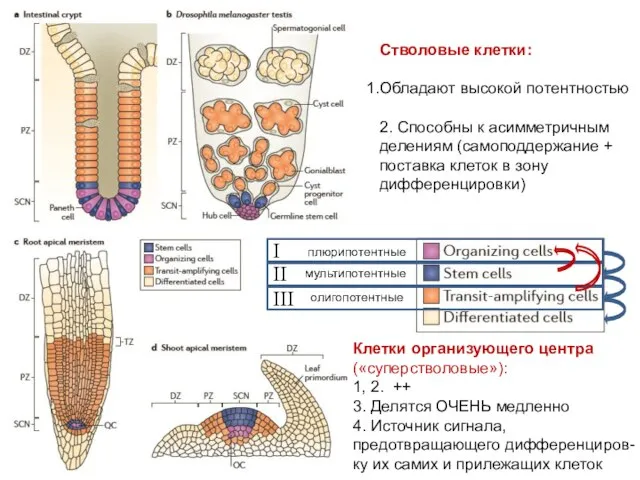
мультипотентные
плюрипотентные
II
III
I
олигопотентные
Стволовые клетки:
Обладают высокой потентностью
2. Способны к асимметричным делениям (самоподдержание + поставка
мультипотентные
плюрипотентные
II
III
I
олигопотентные
Стволовые клетки:
Обладают высокой потентностью
2. Способны к асимметричным делениям (самоподдержание + поставка

Стволовая клетка
Дифференцированная клетка
Пополнение популяции столовых клеток
Асимметричное деление стволовых клеток
Асимметричное деление
Стволовая клетка
Дифференцированная клетка
Пополнение популяции столовых клеток
Асимметричное деление стволовых клеток
Асимметричное деление
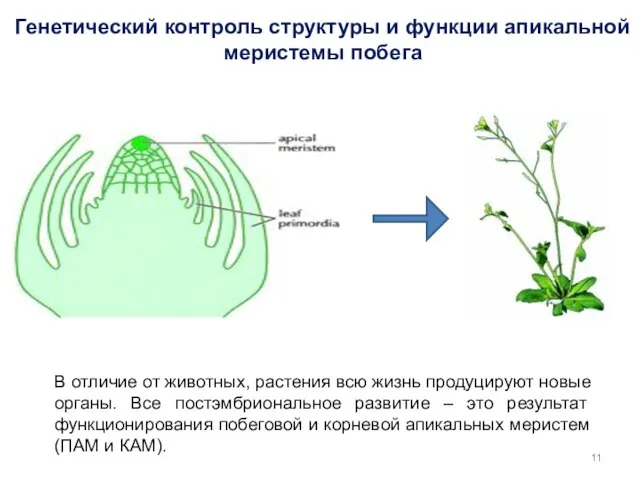
Генетический контроль структуры и функции апикальной меристемы побега
В отличие от животных,
Генетический контроль структуры и функции апикальной меристемы побега
В отличие от животных,

ПАМ
лист
узел
междоузлие
фитомер
Clark, Current Opinion in Plant Biology 2001
Строение апикальной меристемы побега
ПАМ
лист
узел
междоузлие
фитомер
Clark, Current Opinion in Plant Biology 2001
Строение апикальной меристемы побега

Строение апикальной меристемы побега
CZ (central zone, центральная зона) –стволовые клетки (в
Строение апикальной меристемы побега
CZ (central zone, центральная зона) –стволовые клетки (в

«Симпластические поля» в апикальной меристеме побега
«Симпластические поля» в апикальной меристеме побега
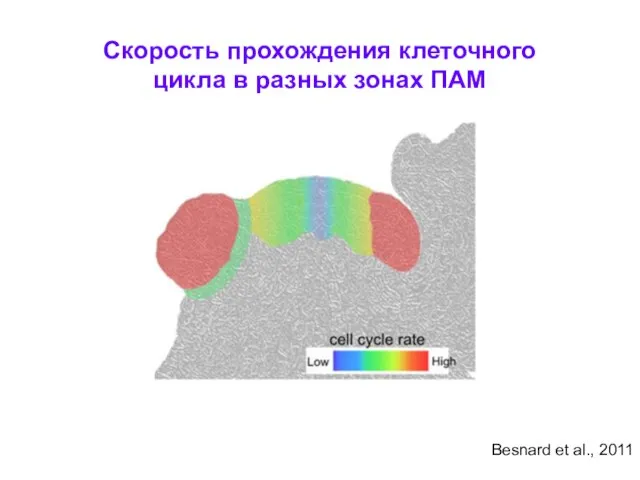
Скорость прохождения клеточного цикла в разных зонах ПАМ
Besnard et al., 2011
Скорость прохождения клеточного цикла в разных зонах ПАМ
Besnard et al., 2011
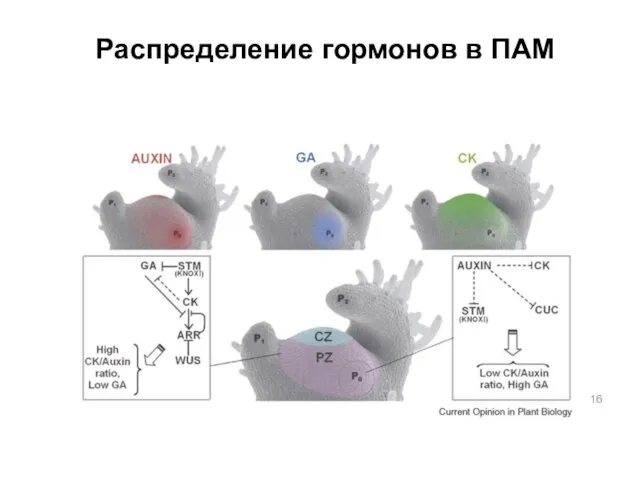
Распределение гормонов в ПАМ
Распределение гормонов в ПАМ
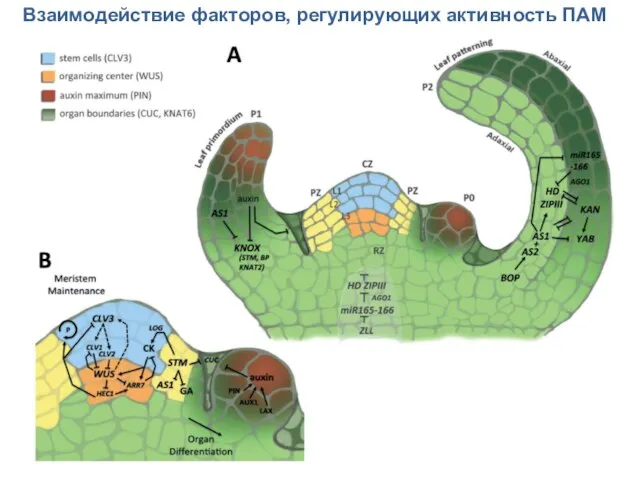
Взаимодействие факторов, регулирующих активность ПАМ
Взаимодействие факторов, регулирующих активность ПАМ

