- Начальные этапы обмена липидов

Содержание
- 2. Липиды – это гетерогенная группа соединений, непосредственно или опосредованно связанных с жирными кислотами. Их общим свойством
- 3. Биомедицинское значение Энергетическая (1г – 39 кДж). Теплоизоляционная. Электроизоляционная. Защитная. Структурная. Всасывание витаминов. Регуляторная. Запас эндогенной
- 5. Липиды по физиологическому значению Резервные – запасаются в больших количествах, а затем расходуются для энергетических нужд
- 6. Жирные кислоты
- 9. Цис – транс- изомерия ненасыщенных жирных кислот
- 10. Ансель Кис (1904-2004)
- 11. Эйкозаноиды Образуются из эйкоза –(С-20) – полиеновых жирных кислот.
- 13. Простагландины
- 14. Простагландины стимулируют Сокращение гладкой мускулатуры. Липолиз. Снижение секреции желудочного сока. Систему свертывания крови. Расширение кровеносных сосудов.
- 15. Триацилглицеролы (нейтральные жиры) Сложные эфиры глицерина и высших жирных кислот. Нейтральные жиры бывают: Твердые – содержат
- 17. Фосфолипиды - это группа липидов, содержащих в своем составе остаток фосфорной кислоты. Глицерофосфолипиды – в их
- 18. Функциональная роль фосфолипидов Являются компонентами клеточных мембран. Выполняют детергентную функцию в кишечнике и желчном пузыре. Источник
- 20. Стероиды
- 21. ПЕРЕВАРИВАНИЕ, ВСАСЫВАНИЕ И ТРАНСПОРТ ЛИПИДОВ.
- 22. С пищей ежедневно поступает от 80 – 150 гр липидов. Основная часть – нейтральные жиры. Переваривание
- 23. Эмульгация жира. Эмульгирование заключается в дроблении крупных липидных частиц на более мелкие. Происходит этот процесс благодаря
- 24. Желчные кислоты Желчные кислоты образуются в печени из эфиров холестерола Образующиеся желчные кислоты поступают из печени
- 27. Таблица Состав желчи человека Липиды и желчные кислоты мол% Холестерол 8* Фосфолипиды 23 Желчные кислоты и
- 28. Рециркуляция желчных кислот между печенью и кишечником. За сутки из печени выделяется 15–30 г желчных кислот
- 30. Переваривание жиров Основным ферментом, который катализирует расщепление липидов пищи, является панкреатическая липаза. Этот фермент присутствует в
- 31. В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды. Эстеразы катализируют преимущественно гидролиз эфиров
- 33. Продукты гидролиза липидов участвуют в образовании мицелл Продукты ферментативного гидролиза жира взаимодействуют с водной средой, объединяются
