Сенсорные системы: общие принципы организации, разнообразие рецепторов, кодировка количества и качества сигналов
- Сенсорные системы: общие принципы организации, разнообразие рецепторов, кодировка количества и качества сигналов

Содержание
- 2. Три составляющие всякой сенсорной системы: Рецепторы (чувствительные клетки или чувствительные отростки нервных клеток) Проводящие нервы (спинномозговые
- 3. Три составляющие всякой сенсорной системы: Рецепторы (чувствительные клетки или чувствительные отростки нервных клеток) Проводящие нервы (спинномозговые
- 4. стимул Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело. В этом случае проводящий нерв образован
- 5. стимул Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело. В этом случае проводящий нерв образован
- 6. стимул Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело. ЦНС Вторично-чувствующие рецепторы: специализированные клетки (не
- 7. Стимул, как правило, вызывает открывание каналов для положительно заряженных ионов (Na+) на мембране рецептора Вход ионов
- 8. время, мс 0 -50 -70 1 2 3 подпороговый; слабый (пороговый); сильный. РП ПД Реакция на
- 9. Предыдущая схема – для первично-чувствующих рецепторов. В случае вторично-чувствующих цепь событий несколько длиннее, но результат тот
- 10. Как происходит передача сигнала от рецепторов к ЦНС? Здесь используется топический принцип: каждый рецептор передает сигнал
- 11. Как происходит передача сигнала от рецепторов в ЦНС? Здесь используется топический принцип: каждый рецептор передает сигнал
- 12. Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и качеством. Нос Ладонь Пятка 1
- 13. Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и качеством. Ситуация усложняется тем, что
- 14. Ситуация усложняется тем, что рецепторы зачастую неравномерно распределены на рецепторной поверхности и сконцентрированы в наиболее значимых
- 15. Прикосновение в точках 1 и 2 в случае спины воспринимает-ся как один стимул, в слу-чае пальца
- 16. Некоторые алгоритмы обработки информации, характерные для сенсор- ных центров головного и спинного мозга: Дивергенция сенсорных сигналов
- 17. Некоторые алгоритмы обработки информации, характерные для сенсор- ных центров головного и спинного мозга: Дивергенция сенсорных сигналов
- 18. Некоторые алгоритмы обработки информации, характерные для сенсор- ных центров головного и спинного мозга: Дивергенция сенсорных сигналов
- 19. Некоторые алгоритмы обработки информации, характерные для сенсор- ных центров головного и спинного мозга: Дивергенция сенсорных сигналов
- 20. Некоторые алгоритмы обработки информации, характерные для сенсор- ных центров головного и спинного мозга: Дивергенция сенсорных сигналов
- 21. СЕТЧАТКА: палочки и колбочки (rods and cones) – фоторецепторы; кроме того, в сетчатке находится несколько типов
- 22. Три типа колбочек (и три типа йодопсинов): красно-, зелено- и сине-чувствительные. Наследование «красного» и «зеленого» (но
- 23. Жорж Сера, «Воскресенье после полудня на острове Гранд-Жатт», 1885
- 24. Палочки и колбочки передают сигнал на ~ 1 млн. проводящих (ганглионарных) нейронов сетчатки (в зрительном нерве
- 25. Равномерная матрица фотокамеры: 144 «пикселя» Неравномерная матрица сетчатки: 72 «пикселя». В центральной ямке (Fovea) наибольшая концентрация
- 26. Зрительные центры головного мозга. Супрахиазменные ядра (передний гипоталамус) Верхние холмики четверохолмия Латеральные коленчатые тела и подушка