Изучение генетического контроля структуры и функции ПАМ методами классической («прямой») генетики:
Выявлены
Изучение генетического контроля структуры и функции ПАМ методами классической («прямой») генетики:
Выявлены
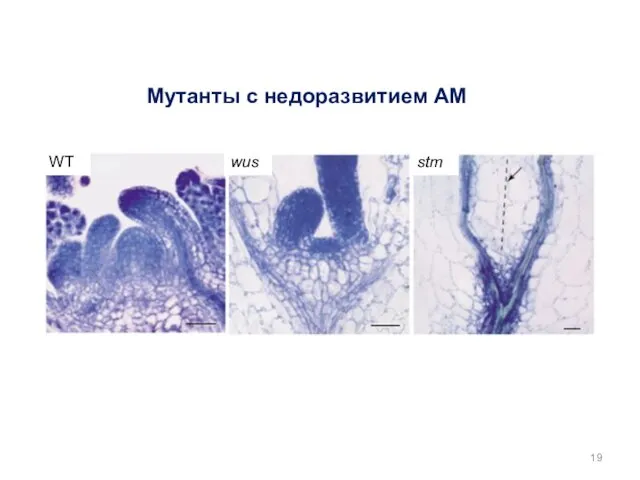
Мутанты с недоразвитием АМ
WT
wus
stm
Мутанты с недоразвитием АМ
WT
wus
stm

stm-5/+ x wus-1/+
3 stm-5/+
9 +/+
3 wus-1/+
1 stm-5/ wus-1
:
:
:
По генотипу
По фенотипу
4
stm-5/+ x wus-1/+
3 stm-5/+
9 +/+
3 wus-1/+
1 stm-5/ wus-1
:
:
:
По генотипу
По фенотипу
4

Изучение взаимодействия генов STM и WUS:
Анализ трансгенных растений с измененной экспрессией
Изучение взаимодействия генов STM и WUS:
Анализ трансгенных растений с измененной экспрессией
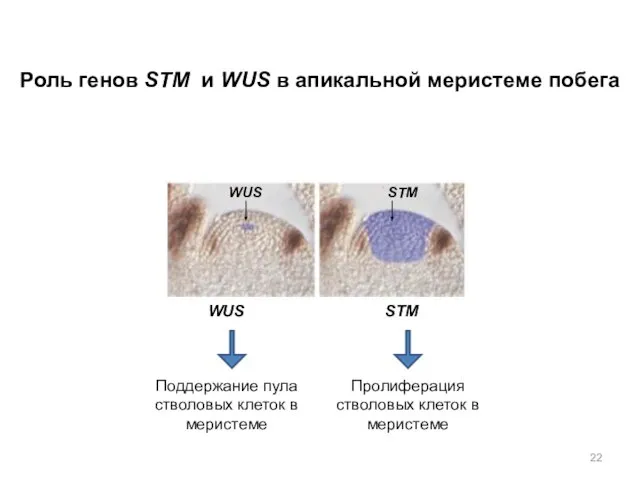
Поддержание пула стволовых клеток в меристеме
Пролиферация
стволовых клеток в меристеме
Роль генов STM
Поддержание пула стволовых клеток в меристеме
Пролиферация
стволовых клеток в меристеме
Роль генов STM

Мутанты по гену WUS (WUSCHEL)
дикий
тип
wus
Mayer et al., Cell 1998
в эмбриогенезе
Мутанты по гену WUS (WUSCHEL)
дикий
тип
wus
Mayer et al., Cell 1998
в эмбриогенезе
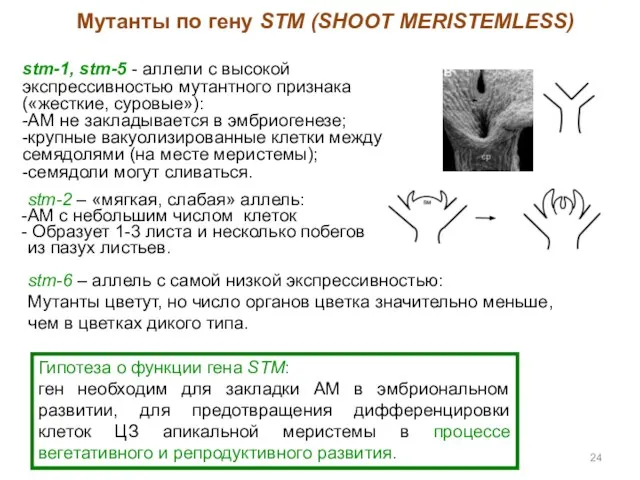
stm-1, stm-5 - аллели с высокой экспрессивностью мутантного признака («жесткие, суровые»):
stm-1, stm-5 - аллели с высокой экспрессивностью мутантного признака («жесткие, суровые»):
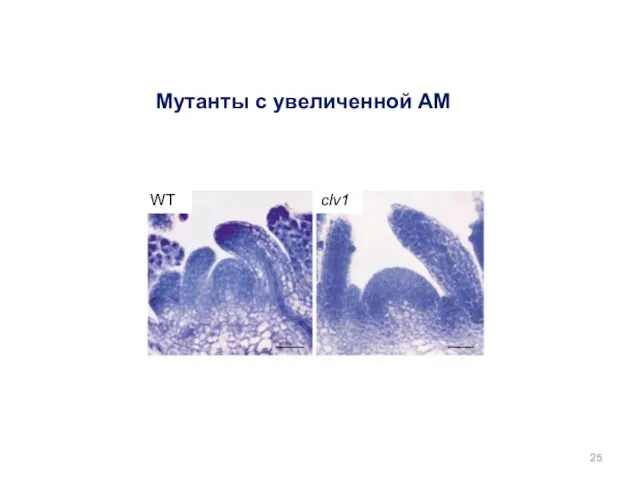
Мутанты с увеличенной АМ
WT
clv1
Мутанты с увеличенной АМ
WT
clv1

Мутанты по генам СLV1, CLV2, CLV3 (CLAVATA)
многолистная розетка
увеличение числа органов цветка
фасциация
Мутанты по генам СLV1, CLV2, CLV3 (CLAVATA)
многолистная розетка
увеличение числа органов цветка
фасциация
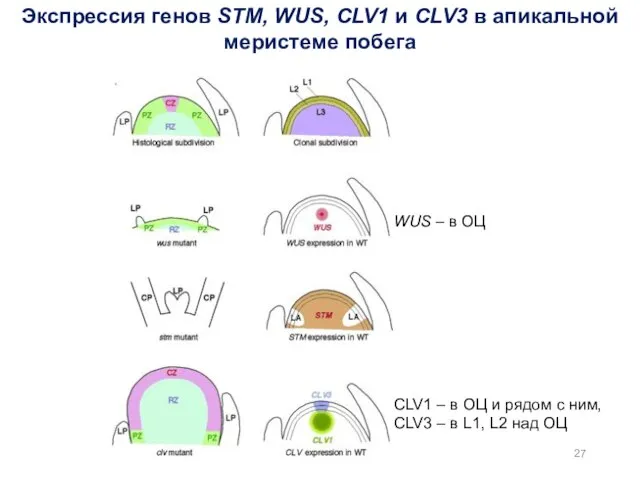
Экспрессия генов STM, WUS, CLV1 и CLV3 в апикальной меристеме побега
WUS
Экспрессия генов STM, WUS, CLV1 и CLV3 в апикальной меристеме побега
WUS