- 35. Ресинтез липидов. В клетках имеются ферментные системы, которые могут превращать СЖК, моно- и диацилглицеролы в ТАГ,
- 36. Транспорт липидов из кишечника Новосинтезированные ТАГ, фосфолипиды и другие всосавшиеся липиды покидают клетки слизистой, попадая сначала
- 39. Нарушение всасывания липидов. Стеаторея. Для обеспечения нормальной функции кишечника в отношении всасывания жиров определяющее значение имеет
- 40. Нарушение любого фактора приводит к стеаторее – выделению непереваренных липидов с калом, который приобретает серовато-белый цвет.
- 41. Формы стеаторей. Панкреатическая стеаторея (дефицит панкреатической липазы) – наблюдается при хроническом панкреатите, гипоплазии клеток и т.д.
- 42. ТКАНЕВЫЙ ЛИПОЛИЗ ТАГ.
- 43. Гидролиз ТАГ Осуществляется липазами. Активность липазы в клетках жировой ткани находится под строгим регуляторным контролем (отсюда
- 46. В результате полного гидролиза молекулы ТАГ образуется три молекулы жирных кислот и одна молекула глицерола. Последний
- 47. ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
- 49. Транспорт активированных жирных кислот в митохондрии Ацил-КоА не может проникнуть в митохондрию без помощи карнитина, который
- 51. Структура карнитина
- 52. Современная теория β-окисления Локализация – матрикс митохондрий Генерирование энергии в виде: Ацетил-коА НАДН·Н + ФАД Н
- 55. Большой вклад в изучение данной последовательности реакций внесли Д. Грин, С. Очоа и Ф. Линен. Они
- 57. Энергетический выход β-окисления жирных кислот зависит от длины углеводородной цепи. Расчет в более общей форме для
- 58. Суммарное уравнение окисления пальмитиновой кислоты. Пальмитоил S-коА +7КоАSН + 7ФАД +7НАД + +7Н 2 О =
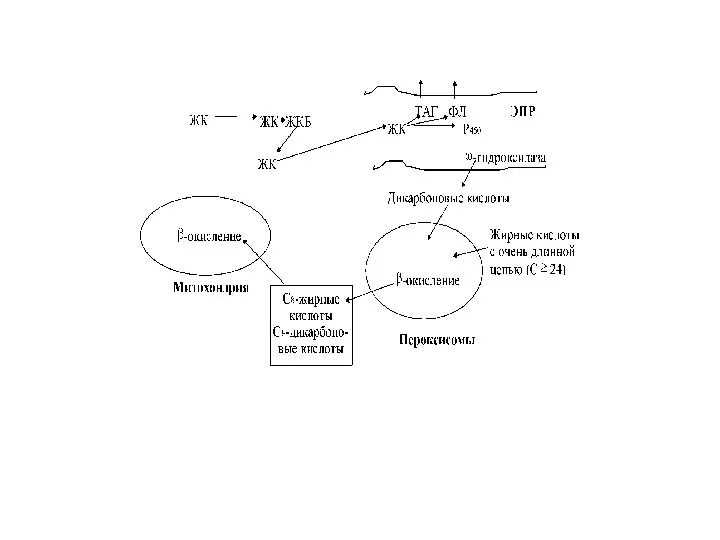
- 60. Окисление жирных кислот в пероксисомах Жирные кислоты с очень длинной цепью. Особенностью метаболизма жирных кислот в
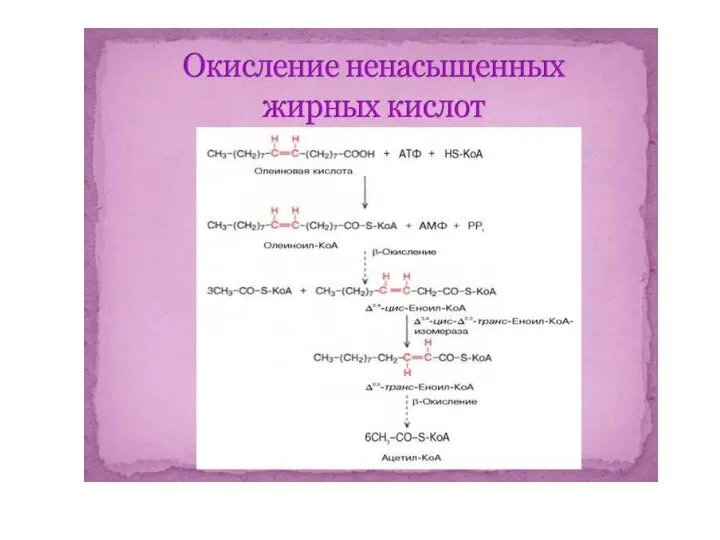
- 62. β-окисление ненасыщенных жирных кислот β-окисление ненасыщенных жирных кислот во многом подобно окислению насыщенных жирных кислот. В
- 64. β-Окисление жирных кислот с нечетным числом углеродных атомов Жирные кислоты с нечетным числом углеродных атомов в
- 67. Пропионат также образуется при расщеплении четырех аминокислот: валина, изолейцина, метионина и треонина, а также из боковой
- 68. Ключевой процесс – поступление жирных кислот в митохондрию. Карнитинацилтрансфераза – аллостерический фермент, его ингибитором выступает малонилкоА.
- 70. Скачать презентацию

