- Обмен липидов. Часть 2

Содержание
- 2. Ожирение Среди человеческой популяции ожирение представляет собой наиболее частое отклонение в обмене ТАГ. Эту патологию диагносцируют
- 3. Синтез и использование кетоновых тел В постабсорбтивный период и при голодании мобилизация ТАГ сопровождается повышением концентрации
- 4. Синтез и использование кетоновых тел Когда концентрация ацетил-КоАв матриксе митохондрий становится высокой,тиолаза катализирует обращение последней реакцииβ-окисленияи
- 5. Рис. 9.23. Метаболизм ВЖК в митохондриях печени в условиях голодания
- 6. Рис. 9.24. Синтез кетоновых тел в митохондриях печени
- 7. Синтез и использование кетоновых тел – при сахарном диабете всеми инсулин зависимыми тканями, за исключением печени.
- 8. Рис. 9.25. Окисление кетоновых тел
- 9. Метаболизм эйкозаноидов Эйкозаноиды — это группа сигнальных молекул местного действия, которые синтезируются практически во всех дифференцированных
- 10. Рис. 9.26. Освобождение арахидоновой кислоты из мембранных липидов. Связывание активаторов с рецептором плазматической мембраны активирует пути
- 11. Метаболизм эйкозаноидов В разных тканях арахидоновая и другие эйкозановые кислоты могут использоваться по трем основным направлениям:
- 12. Рис. 9.27. Пути использования арахидоновой кислоты на синтез эйкозаноидов. PG от английского слова prostaglandins — простагландины,
- 13. Схема. ЦОГ-зависимый путь метаболизма арахидоновой кислоты. Превращение арахидоновой кислоты (АХК) в эндоперикиси PgG2 и PgH2 (циклооксигеназная
- 14. Схема. ЛОГ-зависимый путь метаболизма арахидоновой кислоты. Липоксигеназная и дегидразная стадии контролируются ферментом 5-ЛОГ.
- 15. Метаболизм эйкозаноидов Основным общим предшественником простагландинов и тромбоксанов из семейства арахидоновой кислоты является PGН2, который синтезируется
- 16. Метаболизм эйкозаноидов Другой путь превращений арахидоновой кислоты катализируют липоксигеназы — группа ферментов, присоединяющих молекулу кислорода к
- 17. Метаболизм эйкозаноидов Под влиянием монооксигеназ, работающих с участием цитохромов Р 450, образуются эпоксиды, некоторые виды ГЭТЕ,
- 18. Метаболизм эйкозаноидов
- 19. Обмен холестерола Холестерол — основной стероид организма животных. У взрослого человека содержание холестерола составляет 140–150г. Около
- 20. Обмен холестерола Путь поступления экзогенного холестерола Холестерол содержится только в жирах животного происхождения в свободном виде
- 21. Рис. 9.28. Образования эфиров холестерола: АХАТ — ацилхолестеролацилтрансфераза
- 22. Обмен холестерола Синтез холестерола de novo Ферменты синтеза холестерола имеются практически во всех клетках организма, но
- 23. Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната; Б — превращение мевалоната в
- 24. Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната; Б — превращение мевалоната в
- 25. Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната; Б — превращение мевалоната в
- 26. Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната; Б — превращение мевалоната в
- 27. Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната; Б — превращение мевалоната в
- 28. Обмен холестерола. Синтез холестерола de novo Ключевой регуляторный фермент —ГМГ-КоА-редуктаза,активность которого в печени регулируется трояким способом
- 29. Рис. 9.30. Регуляция активности ГМГ-КоА-редуктазыв печени
- 30. Обмен холестерола. Синтез и функции желчных кислот В печени ежесуточно около 0,5 г холестерола используется на
- 31. Рис. 9.31. Реакция образования 7-α-холестерола
- 32. Обмен холестерола. Синтез и функции желчных кислот Затем происходит восстановление двойной связи В-кольцаи часть молекул подвергается
- 33. Рис. 9.32. Конъюгация желчных кислот
- 34. Обмен холестерола. Синтез и функции желчных кислот Свободные и парные или конъюгированные желчные кислоты, синтезирующиеся печенью,
- 36. Рис. 9.33. Энтерогепатическая циркуляция желчных кислот
- 37. Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов В транспорте холестерола и его эфиров по
- 38. Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов Важную роль в обмене холестерола играют ЛПВП.
- 39. Рис. 9.34. Реакция, катализируемая ЛХАТ: ЛХАТ — лецитинхолестеролацилтрансфераза
- 40. Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов Второй продукт реакции — лизолецитин связывается с
- 41. Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов Ворганизме ХМ и ЛПОНП с помощью ЛП-липазыобеспечивают
- 42. Рис. 9.35. Участие ЛПВП в транспорте холестерола
- 43. Обмен холестерола. Гиперлипидемии и атеросклероз В норме общая концентрация холестерола ( свободный Хс + ЭХс )
- 44. Обмен холестерола. Гиперлипидемии и атеросклероз Таблица 9.3
- 45. Обмен холестерола. Гиперлипидемии и атеросклероз Таблица 9.3
- 46. Обмен холестерола. Гиперлипидемии и атеросклероз в норме у взрослого человека этот показатель не должен превышать 3–4.
- 48. Скачать презентацию

