- Потенциал покоя. Потенциал действия. Проведение нервного импульса

Содержание
- 2. Механизм распространения нервных импульсов был выяснен лишь в последние полвека, после того как у кальмара обнаружили
- 3. Когда кончик микроэлектрода проходит сквозь плазматическую мембрану аксона, лучи осциллографа раздвигаются. Расстояние между лучами на экране
- 4. Мышечные клетки Секреторные клетки Рецепторные клетки Нейроны Потенциал покоя (ПП)
- 5. Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его
- 6. В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40 раз больше, чем вне клетки;
- 7. В клетке находятся также крупномолекулярные анионы; главным образом это отрицательно заряженные белковые молекулы, например глутамат, аспартат,
- 8. Проницаемость клеточной мембраны – это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам
- 9. Проводимость иона зависит от электрохимического градиента и от проницаемости мембраны: чем они больше, тем лучше проводимость
- 10. Потенциал покоя (ПП)
- 11. Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам диффузии, при этом К+ из
- 12. Главным ионом, обеспечивающим формирование ПП, является ион К+. Об этом свидетельствуют результаты опыта с перфузией внутреннего
- 13. Модельный опыт. Сосуд разделен полупроницаемой мембраной. Обе его половины заполнены растворами K2SO4 различной концентрации (С1, и
- 14. В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно
- 15. Величина ПП изменяется в различных условиях деятельности клетки, вследствие этого колеблется и ее возбудимость, например при
- 16. Какие клетки относятся к возбудимым? Нервные, мышечные, рецепторные и секреторные клетки. Какова концентрация ионов натрия снаружи
- 17. Что характерно для калиевых каналов когда клетка находится в состоянии покоя? Калиевые каналы в состоянии покоя
- 18. Как калий-натриевый насос поддерживает потенциал покоя? Для работы этого насоса необходима энергия АТФ, а действует он
- 19. Потенциал действия – это процесс, выражающийся в быстром колебании мембранного потенциала покоя вследствие перемещения ионов в
- 20. Амплитуда ПД не зависит от силы раздражения – она всегда максимальна для данной клетки в конкретных
- 21. В составе ПД различают четыре фазы: 1 – деполяризацию, т.е. исчезновение заряда клетки – уменьшение мембранного
- 22. Процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение Na+ в клетку.
- 23. Деполяризация. При действии деполяризующего раздражителя на клетку (медиатор, электрический ток). Условием, обеспечивающим вход Na+ в клетку,
- 24. В каналах для Na+ имеются активационные m-ворота, которые расположены с внешней стороны клеточной мембраны, и инактивационные
- 25. Инверсия. Примерно через 0,5—2 мс и более после начала деполяризации (это время зависит от вида клетки)
- 26. Выходу К+ из клетки способствует также и электрический градиент. К+ выталкивается положительным зарядом из клетки и
- 27. Фаза реполяризации связана с тем, что проницаемость клеточной мембраны для К+ все еще высока, ворота калиевых
- 28. Фактически мембранный потенциал снижается до более отрицательного, чем в покое, значения. Происходит гиперполяризация, обусловленная тем, что
- 29. Абсолютная рефрактерная фаза – это полная невозбудимость клетки (возбудимость равна нулю), она соответствует пику ПД и
- 30. Фазы ПД: 1 – порог (около 50 мв, ток Na>K) 1-2 – деполяризация 0,5 мс (вход
- 31. 1 Ионные токи во время ПД 2 3 4 5 Потенциал действия (ПД)
- 33. Какому закону подчиняется потенциал действия? ПД подчиняется закону «все или ничего». ПД либо совсем не возникает
- 34. Что происходит с внешними (m-воротами) и внутренними (h-воротами) натриевых каналов во время покоя и при деполяризации?
- 35. Возбудимость повышена. Абсолютная рефрактерная фаза. Относительная рефрактерная фаза. Возбудимость повышена. Подведем итоги. Что обозначено цифрами 1-4:
- 36. В немиелинизированных аксонах, типичных для беспозвоночных, скорость распространения потенциалов действия зависит от сопротивления аксоплазмы. Это сопротивление
- 37. У позвоночных подавляющая часть нервных волокон, особенно в спинномозговых и черепных нервах, окружены миелиновой оболочкой, образованной
- 38. По мере повышения температуры примерно до 40 °С скорость проведения импульсов возрастает. Проведение нервного импульса
- 40. Основные законы проведения нервного импульса: При раздражении нервного волокна возбуждение по нерву распространяется в обе стороны.
- 41. Нервные окончания могут быть рецепторными (экстерорецепторы и интерорецепторы) и эффекторными, например химические синапсы. Строение синапса? Биохимическая
- 44. Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу
- 45. Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для
- 46. Инициатором сокращения миокарда, как и в скелетной мышце, является потенциал действия, распространяющийся вдоль поверхностной мембраны кардиомиоцита.
- 47. Считается, что потенциал действия распространяется с поверхностной мембраны кардиомиоцита вдоль Т-трубочки вглубь волокна и вызывает деполяризацию
- 48. Сократительная система сердца представлена сократительными белками – актином и миозином, а также тропомиозином и тропонином. На
- 49. Удаление кальция из межклеточного пространства приводит к разобщению процессов возбуждения и сокращения миокарда. Потенциалы действия при
- 50. Выход потенциала действия на плато в мышечной клетке сердца связан с: а) вхождением в клетку ионов
- 51. Почему миелинизированные аксоны лягушки 3,5 мкм проводят импульсы со скоростью 30 м/сек, тогда как аксоны кошки
- 52. Подведем итоги. Поясните рисунки:
- 54. Скачать презентацию