Липиды – это гетерогенная группа соединений, непосредственно или опосредованно связанных с
Липиды – это гетерогенная группа соединений, непосредственно или опосредованно связанных с

Биомедицинское значение
Энергетическая (1г – 39 кДж).
Теплоизоляционная.
Электроизоляционная.
Защитная.
Структурная.
Всасывание витаминов.
Регуляторная.
Запас эндогенной воды.
Биомедицинское значение
Энергетическая (1г – 39 кДж).
Теплоизоляционная.
Электроизоляционная.
Защитная.
Структурная.
Всасывание витаминов.
Регуляторная.
Запас эндогенной воды.

Липиды по физиологическому значению
Резервные – запасаются в больших количествах, а
Липиды по физиологическому значению
Резервные – запасаются в больших количествах, а

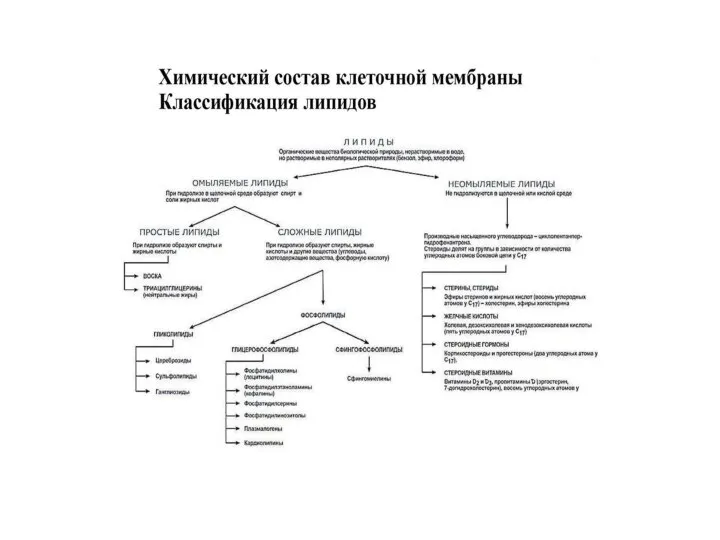
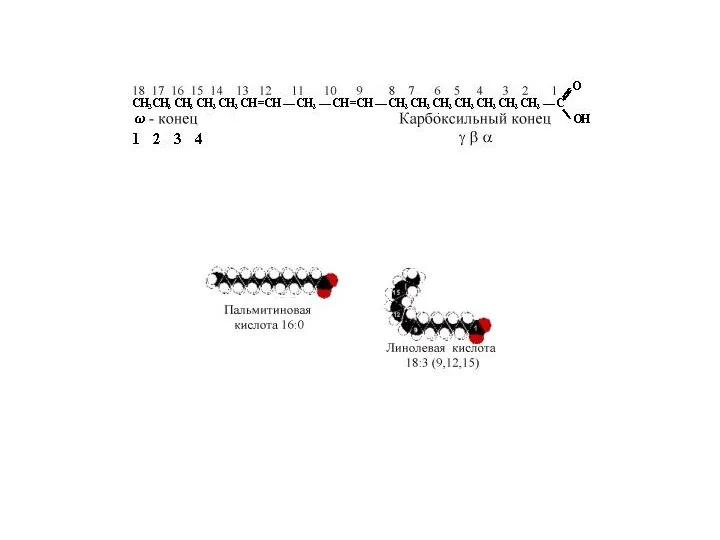
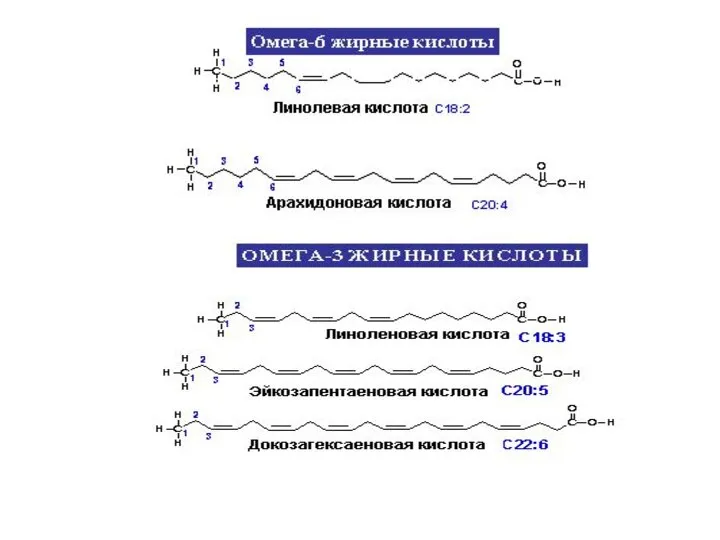
Жирные кислоты
Жирные кислоты
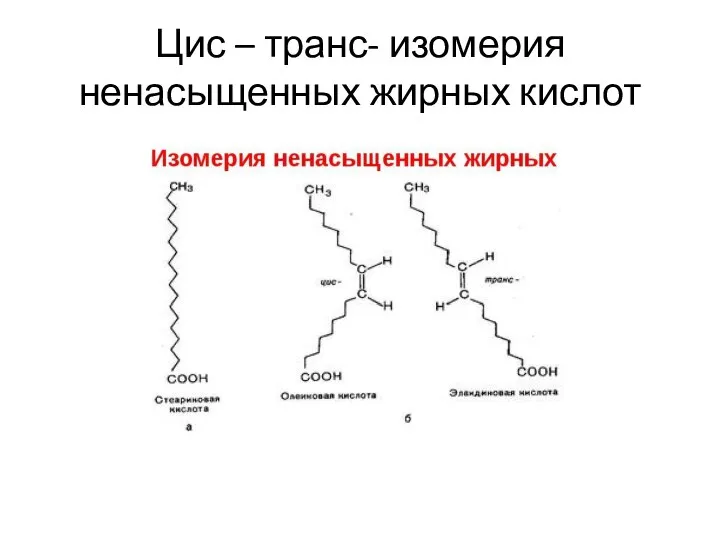
Цис – транс- изомерия ненасыщенных жирных кислот
Цис – транс- изомерия ненасыщенных жирных кислот

Ансель Кис (1904-2004)
Ансель Кис (1904-2004)

Эйкозаноиды
Образуются из эйкоза –(С-20) – полиеновых жирных кислот.
Эйкозаноиды
Образуются из эйкоза –(С-20) – полиеновых жирных кислот.

