- Свойства нервных центров. Лекция 6

Содержание
- 2. Электромеханическое сопряжение в скелетном миоците
- 3. Электромеханическое сопряжение Совокупность явлений, обусловливающих связь между возбуждением (потенциалом действия) и сокращением мышечных волокон
- 4. Виды электромеханического сопряжения В цистернах ретикулума имеется несколько типов Са2+ -каналов, по которым Са2+ выходит в
- 5. Схема ЭМС: А. Расслабленное мышечное волокно с поляризованной клеточной мембраной. Концентрация Ca 2+ внутри клетки ниже
- 6. Последовательность процессов при ЭМС Раздражение. Возникновение ПД. Проведение его вдоль клеточной мембраны и вглубь волокна по
- 7. Цикл возбуждение – сокращение – расслабление в скелетных мышцах
- 8. Расслабление Ca2+‑АТФаза СПР закачивает Са2+ из саркоплазмы в цистерны ретикулума Ca2+ связывается с кальсеквестрином. Са2+ -насосы
- 9. Роль АТФ в мышечном расслаблении Поскольку АТФ поставляет энергию для сокращения, предполагалось, что удаление АТФ должно
- 10. УТОМЛЕНИЕ МЫШЦ Утомление - это временное снижение работоспособности мышцы в результате работы. Утомление изолированной мышцы можно
- 11. 1.Теория Шиффа: - следствие истощения энергетических запасов в мышце. 2. Теория Пфлюгера: накопление в мышце продуктов
- 12. Особенности расположения сократительных филаментов в гладком миоците
- 13. Особенности расположения сократительных филаментов в гладком миоците Нити не организованы в саркомеры. Тонкие филаменты прикреплены к
- 14. Физиологические особенности гладких мышц Менее упорядоченно расположены сократительные белки. ПП -60/ -70 мВ. ПД натрий-кальций-калиевой природы.
- 15. Основные этапы сокращения гладких мышц Figure 12-28: Smooth muscle contraction Увеличение внутриклеточной концентрации иона кальция Са
- 16. Мышечная ткань В организме человека в среднем приходится на долю: - скелетных мышц – 40-50% массы
- 17. Энергетика мышечного сокращения
- 18. ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ Ресинтез АТФ происходит 2 основными путями: анаэробным аэробным
- 19. ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ 1. Фосфогенная, или АТФ-КФ-система; 2. Лактацидная, или гликолитическая; 3. Кислородная, или окислительная.
- 20. Три системы синтеза и ресинтеза АТФ при мышечном сокращении: 1 – фосфогенная, 2 – гликолитическая, 3
- 21. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ, КООРДИНАЦИОННЫЕ ПРИНЦИПЫ в ЦНС
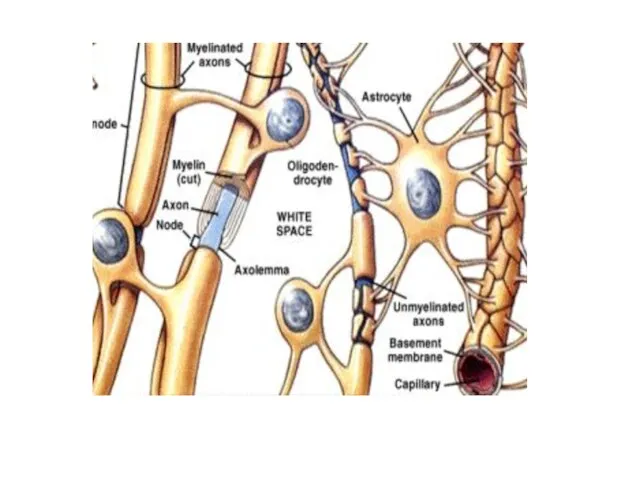
- 23. Нейрогли́я, или просто гли́я — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС
- 24. Клетки нейроглии (астроциты, олигодендроциты, микроглия) заполняют все пространство между нейронами, защищая их от механических повреждений (опорная
- 26. РЕФЛЕКС. РЕФЛЕКТОРНАЯ ДУГА Рефлекс — это ответная реакция организма на раздражение, происходящая при участии нервной системы.
- 29. Классификации рефлексов: По биологической значимости : пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы. По типу рецепторов,: экстероцептивные,
- 30. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ 1. Одностороннее проведение возбуждения. 2. Центральная задержка рефлекса. 3. Последействие (кратковременное и реверберация)
- 31. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ одностороннее проведение возбуждения (химические синапсы), центральная задержка проведения возбуждения (химические синапсы),
- 32. 3. Последействие : кратковременное - связано с особенностью синаптической передачи (медиатор не сразу прекращает действовать, а
- 33. 4. Временная и пространственная суммация
- 34. Центральное облегчение: при одновременном раздражении двух афферентов с перекрывающимися подпороговыми зонами количество возбужденных нейронов больше, чем
- 35. Центральная окклюзия: при одновременном раздражении двух афферентов с перекрывающимися пороговыми зонами количество возбужденных нейронов меньше, чем
- 36. усвоение и трансформация ритма усвоение необходимо для ритмичной работы НЦ (например, для ритмических движений). трансформация ритма
- 37. Повышение ритма может быть связано с циркуляцией возбуждения в нейронных ловушках; Снижение ритма обусловлено лабильностью нейронов
- 38. 6. фоновая активность обеспечивается: поступлением импульсов от соседних НЦ (дивергенция, конвергенция); суммацией МПКП, возникающих за счет
- 39. Опыт Бронджеста Тонус нервного центра При перерезке у лягушки чувствительных корешков сегментов спинного мозга, инервирующих задние
- 40. утомление в НЦ возникает из-за: истощения медиатора; истощения АТФ и глюкозы; снижения чувствительности рецепторов; метаболических сдвигов
- 41. 10. Посттетаническая потенциация Если , после интенсивного раздражения, дать синапсу отдохнуть и нанести одиночное раздражение, то
- 42. 11. Пластичность: Взаимосвязь между нейронами обеспечивается синапсами. Для повышения эффективности синаптической передачи существует два способа: гомосинаптическая
- 43. Гомосинаптическая модуляция частое (тетаническое) раздражение пресинаптической мембраны ↓ повышение [Са2+ ] в синаптическом окончании ↓ увеличение
- 44. Гомосинаптическая модуляция На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к медиатору и, как следствие,
- 45. Гетеросинаптическая модуляция Этапы: Выделение медиатора →↑ цАМФ → инактивация К+-каналов → удлинение фазы деполяризации ПД →
- 46. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС 1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ (по Шеррингтону) 2. ДИВЕРГЕНТНОСТИ
- 48. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
- 49. РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
- 50. Обратные связи Положительные Отрицательные импульсы с периферии, импульсы с периферии, возникающие в возникающие в результате какой-либо
- 51. РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
- 52. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ и гамма-петля
- 53. ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А.А.Ухтомскому, 1931) Доминанта - временно господствующий рефлекс или поведенческий акт, которым трансформируется
- 54. Основные cвойства доминанты ( по А.А.Ухтомскому) 1. Повышенная возбудимость доминантного центра 2. Способность к суммации (подкреплять
- 55. Виды доминант Внутренние доминанты – ведущую роль играет гормональный фон. Внешние доминанты – ведущую роль играет
- 56. Причины прекращения существования доминанты Достижение результата Появление новой, более значимой доминанты Запредельное торможение
- 57. ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
- 58. ПРИНЦИП РЕЦИПРОКНОСТИ + + - - +
- 59. Динамика основных нервных процессов: ИНДУКЦИЯ: Процессы иррадиации и концентрации составляют основу индукционных отношений в ЦНС. Индукция
- 60. 9. Принцип индукции Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию. одновременная отрицательная индукция одновременная положительная
- 61. От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений скоростного характера, требующих быстрой смены
- 63. Скачать презентацию