- 27. Зрительные иллюзии, возникающие благодаря системе латерального торможения ЛКТ.
- 28. Ретинотопические проекции, в первичную зрительную кору
- 29. Первич- ная зрит. кора (17) Вторичная и третичная зрительная кора Первичная зрительная кора получает проекции от
- 30. Жорж Брак «Кувшин и скрипка» Пабло Пикассо «Портрет женщины» Аристарх Лентулов Фильтр «Кубизм»
- 31. Первич- ная зрит. кора (17) Вторичная и третичная зрительная кора Вторичная зрительная кора: узнавание геометрических фигур,
- 32. Вторичная зрительная кора: реакция на обобщенный образ руки и отсутствие реакции на целый ряд других стимулов
- 33. Первич- ная зрит. кора (17) Вторичная и третичная зрительная кора Третичная зрительная кора: узнавание наиболее сложных
- 34. «Гештальт» – форма, структура, целостный образ. Мы узнаем сенсорный образ по сумме признаков, но для узнавания
- 35. Общий «орган чувства» (орган, где расположены рецепторы) – внутреннее ухо. Состоит из улитки (слуховая часть), а
- 36. Наружное ухо: «рупор» для сбора колебаний воздуха. Среднее ухо: энергия колебаний воздуха улавливается барабанной перепон-кой и
- 37. Улитка: чем ниже частота колебаний, тем дальше от овального окна оказывается пик «бегущей волны». В результате
- 38. Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС – тонотопические карты. Активность волосковых
- 39. Слуховые центры головного мозга. 1. Дорзальные и вентральные улитковые ядра; ядра верхней оливы. Первая стадия латерального
- 40. Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности частот (шумы, «звуки природы» и
- 41. Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных слуховых образов (музыки, речи). Узнавание
- 42. Функции различных зон новой коры: 1. Затылочная доля – зрительная кора 2. Височная доля – слуховая
- 43. 1 2 3 Нейрон, воспринима-ющий слух. образ Нейрон, воспринима-ющий зрит. образ Ассоциативный «речевой» нейрон Нейрон слухового
- 44. В 2 года в мозге ребенка около 500 речевых центров; В 3 года – около 2000:
- 45. Формирование речевой модели – еще один пример процессов обучения, то есть появления в коре новых каналов
- 47. Скачать презентацию

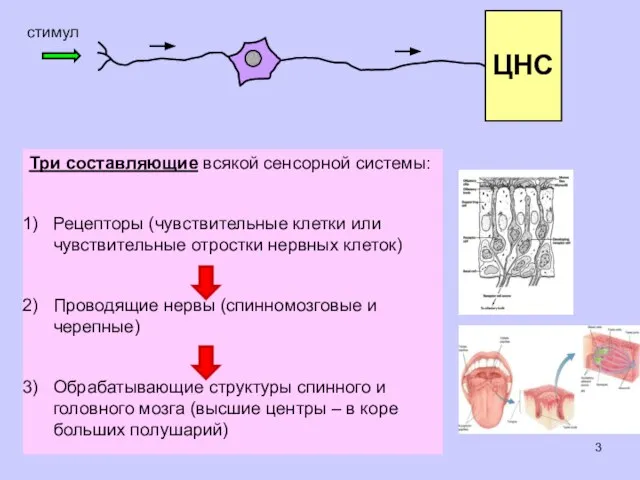
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или чувствительные отростки нервных
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или чувствительные отростки нервных
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или чувствительные отростки нервных
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или чувствительные отростки нервных
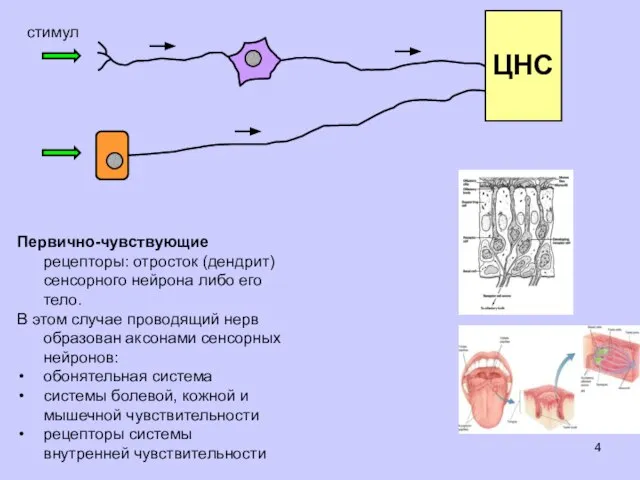
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
В этом
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
В этом
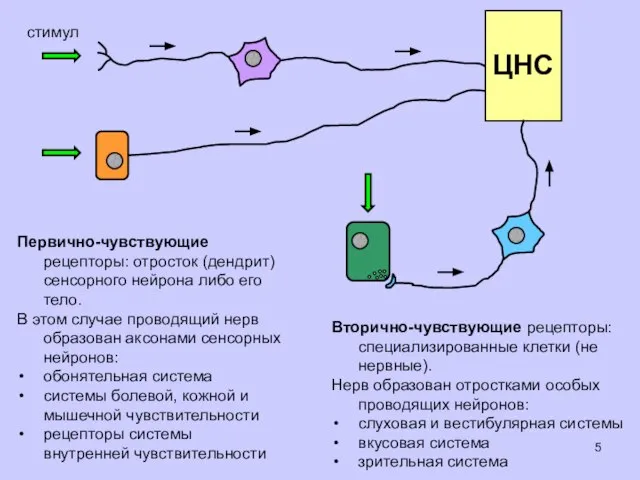
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
В этом
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
В этом
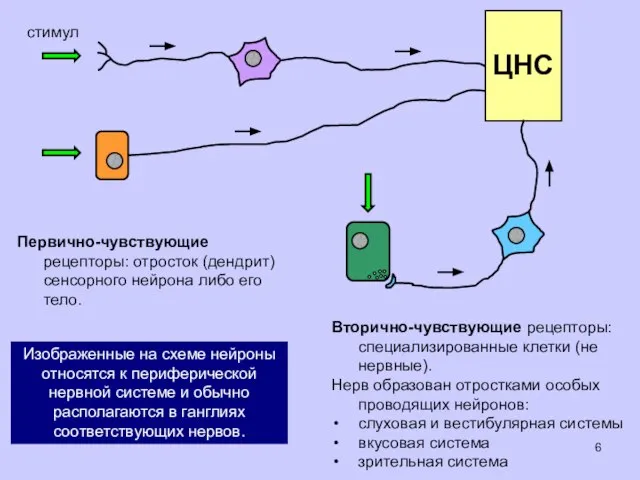
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
ЦНС
Вторично-чувствующие рецепторы:
стимул
Первично-чувствующие рецепторы: отросток (дендрит) сенсорного нейрона либо его тело.
ЦНС
Вторично-чувствующие рецепторы:
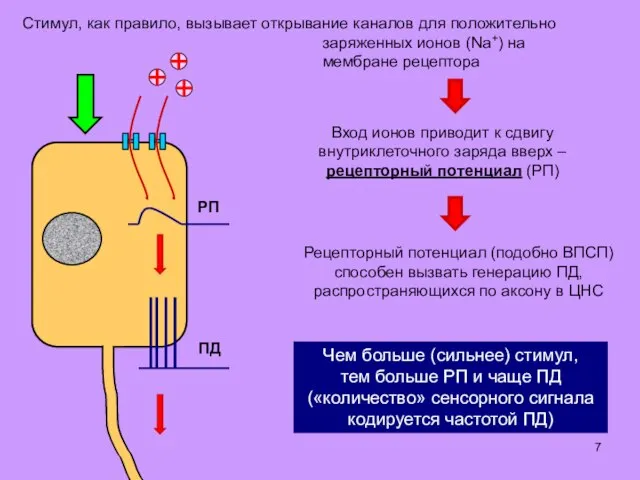
Стимул, как правило, вызывает открывание каналов для положительно заряженных ионов (Na+)
Стимул, как правило, вызывает открывание каналов для положительно заряженных ионов (Na+)
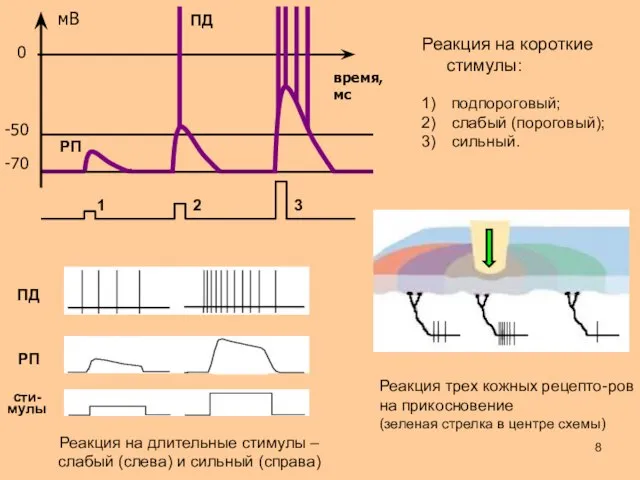
время,
мс
0
-50
-70
1
2
3
подпороговый;
слабый (пороговый);
сильный.