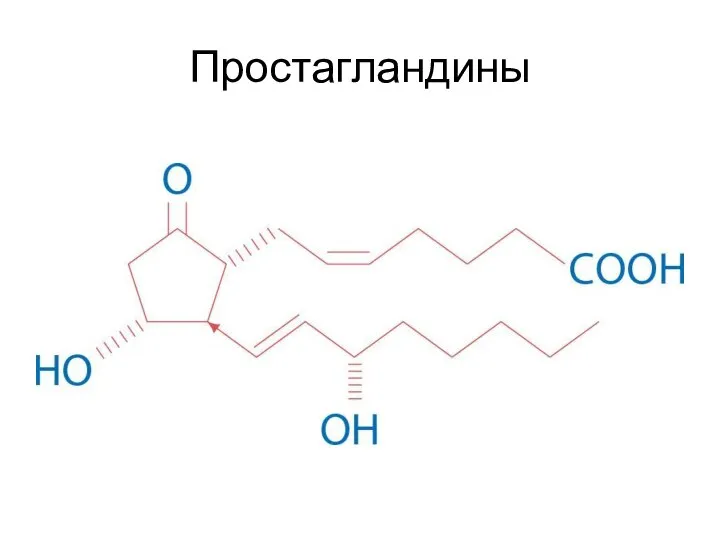
Простагландины
Простагландины

Простагландины стимулируют
Сокращение гладкой мускулатуры.
Липолиз.
Снижение секреции желудочного сока.
Систему свертывания крови.
Расширение кровеносных сосудов.
Активность
Простагландины стимулируют
Сокращение гладкой мускулатуры.
Липолиз.
Снижение секреции желудочного сока.
Систему свертывания крови.
Расширение кровеносных сосудов.
Активность

Триацилглицеролы (нейтральные жиры)
Сложные эфиры глицерина и высших жирных кислот.
Нейтральные
Триацилглицеролы (нейтральные жиры)
Сложные эфиры глицерина и высших жирных кислот.
Нейтральные


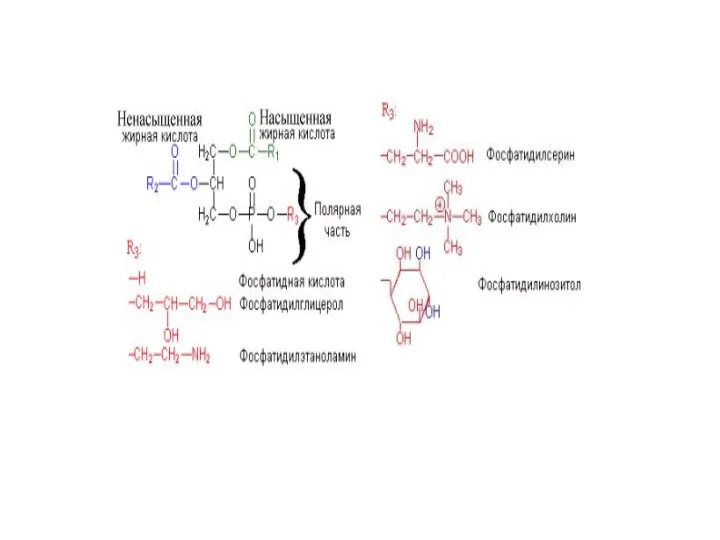
Фосфолипиды
- это группа липидов, содержащих в своем составе остаток фосфорной
Фосфолипиды
- это группа липидов, содержащих в своем составе остаток фосфорной

Функциональная роль фосфолипидов
Являются компонентами клеточных мембран.
Выполняют детергентную функцию в кишечнике и
Функциональная роль фосфолипидов
Являются компонентами клеточных мембран.
Выполняют детергентную функцию в кишечнике и

Стероиды
Стероиды

ПЕРЕВАРИВАНИЕ, ВСАСЫВАНИЕ И ТРАНСПОРТ ЛИПИДОВ.
ПЕРЕВАРИВАНИЕ, ВСАСЫВАНИЕ И ТРАНСПОРТ ЛИПИДОВ.

С пищей ежедневно поступает от 80 – 150 гр липидов.
Основная
С пищей ежедневно поступает от 80 – 150 гр липидов.
Основная

Эмульгация жира.
Эмульгирование заключается в дроблении крупных липидных частиц на более мелкие.
Эмульгация жира.
Эмульгирование заключается в дроблении крупных липидных частиц на более мелкие.