Ожирение
Среди человеческой популяции ожирение представляет собой наиболее частое отклонение в
Ожирение
Среди человеческой популяции ожирение представляет собой наиболее частое отклонение в

Синтез и использование кетоновых тел
В постабсорбтивный период и при голодании
Синтез и использование кетоновых тел
В постабсорбтивный период и при голодании

Синтез и использование кетоновых тел
Когда концентрация ацетил-КоАв матриксе митохондрий становится высокой,тиолаза катализирует
Синтез и использование кетоновых тел
Когда концентрация ацетил-КоАв матриксе митохондрий становится высокой,тиолаза катализирует
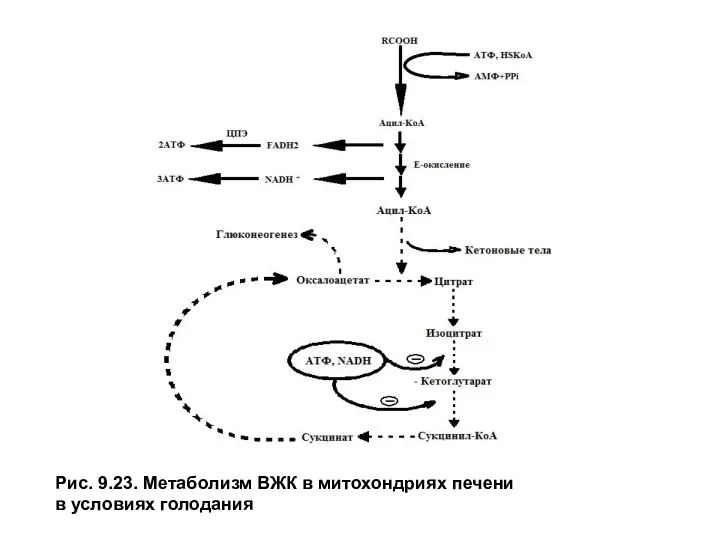
Рис. 9.23. Метаболизм ВЖК в митохондриях печени
в условиях голодания
Рис. 9.23. Метаболизм ВЖК в митохондриях печени
в условиях голодания
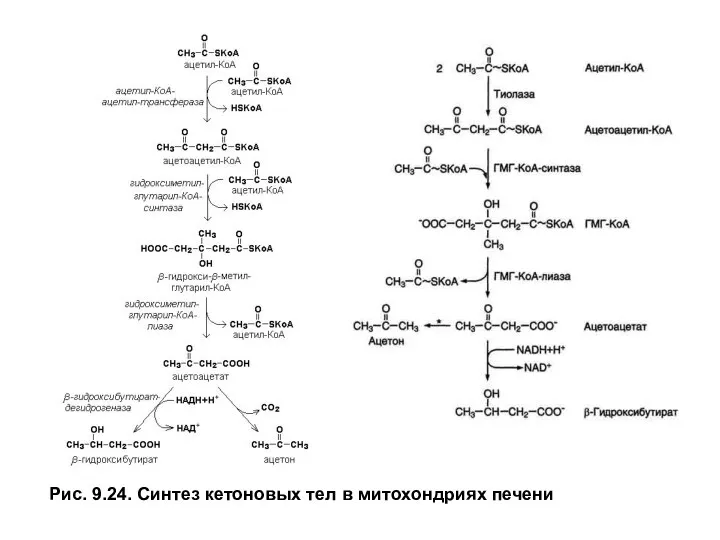
Рис. 9.24. Синтез кетоновых тел в митохондриях печени
Рис. 9.24. Синтез кетоновых тел в митохондриях печени

