- Лекции3 Генная Инженерия 2 курс Био Маги

Содержание
- 2. 25-30 мышиных яйцеклеток с чужеродным геном после микроинъекции имплантируются в псевдобеоременную самку, которая была предварительно спарена
- 3. Использование вирусов для переноса генов Вирусные частицы имеют природное свойство адсорбироваться на поверхности клеток и внедрять
- 4. Преимущества вирусных векторов Большая емкость для чужеродной ДНК У вируса нет цитотоксического эффекта, так как некоторые
- 5. Адено-ассоциированные вирусы (AAV) Эти вирусы генетически не ролдственны аденовирусам, но называются так, потому что впервые были
- 6. Зависимость AAV от гетерологичного вируса-помощника, например, аденовируса, обеспечивает высокий уровень контроля над вирусной репликацией, делая AAV
- 7. Геном AAV является мальньким (5 тыс.п.о.) и включает центральный район, содержащий ген rep (репликаза) и cap
- 8. Созданы новые векторы с делецией генов rep и cap genes и использующие только повторяющиеся элементы, необходимые
- 9. Бакуловирусные векторы (BV) Бакуловирусы обеспечивают высокий уровень экспрессии трансгена в клетках насекомых, однако их можно использовать
- 10. Одна из проблем экспрессии белков млекопитающих в клетках насекомых – различный тип гликозилирования белков. Одним из
- 11. Протокол для получения трансгенных телят Сбор ооцитов с мясокомбината (забой коров); in vitro созревание этих ооцитов;
- 12. Цели получения трансгенных коров Изменить состав молока. Например, получать молока с желательными аллелями по гену каппа-казеина.
- 13. Трансгенная птица Птичьи яйцеклетки обычно оплодотворяются через 30 минут после овуляции. Деление клеток происходит в яйцеводе
- 14. Протокол для птицы Бластодермальные клетки удаляют из донорного яйца. Эти клетки трансфицируют катионными липидами (липосомами) с
- 15. Цели получения трансгенной птицы Улучшить генетику существующих линий птицы в отношении: Устойчивость к птичьим вирусным и
- 16. Методы получения трансгенных растений
- 17. Агробактерии - природные геномодификаторы По мнению древних ученых-философов, ни один человек не способен придумать что-либо, чего
- 18. АГРОБАКТЕРИИ Род грамотрицательных аэробных бактерий. 4 вида, обитают в почве, главным образом в ризосфере. Способны вызывать
- 19. Клетки растений можно обработать лизоцимом для удаления клеточной стенки с получением протопластов, которые осторожно центрифугируют, наслаивая
- 20. Получив такой подарок, клетки начинают бурно делиться, превращаясь в разрастание рыхлой ткани — корончатый галл, и
- 21. Генетическая колонизация растения 1- агробактерии существуют в ризосфере; 2 - строение A. tumefaciens; 3 – встраивание
- 22. Локализация корончатого галла на шейке корней растений. Опины (необычное для растений соединение) синтезируются трансформированными растениями. Не
- 23. Корневой рак плодовых культур.
- 26. Скачать презентацию

25-30 мышиных яйцеклеток с чужеродным геном после микроинъекции имплантируются в псевдобеоременную
25-30 мышиных яйцеклеток с чужеродным геном после микроинъекции имплантируются в псевдобеоременную

Использование вирусов для переноса генов
Вирусные частицы имеют природное свойство адсорбироваться на
Использование вирусов для переноса генов
Вирусные частицы имеют природное свойство адсорбироваться на

Преимущества вирусных векторов
Большая емкость для чужеродной ДНК
У вируса нет цитотоксического эффекта,
Преимущества вирусных векторов
Большая емкость для чужеродной ДНК
У вируса нет цитотоксического эффекта,

Адено-ассоциированные вирусы (AAV)
Эти вирусы генетически не ролдственны аденовирусам, но называются так,
Адено-ассоциированные вирусы (AAV)
Эти вирусы генетически не ролдственны аденовирусам, но называются так,