Экспрессия генов STM и WUS в апикальной меристеме побега
Bowman J., Yuval
Экспрессия генов STM и WUS в апикальной меристеме побега
Bowman J., Yuval
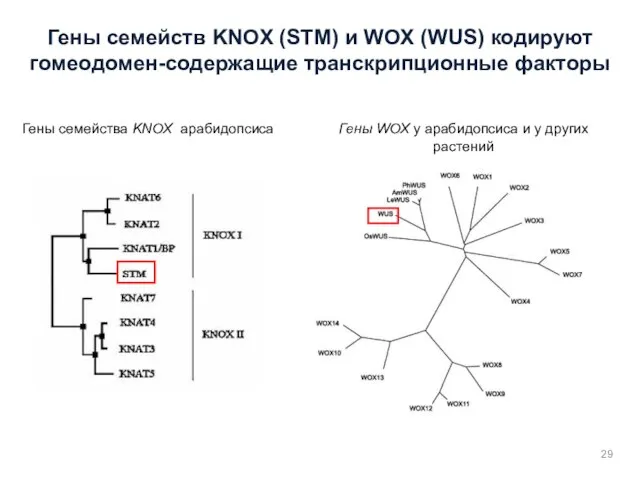
Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы
Гены
Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы
Гены

Гомеодомен-ДНК - комплекс
вариабельный уч.
гомеодомен
N
С
Структура транскрипционного фактора с ДНК-связывающим гомеодоменом
«Атипичный» гомеодомен ТФ
Гомеодомен-ДНК - комплекс
вариабельный уч.
гомеодомен
N
С
Структура транскрипционного фактора с ДНК-связывающим гомеодоменом
«Атипичный» гомеодомен ТФ
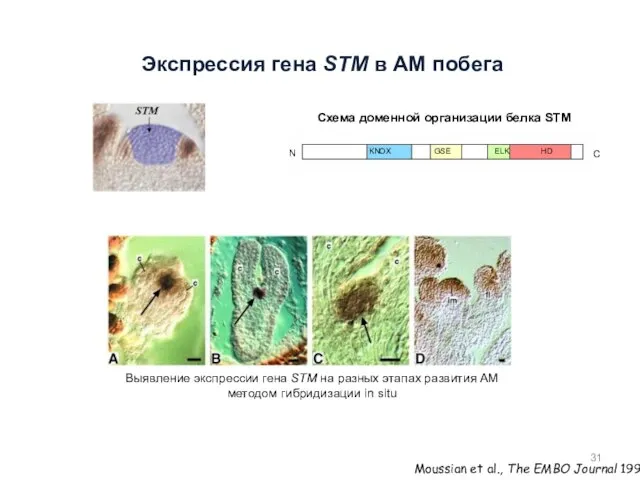
Экспрессия гена STM в АМ побега
Moussian et al., The EMBO Journal
Экспрессия гена STM в АМ побега
Moussian et al., The EMBO Journal

Транспорт продуктов генов KNOX в апикальной меристеме побега
Транспорт продуктов генов KNOX в апикальной меристеме побега

Нарушение транспорта белка STM вызывает дефекты развития ПАМ
Plant Journal, 2017
Нарушение транспорта белка STM вызывает дефекты развития ПАМ
Plant Journal, 2017

ТФ BELL (тоже из семейства TALE) – партнеры ТФ KNOX
(обеспечивают транспорт
ТФ BELL (тоже из семейства TALE) – партнеры ТФ KNOX
(обеспечивают транспорт
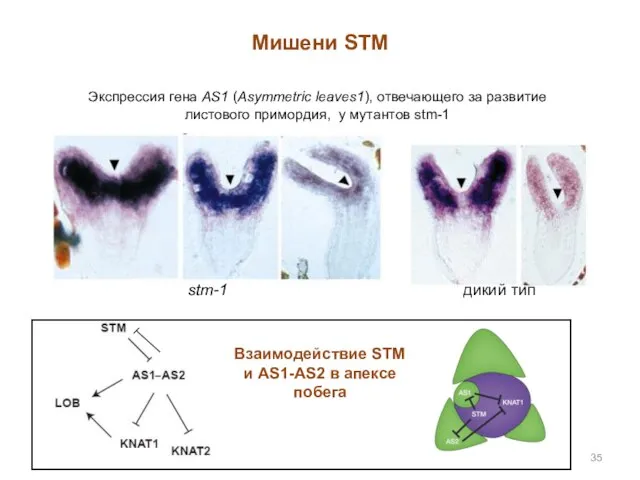
Мишени STM
Экспрессия гена AS1 (Asymmetric leaves1), отвечающего за развитие листового примордия,
Мишени STM
Экспрессия гена AS1 (Asymmetric leaves1), отвечающего за развитие листового примордия,
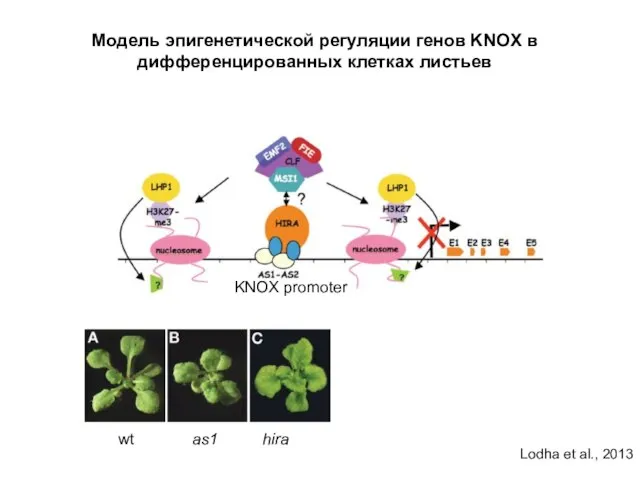
Модель эпигенетической регуляции генов KNOX в дифференцированных клетках листьев
wt as1
Модель эпигенетической регуляции генов KNOX в дифференцированных клетках листьев
wt as1
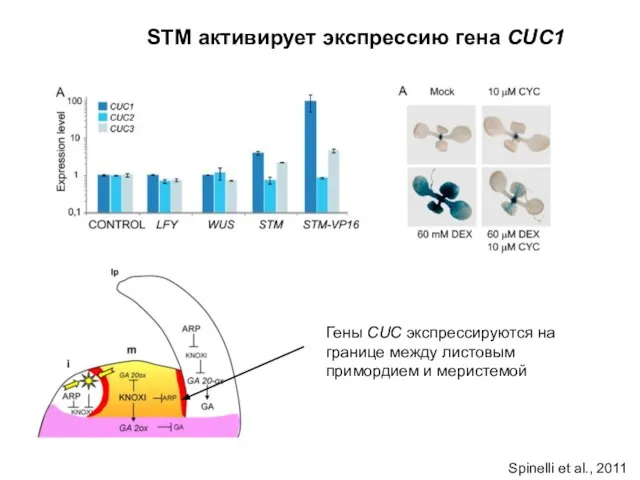
STM активирует экспрессию гена CUC1
Гены CUC экспрессируются на границе между листовым
STM активирует экспрессию гена CUC1
Гены CUC экспрессируются на границе между листовым