Синтез и использование кетоновых тел
– при сахарном диабете всеми инсулин
Синтез и использование кетоновых тел
– при сахарном диабете всеми инсулин
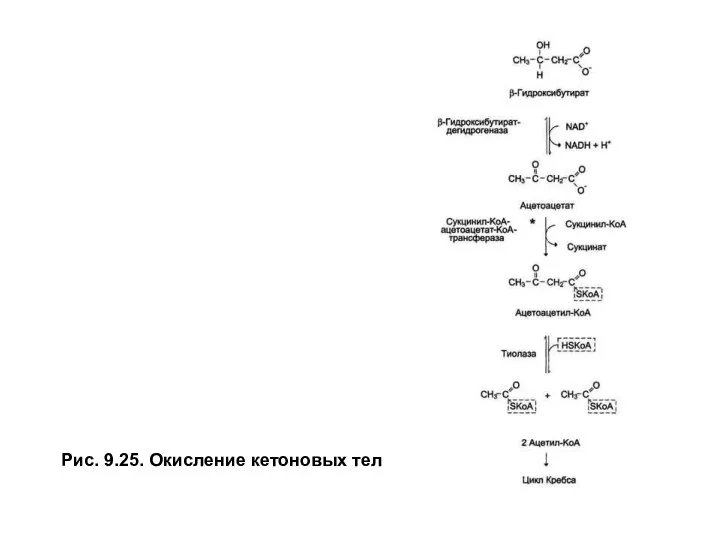
Рис. 9.25. Окисление кетоновых тел
Рис. 9.25. Окисление кетоновых тел

Метаболизм эйкозаноидов
Эйкозаноиды — это группа сигнальных молекул местного действия, которые синтезируются
Метаболизм эйкозаноидов
Эйкозаноиды — это группа сигнальных молекул местного действия, которые синтезируются
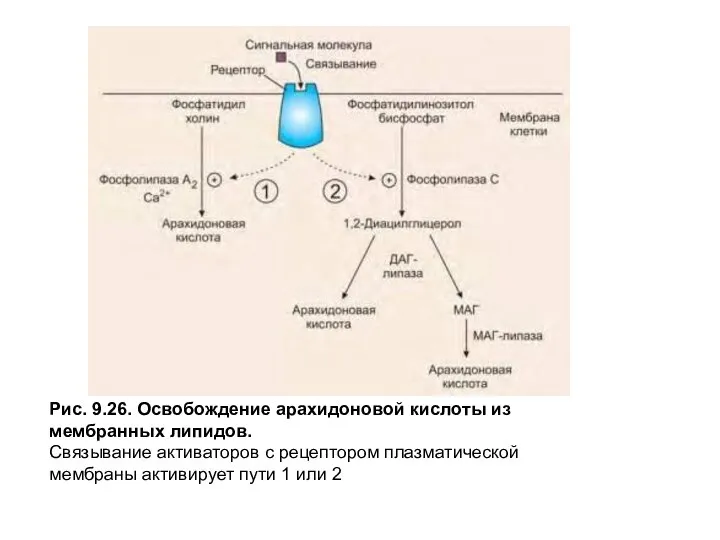
Рис. 9.26. Освобождение арахидоновой кислоты из мембранных липидов.
Связывание активаторов с рецептором
Рис. 9.26. Освобождение арахидоновой кислоты из мембранных липидов.
Связывание активаторов с рецептором