РП
ПД
Реакция на короткие стимулы:
время,
мс
0
-50
-70
1
2
3
подпороговый;
слабый (пороговый);
сильный.
РП
ПД
Реакция на короткие стимулы:
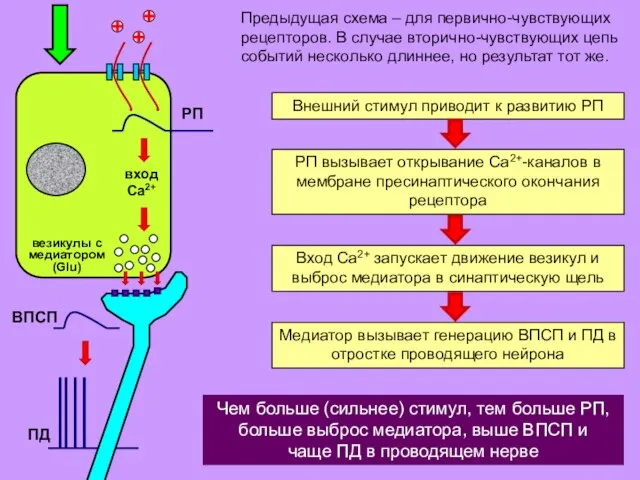
Предыдущая схема – для первично-чувствующих рецепторов. В случае вторично-чувствующих цепь событий
Предыдущая схема – для первично-чувствующих рецепторов. В случае вторично-чувствующих цепь событий
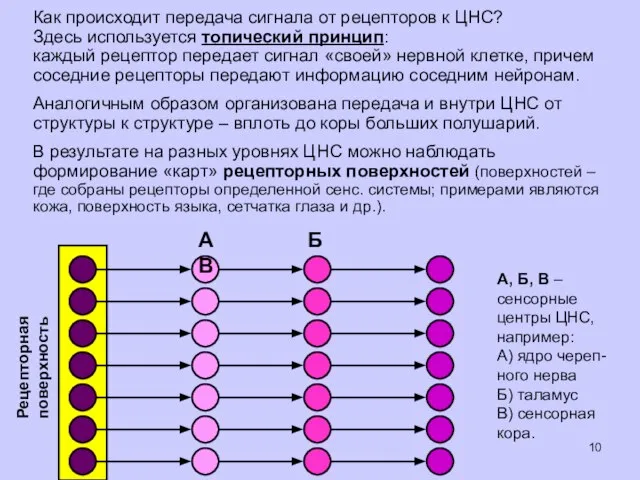
Как происходит передача сигнала от рецепторов к ЦНС?
Здесь используется топический принцип:
каждый
Как происходит передача сигнала от рецепторов к ЦНС?
Здесь используется топический принцип:
каждый
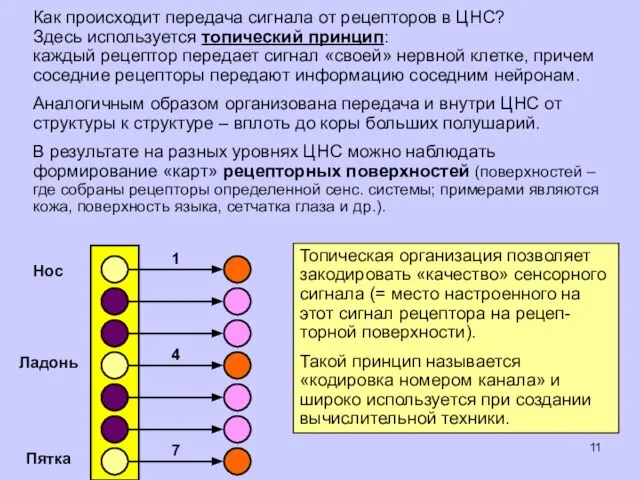
Как происходит передача сигнала от рецепторов в ЦНС?
Здесь используется топический принцип:
каждый
Как происходит передача сигнала от рецепторов в ЦНС?
Здесь используется топический принцип:
каждый
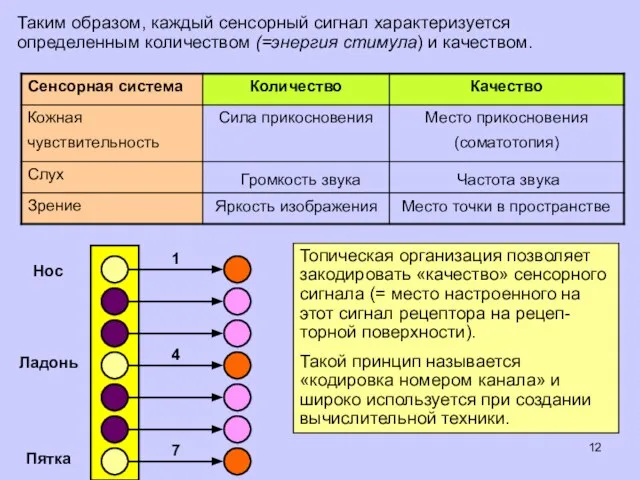
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и
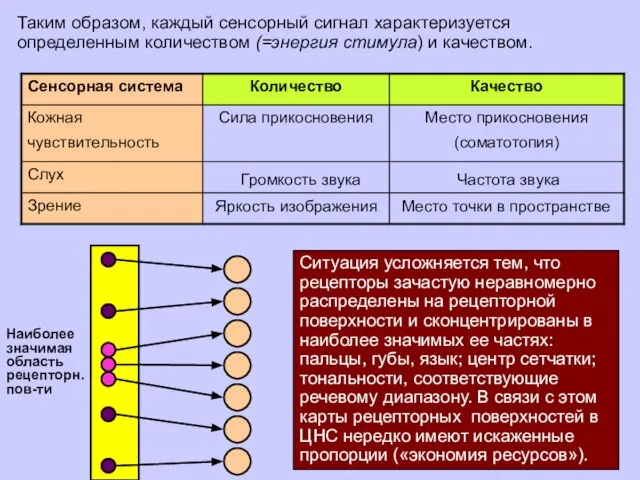
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и
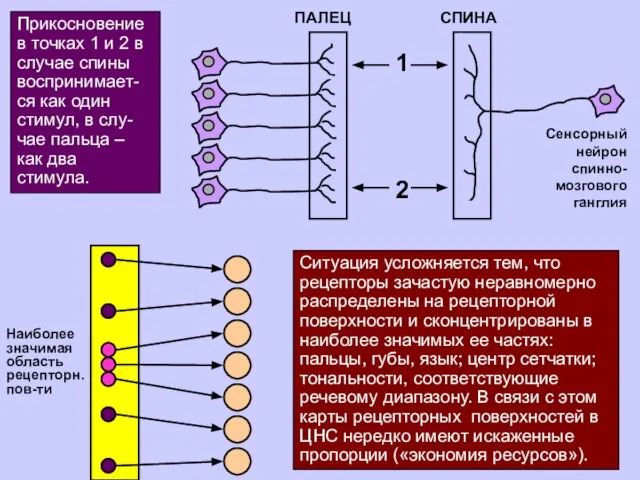
Ситуация усложняется тем, что рецепторы зачастую неравномерно распределены на рецепторной поверхности
Ситуация усложняется тем, что рецепторы зачастую неравномерно распределены на рецепторной поверхности