Мишени STM в ПАМ
STM
STM
IPT (биосинтез цитокинина)
GA2ox (дезактивация гибереллинов)
Jasinski et al. 2005
Мишени STM в ПАМ
STM
STM
IPT (биосинтез цитокинина)
GA2ox (дезактивация гибереллинов)
Jasinski et al. 2005

Взаимодействие STM (генов KNOX) и генов, вовлеченных в метаболизм цитокинина и
Взаимодействие STM (генов KNOX) и генов, вовлеченных в метаболизм цитокинина и
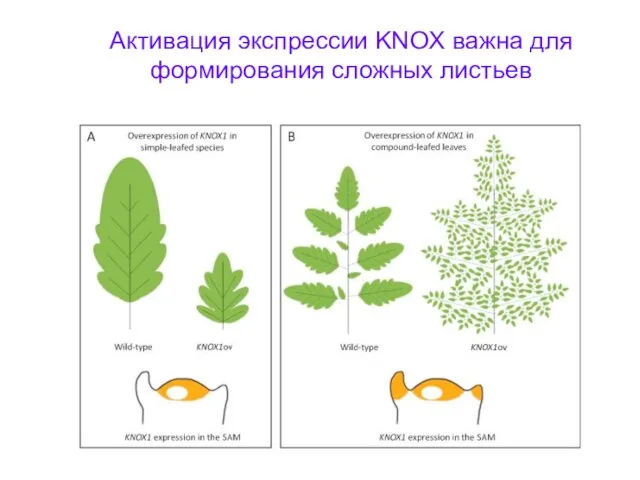
Активация экспрессии KNOX важна для формирования сложных листьев
Активация экспрессии KNOX важна для формирования сложных листьев
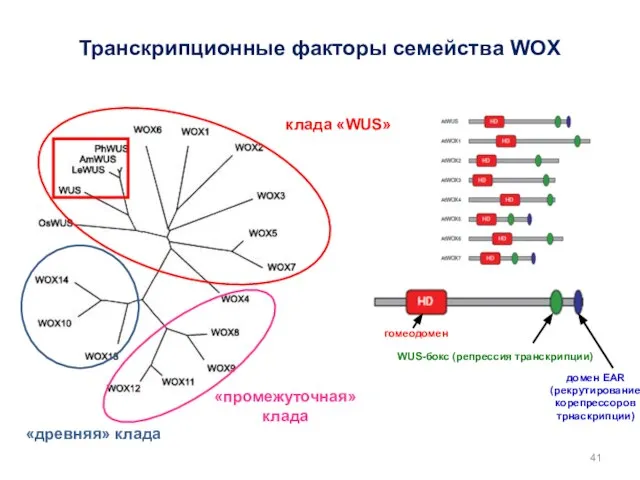
Транскрипционные факторы семейства WOX
WUS-бокс (репрессия транскрипции)
домен EAR
(рекрутирование корепрессоров трнаскрипции)
гомеодомен
«древняя» клада
«промежуточная»
Транскрипционные факторы семейства WOX
WUS-бокс (репрессия транскрипции)
домен EAR
(рекрутирование корепрессоров трнаскрипции)
гомеодомен
«древняя» клада
«промежуточная»

Ядерная локализация гибридного белка WUS-GUS
(в клетках эпидермиса лука)
Экспрессия гена WUS
Ядерная локализация гибридного белка WUS-GUS
(в клетках эпидермиса лука)
Экспрессия гена WUS

мхи
зеленые водоросли
WUS/WOX9
про-ортолог
линия WUS
линия WOX9
линия WOX13
WOX13
Плауны
Семенные растения
Папоротники
Покрытосеменные растения
Голосеменные растения
WUS/WOX5
про-ортолог
WUS WOX5
Усложнение организации меристемы
мхи
зеленые водоросли
WUS/WOX9
про-ортолог
линия WUS
линия WOX9
линия WOX13
WOX13
Плауны
Семенные растения
Папоротники
Покрытосеменные растения
Голосеменные растения
WUS/WOX5
про-ортолог
WUS WOX5
Усложнение организации меристемы

Геном Gnetum gnemon – содержит про-ортолог GgWUS/WOX5 (до дупликации генов WUS/WOX5
Геном Gnetum gnemon – содержит про-ортолог GgWUS/WOX5 (до дупликации генов WUS/WOX5

Взаимодействие WUS и системы генов CLAVATA
увеличение числа органов цветка
фасциация стебля
увеличение числа
Взаимодействие WUS и системы генов CLAVATA
увеличение числа органов цветка
фасциация стебля
увеличение числа

CLV1 и CLV2 кодируют рецепторные белки (c лейцин-богатым LRR–доменом), работающие в
CLV1 и CLV2 кодируют рецепторные белки (c лейцин-богатым LRR–доменом), работающие в
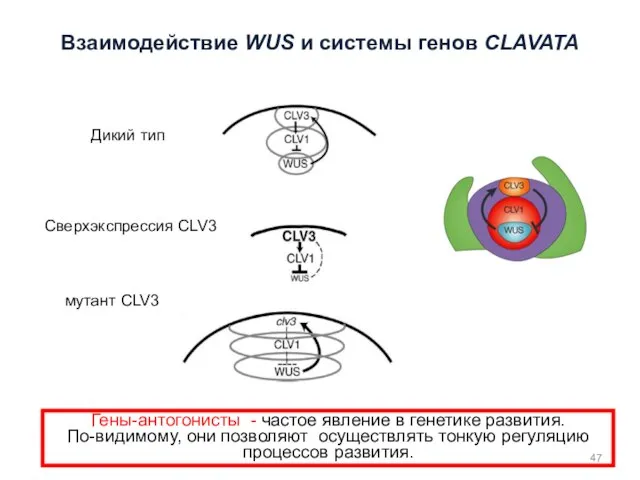
Гены-антогонисты - частое явление в генетике развития.
По-видимому, они позволяют осуществлять тонкую
Гены-антогонисты - частое явление в генетике развития.
По-видимому, они позволяют осуществлять тонкую