Желчные кислоты
Желчные кислоты образуются в печени из эфиров холестерола
Образующиеся желчные
Желчные кислоты
Желчные кислоты образуются в печени из эфиров холестерола
Образующиеся желчные
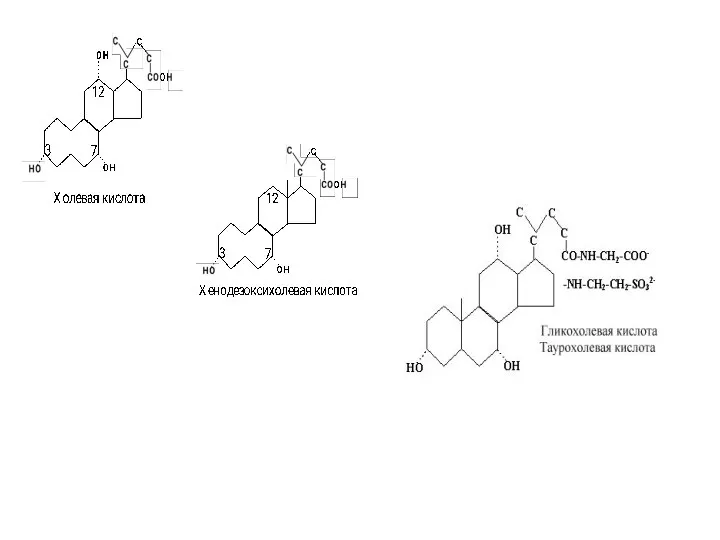


Таблица Состав желчи человека
Липиды и желчные кислоты мол%
Холестерол 8*
Фосфолипиды 23
Желчные
Таблица Состав желчи человека
Липиды и желчные кислоты мол%
Холестерол 8*
Фосфолипиды 23
Желчные

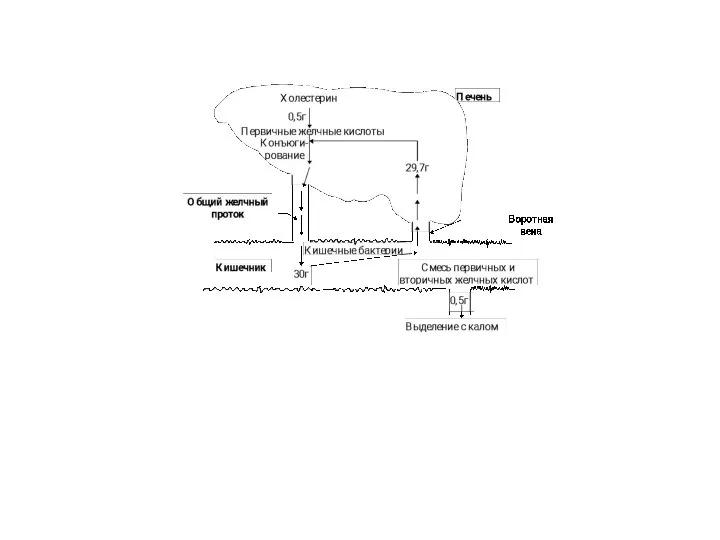
Рециркуляция желчных кислот между печенью и кишечником.
За сутки из печени выделяется
Рециркуляция желчных кислот между печенью и кишечником.
За сутки из печени выделяется

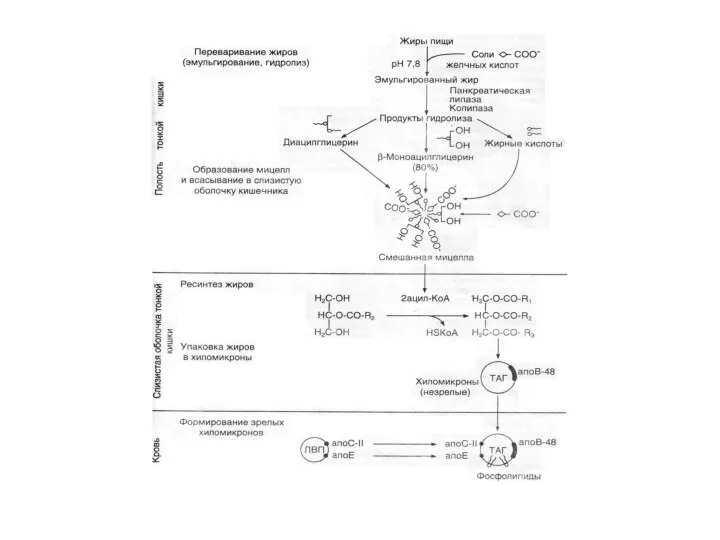
Переваривание жиров
Основным ферментом, который катализирует расщепление липидов пищи, является панкреатическая
Переваривание жиров
Основным ферментом, который катализирует расщепление липидов пищи, является панкреатическая