Метаболизм эйкозаноидов
В разных тканях арахидоновая и другие эйкозановые кислоты могут
Метаболизм эйкозаноидов
В разных тканях арахидоновая и другие эйкозановые кислоты могут
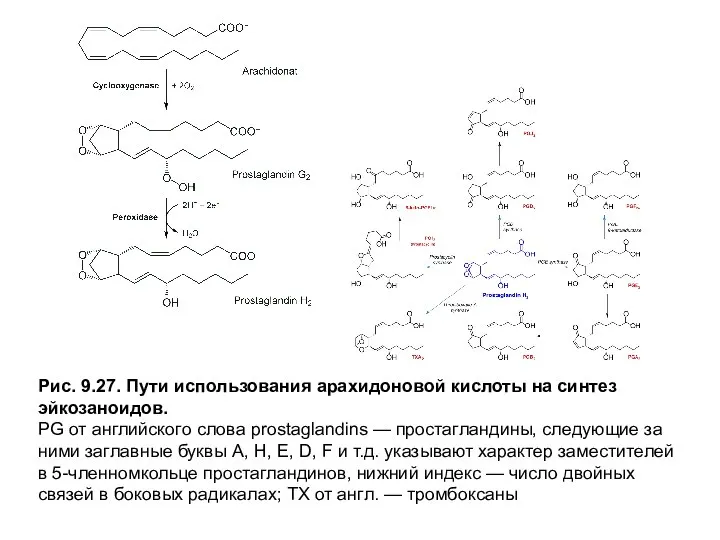
Рис. 9.27. Пути использования арахидоновой кислоты на синтез эйкозаноидов.
PG от английского
Рис. 9.27. Пути использования арахидоновой кислоты на синтез эйкозаноидов.
PG от английского
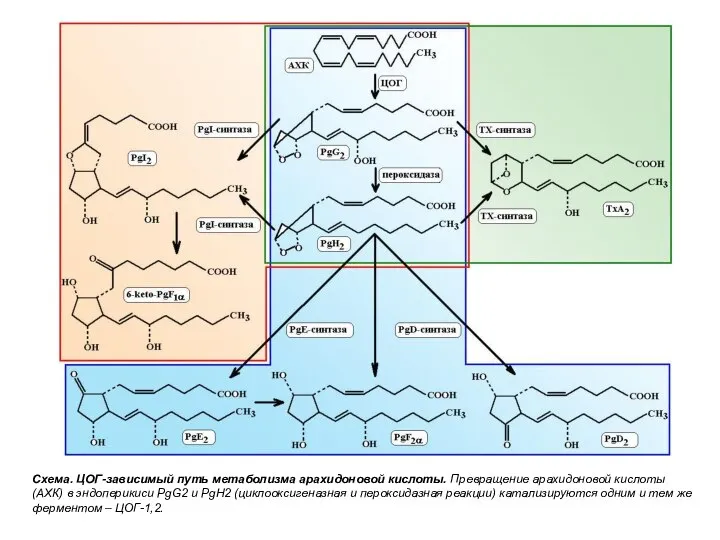
Схема. ЦОГ-зависимый путь метаболизма арахидоновой кислоты. Превращение арахидоновой кислоты (АХК) в
Схема. ЦОГ-зависимый путь метаболизма арахидоновой кислоты. Превращение арахидоновой кислоты (АХК) в
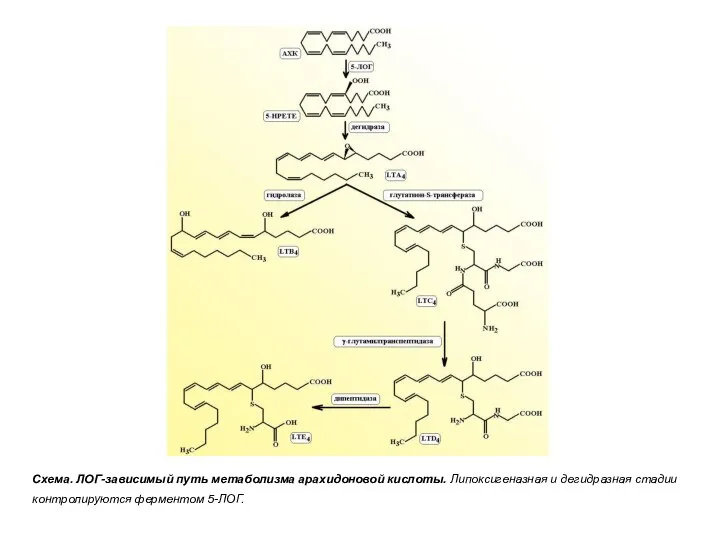
Схема. ЛОГ-зависимый путь метаболизма арахидоновой кислоты. Липоксигеназная и дегидразная стадии контролируются
Схема. ЛОГ-зависимый путь метаболизма арахидоновой кислоты. Липоксигеназная и дегидразная стадии контролируются