дикий тип мутант clv3 35S::CLV3
CLV3
WUS
CLV3
WUS
Увеличение домена экспрессии
дикий тип мутант clv3 35S::CLV3
CLV3
WUS
CLV3
WUS
Увеличение домена экспрессии
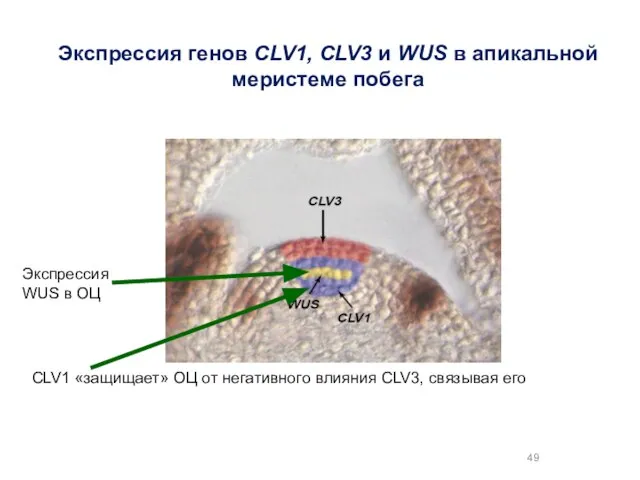
Экспрессия генов CLV1, CLV3 и WUS в апикальной меристеме побега
Экспрессия
Экспрессия генов CLV1, CLV3 и WUS в апикальной меристеме побега
Экспрессия

Рецепторные киназы растений c LRR–доменом
(Leucine-rich repeat receptor kinases)
Haffani et al., Can.
Рецепторные киназы растений c LRR–доменом
(Leucine-rich repeat receptor kinases)
Haffani et al., Can.
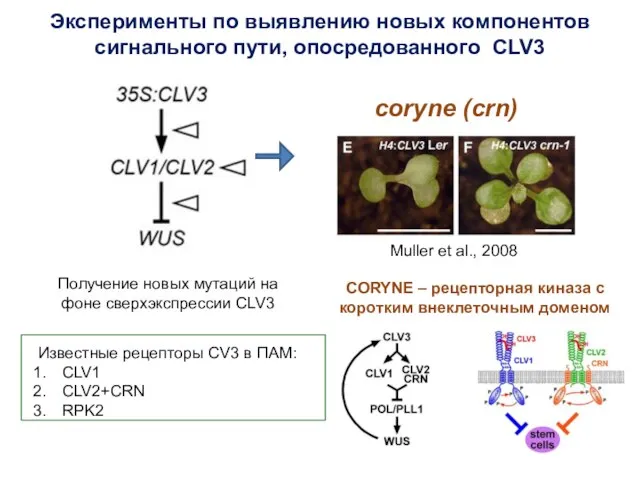
Эксперименты по выявлению новых компонентов
сигнального пути, опосредованного CLV3
Получение новых мутаций
Эксперименты по выявлению новых компонентов
сигнального пути, опосредованного CLV3
Получение новых мутаций
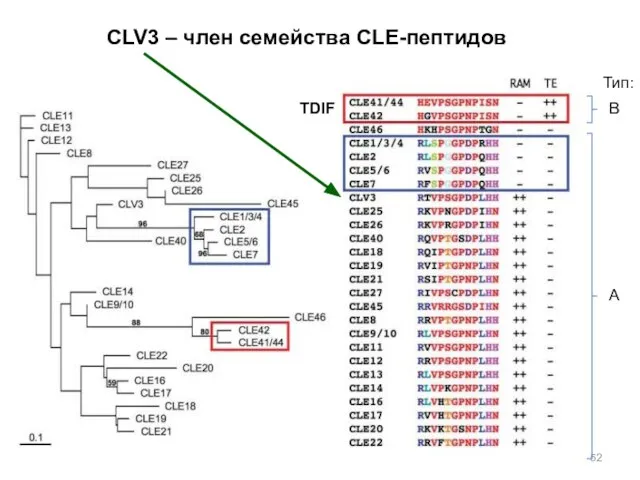
TDIF
CLV3 – член семейства CLE-пептидов
В
А
Тип:
TDIF
CLV3 – член семейства CLE-пептидов
В
А
Тип:

Участие CLE-пептидов в развитии Arabidopsis
Jun J et al. Plant Physiol. 2010;154:1721-1736
©2010
Участие CLE-пептидов в развитии Arabidopsis
Jun J et al. Plant Physiol. 2010;154:1721-1736
©2010
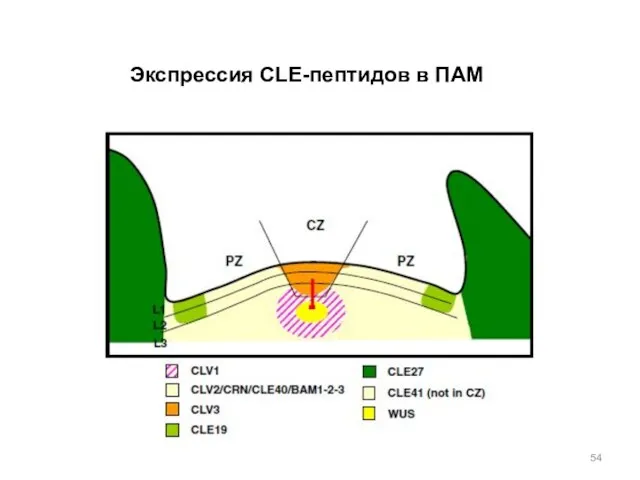
Экспрессия CLE-пептидов в ПАМ
Экспрессия CLE-пептидов в ПАМ

