- Сцепленное наследование

Содержание
- 2. Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один
- 3. Пример нарушения закона независимого комбинирования признаков впервые был обнаружен У. Бэтсоном и Р. Пеннетом при изучении
- 4. В 1909 г. к детальному изучению этого вопроса приступил Т. Г. Морган. Прежде всего он четко
- 5. Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и
- 6. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны
- 7. Кроссинговер Кроссинговер (от англ. crossingover), перекрест, взаимный обмен участками парных хромосом, происходящий в результате разрыва и
- 8. Кроссинговер в грубом приближении зависит от линейного расстояния между генами. В случае, если на участке между
- 9. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит
- 10. Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел. Кроссоверные гаметы — гаметы, в
- 11. Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей. Рекомбинанты —
- 12. Внутрихромосомная локализация генов. Генетические карты По мере обнаружения все новых и новых мутаций увеличивался объем сведений
- 13. Частота рекомбинаций отражает относительное расположение генов в хромосоме и вычисляется по формуле Данные о частотах рекомбинаций
- 14. На практике обычно определяют частоту рекомбинации по меньшей мере для трех генов одновременно; этот метод, называемый
- 15. При построении хромосомных карт могут возникнуть затруднения, создаваемые двойным кроссинговером; особенно это касается тех случаев, когда
- 16. Генетические карты (группы сцепления) дрозофилы. Номера групп сцепления обозначены римскими цифрами. Цифры на генетических картах обозначают
- 17. Мутация vg(рецессивная мутация) Мухи с такой мутацией неспособны летать. Мутация lac (возникновение вместо антенн на голове
- 18. Факты, создавшие фундамент хромосомной теории наследственности Они были установлены при исследовании таких проблем, как: хромосомный механизм
- 20. Скачать презентацию

Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно,
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно,

Пример нарушения закона независимого комбинирования признаков впервые был обнаружен У. Бэтсоном
Пример нарушения закона независимого комбинирования признаков впервые был обнаружен У. Бэтсоном

В 1909 г. к детальному изучению этого вопроса приступил Т. Г.
В 1909 г. к детальному изучению этого вопроса приступил Т. Г.
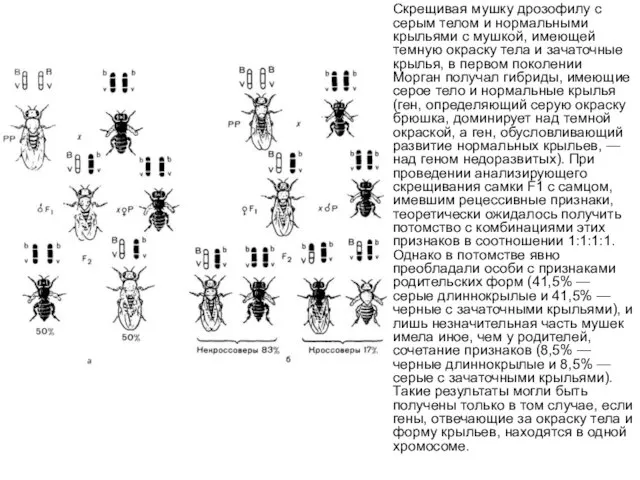
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой,
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой,
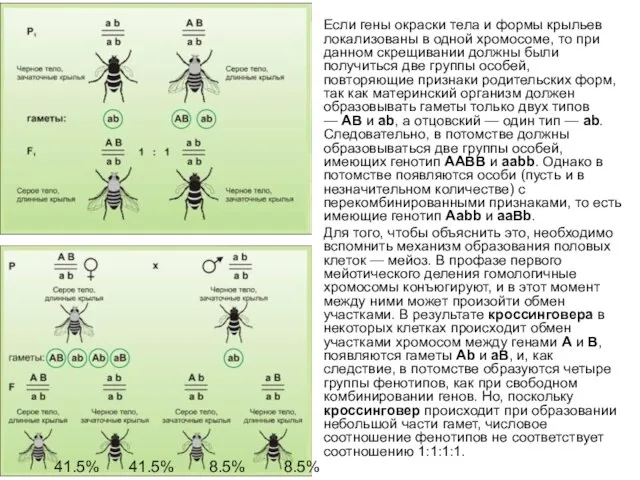
Если гены окраски тела и формы крыльев локализованы в одной хромосоме,
Если гены окраски тела и формы крыльев локализованы в одной хромосоме,
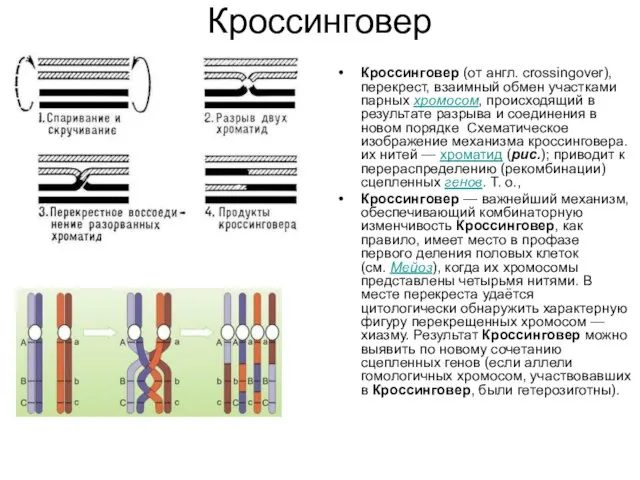
Кроссинговер
Кроссинговер (от англ. crossingover), перекрест, взаимный обмен участками парных хромосом, происходящий в результате
Кроссинговер
Кроссинговер (от англ. crossingover), перекрест, взаимный обмен участками парных хромосом, происходящий в результате