Механизм распространения нервных импульсов был выяснен лишь в последние полвека, после
Механизм распространения нервных импульсов был выяснен лишь в последние полвека, после
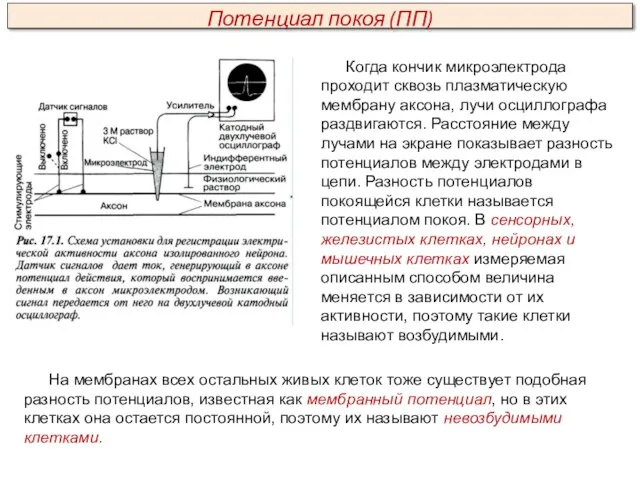
Когда кончик микроэлектрода проходит сквозь плазматическую мембрану аксона, лучи осциллографа раздвигаются.
Когда кончик микроэлектрода проходит сквозь плазматическую мембрану аксона, лучи осциллографа раздвигаются.
Мышечные клетки
Секреторные клетки
Рецепторные клетки
Нейроны
Потенциал покоя (ПП)
Мышечные клетки
Секреторные клетки
Рецепторные клетки
Нейроны
Потенциал покоя (ПП)

Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и
Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и
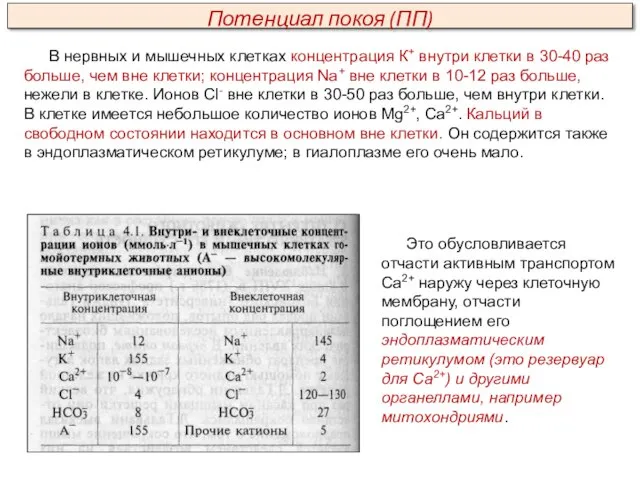
В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40
В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40