Электромеханическое сопряжение в скелетном миоците
Электромеханическое сопряжение в скелетном миоците

Электромеханическое сопряжение
Совокупность явлений, обусловливающих связь между возбуждением (потенциалом действия) и сокращением
Электромеханическое сопряжение
Совокупность явлений, обусловливающих связь между возбуждением (потенциалом действия) и сокращением
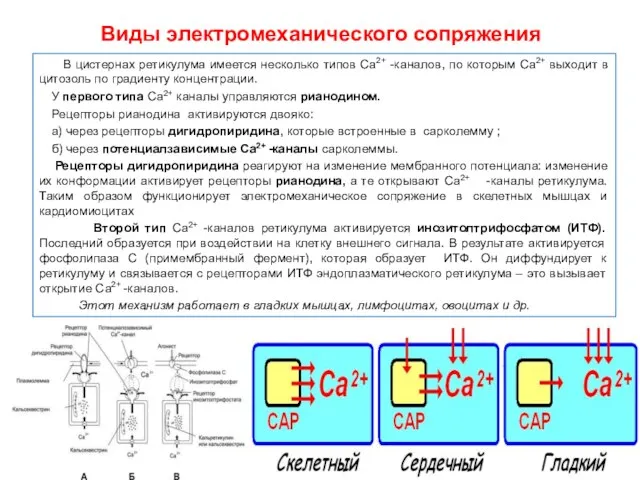
Виды электромеханического сопряжения
В цистернах ретикулума имеется несколько типов Са2+ -каналов,
Виды электромеханического сопряжения
В цистернах ретикулума имеется несколько типов Са2+ -каналов,
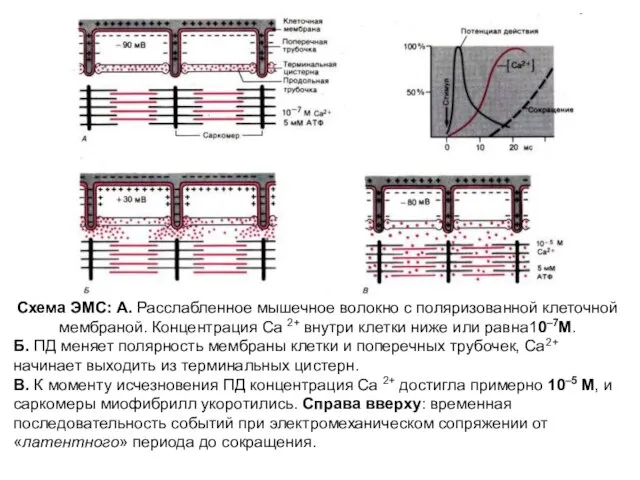
Схема ЭМС: А. Расслабленное мышечное волокно с поляризованной клеточной мембраной. Концентрация
Схема ЭМС: А. Расслабленное мышечное волокно с поляризованной клеточной мембраной. Концентрация