В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды.
В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды.
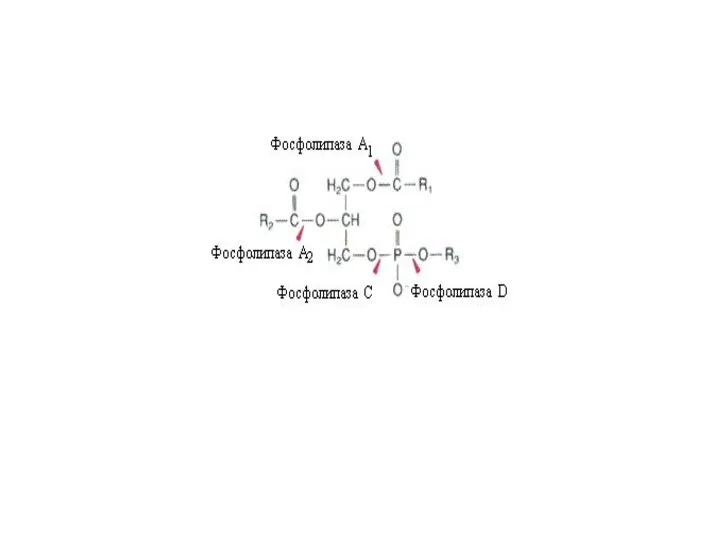

Продукты гидролиза липидов участвуют в образовании мицелл
Продукты ферментативного гидролиза жира
Продукты гидролиза липидов участвуют в образовании мицелл
Продукты ферментативного гидролиза жира


Ресинтез липидов.
В клетках имеются ферментные системы, которые могут превращать СЖК, моно-
Ресинтез липидов.
В клетках имеются ферментные системы, которые могут превращать СЖК, моно-

Транспорт липидов из кишечника
Новосинтезированные ТАГ, фосфолипиды и другие всосавшиеся липиды
Транспорт липидов из кишечника
Новосинтезированные ТАГ, фосфолипиды и другие всосавшиеся липиды


Нарушение всасывания липидов.
Стеаторея.
Для обеспечения нормальной функции кишечника в отношении всасывания жиров
Нарушение всасывания липидов.
Стеаторея.
Для обеспечения нормальной функции кишечника в отношении всасывания жиров

Нарушение любого фактора приводит к стеаторее – выделению непереваренных липидов с

Формы стеаторей.
Панкреатическая стеаторея (дефицит панкреатической липазы) – наблюдается при хроническом панкреатите,
Формы стеаторей.
Панкреатическая стеаторея (дефицит панкреатической липазы) – наблюдается при хроническом панкреатите,

ТКАНЕВЫЙ ЛИПОЛИЗ ТАГ.
ТКАНЕВЫЙ ЛИПОЛИЗ ТАГ.

Гидролиз ТАГ
Осуществляется липазами. Активность липазы в клетках жировой ткани находится под
Гидролиз ТАГ
Осуществляется липазами. Активность липазы в клетках жировой ткани находится под



В результате полного гидролиза молекулы ТАГ образуется три молекулы жирных кислот
В результате полного гидролиза молекулы ТАГ образуется три молекулы жирных кислот

ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
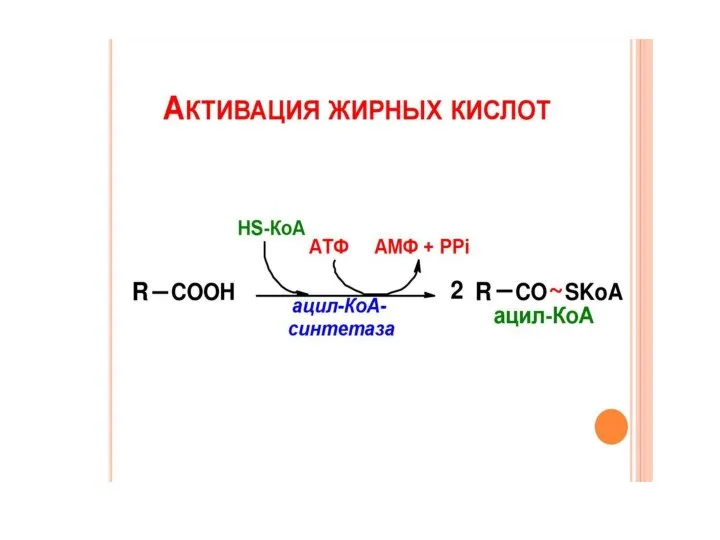

Транспорт активированных жирных кислот в митохондрии
Ацил-КоА не может проникнуть в митохондрию
Транспорт активированных жирных кислот в митохондрии
Ацил-КоА не может проникнуть в митохондрию