В клетке находятся также крупномолекулярные анионы; главным образом это отрицательно заряженные
В клетке находятся также крупномолекулярные анионы; главным образом это отрицательно заряженные

Проницаемость клеточной мембраны – это ее способность пропускать воду, незаряженные и
Проницаемость клеточной мембраны – это ее способность пропускать воду, незаряженные и

Проводимость иона зависит от электрохимического градиента и от проницаемости мембраны: чем
Проводимость иона зависит от электрохимического градиента и от проницаемости мембраны: чем
Потенциал покоя (ПП)
Потенциал покоя (ПП)
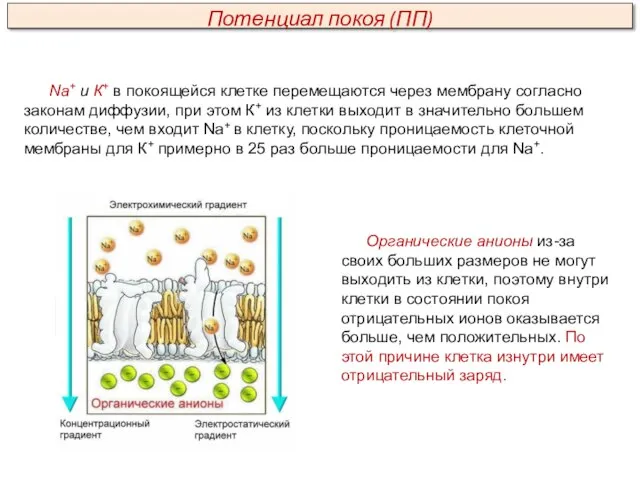
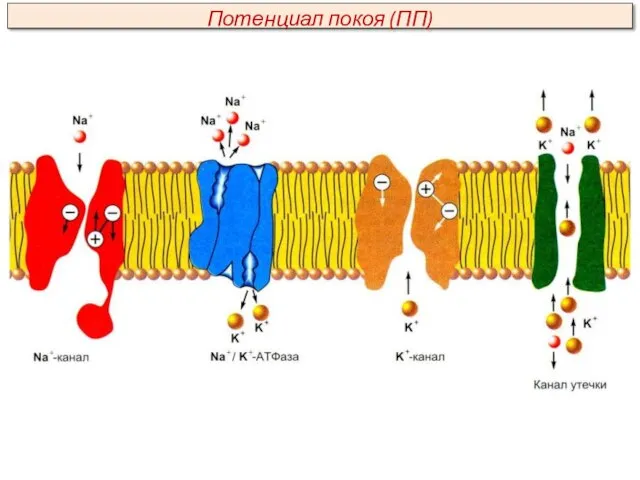
Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам
Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам

Главным ионом, обеспечивающим формирование ПП, является ион К+. Об этом свидетельствуют
Главным ионом, обеспечивающим формирование ПП, является ион К+. Об этом свидетельствуют

Модельный опыт. Сосуд разделен полупроницаемой мембраной. Обе его половины заполнены растворами
Модельный опыт. Сосуд разделен полупроницаемой мембраной. Обе его половины заполнены растворами

В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация
В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация

Величина ПП изменяется в различных условиях деятельности клетки, вследствие этого колеблется
Величина ПП изменяется в различных условиях деятельности клетки, вследствие этого колеблется

Какие клетки относятся к возбудимым?
Нервные, мышечные, рецепторные и секреторные клетки.
Какова концентрация
Какие клетки относятся к возбудимым?
Нервные, мышечные, рецепторные и секреторные клетки.
Какова концентрация

Что характерно для калиевых каналов когда клетка находится в состоянии покоя?
Калиевые
Что характерно для калиевых каналов когда клетка находится в состоянии покоя?
Калиевые

