- Выход позвоночных на сушу

Содержание
- 2. ранний девон
- 3. Континентальный климат с резкими колебаниями температуры, ветром, нерегулярными осадками создавал жесткие условия для жизни растений. Многие
- 4. средний девон
- 5. Тело рыб того времени было покрыто костным панцирем и кожное дыхание было невозможно. Вероятно, многие (или
- 6. Целакант = латимерия
- 7. Эустеноптерон Эустеноптерон был подвижной хищной рыбой длиной около 40 см, обитавшей в пресных водоемах. Имел тело
- 8. Пандерихтис У большинства древних рыб имеется узкий канал от свода черепа ко рту (брыз-гальце), опирающийся на
- 9. Пресноводные рыбы приобрели приспособления, позволившие их потомкам, четвероногим позвоночным, перейти к жизни на суше. Грудные плавники
- 10. Акантостега Акантостега - род ископаемых четвероногих, живших в позднем девоне (416-360 млн л н). Считается промежуточным
- 11. Ихтиостега Ихтиостега - род ранних четвероногих из верхнего девона (416-360 млн л н ). Считается представителем
- 12. Тулерпетон Тулерпетон - род ископаемых земноводных из позднего девона(416-360 млн л н), впервые обнаруженный в Тульской
- 13. Эволюционная схема происхождения наземных позвоночных
- 14. Развитие конечности наземного типа От 385 до 350 млн л н от кистепёрых рыб произошли чет-вероногие,
- 15. Стегоцефалы Группа ранних амфибий из позднего девона (416-360 млн л н). Стегоцефалы (панцирного-ловые) имели поверх черепа
- 16. У многих стегоцефплов тело было покрыто рыбообразной чешуёй, у других покровные кости образовывали защитную броню на
- 17. циклотозавр утегения локсомма эриопс протерогиринус эукритта крассигиринус элгинерпетон ихтиостега греерерпетон дипловертеброн хинерпетон сеймурия двинозавр ископаемые стегоцефалы
- 18. Риббо Находки примитивных амфибий, принадлежавших разным видам и особям были сделаны на территории Шотландии. Они относятся
- 19. Педерпес Встречался в раннем карбоне - каменноугольном периоде (около 354-344 млн л н). Он достигал в
- 20. Крассигиринус Жил в раннем карбоне - каменноугольном периоде (354-344 млн л н). Он обитал в болотах
- 21. Эриопс Крупное животное, с длиной черепа до 50 см, общей длиной до 2 метров. Череп массивный,
- 22. Надкласс Четвероногие Tetrapoda Класс Земноводные Amphbia хвостатые безногие бесхвостые
- 23. Изменения локомоции в наземной среде Прямое строение опорных частей грудных плавников не позволяет двигаться вперед, поддерживая
- 24. Скелет земноводных состоит из кости и хряща. У современных представителей, особенно наземных, доля кости выше, чем,
- 25. 1- шейный позвонок 2- туловищный позвонок 3- рёбра 4- крестцовый позвонок 5- хволстовые позвонки 6- уростиль
- 26. Строение конечностей лягушки Передняя 1 — плечевая кость, 2 — головка плеча, 3— предплечье, 4 —
- 27. Мышечная системы амфибий существенно отличается от мышц рыб. Часть туловищной мускулатуры сохраняет метамерное рас-положение, особенно заметно
- 28. 1 — лопатка, 2 — надлопаточный хрящ, 3 — коракоид (воронья кость) 4 — суставная впадина
- 29. 1 — подвздошная кость, 2 — седалищная кость, 3— лобковый хрящ, 4 — вертлужная впадина, Тазовый
- 30. 1 — боковая затылочная кость, 2 — затылочный мыщелок, 3 — переднеушная кость, 4 — клиновидно-обонятельная
- 31. Для большинства рыб характерна гиостилия (вариант, при котором с мозговым черепом связана только гиоидная дуга, а
- 32. 1 — правое предсердие, 2 — левое предсердие, 3 — желудочек, 4 — клапаны, закрывающие общее
- 33. Сосудистая система земноводных
- 34. Внутреннее строение лягушки
- 35. Механизм дыхания земноводных В дыхательную систему входят и проводящие пути и лёгкие. Наружные ноздри ведут в
- 36. 1 — почка, 2 — мочеточник (он же семяпровод), 3 — полость клоаки, 4 — мочеполовое
- 37. 1 — почка, 2 — мочеточник, 3 — полость клоаки, 4 — мочевое отверстие, 5 —
- 38. Водно-солевой обмен у земноводных БОльшая часть видов - наземная, часть - водная. Наземные выделяют мочевину -
- 39. при недостатке влаги в почках - растёт реабсорбция воды при повышении солёности в почках - ослабевает
- 40. Размножение и развитие амфибий Строение половой системы сходно с другими первичноводными: у самцов вольфов канал служит
- 41. Вид кладок амфибий. А - лягушка, Б- квакша, В- чесночница, Г- жаба, Д- жерлянка, Е- жаба-повитуха
- 42. Развитие яйца лягушки 1- бластоцель, 2- желточная пробка, 3- эктодерма, 4- медулярная бороздка, 5- энтодерма, 6-
- 43. Развитие остромордой лягушки 1- икра, 2- вылупившаяся личинка, 3 и 4- рост наружных жабр и плавниковой
- 45. Скачать презентацию