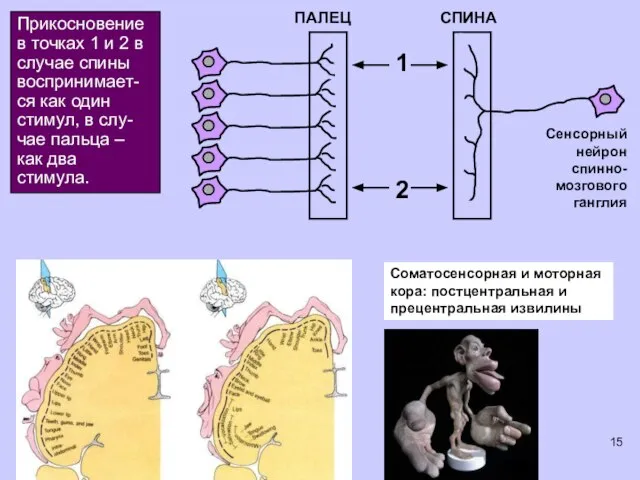
Прикосновение в точках 1 и 2 в случае спины воспринимает-ся как
Прикосновение в точках 1 и 2 в случае спины воспринимает-ся как
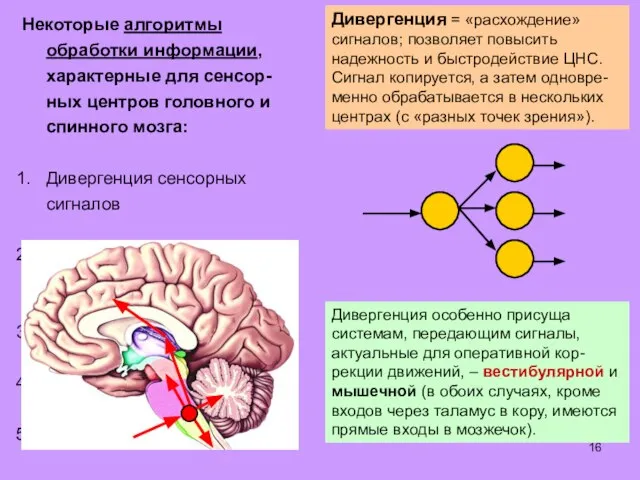
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного

Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного
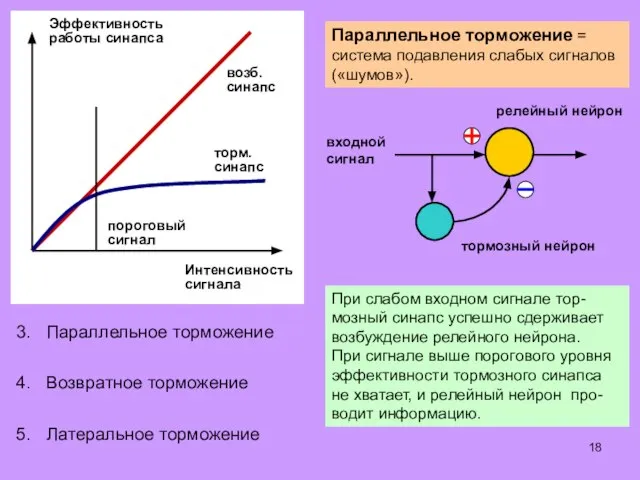
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и спинного
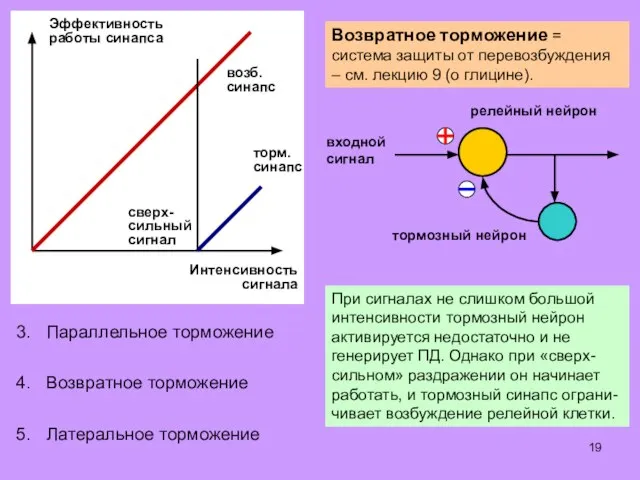
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и
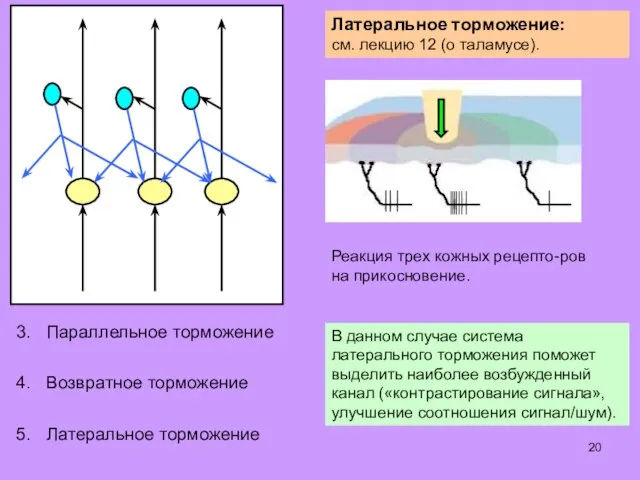
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и
Некоторые алгоритмы обработки информации, характерные для сенсор-
ных центров головного и
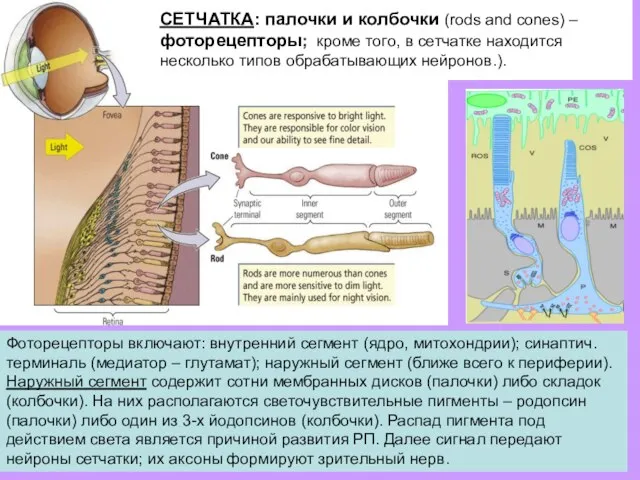
СЕТЧАТКА: палочки и колбочки (rods and cones) – фоторецепторы; кроме того,
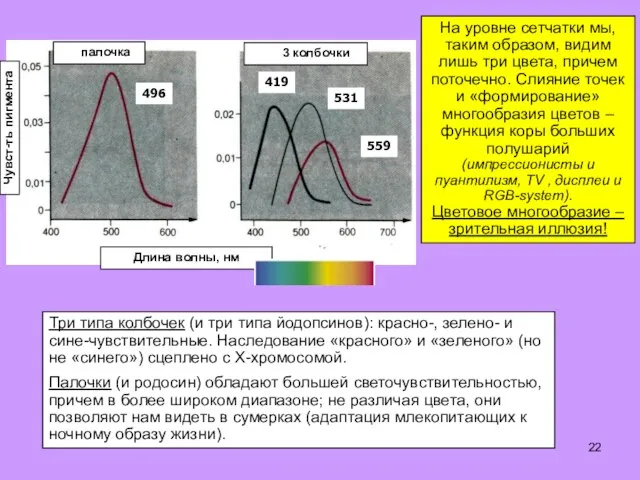
Три типа колбочек (и три типа йодопсинов): красно-, зелено- и сине-чувствительные.
Три типа колбочек (и три типа йодопсинов): красно-, зелено- и сине-чувствительные.

Жорж Сера, «Воскресенье после полудня на острове Гранд-Жатт», 1885
Жорж Сера, «Воскресенье после полудня на острове Гранд-Жатт», 1885

Палочки и колбочки передают сигнал на
~ 1 млн. проводящих (ганглионарных)