Стволовые клетки
WUS
?
Каким образом WUS регулирует поддержание запаса стволовых клеток?
Стволовые клетки
WUS
?
Каким образом WUS регулирует поддержание запаса стволовых клеток?
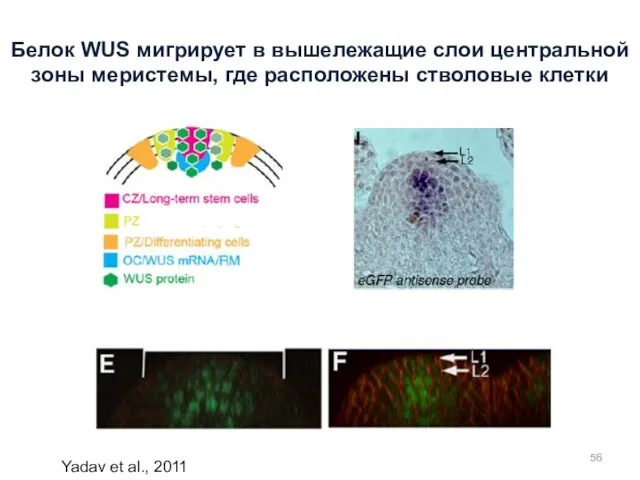
Yadav et al., 2011
Белок WUS мигрирует в вышележащие слои центральной зоны
Yadav et al., 2011
Белок WUS мигрирует в вышележащие слои центральной зоны
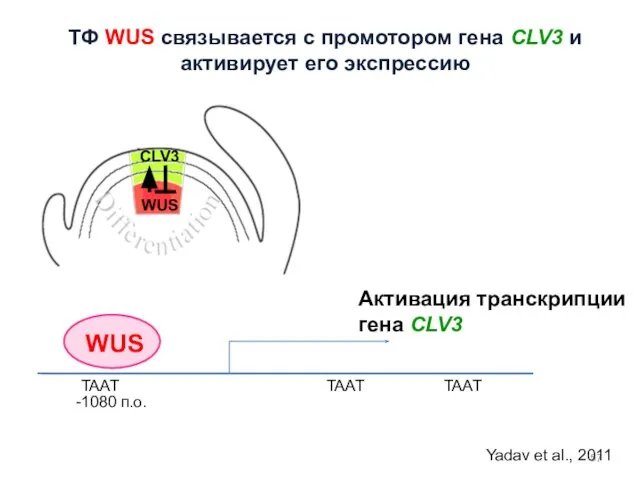
WUS
TAAT
TAAT
TAAT
-1080 п.о.
Активация транскрипции гена CLV3
Yadav et al., 2011
ТФ WUS связывается с
WUS
TAAT
TAAT
TAAT
-1080 п.о.
Активация транскрипции гена CLV3
Yadav et al., 2011
ТФ WUS связывается с
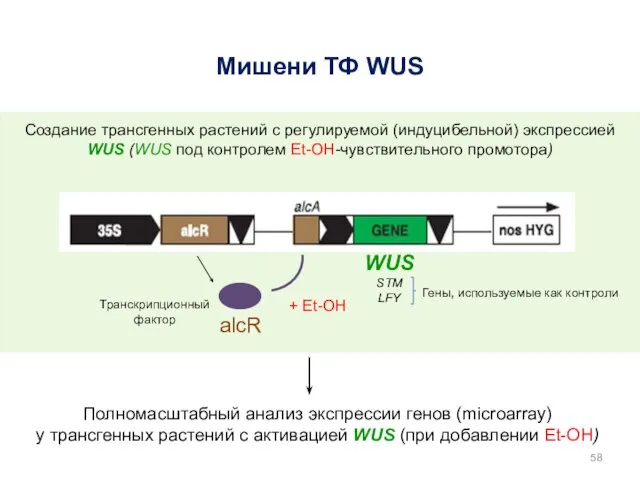
WUS STM
LFY
Мишени ТФ WUS
Создание трансгенных растений с регулируемой (индуцибельной) экспрессией WUS
WUS STM
LFY
Мишени ТФ WUS
Создание трансгенных растений с регулируемой (индуцибельной) экспрессией WUS

Yadav et al., 2013
WUS негативно регулирует экспрессию генов многих ТФ, регулирующих
Yadav et al., 2013
WUS негативно регулирует экспрессию генов многих ТФ, регулирующих
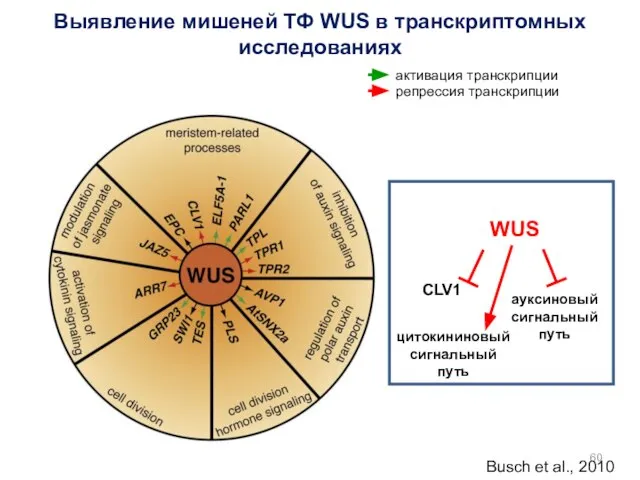
Busch et al., 2010
Выявление мишеней ТФ WUS в транскриптомных исследованиях
WUS
CLV1
ауксиновый сигнальный
Busch et al., 2010
Выявление мишеней ТФ WUS в транскриптомных исследованиях
WUS
CLV1
ауксиновый сигнальный

WUS негативно регулирует экспрессию генов
ARR А-типа (гены первичного ответа на
WUS негативно регулирует экспрессию генов
ARR А-типа (гены первичного ответа на

WUS негативно регулирует экспрессию генов LOG, кодирующих цитокинин-активирующие ферменты
Активность пролиферации
WUS негативно регулирует экспрессию генов LOG, кодирующих цитокинин-активирующие ферменты
Активность пролиферации
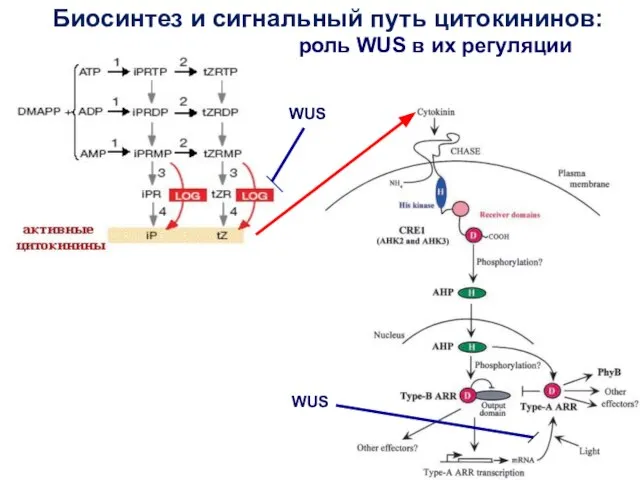
Биосинтез и сигнальный путь цитокининов:
WUS
WUS
роль WUS в их регуляции
Биосинтез и сигнальный путь цитокининов:
WUS
WUS
роль WUS в их регуляции
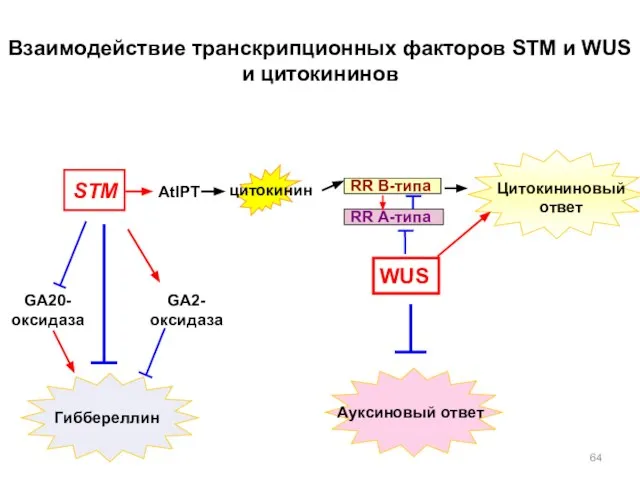
WUS
STM
AtIPT
Цитокининовый
ответ
цитокинин
Взаимодействие транскрипционных факторов STM и WUS и цитокининов
Ауксиновый
WUS
STM
AtIPT
Цитокининовый
ответ
цитокинин
Взаимодействие транскрипционных факторов STM и WUS и цитокининов
Ауксиновый