ранний девон
ранний девон

Континентальный климат с
резкими колебаниями температуры,
ветром,
нерегулярными осадками
создавал жесткие
Континентальный климат с
резкими колебаниями температуры,
ветром,
нерегулярными осадками
создавал жесткие

средний девон
средний девон

Тело рыб того времени было покрыто костным панцирем и кожное дыхание
Тело рыб того времени было покрыто костным панцирем и кожное дыхание

Целакант = латимерия
Целакант = латимерия

Эустеноптерон
Эустеноптерон был подвижной хищной рыбой длиной около 40 см, обитавшей в
Эустеноптерон
Эустеноптерон был подвижной хищной рыбой длиной около 40 см, обитавшей в

Пандерихтис
У большинства древних рыб имеется узкий канал от свода черепа ко
Пандерихтис
У большинства древних рыб имеется узкий канал от свода черепа ко
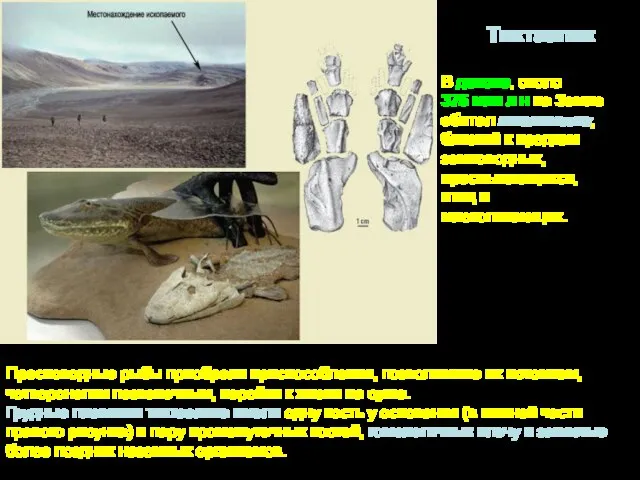
Пресноводные рыбы приобрели приспособления, позволившие их потомкам, четвероногим позвоночным, перейти к
Пресноводные рыбы приобрели приспособления, позволившие их потомкам, четвероногим позвоночным, перейти к

Акантостега
Акантостега - род ископаемых четвероногих, живших в позднем девоне (416-360 млн
Акантостега
Акантостега - род ископаемых четвероногих, живших в позднем девоне (416-360 млн

Ихтиостега
Ихтиостега - род ранних четвероногих из верхнего девона (416-360 млн л н
Ихтиостега
Ихтиостега - род ранних четвероногих из верхнего девона (416-360 млн л н

Тулерпетон
Тулерпетон - род ископаемых земноводных из позднего девона(416-360 млн л н), впервые
Тулерпетон
Тулерпетон - род ископаемых земноводных из позднего девона(416-360 млн л н), впервые

Эволюционная схема происхождения наземных
позвоночных
Эволюционная схема происхождения наземных
позвоночных

Развитие конечности наземного типа
От 385 до 350 млн л н от
Развитие конечности наземного типа
От 385 до 350 млн л н от

Стегоцефалы
Группа ранних амфибий из позднего девона (416-360 млн л н).
Стегоцефалы
Стегоцефалы
Группа ранних амфибий из позднего девона (416-360 млн л н).
Стегоцефалы

У многих стегоцефплов тело было покрыто рыбообразной чешуёй, у других покровные
У многих стегоцефплов тело было покрыто рыбообразной чешуёй, у других покровные

циклотозавр
утегения
локсомма
эриопс
протерогиринус
эукритта
крассигиринус
элгинерпетон
ихтиостега
греерерпетон
дипловертеброн
хинерпетон
сеймурия
двинозавр
ископаемые стегоцефалы
циклотозавр
утегения
локсомма
эриопс
протерогиринус
эукритта
крассигиринус
элгинерпетон
ихтиостега
греерерпетон
дипловертеброн
хинерпетон
сеймурия
двинозавр
ископаемые стегоцефалы