Последовательность процессов при ЭМС
Раздражение.
Возникновение ПД.
Проведение его вдоль клеточной мембраны
Последовательность процессов при ЭМС
Раздражение.
Возникновение ПД.
Проведение его вдоль клеточной мембраны
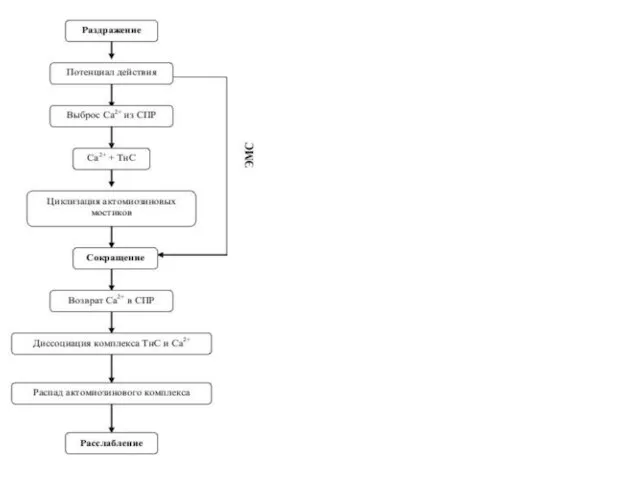
Цикл
возбуждение – сокращение – расслабление
в скелетных мышцах
Цикл
возбуждение – сокращение – расслабление
в скелетных мышцах

Расслабление
Ca2+‑АТФаза СПР закачивает Са2+ из саркоплазмы в цистерны ретикулума
Ca2+ связывается с
Расслабление
Ca2+‑АТФаза СПР закачивает Са2+ из саркоплазмы в цистерны ретикулума
Ca2+ связывается с

Роль АТФ в мышечном расслаблении
Поскольку АТФ поставляет энергию для сокращения,
Роль АТФ в мышечном расслаблении
Поскольку АТФ поставляет энергию для сокращения,

УТОМЛЕНИЕ МЫШЦ
Утомление - это временное снижение работоспособности мышцы в результате
УТОМЛЕНИЕ МЫШЦ
Утомление - это временное снижение работоспособности мышцы в результате

1.Теория Шиффа: - следствие истощения энергетических запасов в мышце.
2. Теория Пфлюгера:
1.Теория Шиффа: - следствие истощения энергетических запасов в мышце. 2. Теория Пфлюгера:

Особенности расположения сократительных филаментов в гладком миоците
Особенности расположения сократительных филаментов в гладком миоците

Особенности расположения сократительных филаментов в гладком миоците
Нити не организованы в саркомеры.
Тонкие
Особенности расположения сократительных филаментов в гладком миоците
Нити не организованы в саркомеры.
Тонкие

Физиологические
особенности гладких мышц
Менее упорядоченно расположены сократительные белки.
ПП -60/ -70
Физиологические
особенности гладких мышц
Менее упорядоченно расположены сократительные белки.
ПП -60/ -70
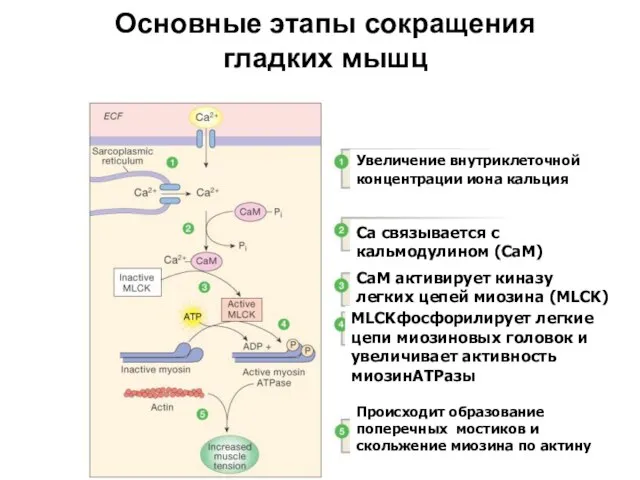
Основные этапы сокращения гладких мышц
Figure 12-28: Smooth muscle contraction
Увеличение внутриклеточной концентрации
Основные этапы сокращения гладких мышц
Figure 12-28: Smooth muscle contraction
Увеличение внутриклеточной концентрации

Мышечная ткань
В организме человека в среднем приходится на долю:
- скелетных
Мышечная ткань
В организме человека в среднем приходится на долю:
- скелетных

Энергетика мышечного сокращения
Энергетика мышечного сокращения

ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
Ресинтез АТФ происходит 2 основными путями:
анаэробным
аэробным
ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
Ресинтез АТФ происходит 2 основными путями:
анаэробным
аэробным

ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
1. Фосфогенная, или АТФ-КФ-система;
2. Лактацидная, или гликолитическая;
3. Кислородная, или
ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
1. Фосфогенная, или АТФ-КФ-система;
2. Лактацидная, или гликолитическая;
3. Кислородная, или
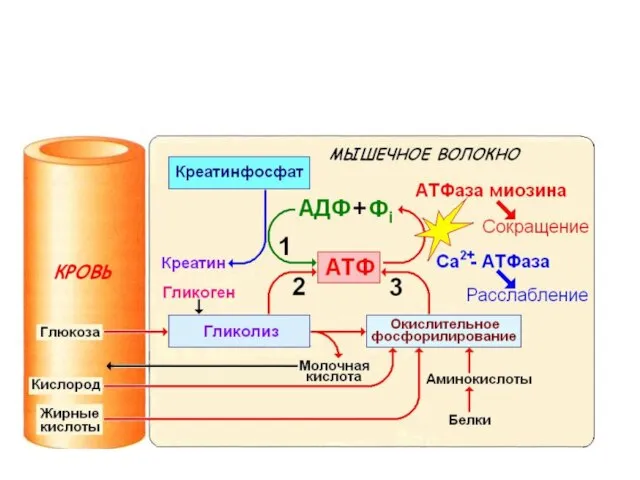
Три системы синтеза и ресинтеза АТФ при мышечном сокращении: 1 –
Три системы синтеза и ресинтеза АТФ при мышечном сокращении: 1 –
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ,
КООРДИНАЦИОННЫЕ ПРИНЦИПЫ в ЦНС
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ,
КООРДИНАЦИОННЫЕ ПРИНЦИПЫ в ЦНС
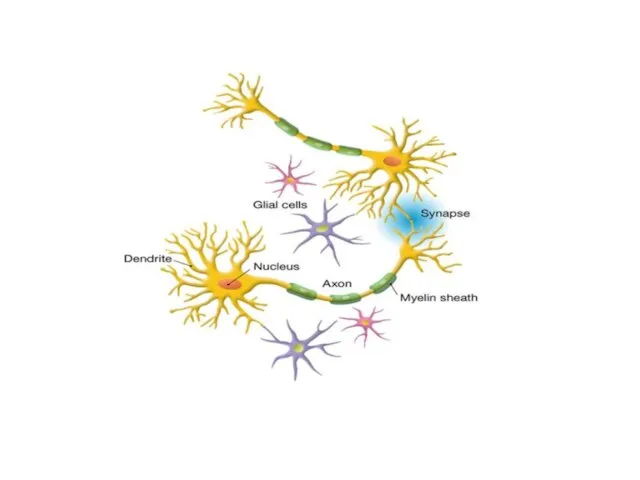

Нейрогли́я, или просто гли́я — совокупность вспомогательных клеток нервной ткани. Составляет около 40 %
Нейрогли́я, или просто гли́я — совокупность вспомогательных клеток нервной ткани. Составляет около 40 %

Клетки нейроглии (астроциты, олигодендроциты, микроглия) заполняют все пространство между нейронами, защищая
Клетки нейроглии (астроциты, олигодендроциты, микроглия) заполняют все пространство между нейронами, защищая

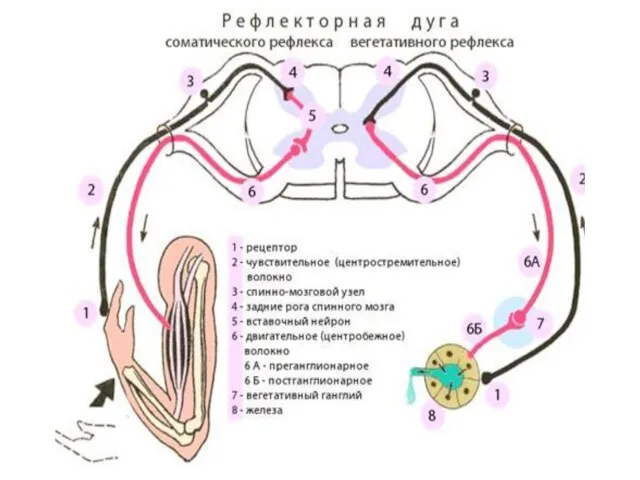
РЕФЛЕКС. РЕФЛЕКТОРНАЯ ДУГА
Рефлекс — это ответная реакция организма на раздражение, происходящая при
РЕФЛЕКС. РЕФЛЕКТОРНАЯ ДУГА
Рефлекс — это ответная реакция организма на раздражение, происходящая при

Классификации рефлексов:
По биологической значимости : пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы.
По
Классификации рефлексов:
По биологической значимости : пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы.
По

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. Одностороннее проведение возбуждения.
2. Центральная задержка рефлекса.
3. Последействие
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. Одностороннее проведение возбуждения.
2. Центральная задержка рефлекса.
3. Последействие

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
одностороннее проведение возбуждения (химические синапсы),
центральная задержка проведения возбуждения (химические
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
одностороннее проведение возбуждения (химические синапсы),
центральная задержка проведения возбуждения (химические