Роль гормонов в ПАМ
Роль гормонов в ПАМ

DR5:GFP
Ауксины как регуляторы формирования листовых примордиев
DR5:GFP
Ауксины как регуляторы формирования листовых примордиев
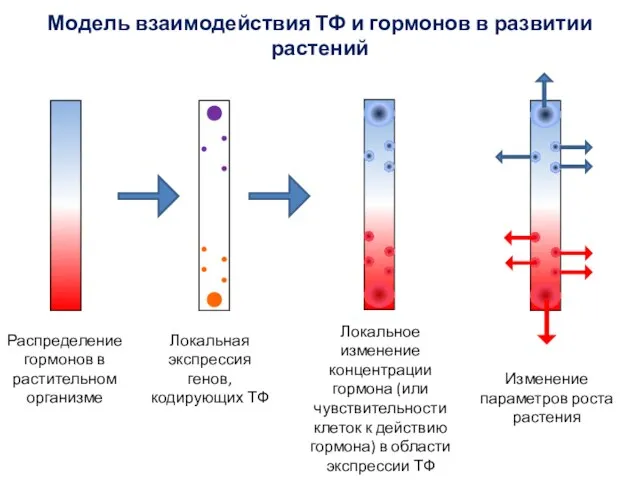
Локальная экспрессия генов, кодирующих ТФ
Локальное изменение концентрации гормона (или чувствительности клеток
Локальная экспрессия генов, кодирующих ТФ
Локальное изменение концентрации гормона (или чувствительности клеток
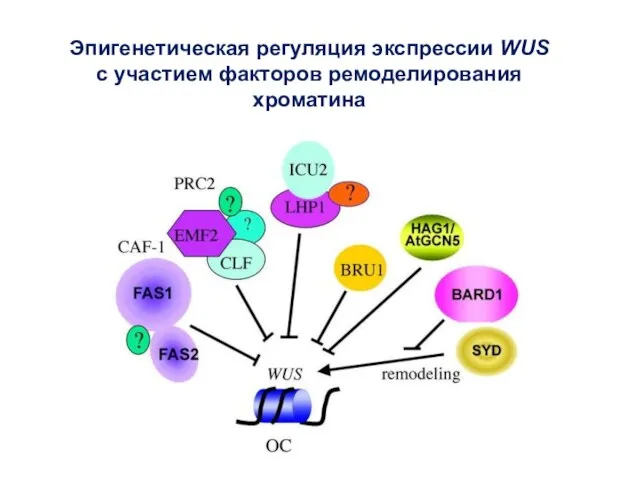
Эпигенетическая регуляция экспрессии WUS с участием факторов ремоделирования хроматина
Эпигенетическая регуляция экспрессии WUS с участием факторов ремоделирования хроматина
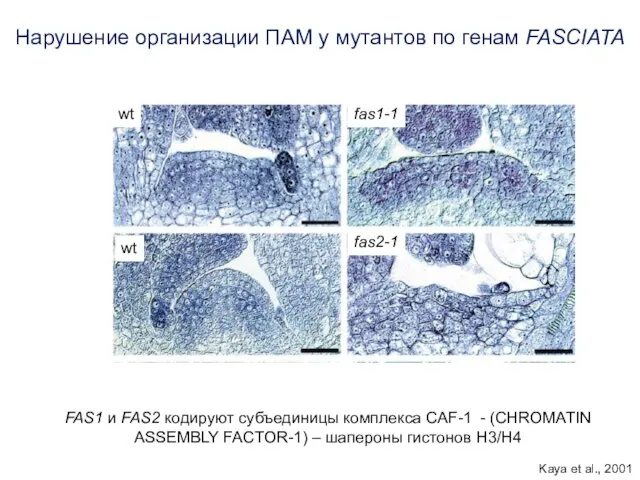
fas1-1
fas2-1
wt
wt
Нарушение организации ПАМ у мутантов по генам FASCIATA
FAS1 и FAS2
fas1-1
fas2-1
wt
wt
Нарушение организации ПАМ у мутантов по генам FASCIATA
FAS1 и FAS2

WUS
CLV3
before just after 2 days after
При выжигании зон экспрессии WUS
WUS
CLV3
before just after 2 days after
При выжигании зон экспрессии WUS
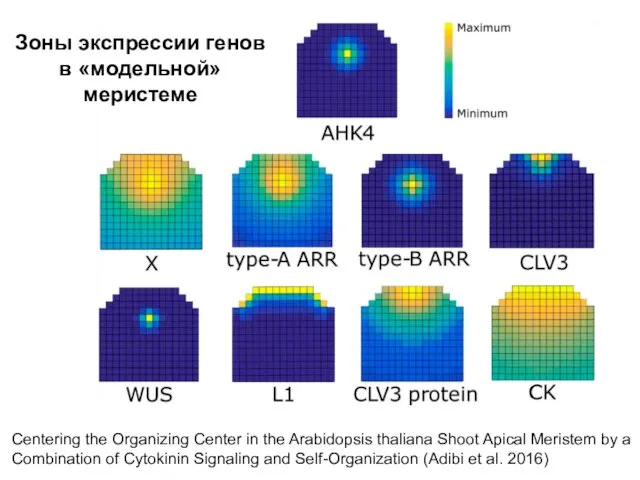
Зоны экспрессии генов в «модельной» меристеме
Centering the Organizing Center in the
Зоны экспрессии генов в «модельной» меристеме
Centering the Organizing Center in the