Кроссинговер в грубом приближении зависит от линейного расстояния между генами. В случае,
Кроссинговер в грубом приближении зависит от линейного расстояния между генами. В случае,
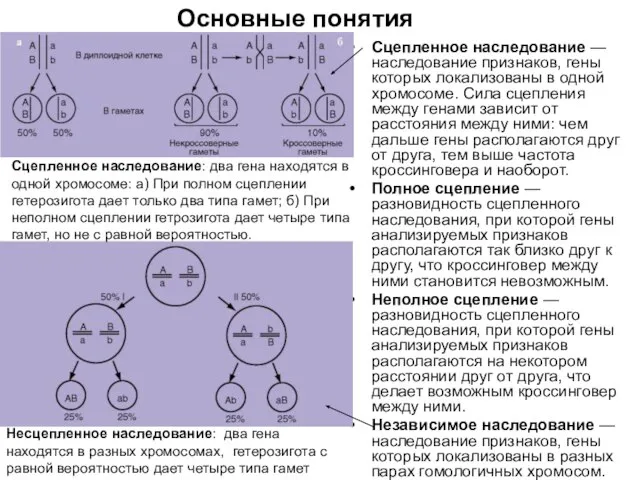
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила
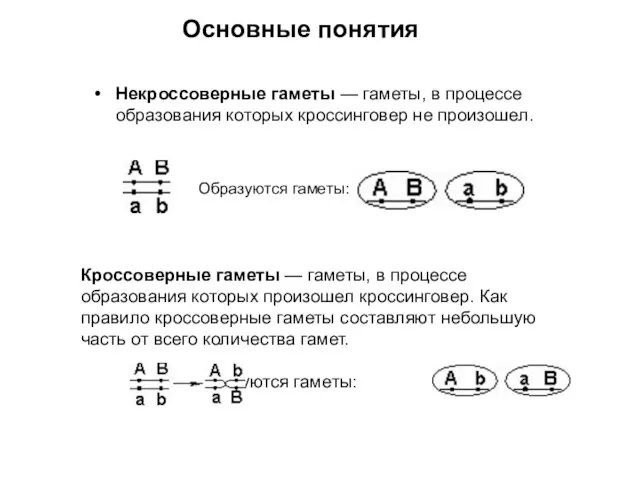
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Кроссоверные гаметы —
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Кроссоверные гаметы —

Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и

Внутрихромосомная локализация генов. Генетические карты
По мере обнаружения все новых и новых
Внутрихромосомная локализация генов. Генетические карты
По мере обнаружения все новых и новых