Риббо
Находки примитивных амфибий, принадлежавших разным видам и особям были сделаны на
Риббо
Находки примитивных амфибий, принадлежавших разным видам и особям были сделаны на

Педерпес
Встречался в раннем карбоне - каменноугольном периоде (около 354-344 млн л
Педерпес
Встречался в раннем карбоне - каменноугольном периоде (около 354-344 млн л

Крассигиринус
Жил в раннем карбоне - каменноугольном периоде (354-344 млн л н).
Он
Крассигиринус
Жил в раннем карбоне - каменноугольном периоде (354-344 млн л н).
Он

Эриопс
Крупное животное, с длиной черепа до 50 см, общей длиной
Эриопс
Крупное животное, с длиной черепа до 50 см, общей длиной

Надкласс Четвероногие Tetrapoda
Класс Земноводные Amphbia
хвостатые
безногие
бесхвостые
Надкласс Четвероногие Tetrapoda
Класс Земноводные Amphbia
хвостатые
безногие
бесхвостые
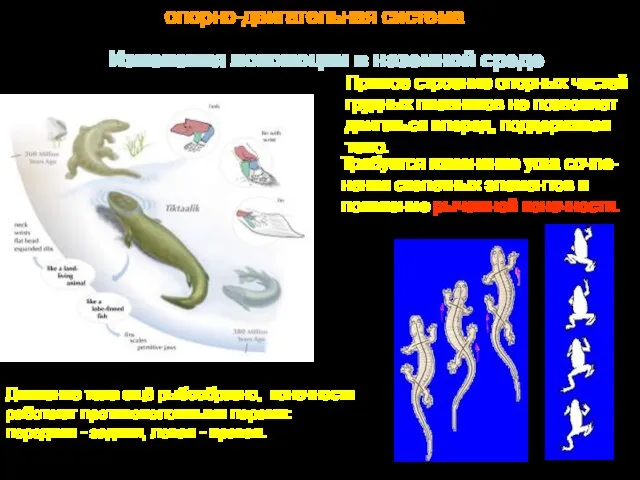
Изменения локомоции в наземной среде
Прямое строение опорных частей
грудных плавников не
Изменения локомоции в наземной среде
Прямое строение опорных частей
грудных плавников не

Скелет земноводных состоит из кости и хряща.
У современных представителей, особенно наземных,
Скелет земноводных состоит из кости и хряща.
У современных представителей, особенно наземных,
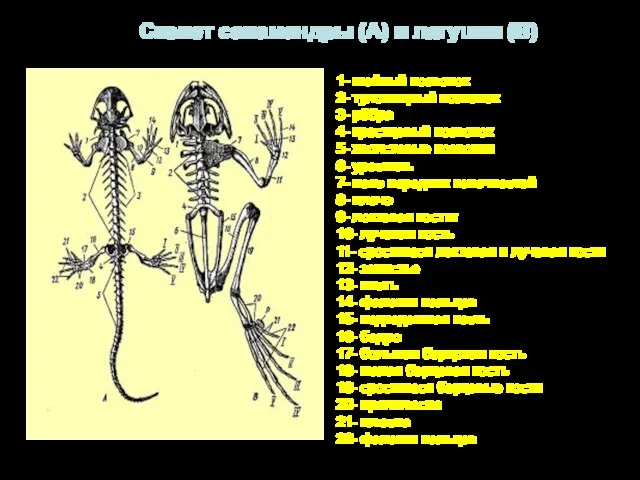
1- шейный позвонок
2- туловищный позвонок
3- рёбра
4- крестцовый позвонок
5- хволстовые позвонки
6- уростиль
7-
1- шейный позвонок
2- туловищный позвонок
3- рёбра
4- крестцовый позвонок
5- хволстовые позвонки
6- уростиль
7-

Строение конечностей лягушки
Передняя
1 — плечевая кость,
2 — головка плеча,
Строение конечностей лягушки
Передняя
1 — плечевая кость,
2 — головка плеча,

Мышечная системы амфибий существенно отличается от мышц рыб.
Часть туловищной мускулатуры
Мышечная системы амфибий существенно отличается от мышц рыб.
Часть туловищной мускулатуры