Как калий-натриевый насос поддерживает потенциал покоя?
Для работы этого насоса необходима энергия
Как калий-натриевый насос поддерживает потенциал покоя?
Для работы этого насоса необходима энергия
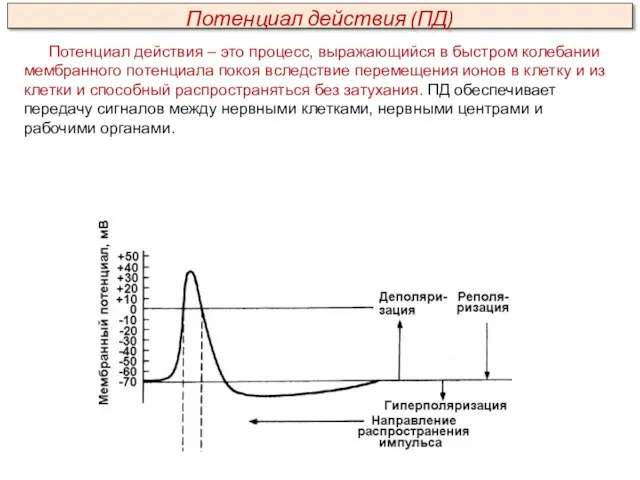
Потенциал действия – это процесс, выражающийся в быстром колебании мембранного потенциала
Потенциал действия – это процесс, выражающийся в быстром колебании мембранного потенциала

Амплитуда ПД не зависит от силы раздражения – она всегда максимальна
Амплитуда ПД не зависит от силы раздражения – она всегда максимальна
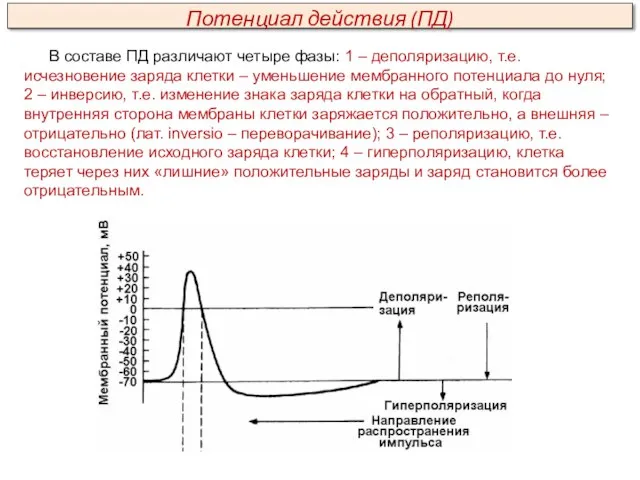
В составе ПД различают четыре фазы: 1 – деполяризацию, т.е. исчезновение
В составе ПД различают четыре фазы: 1 – деполяризацию, т.е. исчезновение
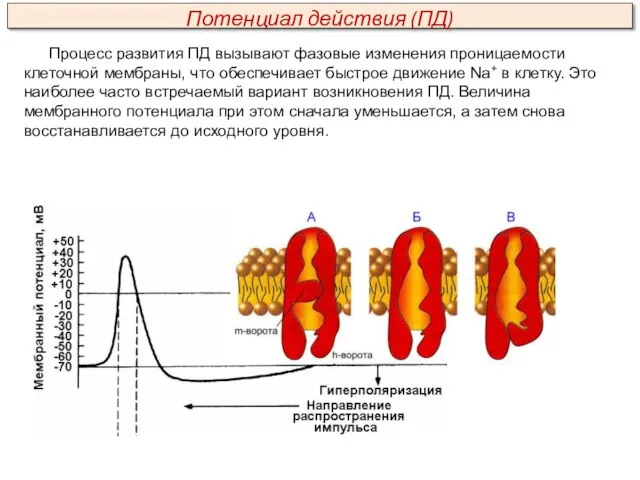
Процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает
Процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает

Деполяризация. При действии деполяризующего раздражителя на клетку (медиатор, электрический ток). Условием,
Деполяризация. При действии деполяризующего раздражителя на клетку (медиатор, электрический ток). Условием,
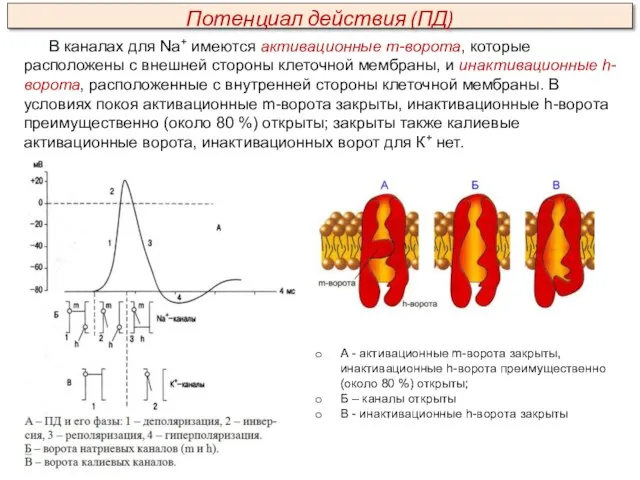
В каналах для Na+ имеются активационные m-ворота, которые расположены с внешней
В каналах для Na+ имеются активационные m-ворота, которые расположены с внешней
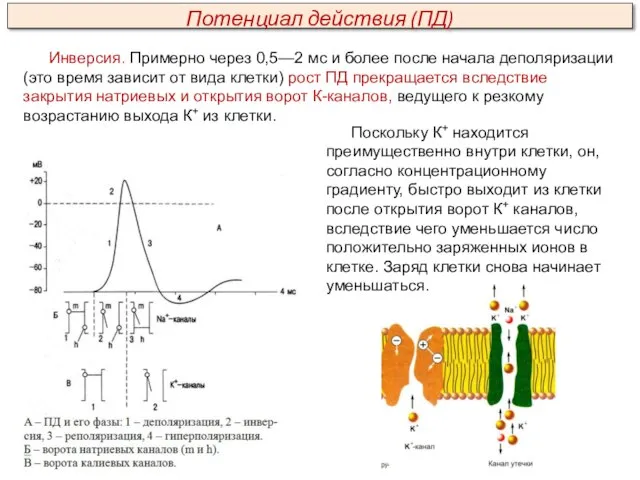
Инверсия. Примерно через 0,5—2 мс и более после начала деполяризации (это
Инверсия. Примерно через 0,5—2 мс и более после начала деполяризации (это

Выходу К+ из клетки способствует также и электрический градиент. К+ выталкивается
Выходу К+ из клетки способствует также и электрический градиент. К+ выталкивается
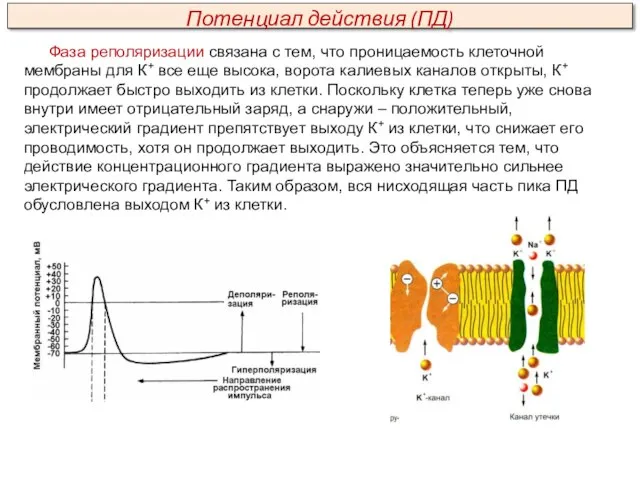
Фаза реполяризации связана с тем, что проницаемость клеточной мембраны для К+
Фаза реполяризации связана с тем, что проницаемость клеточной мембраны для К+

Фактически мембранный потенциал снижается до более отрицательного, чем в покое, значения.
Фактически мембранный потенциал снижается до более отрицательного, чем в покое, значения.