Палочки и колбочки передают сигнал на
~ 1 млн. проводящих (ганглионарных)

Равномерная матрица
фотокамеры: 144 «пикселя»
Неравномерная матрица
сетчатки: 72 «пикселя».
В центральной ямке
(Fovea)
Равномерная матрица
фотокамеры: 144 «пикселя»
Неравномерная матрица
сетчатки: 72 «пикселя».
В центральной ямке
(Fovea)

Зрительные центры головного мозга.
Супрахиазменные ядра (передний гипоталамус)
Верхние холмики четверохолмия
Латеральные коленчатые тела
Зрительные центры головного мозга.
Супрахиазменные ядра (передний гипоталамус)
Верхние холмики четверохолмия
Латеральные коленчатые тела

Зрительные иллюзии, возникающие благодаря системе латерального торможения ЛКТ.
Зрительные иллюзии, возникающие благодаря системе латерального торможения ЛКТ.
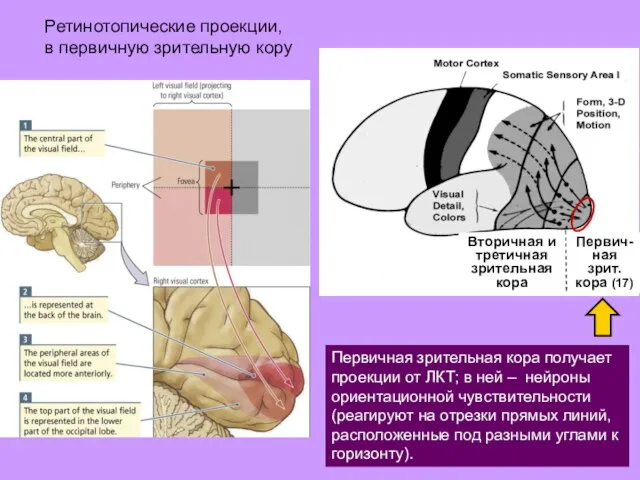
Ретинотопические проекции,
в первичную зрительную кору
Ретинотопические проекции,
в первичную зрительную кору
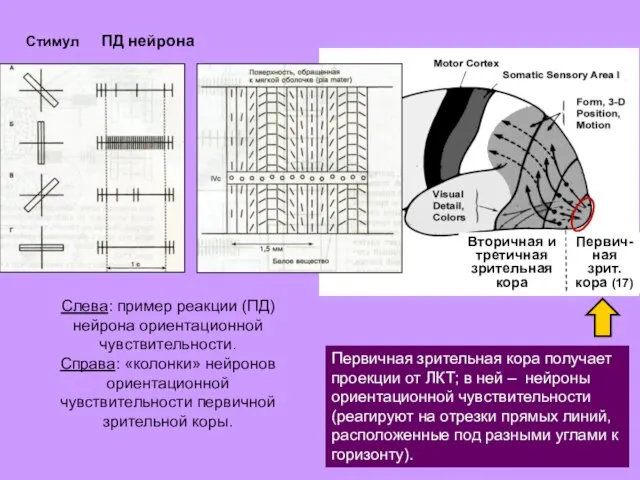
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Первичная зрительная кора получает проекции
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Первичная зрительная кора получает проекции

Жорж Брак «Кувшин и скрипка»
Пабло Пикассо «Портрет женщины»
Аристарх Лентулов
Фильтр «Кубизм»
Жорж Брак «Кувшин и скрипка»
Пабло Пикассо «Портрет женщины»