3. Последействие :
кратковременное - связано с особенностью синаптической передачи (медиатор не
3. Последействие :
кратковременное - связано с особенностью синаптической передачи (медиатор не
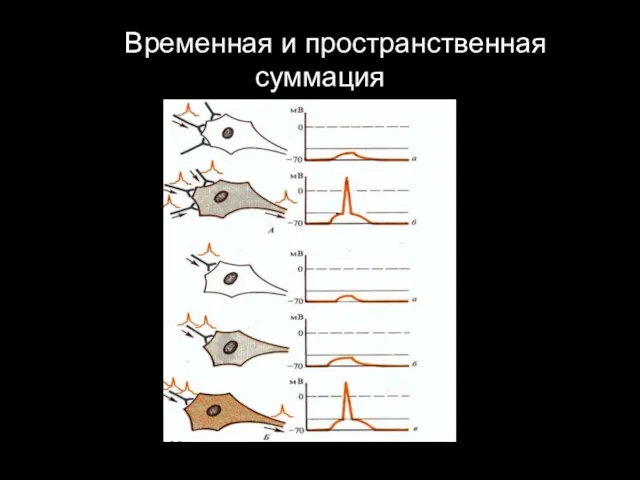
4. Временная и пространственная суммация
4. Временная и пространственная суммация
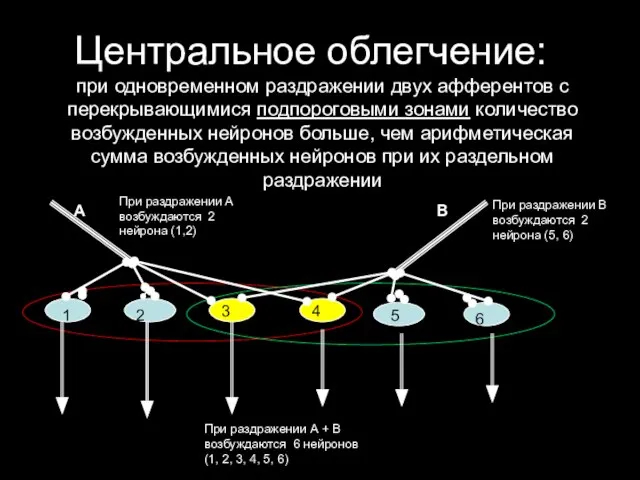
Центральное облегчение:
при одновременном раздражении двух афферентов с перекрывающимися подпороговыми зонами количество
Центральное облегчение: при одновременном раздражении двух афферентов с перекрывающимися подпороговыми зонами количество
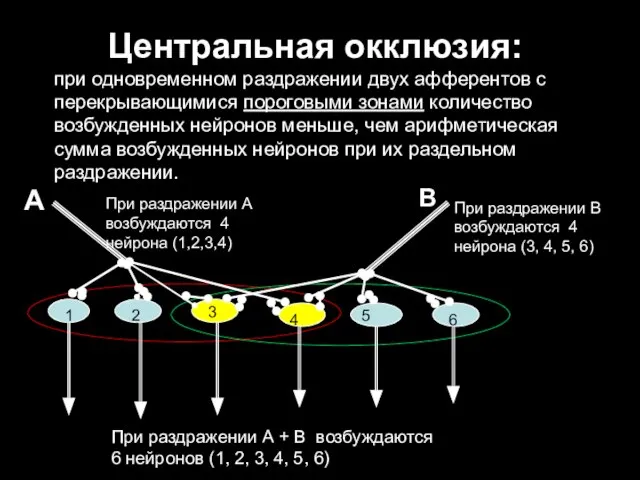
Центральная окклюзия:
при одновременном раздражении двух афферентов с перекрывающимися пороговыми зонами количество
Центральная окклюзия:
при одновременном раздражении двух афферентов с перекрывающимися пороговыми зонами количество
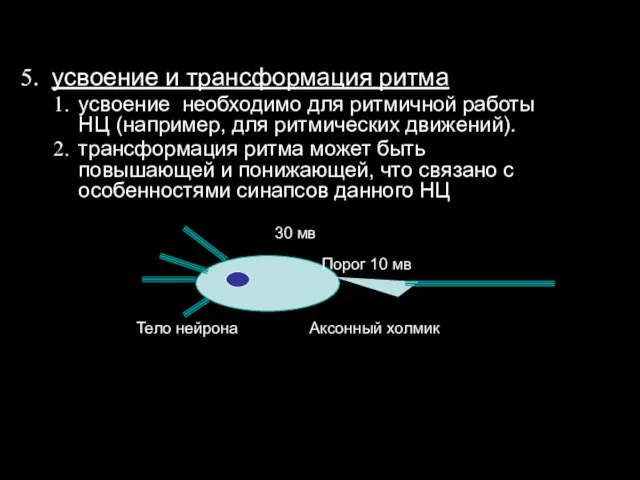
усвоение и трансформация ритма
усвоение необходимо для ритмичной работы НЦ (например,
усвоение и трансформация ритма
усвоение необходимо для ритмичной работы НЦ (например,

Повышение ритма может быть связано с циркуляцией возбуждения в нейронных ловушках;
Снижение
Повышение ритма может быть связано с циркуляцией возбуждения в нейронных ловушках;
Снижение

6. фоновая активность обеспечивается:
поступлением импульсов от соседних НЦ (дивергенция, конвергенция);
6. фоновая активность обеспечивается:
поступлением импульсов от соседних НЦ (дивергенция, конвергенция);

Опыт Бронджеста
Тонус нервного центра
При перерезке у лягушки чувствительных корешков сегментов спинного
Опыт Бронджеста
Тонус нервного центра
При перерезке у лягушки чувствительных корешков сегментов спинного

утомление в НЦ возникает из-за:
истощения медиатора;
истощения АТФ и глюкозы;
снижения чувствительности рецепторов;
метаболических
утомление в НЦ возникает из-за:
истощения медиатора;
истощения АТФ и глюкозы;
снижения чувствительности рецепторов;
метаболических