Частота рекомбинаций отражает относительное расположение генов в хромосоме и вычисляется по
Частота рекомбинаций отражает относительное расположение генов в хромосоме и вычисляется по
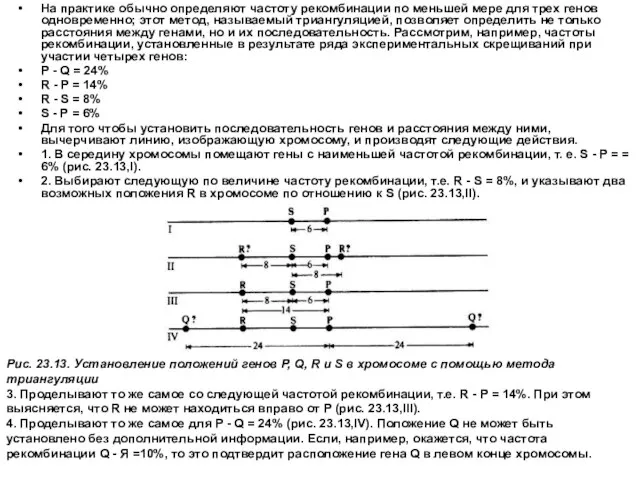
На практике обычно определяют частоту рекомбинации по меньшей мере для трех
На практике обычно определяют частоту рекомбинации по меньшей мере для трех
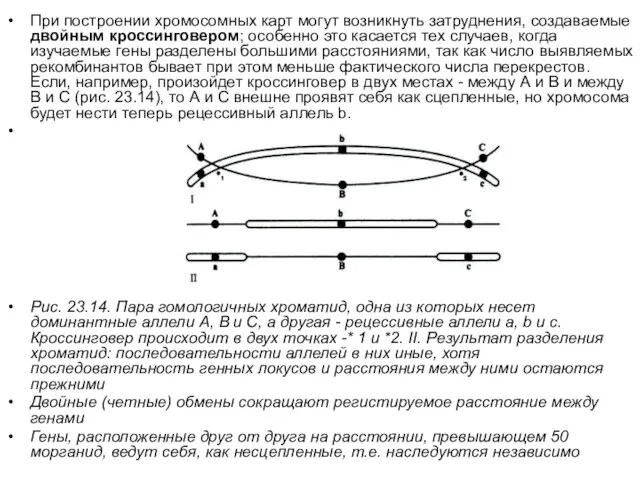
При построении хромосомных карт могут возникнуть затруднения, создаваемые двойным кроссинговером; особенно
При построении хромосомных карт могут возникнуть затруднения, создаваемые двойным кроссинговером; особенно

Генетические карты (группы сцепления) дрозофилы.
Номера групп сцепления обозначены римскими цифрами. Цифры на
Генетические карты (группы сцепления) дрозофилы.
Номера групп сцепления обозначены римскими цифрами. Цифры на