Зависимость AAV от гетерологичного вируса-помощника, например, аденовируса, обеспечивает высокий уровень контроля
Зависимость AAV от гетерологичного вируса-помощника, например, аденовируса, обеспечивает высокий уровень контроля

Геном AAV является мальньким (5 тыс.п.о.) и включает центральный район, содержащий
Геном AAV является мальньким (5 тыс.п.о.) и включает центральный район, содержащий

Созданы новые векторы с делецией генов rep и cap genes и
Созданы новые векторы с делецией генов rep и cap genes и

Бакуловирусные векторы (BV)
Бакуловирусы обеспечивают высокий уровень экспрессии трансгена в клетках насекомых,
Бакуловирусные векторы (BV)
Бакуловирусы обеспечивают высокий уровень экспрессии трансгена в клетках насекомых,

Одна из проблем экспрессии белков млекопитающих в клетках насекомых – различный
Одна из проблем экспрессии белков млекопитающих в клетках насекомых – различный

Протокол для получения трансгенных телят
Сбор ооцитов с мясокомбината (забой коров);
in vitro
Протокол для получения трансгенных телят
Сбор ооцитов с мясокомбината (забой коров);
in vitro

Цели получения трансгенных коров
Изменить состав молока. Например, получать молока с желательными
Цели получения трансгенных коров
Изменить состав молока. Например, получать молока с желательными

Трансгенная птица
Птичьи яйцеклетки обычно оплодотворяются через 30 минут после овуляции. Деление
Трансгенная птица
Птичьи яйцеклетки обычно оплодотворяются через 30 минут после овуляции. Деление

Протокол для птицы
Бластодермальные клетки удаляют из донорного яйца.
Эти клетки
Протокол для птицы
Бластодермальные клетки удаляют из донорного яйца.
Эти клетки

Цели получения трансгенной птицы
Улучшить генетику существующих линий птицы в
Цели получения трансгенной птицы
Улучшить генетику существующих линий птицы в

Методы получения трансгенных растений
Методы получения трансгенных растений

Агробактерии - природные геномодификаторы
По мнению древних ученых-философов, ни один человек не
Агробактерии - природные геномодификаторы
По мнению древних ученых-философов, ни один человек не

АГРОБАКТЕРИИ
Род грамотрицательных аэробных бактерий. 4 вида, обитают в почве, главным
АГРОБАКТЕРИИ
Род грамотрицательных аэробных бактерий. 4 вида, обитают в почве, главным

Клетки растений можно обработать лизоцимом для удаления клеточной стенки с получением
Клетки растений можно обработать лизоцимом для удаления клеточной стенки с получением
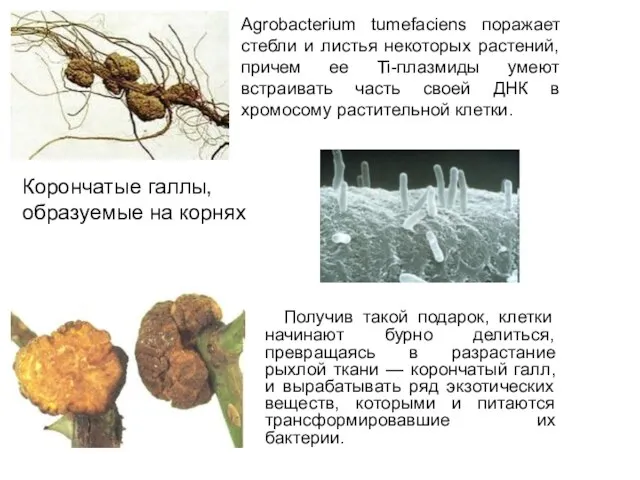
Получив такой подарок, клетки начинают бурно делиться, превращаясь в разрастание
Получив такой подарок, клетки начинают бурно делиться, превращаясь в разрастание
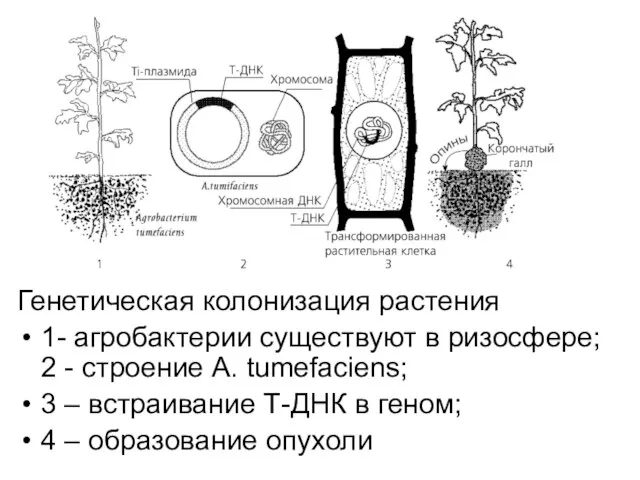
Генетическая колонизация растения
1- агробактерии существуют в ризосфере; 2 - строение
Генетическая колонизация растения
1- агробактерии существуют в ризосфере; 2 - строение