10. Посттетаническая потенциация
Если , после интенсивного раздражения, дать синапсу отдохнуть и
10. Посттетаническая потенциация
Если , после интенсивного раздражения, дать синапсу отдохнуть и
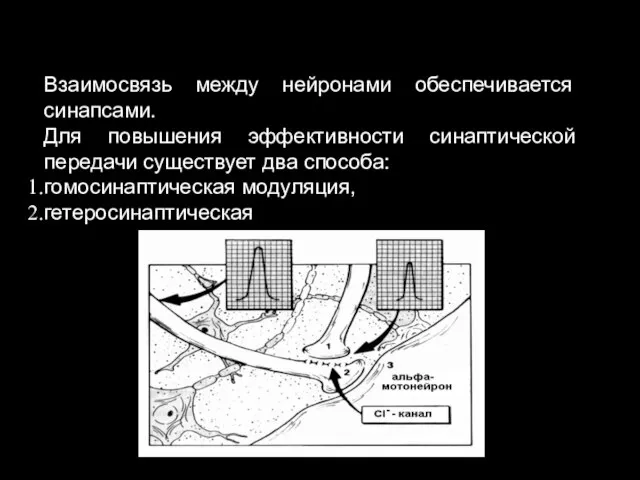
11. Пластичность:
Взаимосвязь между нейронами обеспечивается синапсами.
Для повышения эффективности синаптической передачи существует
11. Пластичность:
Взаимосвязь между нейронами обеспечивается синапсами.
Для повышения эффективности синаптической передачи существует
![Гомосинаптическая модуляция частое (тетаническое) раздражение пресинаптической мембраны ↓ повышение [Са2+ ]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/603023/slide-42.jpg)
Гомосинаптическая
модуляция
частое (тетаническое) раздражение пресинаптической мембраны
↓
повышение [Са2+ ] в
Гомосинаптическая
модуляция
частое (тетаническое) раздражение пресинаптической мембраны
↓
повышение [Са2+ ] в

Гомосинаптическая
модуляция
На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к
Гомосинаптическая
модуляция
На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к
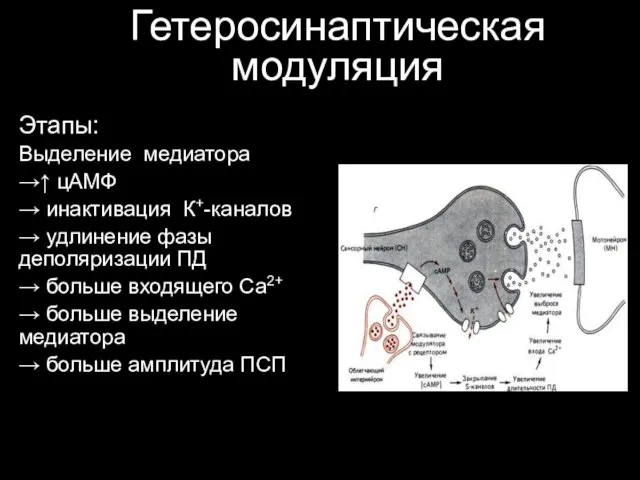
Гетеросинаптическая модуляция
Этапы:
Выделение медиатора
→↑ цАМФ
→ инактивация К+-каналов
→ удлинение фазы деполяризации
Гетеросинаптическая модуляция
Этапы:
Выделение медиатора
→↑ цАМФ
→ инактивация К+-каналов
→ удлинение фазы деполяризации

ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ

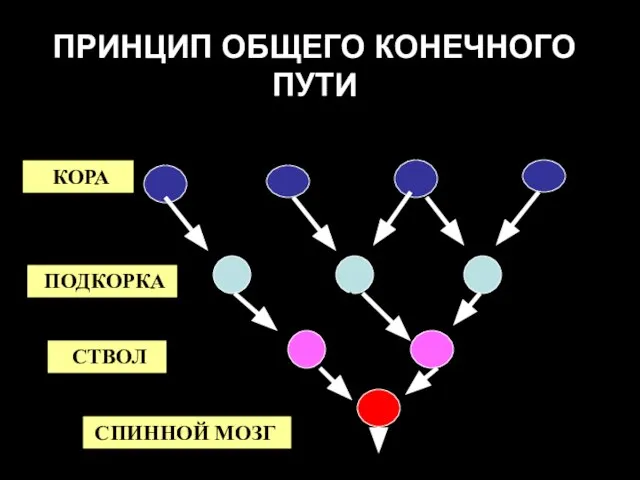
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ

РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
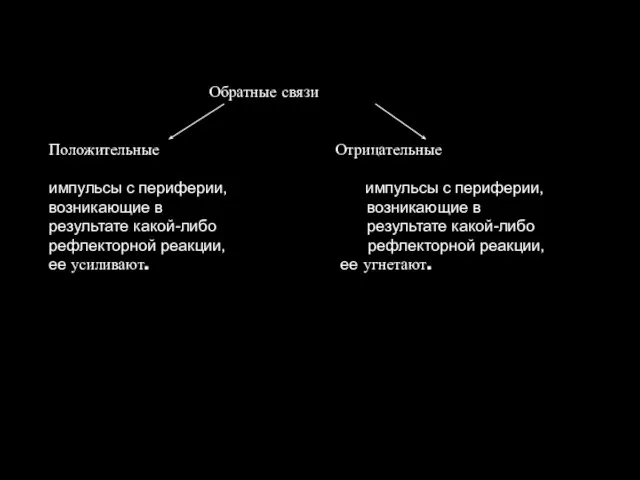
Обратные связи
Положительные Отрицательные
импульсы с периферии, импульсы с периферии,
возникающие в возникающие
Обратные связи
Положительные Отрицательные
импульсы с периферии, импульсы с периферии,
возникающие в возникающие
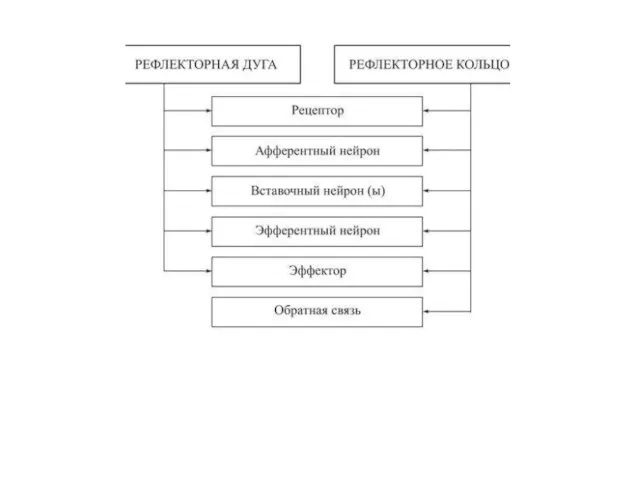
РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
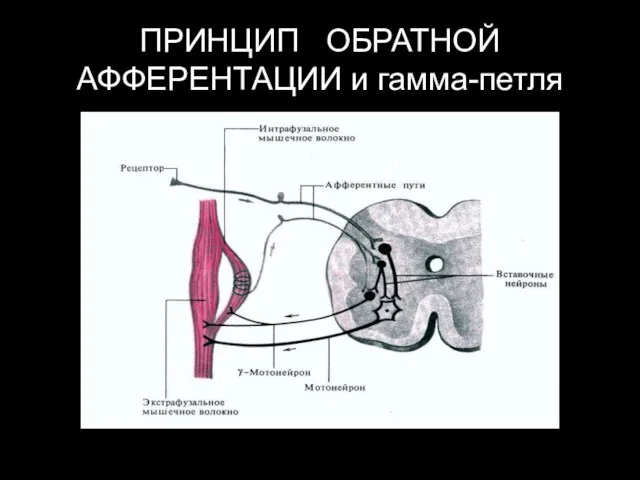
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ и гамма-петля
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ и гамма-петля

ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
Доминанта - временно господствующий рефлекс или поведенческий
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
Доминанта - временно господствующий рефлекс или поведенческий

Основные cвойства доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Способность к
Основные cвойства доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Способность к

Виды доминант
Внутренние доминанты – ведущую роль играет гормональный фон.
Внешние доминанты –
Виды доминант
Внутренние доминанты – ведущую роль играет гормональный фон.
Внешние доминанты –

Причины прекращения существования доминанты
Достижение результата
Появление новой, более значимой доминанты
Запредельное торможение
Причины прекращения существования доминанты
Достижение результата
Появление новой, более значимой доминанты
Запредельное торможение
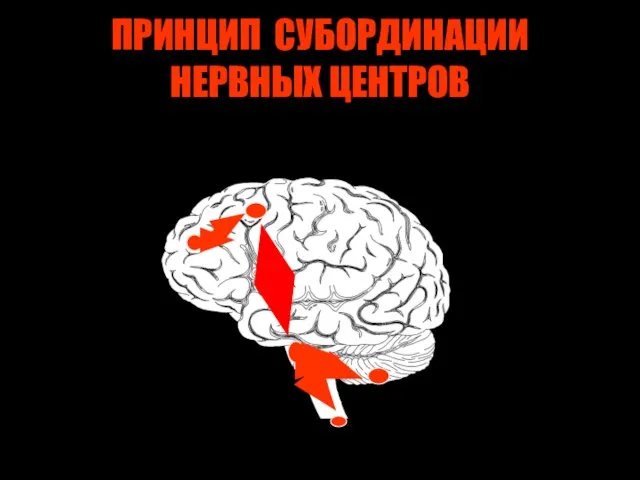
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
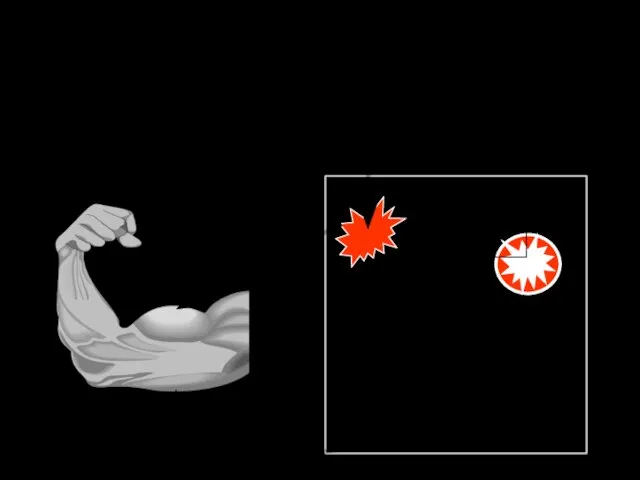
ПРИНЦИП РЕЦИПРОКНОСТИ
+
+
-
-
+
ПРИНЦИП РЕЦИПРОКНОСТИ
+
+
-
-
+