Регуляции функции меристемы с участием сигналов от дифференцированных клеток
дикий тип
ham
(hairy
Регуляции функции меристемы с участием сигналов от дифференцированных клеток
дикий тип
ham
(hairy
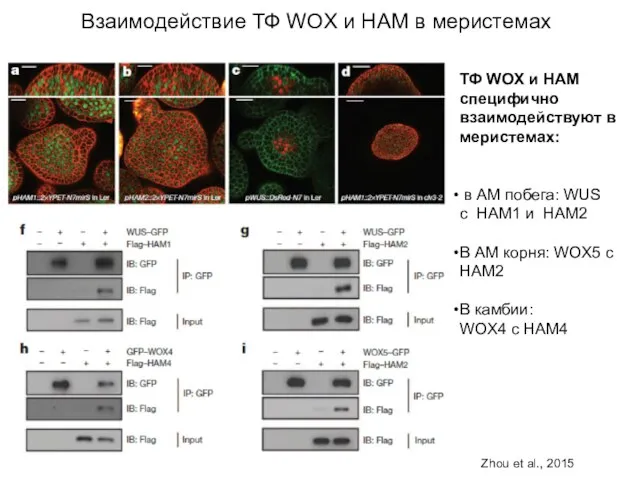
Взаимодействие ТФ WOX и HAM в меристемах
ТФ WOX и НАМ специфично
Взаимодействие ТФ WOX и HAM в меристемах
ТФ WOX и НАМ специфично

Филлотаксис
Золотое сечение (1,61803…)
Золотой угол (137,5°…) – «золотое сечение» окружности
Филлотаксис
Золотое сечение (1,61803…)
Золотой угол (137,5°…) – «золотое сечение» окружности
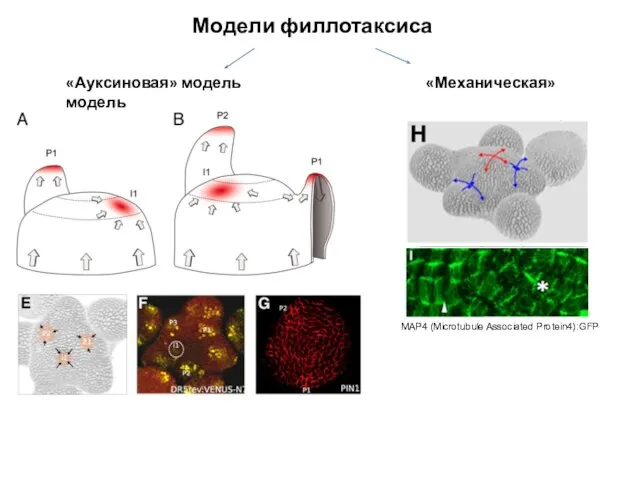
Модели филлотаксиса
«Ауксиновая» модель «Механическая» модель
MAP4 (Microtubule Associated Protein4):GFP
Модели филлотаксиса
«Ауксиновая» модель «Механическая» модель
MAP4 (Microtubule Associated Protein4):GFP
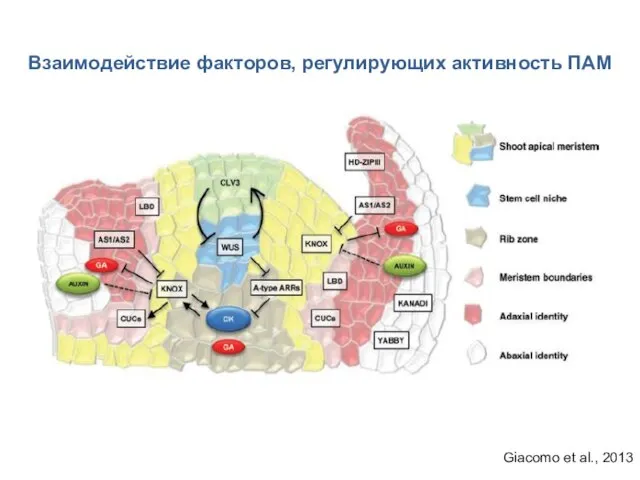
Giacomo et al., 2013
Взаимодействие факторов, регулирующих активность ПАМ
Giacomo et al., 2013
Взаимодействие факторов, регулирующих активность ПАМ
 Доказательство научности факта. Этапы научного эксперимента на примере изучения условий прорастания семян иссл
Доказательство научности факта. Этапы научного эксперимента на примере изучения условий прорастания семян иссл Питание человека
Питание человека Презентация на тему "Здоровое питание или о пользе молочных продуктов" - скачать презентации по Биологии
Презентация на тему "Здоровое питание или о пользе молочных продуктов" - скачать презентации по Биологии Животные эдиакария
Животные эдиакария Презентация на тему "Смешанные леса " - скачать презентации по Биологии
Презентация на тему "Смешанные леса " - скачать презентации по Биологии Презентация на тему "СТРУКТУРНЫЕ ОСНОВЫ БИОЛОГИИ РАЗВИТИЯ" - скачать презентации по Биологии
Презентация на тему "СТРУКТУРНЫЕ ОСНОВЫ БИОЛОГИИ РАЗВИТИЯ" - скачать презентации по Биологии Презентацию подготовила Семибратова Л.П. Учитель биологии МОУ «Чебаклинская СОШ»
Презентацию подготовила Семибратова Л.П. Учитель биологии МОУ «Чебаклинская СОШ» Физиология высшей нервной деятельности
Физиология высшей нервной деятельности Минеральное питание растений. (6 класс)
Минеральное питание растений. (6 класс) Презентация на тему Группы растений
Презентация на тему Группы растений  Рослини-хижаки
Рослини-хижаки История развития жизни на Земле
История развития жизни на Земле Генетика как наука. Закономерности наследственности установленные Г. Менделем
Генетика как наука. Закономерности наследственности установленные Г. Менделем Функциональная анатомия ствола головного мозга и конечного мозга
Функциональная анатомия ствола головного мозга и конечного мозга Презентация на тему Покровы тела. Сравнительная характеристика.
Презентация на тему Покровы тела. Сравнительная характеристика. La fase líquida y gaseosa del suelo la humedad del suelo
La fase líquida y gaseosa del suelo la humedad del suelo Презентация на тему "Помогите мне увидеть мир" - скачать презентации по Биологии
Презентация на тему "Помогите мне увидеть мир" - скачать презентации по Биологии Доказательства эволюции животных
Доказательства эволюции животных Презентация Использование фитонцидов растений для оздоровления воздуха помещений
Презентация Использование фитонцидов растений для оздоровления воздуха помещений  Посев зерновых и зернобобовых культур
Посев зерновых и зернобобовых культур Пищеварительная система
Пищеварительная система Schnuffi, der kleine Igel
Schnuffi, der kleine Igel Презентация на тему "Значение растений в природе и в жизни человека" - скачать презентации по Биологии
Презентация на тему "Значение растений в природе и в жизни человека" - скачать презентации по Биологии Симбиоз
Симбиоз Диагностика дикроцелиоза и парамфистоматоза жвачных. Метод последовательных смывов
Диагностика дикроцелиоза и парамфистоматоза жвачных. Метод последовательных смывов Отряд Двукрылые Составила: учитель биологии и химии высшей квалификационной категории Бочкова Ирина Анатольевна
Отряд Двукрылые Составила: учитель биологии и химии высшей квалификационной категории Бочкова Ирина Анатольевна  Биоритмы как основа рациональной организации порядка человека
Биоритмы как основа рациональной организации порядка человека Пластический и энергетический обмен
Пластический и энергетический обмен