Локализация корончатого галла на шейке корней растений.
Опины (необычное для растений соединение)
Локализация корончатого галла на шейке корней растений.
Опины (необычное для растений соединение)
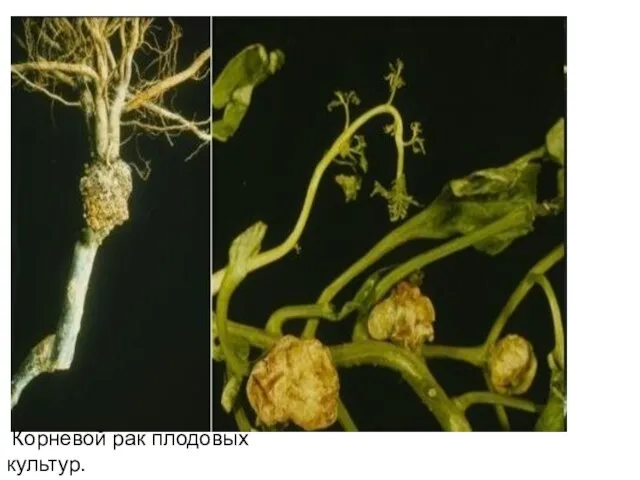
Корневой рак плодовых культур.
Корневой рак плодовых культур.