Аристарх Лентулов
Фильтр «Кубизм»
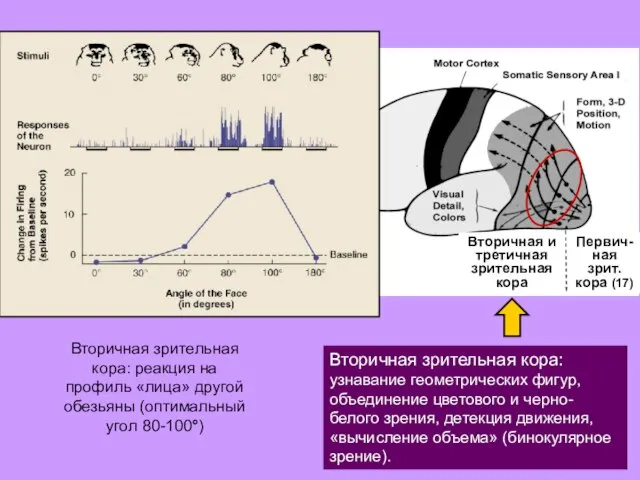
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Вторичная зрительная кора: узнавание геометрических
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Вторичная зрительная кора: узнавание геометрических
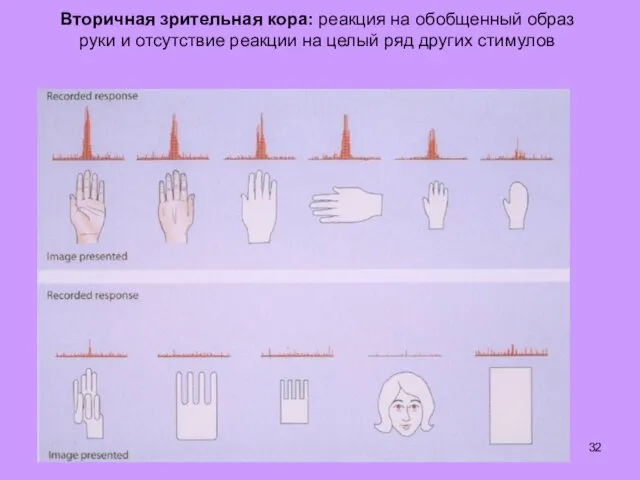
Вторичная зрительная кора: реакция на обобщенный образ руки и отсутствие реакции
Вторичная зрительная кора: реакция на обобщенный образ руки и отсутствие реакции
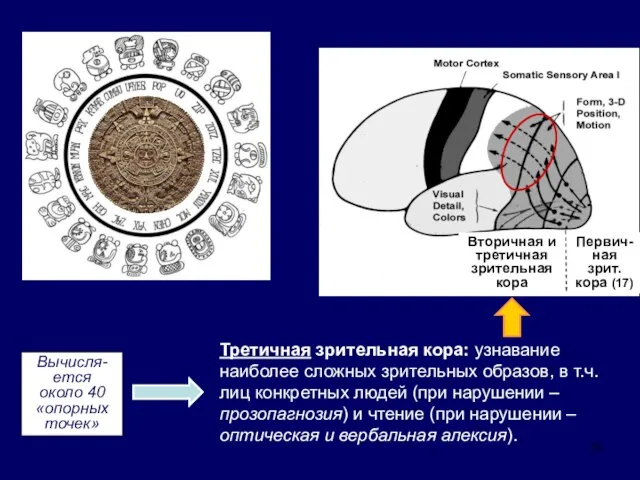
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Третичная зрительная кора: узнавание наиболее
Первич-
ная
зрит.
кора (17)
Вторичная и
третичная
зрительная
кора
Третичная зрительная кора: узнавание наиболее

«Гештальт» – форма, структура, целостный образ.
Мы узнаем сенсорный образ по сумме
«Гештальт» – форма, структура, целостный образ.
Мы узнаем сенсорный образ по сумме
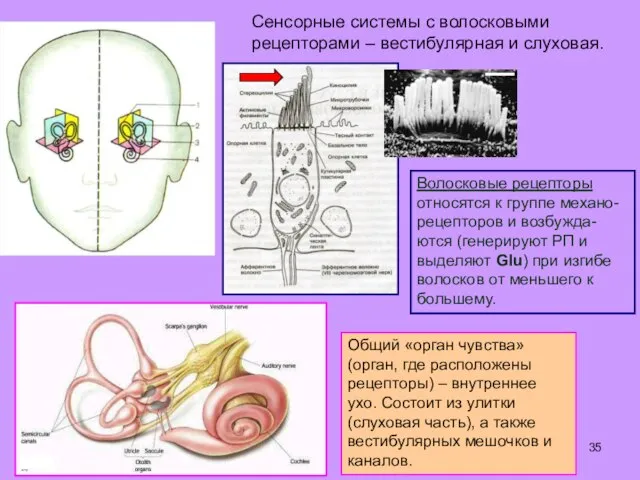
Общий «орган чувства» (орган, где расположены рецепторы) – внутреннее ухо. Состоит
Общий «орган чувства» (орган, где расположены рецепторы) – внутреннее ухо. Состоит

Наружное ухо: «рупор» для сбора колебаний воздуха.
Среднее ухо: энергия колебаний воздуха
Наружное ухо: «рупор» для сбора колебаний воздуха.