Динамика основных нервных процессов: ИНДУКЦИЯ:
Процессы иррадиации и концентрации составляют основу индукционных
Динамика основных нервных процессов: ИНДУКЦИЯ:
Процессы иррадиации и концентрации составляют основу индукционных
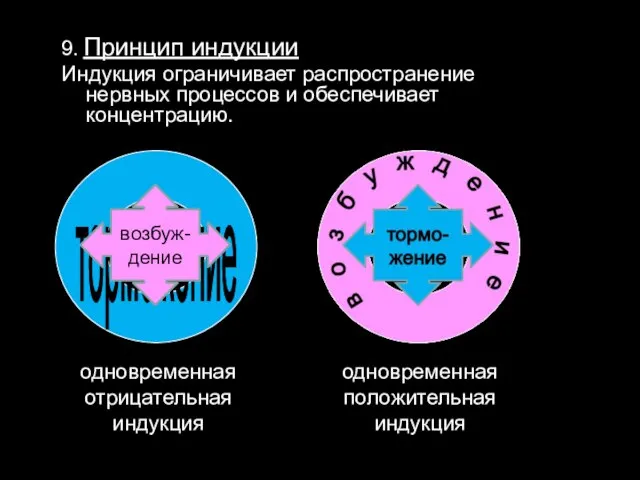
9. Принцип индукции
Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию.
одновременная
отрицательная
9. Принцип индукции
Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию.
одновременная
отрицательная
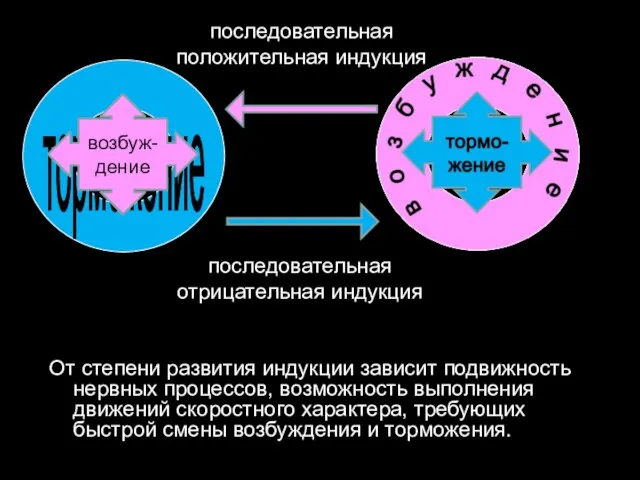
От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений
От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений
 Среды обитания организмов
Среды обитания организмов Китообразные
Китообразные Изъятие и направление трупного материала на лабораторное исследование
Изъятие и направление трупного материала на лабораторное исследование Репрезентативность геоботанических описаний лесных сообществ природного парка Кумысная поляна
Репрезентативность геоботанических описаний лесных сообществ природного парка Кумысная поляна Зоология
Зоология Газообмен в легких. Транспорт газов кровью
Газообмен в легких. Транспорт газов кровью Прыткая ящерица. Отряд чешуйчатые
Прыткая ящерица. Отряд чешуйчатые Приспособленность организмов к условиям внешней среды как результат действия естественного отбора
Приспособленность организмов к условиям внешней среды как результат действия естественного отбора Формирование классической биологии в xix веке
Формирование классической биологии в xix веке Human Body
Human Body § 37. Строение и деятельность внутренних органов 7 класс биология
§ 37. Строение и деятельность внутренних органов 7 класс биология  Презентация на тему Беспозвоночные Презентация к уроку биологии, 5 класс
Презентация на тему Беспозвоночные Презентация к уроку биологии, 5 класс Разнообразен и прекрасен мир растений Мхи, водоросли, папоротники, плауны И семенные: сосны, туя, пихты, ели Но властелин
Разнообразен и прекрасен мир растений Мхи, водоросли, папоротники, плауны И семенные: сосны, туя, пихты, ели Но властелин Презентация на тему "Лист и видоизменения листа" - скачать презентации по Биологии
Презентация на тему "Лист и видоизменения листа" - скачать презентации по Биологии Осень золотая. Лес. Деревья
Осень золотая. Лес. Деревья Вирустардың нәсілдік қасиеттері, олардың өзгергіштігі.мутация түрлері. Гендік инженерия
Вирустардың нәсілдік қасиеттері, олардың өзгергіштігі.мутация түрлері. Гендік инженерия Самые яркие аквариумные рыбки Аквариумные рыбки - популярные домашние питомцы. Наблюдения за поведением рыб успокаивают, расс
Самые яркие аквариумные рыбки Аквариумные рыбки - популярные домашние питомцы. Наблюдения за поведением рыб успокаивают, расс Классификация организмов. Принципы классификации
Классификация организмов. Принципы классификации Определение витамина С в цитрусовых фруктах и их влияние на организм человека
Определение витамина С в цитрусовых фруктах и их влияние на организм человека Чарльз Дарвин о причинах эволюции животного мира
Чарльз Дарвин о причинах эволюции животного мира 01_Vvedenie_Biologia_kak_nauka_Obschaya_kharakteristika_zhizni
01_Vvedenie_Biologia_kak_nauka_Obschaya_kharakteristika_zhizni Тема урока: Обработка клубнеплодов
Тема урока: Обработка клубнеплодов Обмен веществ. Биологическое окисление
Обмен веществ. Биологическое окисление Почка растений (7 класс)
Почка растений (7 класс) Биотические факторы среды. Взаимоотношения между организмами
Биотические факторы среды. Взаимоотношения между организмами Презентация "Алкалоиды" - презентации по Биологии
Презентация "Алкалоиды" - презентации по Биологии Эволюционная роль мутаций, 9 класс
Эволюционная роль мутаций, 9 класс Видовой состав растений горы Большой Башарт
Видовой состав растений горы Большой Башарт