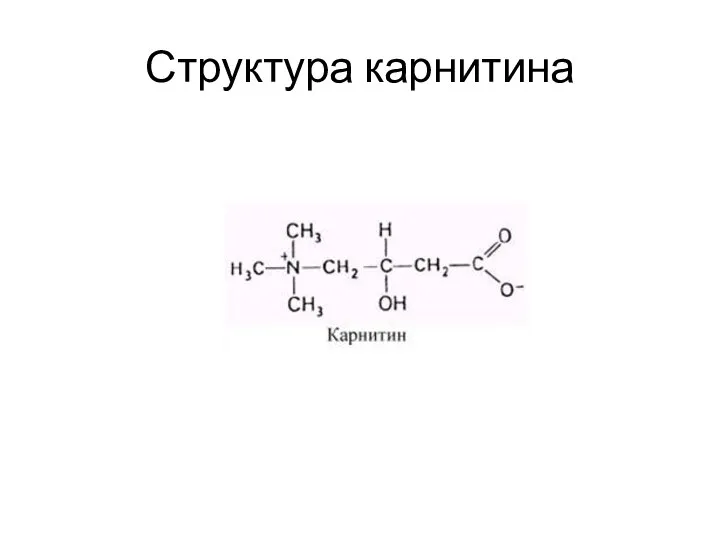
Структура карнитина
Структура карнитина

Современная теория β-окисления
Локализация – матрикс митохондрий
Генерирование энергии в виде:
Ацетил-коА
НАДН·Н +
ФАД
Современная теория β-окисления
Локализация – матрикс митохондрий
Генерирование энергии в виде:
Ацетил-коА
НАДН·Н +
ФАД
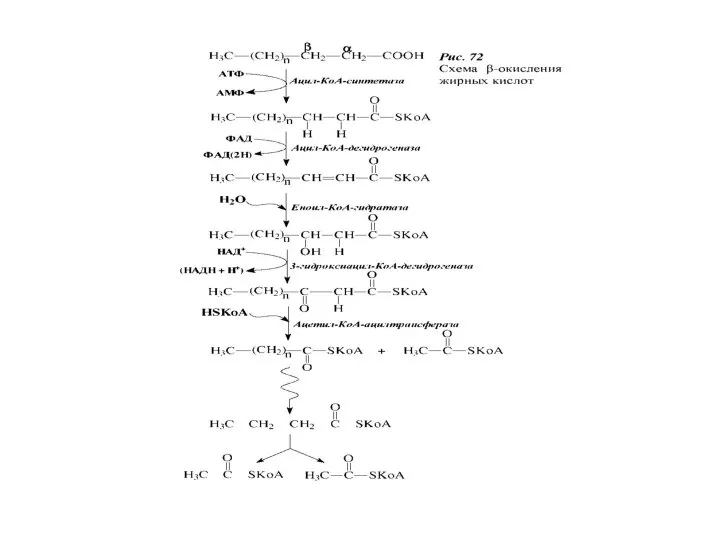


Большой вклад в изучение данной последовательности реакций внесли Д. Грин, С.
Большой вклад в изучение данной последовательности реакций внесли Д. Грин, С.
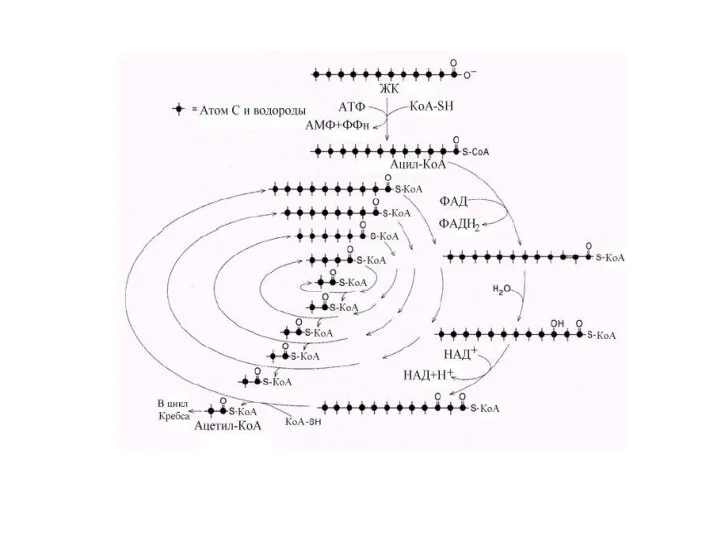

Энергетический выход β-окисления жирных кислот зависит от длины углеводородной цепи.
Расчет в
Энергетический выход β-окисления жирных кислот зависит от длины углеводородной цепи.
Расчет в

Суммарное уравнение окисления пальмитиновой кислоты.
Пальмитоил S-коА +7КоАSН + 7ФАД +7НАД +
Суммарное уравнение окисления пальмитиновой кислоты.
Пальмитоил S-коА +7КоАSН + 7ФАД +7НАД +


Окисление жирных кислот в пероксисомах
Жирные кислоты с очень длинной цепью. Особенностью
Окисление жирных кислот в пероксисомах
Жирные кислоты с очень длинной цепью. Особенностью