Мутация vg(рецессивная мутация)
Мухи с такой мутацией неспособны
летать. Мутация lac (возникновение
вместо антенн
Мутация vg(рецессивная мутация) Мухи с такой мутацией неспособны летать. Мутация lac (возникновение вместо антенн
Факты, создавшие фундамент хромосомной теории наследственности
Они были установлены при исследовании таких
Факты, создавшие фундамент хромосомной теории наследственности
Они были установлены при исследовании таких
 МИКРОСКОПИЧЕСКОЕ СТРОЕНИЕ ЗУБА План лекции: Общий план строения зуба. Тканевой состав зуба. Источники формирования зу
МИКРОСКОПИЧЕСКОЕ СТРОЕНИЕ ЗУБА План лекции: Общий план строения зуба. Тканевой состав зуба. Источники формирования зу История развития животного мира
История развития животного мира Презентация на тему СТРУКТУРА И ФУНКЦИИ ХРОМОСОМ
Презентация на тему СТРУКТУРА И ФУНКЦИИ ХРОМОСОМ Презентация Экология человека и промышленное загрязнение окружающей среды
Презентация Экология человека и промышленное загрязнение окружающей среды  Лекция 6-7. Развитие половых клеток. Оплодотворение. Развитие зародыша
Лекция 6-7. Развитие половых клеток. Оплодотворение. Развитие зародыша Изменчивость. Готовимся к ЕГЭ
Изменчивость. Готовимся к ЕГЭ Презентация на тему Антропоген
Презентация на тему Антропоген 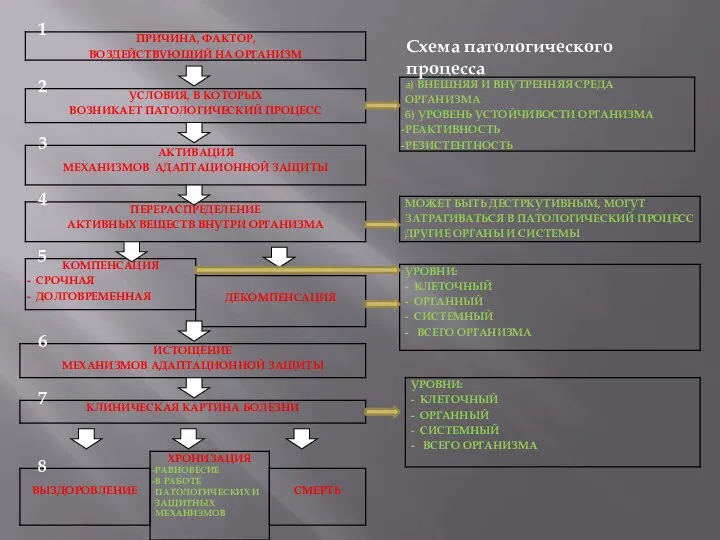 Схема патологического процесса
Схема патологического процесса Картофель. Сортовые признаки
Картофель. Сортовые признаки Медицинская арахноэнтомология. Тип членистоногие. Класс ракообразные. Класс паукообразные. Класс насекомые
Медицинская арахноэнтомология. Тип членистоногие. Класс ракообразные. Класс паукообразные. Класс насекомые Общая характеристика червей. Тип Плоские Черви. Класс Ресничные черви
Общая характеристика червей. Тип Плоские Черви. Класс Ресничные черви Фотосистема ІІ
Фотосистема ІІ  Основные типы экологических взаимодействий. Конкуренция
Основные типы экологических взаимодействий. Конкуренция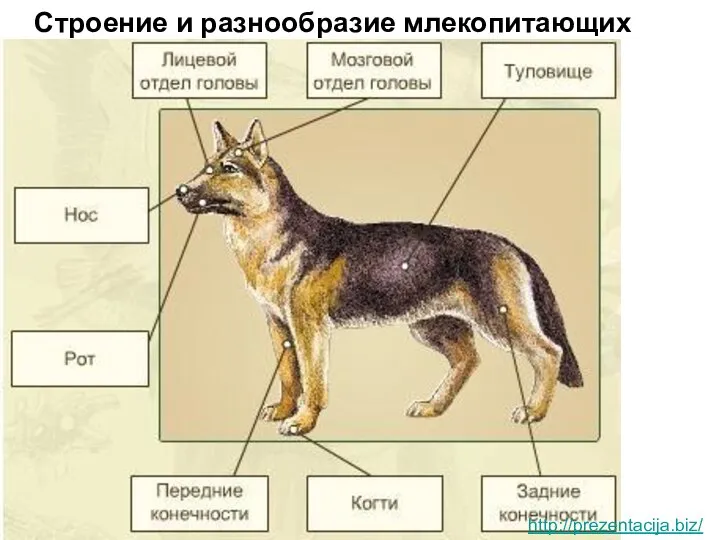 Строение и разнообразие млекопитающих
Строение и разнообразие млекопитающих 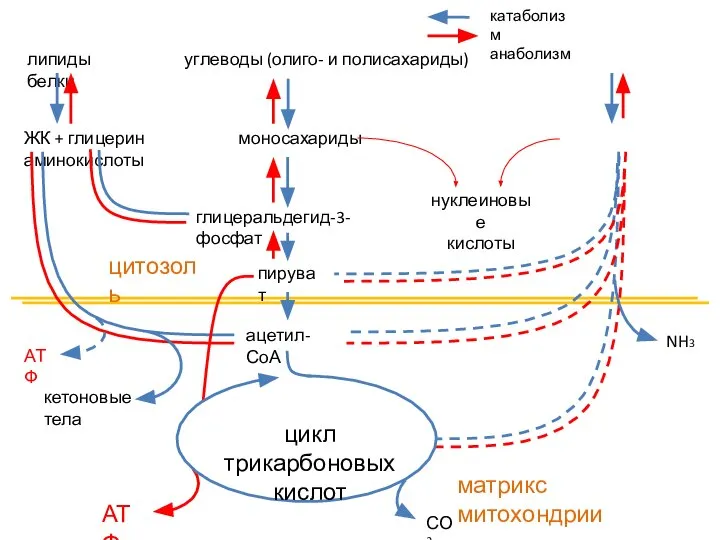 Биоокисление и цикл Кребса
Биоокисление и цикл Кребса Углеводы простые (быстрые) и сложные(медленные)
Углеводы простые (быстрые) и сложные(медленные) Как появился человек на земле
Как появился человек на земле Природные ресурсы. Рациональное природопользование. Тема 59
Природные ресурсы. Рациональное природопользование. Тема 59 Тема урока : «Строение и функции нервной системы»
Тема урока : «Строение и функции нервной системы» Отдел Glomeromycota
Отдел Glomeromycota Щука, квакша, крокодил, голубь, медведь
Щука, квакша, крокодил, голубь, медведь Тест по теме Пресмыкающиеся
Тест по теме Пресмыкающиеся Основы нейронных сетей
Основы нейронных сетей Презентация на тему "Алоэ. Зелёный доктор на вашем окне" - скачать презентации по Биологии
Презентация на тему "Алоэ. Зелёный доктор на вашем окне" - скачать презентации по Биологии Атлас редких и краснокнижных видов Центрально-Чернозёмного заповедника
Атлас редких и краснокнижных видов Центрально-Чернозёмного заповедника Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы
Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы Соединения костей черепа, позвонков, ребер, грудины
Соединения костей черепа, позвонков, ребер, грудины Анатомия и физиология женских половых органов. Женский таз
Анатомия и физиология женских половых органов. Женский таз