Метаболизм эйкозаноидов
Основным общим предшественником простагландинов и тромбоксанов из семейства арахидоновой
Метаболизм эйкозаноидов
Основным общим предшественником простагландинов и тромбоксанов из семейства арахидоновой

Метаболизм эйкозаноидов
Другой путь превращений арахидоновой кислоты катализируют липоксигеназы — группа ферментов, присоединяющих
Метаболизм эйкозаноидов
Другой путь превращений арахидоновой кислоты катализируют липоксигеназы — группа ферментов, присоединяющих

Метаболизм эйкозаноидов
Под влиянием монооксигеназ, работающих с участием цитохромов Р 450,
Метаболизм эйкозаноидов
Под влиянием монооксигеназ, работающих с участием цитохромов Р 450,
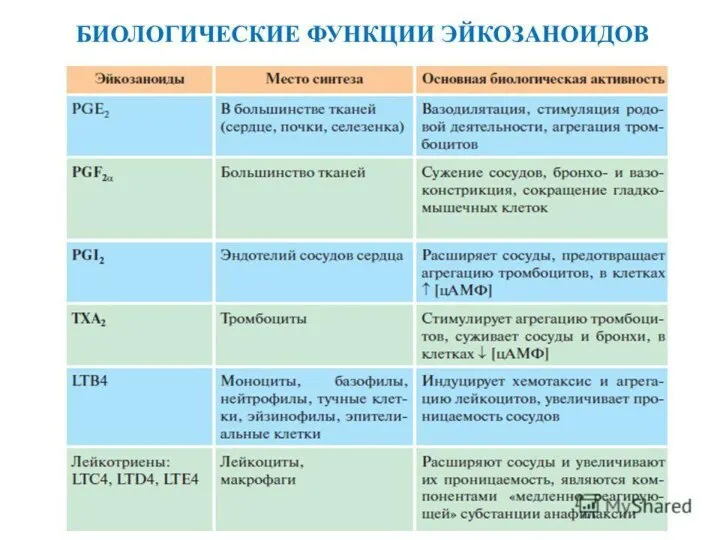
Метаболизм эйкозаноидов
Метаболизм эйкозаноидов

Обмен холестерола
Холестерол — основной стероид организма животных. У взрослого человека
Обмен холестерола
Холестерол — основной стероид организма животных. У взрослого человека