Среднее ухо: энергия колебаний воздуха
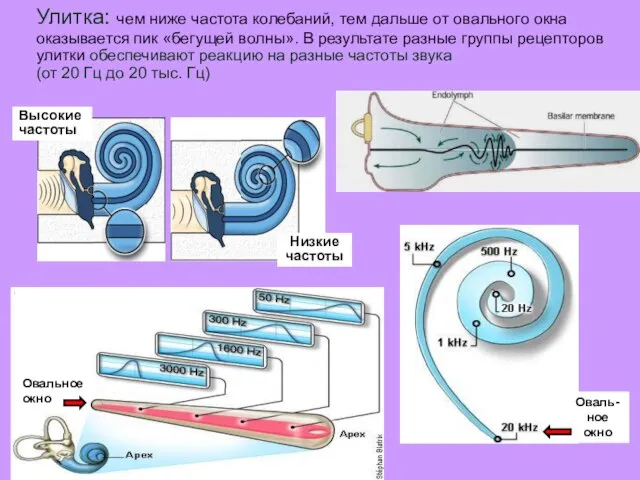
Улитка: чем ниже частота колебаний, тем дальше от овального окна оказывается
Улитка: чем ниже частота колебаний, тем дальше от овального окна оказывается
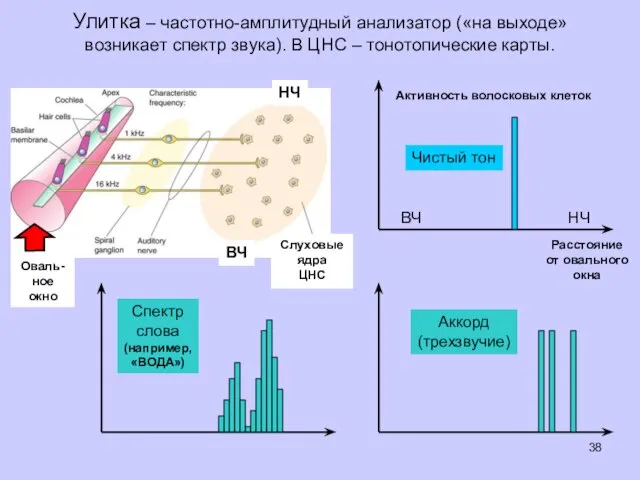
Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС
Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС
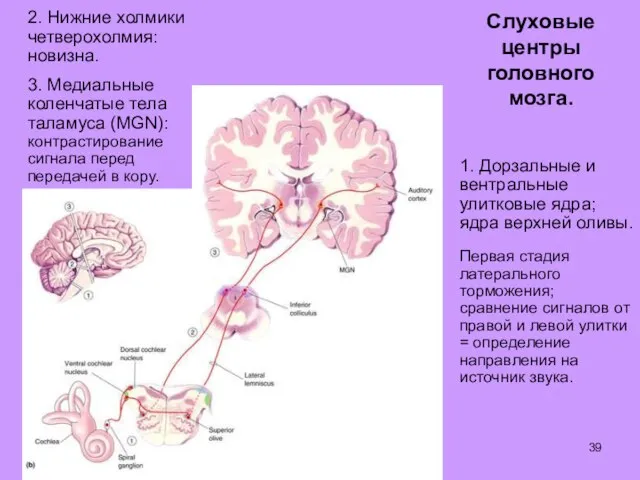
Слуховые
центры
головного
мозга.
1. Дорзальные и вентральные улитковые ядра; ядра верхней оливы.
Первая стадия
Слуховые
центры
головного
мозга.
1. Дорзальные и вентральные улитковые ядра; ядра верхней оливы.
Первая стадия
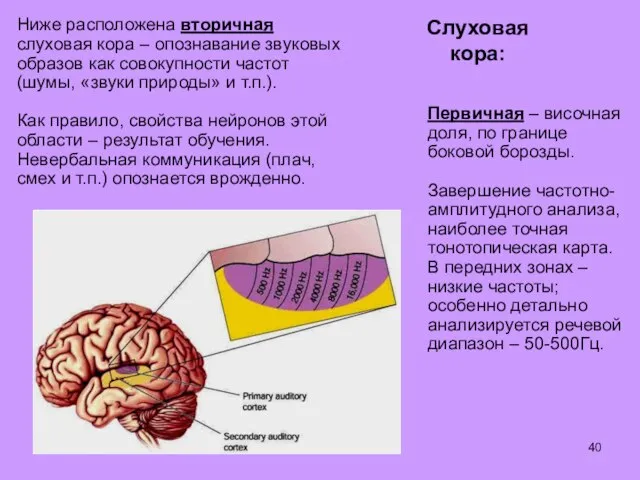
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
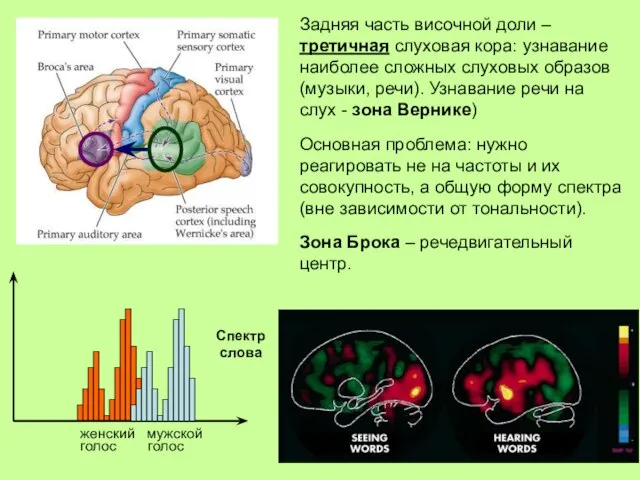
Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных
Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных
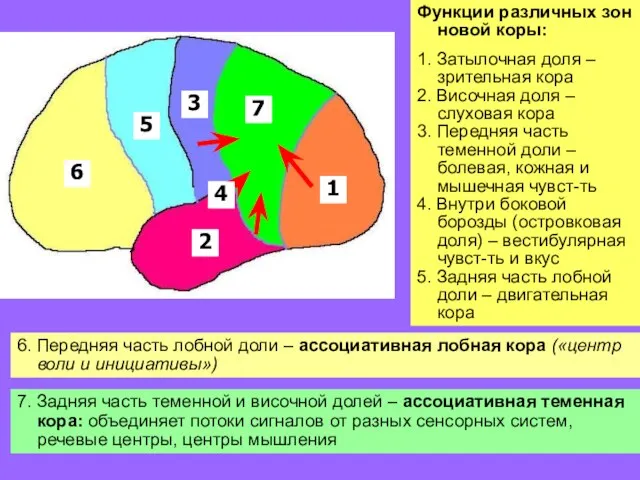
Функции различных зон новой коры:
1. Затылочная доля – зрительная кора
2. Височная
Функции различных зон новой коры:
1. Затылочная доля – зрительная кора
2. Височная
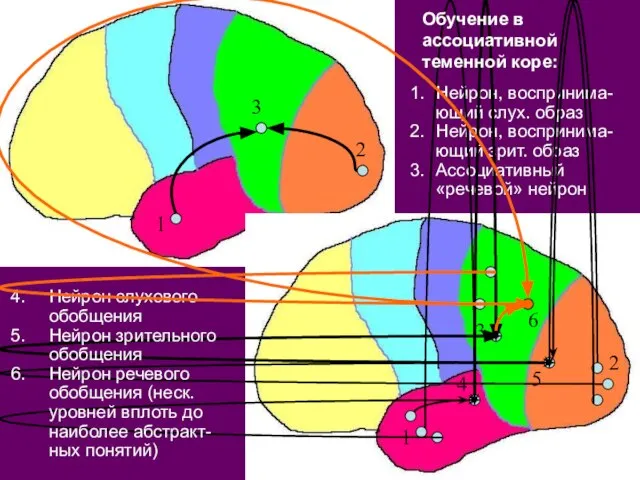
1
2
3
Нейрон, воспринима-ющий слух. образ
Нейрон, воспринима-ющий зрит. образ
Ассоциативный «речевой» нейрон
Нейрон слухового обобщения
Нейрон
1
2
3
Нейрон, воспринима-ющий слух. образ
Нейрон, воспринима-ющий зрит. образ
Ассоциативный «речевой» нейрон
Нейрон слухового обобщения
Нейрон
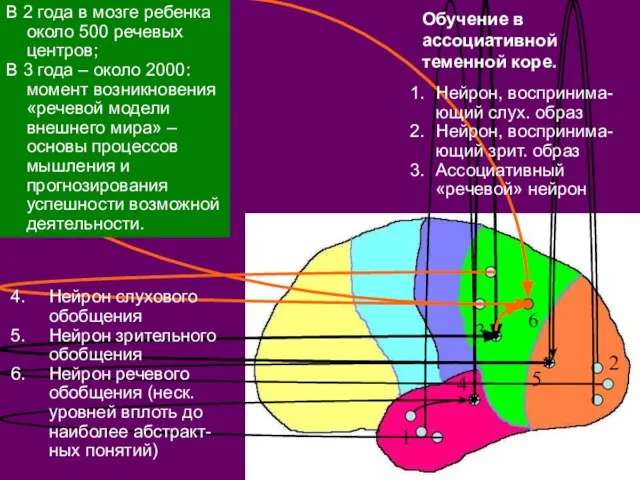
В 2 года в мозге ребенка около 500 речевых центров;
В 3
В 2 года в мозге ребенка около 500 речевых центров;
В 3
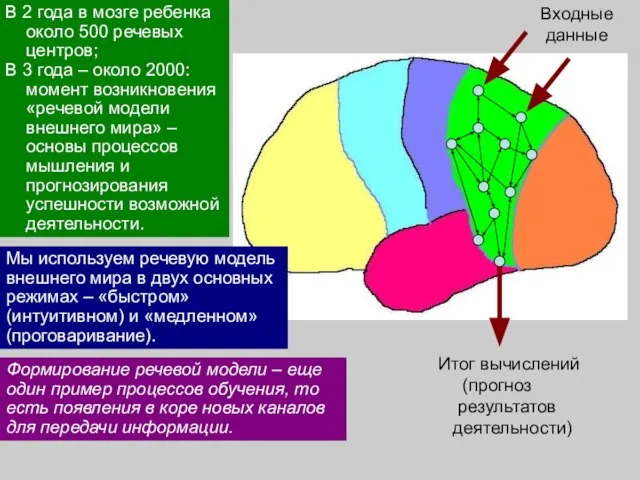
Формирование речевой модели – еще один пример процессов обучения, то есть
Формирование речевой модели – еще один пример процессов обучения, то есть
 Развитие представлений о происхождении жизни на Земле. Геологические эры и развитие органического мира
Развитие представлений о происхождении жизни на Земле. Геологические эры и развитие органического мира Презентация на тему "Карл Линней" - скачать презентации по Биологии
Презентация на тему "Карл Линней" - скачать презентации по Биологии Питьевой режим