 ЭКОЛОГИЧЕСКИЙ ЭРУДИЦИОН
ЭКОЛОГИЧЕСКИЙ ЭРУДИЦИОН Презентация на тему "Рост и развитие животных" - скачать презентации по Биологии
Презентация на тему "Рост и развитие животных" - скачать презентации по Биологии Печень, строение и функции.
Печень, строение и функции. Презентация Фотосинтез
Презентация Фотосинтез Выращивание кактуса в домашних условиях
Выращивание кактуса в домашних условиях Исследование: Жизнь сирийского хомячка в домашних условиях
Исследование: Жизнь сирийского хомячка в домашних условиях Биологическая роль элементов IIА группы и их применение в медицине
Биологическая роль элементов IIА группы и их применение в медицине Презентация на тему "Анатомо-физиологические основы полового воспитания" - скачать презентации по Биологии
Презентация на тему "Анатомо-физиологические основы полового воспитания" - скачать презентации по Биологии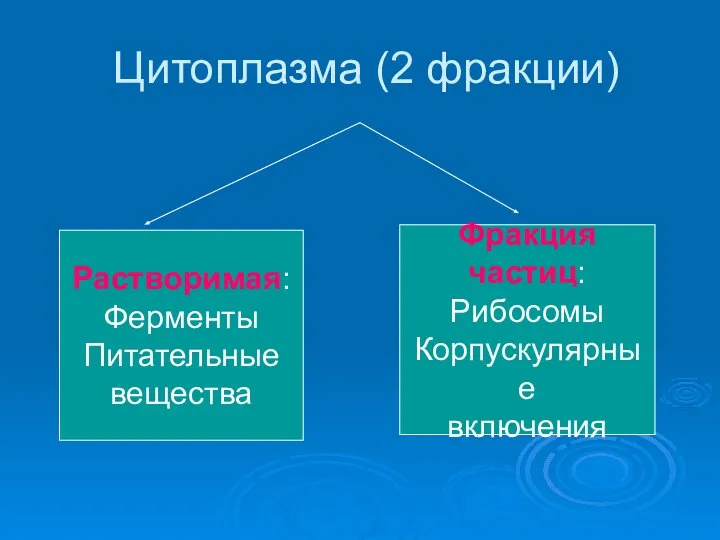 Цитоплазма (2 фракции)
Цитоплазма (2 фракции) Презентация на тему "Тип Моллюски" - скачать бесплатно презентации по Биологии
Презентация на тему "Тип Моллюски" - скачать бесплатно презентации по Биологии Отряд Зайцеобразные
Отряд Зайцеобразные Экскурсия. Океанариум
Экскурсия. Океанариум Презентация на тему Свертывание крови. Гемофилия.
Презентация на тему Свертывание крови. Гемофилия. Физиология целенаправленного поведения
Физиология целенаправленного поведения Muscular tissue
Muscular tissue Отдел Моховидные
Отдел Моховидные Микрофлора окружающей среды. Санитарно-микробиологический контроль воды
Микрофлора окружающей среды. Санитарно-микробиологический контроль воды Горилла Коко
Горилла Коко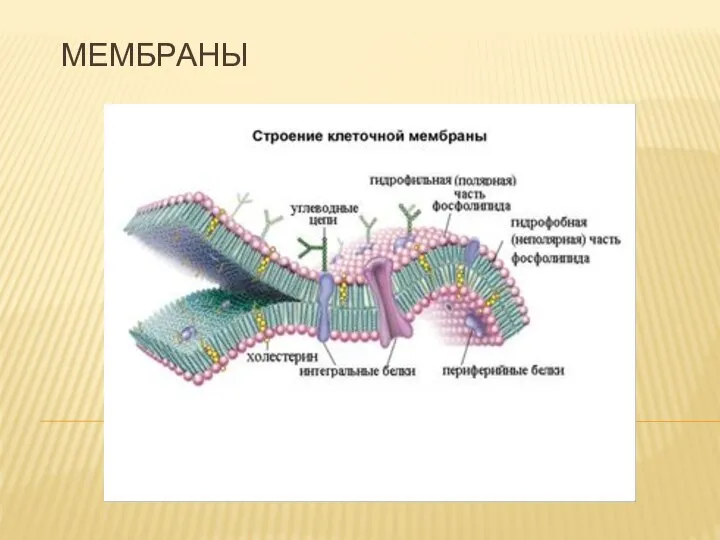 Мембраны. Свойства мембраны
Мембраны. Свойства мембраны Умная домашняя теплица “EcoGrow”
Умная домашняя теплица “EcoGrow” Презентация на тему "С уважением к энергосбережению" - скачать бесплатно презентации по Биологии
Презентация на тему "С уважением к энергосбережению" - скачать бесплатно презентации по Биологии Презентация на тему "Необычные породы собак" - скачать бесплатно презентации по Биологии
Презентация на тему "Необычные породы собак" - скачать бесплатно презентации по Биологии Презентация на тему "Цикл Кребса" - скачать презентации по Биологии
Презентация на тему "Цикл Кребса" - скачать презентации по Биологии Белки , жиры и углеводы
Белки , жиры и углеводы Обмен нуклеотидов. (Лекция 13)
Обмен нуклеотидов. (Лекция 13) Презентация на тему "Строение растения кукурузы" - скачать бесплатно презентации по Биологии
Презентация на тему "Строение растения кукурузы" - скачать бесплатно презентации по Биологии Ботаника. Задание 4
Ботаника. Задание 4 Типы наследования признаков у человека
Типы наследования признаков у человека