Абсолютная рефрактерная фаза – это полная невозбудимость клетки (возбудимость равна нулю),
Абсолютная рефрактерная фаза – это полная невозбудимость клетки (возбудимость равна нулю),

Фазы ПД:
1 – порог (около 50 мв, ток Na>K)
1-2 – деполяризация
Фазы ПД:
1 – порог (около 50 мв, ток Na>K)
1-2 – деполяризация
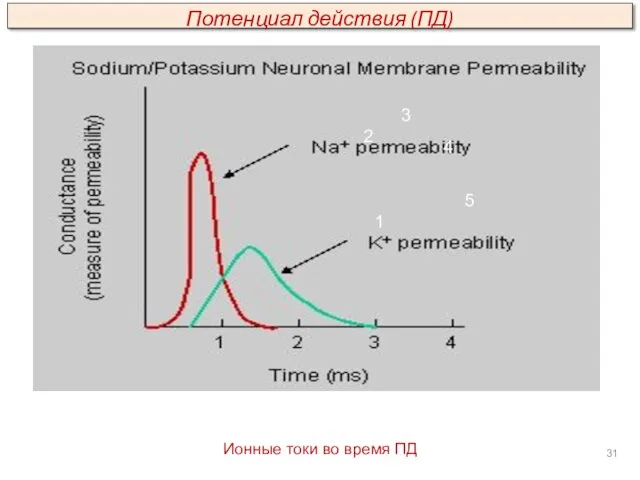
1
Ионные токи во время ПД
2
3
4
5
Потенциал действия (ПД)
1
Ионные токи во время ПД
2
3
4
5
Потенциал действия (ПД)


Какому закону подчиняется потенциал действия?
ПД подчиняется закону «все или ничего». ПД
Какому закону подчиняется потенциал действия?
ПД подчиняется закону «все или ничего». ПД

Что происходит с внешними (m-воротами) и внутренними (h-воротами) натриевых каналов во
Что происходит с внешними (m-воротами) и внутренними (h-воротами) натриевых каналов во

Возбудимость повышена.
Абсолютная рефрактерная фаза.
Относительная рефрактерная фаза.
Возбудимость повышена.
Подведем итоги. Что обозначено цифрами
Возбудимость повышена.
Абсолютная рефрактерная фаза.
Относительная рефрактерная фаза.
Возбудимость повышена.
Подведем итоги. Что обозначено цифрами

В немиелинизированных аксонах, типичных для беспозвоночных, скорость распространения потенциалов действия зависит
В немиелинизированных аксонах, типичных для беспозвоночных, скорость распространения потенциалов действия зависит

У позвоночных подавляющая часть нервных волокон, особенно в спинномозговых и черепных
У позвоночных подавляющая часть нервных волокон, особенно в спинномозговых и черепных

По мере повышения температуры примерно до 40 °С скорость проведения импульсов
По мере повышения температуры примерно до 40 °С скорость проведения импульсов

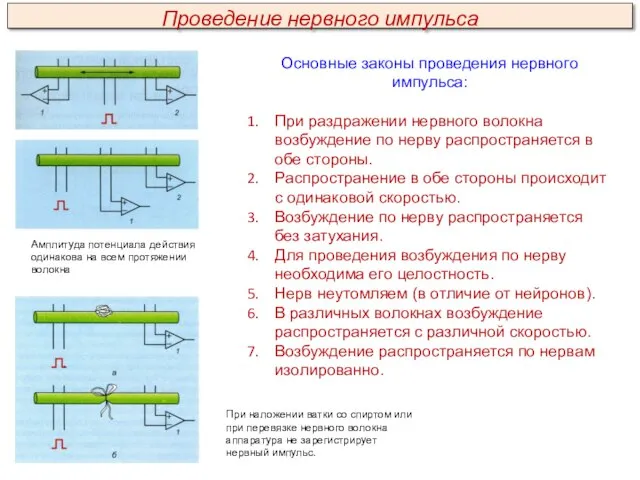
Основные законы проведения нервного импульса:
При раздражении нервного волокна возбуждение по нерву
Основные законы проведения нервного импульса:
При раздражении нервного волокна возбуждение по нерву