1 — лопатка,
2 — надлопаточный хрящ,
3 — коракоид (воронья
1 — лопатка,
2 — надлопаточный хрящ,
3 — коракоид (воронья
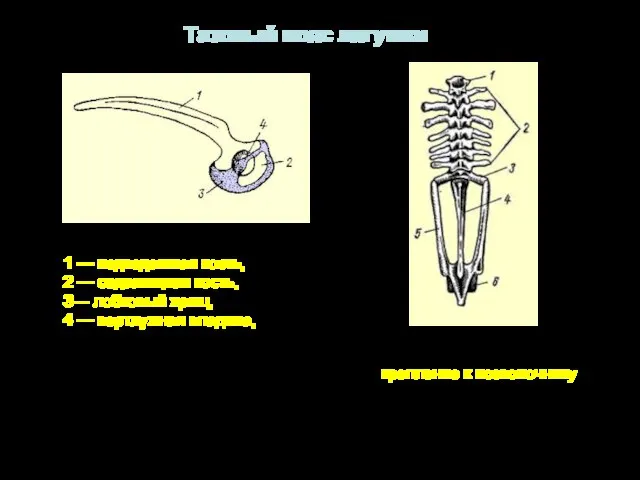
1 — подвздошная кость,
2 — седалищная кость,
3— лобковый хрящ,
4
1 — подвздошная кость,
2 — седалищная кость,
3— лобковый хрящ,
4

1 — боковая затылочная кость, 2 — затылочный мыщелок,
3 —
1 — боковая затылочная кость, 2 — затылочный мыщелок,
3 —

Для большинства рыб характерна гиостилия (вариант, при котором с мозговым черепом
Для большинства рыб характерна гиостилия (вариант, при котором с мозговым черепом

1 — правое предсердие,
2 — левое предсердие,
3 — желудочек,
1 — правое предсердие,
2 — левое предсердие,
3 — желудочек,

Сосудистая система земноводных
Сосудистая система земноводных

Внутреннее строение лягушки
Внутреннее строение лягушки

Механизм дыхания земноводных
В дыхательную систему входят и проводящие пути и лёгкие.
Механизм дыхания земноводных
В дыхательную систему входят и проводящие пути и лёгкие.

1 — почка,
2 — мочеточник (он же семяпровод),
3 —
1 — почка,
2 — мочеточник (он же семяпровод),
3 —

1 — почка,
2 — мочеточник,
3 — полость клоаки,
4
1 — почка,
2 — мочеточник,
3 — полость клоаки,
4

Водно-солевой обмен у земноводных
БОльшая часть видов - наземная, часть - водная.
Наземные
Водно-солевой обмен у земноводных
БОльшая часть видов - наземная, часть - водная.
Наземные

при недостатке влаги
в почках
- растёт реабсорбция воды
при повышении солёности
в почках
при недостатке влаги
в почках
- растёт реабсорбция воды
при повышении солёности
в почках
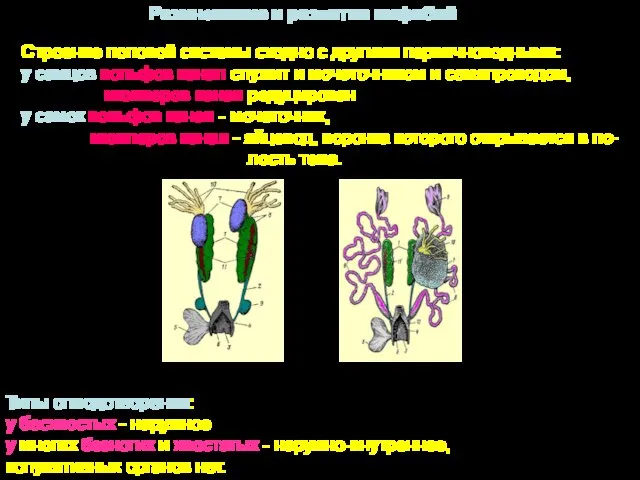
Размножение и развитие амфибий
Строение половой системы сходно с другими первичноводными:
у самцов
Размножение и развитие амфибий
Строение половой системы сходно с другими первичноводными:
у самцов
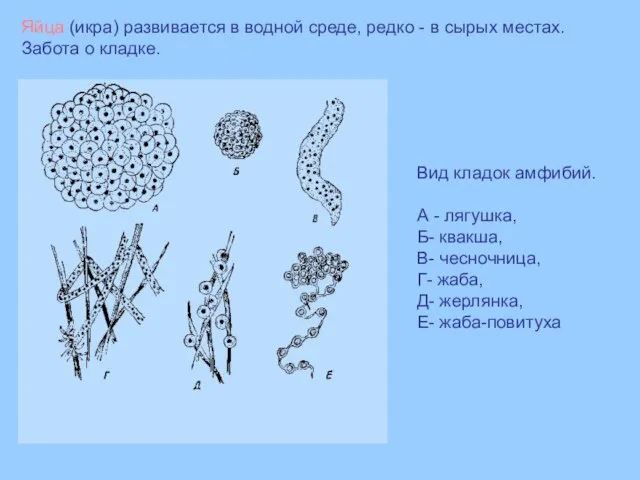
Вид кладок амфибий.
А - лягушка,
Б- квакша,
В- чесночница,
Г- жаба,
Д- жерлянка,
Е- жаба-повитуха
Яйца (икра)
Вид кладок амфибий.
А - лягушка,
Б- квакша,
В- чесночница,
Г- жаба,
Д- жерлянка,
Е- жаба-повитуха
Яйца (икра)