Питьевой режим Профилактика туберкулёза Митихина М.С. г. Курск
Профилактика туберкулёза Митихина М.С. г. Курск  Қолдан сұрыптау
Қолдан сұрыптау Древние растения и животные Кропачева Марина Васильевна учитель начальных классов МАОУ «СОШ №12 с углубленным изучением немец
Древние растения и животные Кропачева Марина Васильевна учитель начальных классов МАОУ «СОШ №12 с углубленным изучением немец История изучения клетки
История изучения клетки Презентация на тему "Тема урока : " Насекомые "" - скачать бесплатно презентации по Биологии
Презентация на тему "Тема урока : " Насекомые "" - скачать бесплатно презентации по Биологии Psikhologia_Mozg
Psikhologia_Mozg Theory of Phylembryogenesis
Theory of Phylembryogenesis Ткани животных
Ткани животных Транспорт веществ в живом организме Урок биологии в 11 классе
Транспорт веществ в живом организме Урок биологии в 11 классе Анатомия органов мочевыделительной системы. Надпочечники. Забрюшинное пространство
Анатомия органов мочевыделительной системы. Надпочечники. Забрюшинное пространство Процесс рибосомной РНК
Процесс рибосомной РНК Введение в историю и методологию биологии
Введение в историю и методологию биологии ДНК в генной инженерии
ДНК в генной инженерии Учение об эволюции
Учение об эволюции Эволюция человека (антропогенез). Движущие силы антропогенеза
Эволюция человека (антропогенез). Движущие силы антропогенеза Историческое развитие растений на Земле
Историческое развитие растений на Земле Презентация на тему Горячая рыба
Презентация на тему Горячая рыба  Презентация на тему Мир растений
Презентация на тему Мир растений  Ткани организма человека (3 и 4 часть)
Ткани организма человека (3 и 4 часть) Аттестационная работа. Элементы исследовательской работы, используемые при изучении дисциплины Биология
Аттестационная работа. Элементы исследовательской работы, используемые при изучении дисциплины Биология Домашнее задание: § 23, 31
Домашнее задание: § 23, 31 Проблема здоровья людей
Проблема здоровья людей Классы грузов
Классы грузов Класс Птицы
Класс Птицы Яснотковые. Биологическое описание
Яснотковые. Биологическое описание