β-окисление ненасыщенных жирных кислот
β-окисление ненасыщенных жирных кислот во многом
β-окисление ненасыщенных жирных кислот
β-окисление ненасыщенных жирных кислот во многом

β-Окисление жирных кислот с нечетным числом углеродных атомов
Жирные кислоты с нечетным
β-Окисление жирных кислот с нечетным числом углеродных атомов
Жирные кислоты с нечетным
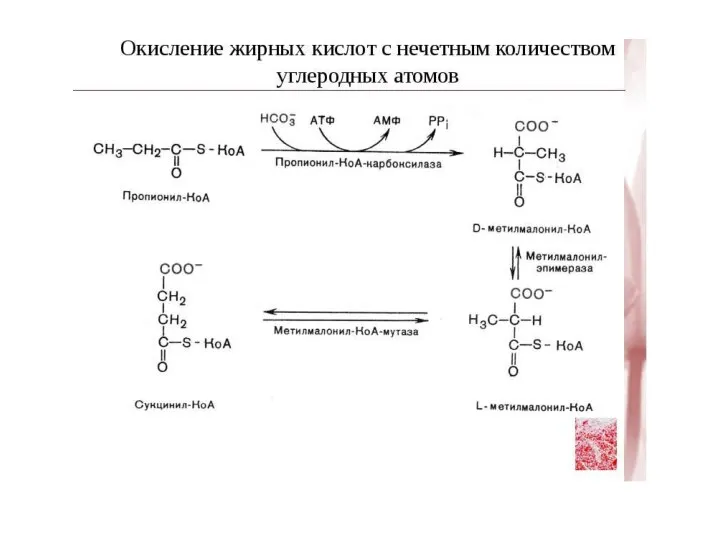

Пропионат также образуется при расщеплении четырех аминокислот: валина, изолейцина, метионина и
Пропионат также образуется при расщеплении четырех аминокислот: валина, изолейцина, метионина и

Ключевой процесс – поступление жирных кислот в митохондрию.
Карнитинацилтрансфераза – аллостерический фермент,
Ключевой процесс – поступление жирных кислот в митохондрию.
Карнитинацилтрансфераза – аллостерический фермент,
 Алюминий и его соединения
Алюминий и его соединения Лабораторная посуда и оборудование
Лабораторная посуда и оборудование Презентация по Химии "Классификация и номенклатура неорганических соединений" - скачать смотреть
Презентация по Химии "Классификация и номенклатура неорганических соединений" - скачать смотреть  Аммиак
Аммиак Формування мотивації до самостійної пізнавальної діяльності учнів на уроках хімії
Формування мотивації до самостійної пізнавальної діяльності учнів на уроках хімії Иондық және ковалентті байланысы бар қосылыстардың электролиттік диссоциациясы
Иондық және ковалентті байланысы бар қосылыстардың электролиттік диссоциациясы Полиэтилен
Полиэтилен Протолитические равновесия в растворах электролитов
Протолитические равновесия в растворах электролитов Сахар – практически чистая сахароза
Сахар – практически чистая сахароза Общая и медицинская химия
Общая и медицинская химия Ферменттер. Зерттелу тарихы
Ферменттер. Зерттелу тарихы The phenomenon of radioactivity. Radioisotopes. The nuclear reaction
The phenomenon of radioactivity. Radioisotopes. The nuclear reaction Вещественный состав магматических горных пород
Вещественный состав магматических горных пород Проведення полімеразної ланцюгової реакції з використанням діагностичних тест-систем
Проведення полімеразної ланцюгової реакції з використанням діагностичних тест-систем Неорганические соединения галогенов
Неорганические соединения галогенов Подгруппа мышьяка
Подгруппа мышьяка Алкадиены Учитель химии МОУ лицея №6 Дробот Светлана Сергеевна
Алкадиены Учитель химии МОУ лицея №6 Дробот Светлана Сергеевна Сера
Сера Підготувала учениця 10-Б класу Березянська Альона
Підготувала учениця 10-Б класу Березянська Альона  Оксиды. Названия оксидов
Оксиды. Названия оксидов Исследование Е. Е. Вагнера в области терпенов и камфоры
Исследование Е. Е. Вагнера в области терпенов и камфоры Приборы и методы исследования в химической технологии
Приборы и методы исследования в химической технологии Пищевые добавки
Пищевые добавки Комбинированные задачи. Базовые формулы
Комбинированные задачи. Базовые формулы Біотехнічні основи технології харчових продуктів
Біотехнічні основи технології харчових продуктів Производство самодельного мыла: технология, экономика
Производство самодельного мыла: технология, экономика Электрохимические методы анализа, часть 2. Кулонометрия. Вольтамперометрия
Электрохимические методы анализа, часть 2. Кулонометрия. Вольтамперометрия Предмет и объекты исследования коллоидной химии. Лекция 01
Предмет и объекты исследования коллоидной химии. Лекция 01