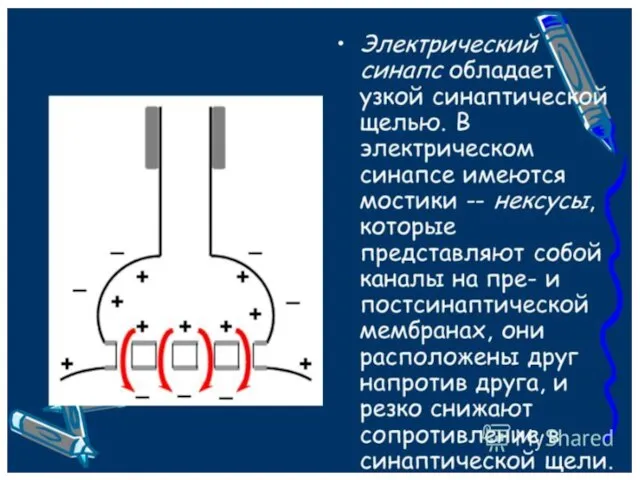
Нервные окончания могут быть рецепторными (экстерорецепторы и интерорецепторы) и эффекторными, например
Нервные окончания могут быть рецепторными (экстерорецепторы и интерорецепторы) и эффекторными, например


Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной

Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для

Инициатором сокращения миокарда, как и в скелетной мышце, является потенциал действия,
Инициатором сокращения миокарда, как и в скелетной мышце, является потенциал действия,

Считается, что потенциал действия распространяется с поверхностной мембраны кардиомиоцита вдоль Т-трубочки
Считается, что потенциал действия распространяется с поверхностной мембраны кардиомиоцита вдоль Т-трубочки

Сократительная система сердца представлена сократительными белками – актином и миозином, а
Сократительная система сердца представлена сократительными белками – актином и миозином, а
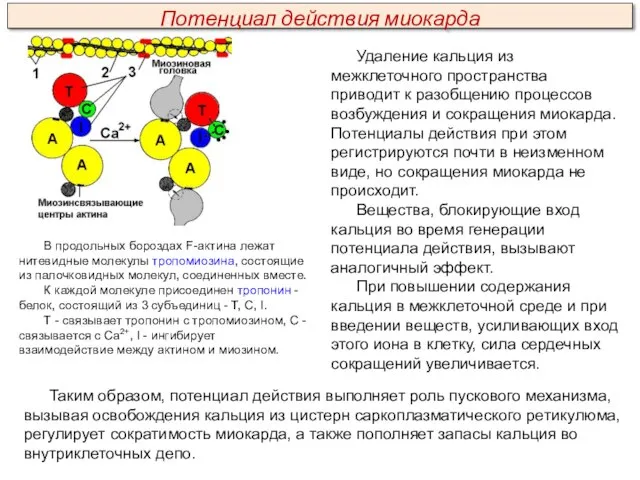
Удаление кальция из межклеточного пространства приводит к разобщению процессов возбуждения и
Удаление кальция из межклеточного пространства приводит к разобщению процессов возбуждения и

Выход потенциала действия на плато в мышечной клетке сердца связан с:
а)
Выход потенциала действия на плато в мышечной клетке сердца связан с:
а)
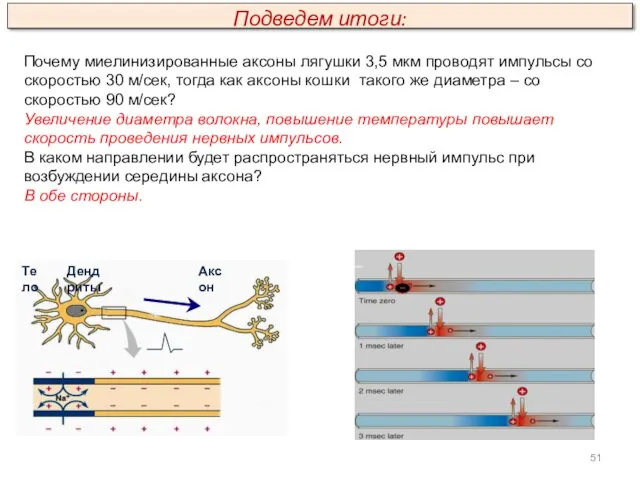
Почему миелинизированные аксоны лягушки 3,5 мкм проводят импульсы со скоростью 30
Почему миелинизированные аксоны лягушки 3,5 мкм проводят импульсы со скоростью 30