Развитие яйца лягушки
1- бластоцель,
2- желточная пробка,
3- эктодерма,
4- медулярная бороздка,
5- энтодерма,
6- мезодерма,
7-
Развитие яйца лягушки
1- бластоцель,
2- желточная пробка,
3- эктодерма,
4- медулярная бороздка,
5- энтодерма,
6- мезодерма,
7-

Развитие остромордой лягушки
1- икра,
2- вылупившаяся личинка,
3 и 4- рост наружных жабр
Развитие остромордой лягушки
1- икра,
2- вылупившаяся личинка,
3 и 4- рост наружных жабр
 Преподаватель: Погребнёв Сергей Сергеевич Лекция № 3. ОРГАНИЧЕСКИЕ ВЕЩЕСТВА. БЕЛКИ.
Преподаватель: Погребнёв Сергей Сергеевич Лекция № 3. ОРГАНИЧЕСКИЕ ВЕЩЕСТВА. БЕЛКИ. Типы клеток
Типы клеток «Влияние загрязнения окружающей среды на половой состав новорожденных.» Исследовательская работа
«Влияние загрязнения окружающей среды на половой состав новорожденных.» Исследовательская работа Balstaudi. Elastīgais skrimslis
Balstaudi. Elastīgais skrimslis Жизненные формы водорослей
Жизненные формы водорослей Грибы - паразиты (5 класс)
Грибы - паразиты (5 класс) Учитель химии и биологии МБОУ СОШ № 9 МО ЩР станица Новощербиновская Степучева Ольга Викторовна
Учитель химии и биологии МБОУ СОШ № 9 МО ЩР станица Новощербиновская Степучева Ольга Викторовна Сущность жизни и свойства живого. Уровни организации живой материи
Сущность жизни и свойства живого. Уровни организации живой материи Методы биологического познания
Методы биологического познания Кровь и остальные компоненты внутренней среды организма. Автор учитель биологии МОУ «Гимназия № 23» г. Троицк Че
Кровь и остальные компоненты внутренней среды организма. Автор учитель биологии МОУ «Гимназия № 23» г. Троицк Че Вегетативные органы растений
Вегетативные органы растений Метаболизм липидов
Метаболизм липидов АТЕРОСКЛЕРОЗ БОЛЕЗНЬ ЦИВИЛИЗАЦИИ Шидьюсова Анастасия 243 группа
АТЕРОСКЛЕРОЗ БОЛЕЗНЬ ЦИВИЛИЗАЦИИ Шидьюсова Анастасия 243 группа  Плодовые культуры. Земляника садовая
Плодовые культуры. Земляника садовая Кожа. Строение, значение и гигиена кожи
Кожа. Строение, значение и гигиена кожи Молюски
Молюски Общая характеристика Типа Круглые или Первичнополостные черви (Nemathelminthes) .
Общая характеристика Типа Круглые или Первичнополостные черви (Nemathelminthes) .  Головной мозг
Головной мозг Презентация на тему "В мире инфекций" - скачать презентации по Биологии
Презентация на тему "В мире инфекций" - скачать презентации по Биологии Заповедники Донского края. Красная книга Ростовской области
Заповедники Донского края. Красная книга Ростовской области Постэмбриональное развитие животных
Постэмбриональное развитие животных Ч.Дарвиннің Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары, эволюцияның синтетикалық теориясы
Ч.Дарвиннің Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары, эволюцияның синтетикалық теориясы Биотехнология
Биотехнология Биологический механизм запахов
Биологический механизм запахов Разнообразие круглых червей
Разнообразие круглых червей Презентация на тему Бионика
Презентация на тему Бионика  Биоритмы - внутренние часы человека Работу выполнили: ученицы 9 класса «Б» Городничева Кристина Алимова Анжелика Климова Регина.
Биоритмы - внутренние часы человека Работу выполнили: ученицы 9 класса «Б» Городничева Кристина Алимова Анжелика Климова Регина. Анатомия ЦНС и физиология ВНД
Анатомия ЦНС и физиология ВНД