Подведем итоги. Поясните рисунки:
Подведем итоги. Поясните рисунки:
 Презентация на тему "Доказательства эволюции" - скачать бесплатно презентации по Биологии
Презентация на тему "Доказательства эволюции" - скачать бесплатно презентации по Биологии К.Линней. Его работы по систематике растений и животных.
К.Линней. Его работы по систематике растений и животных. Всеядные птицы
Всеядные птицы Возникновение жизни на Земле. Эволюция
Возникновение жизни на Земле. Эволюция Презентация на тему Типы экологических взаимодействий
Презентация на тему Типы экологических взаимодействий  Энергетический обмен веществ
Энергетический обмен веществ Сілекей
Сілекей Як утворюються перлини
Як утворюються перлини Презентация на тему "МЕЖДУНАРОДНЫЙ КОДЕКС БОТАНИЧЕСКОЙ НОМЕНКЛАТУРЫ" - скачать презентации по Биологии
Презентация на тему "МЕЖДУНАРОДНЫЙ КОДЕКС БОТАНИЧЕСКОЙ НОМЕНКЛАТУРЫ" - скачать презентации по Биологии Презентация на тему "Транспортные системы организма. Круги кровообращения" - скачать презентации по Биологии
Презентация на тему "Транспортные системы организма. Круги кровообращения" - скачать презентации по Биологии Светлой памяти доктора биологических наук, профессора, заведующего кафедрой биологии и общей генетики Ивановского государственн
Светлой памяти доктора биологических наук, профессора, заведующего кафедрой биологии и общей генетики Ивановского государственн Органы чувств человека
Органы чувств человека Основные понятия генетики
Основные понятия генетики Основы вирусологии
Основы вирусологии Сон и его значение
Сон и его значение Тема урока«Спинной мозг» 8 класс
Тема урока«Спинной мозг» 8 класс 1 апреля - Международный день птиц
1 апреля - Международный день птиц Гельминтология
Гельминтология Дыхание растений, животных и человека учитель
Дыхание растений, животных и человека учитель Великий трудолюбец Ломоносов
Великий трудолюбец Ломоносов Қазіргі жаратылыстану концепциялары. Жаратылыстану терминіне түсінік
Қазіргі жаратылыстану концепциялары. Жаратылыстану терминіне түсінік Энергетическое обеспечение биосинтеза белка и нуклеиновых кислот
Энергетическое обеспечение биосинтеза белка и нуклеиновых кислот Микроэкология тела человека
Микроэкология тела человека Индивидуальное развитие организмов - онтогенез
Индивидуальное развитие организмов - онтогенез ЧУВСТВИТЕЛЬНОСТЬ. СИНДРОМЫ ПОРАЖЕНИЯ. Доцент кафедры неврологии ПГМА, д.м.н. Каракулова Юлия
ЧУВСТВИТЕЛЬНОСТЬ. СИНДРОМЫ ПОРАЖЕНИЯ. Доцент кафедры неврологии ПГМА, д.м.н. Каракулова Юлия Презентация на тему " МЫ ЗА ЗДОРОВЫЙ ОБРАЗ ЖИЗНИ!" - скачать презентации по Биологии
Презентация на тему " МЫ ЗА ЗДОРОВЫЙ ОБРАЗ ЖИЗНИ!" - скачать презентации по Биологии Мышцы - как активная часть опорно-двигательной системы
Мышцы - как активная часть опорно-двигательной системы Тепловой режим
Тепловой режим