Обмен холестерола
Путь поступления экзогенного холестерола
Холестерол содержится только в жирах животного
Обмен холестерола
Путь поступления экзогенного холестерола
Холестерол содержится только в жирах животного
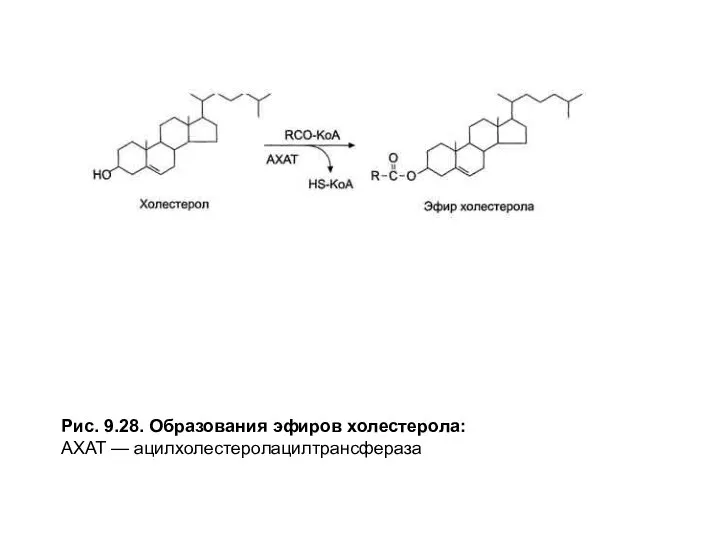
Рис. 9.28. Образования эфиров холестерола:
АХАТ — ацилхолестеролацилтрансфераза
Рис. 9.28. Образования эфиров холестерола:
АХАТ — ацилхолестеролацилтрансфераза

Обмен холестерола
Синтез холестерола de novo
Ферменты синтеза холестерола имеются практически во
Обмен холестерола
Синтез холестерола de novo
Ферменты синтеза холестерола имеются практически во
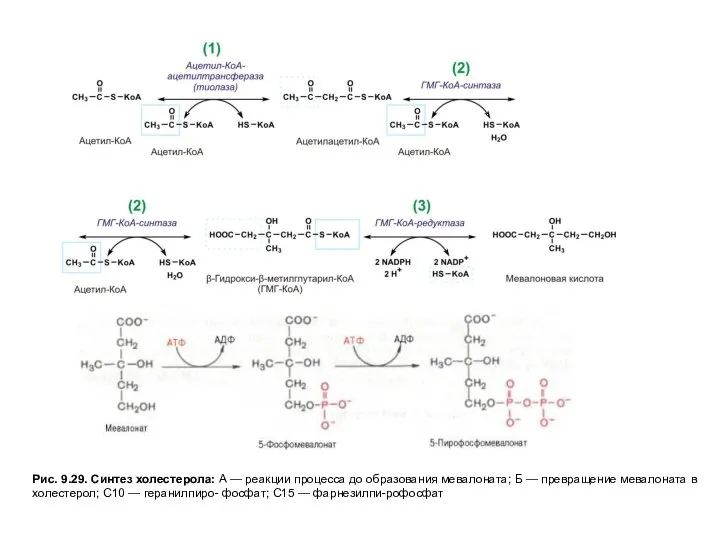
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
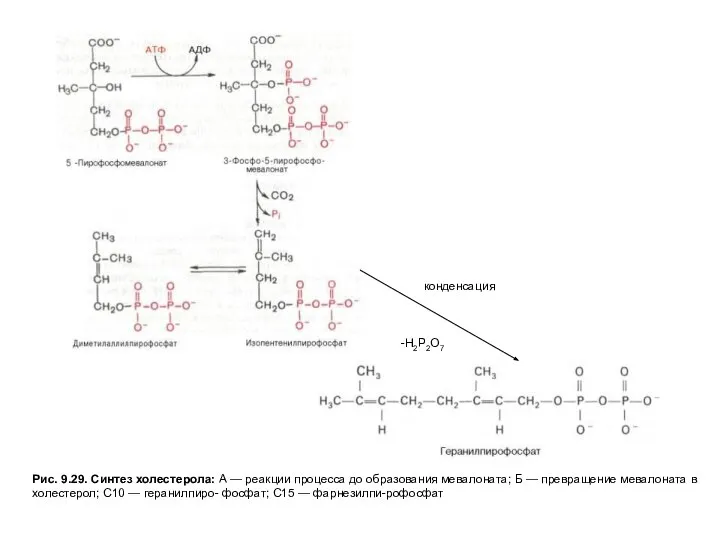
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
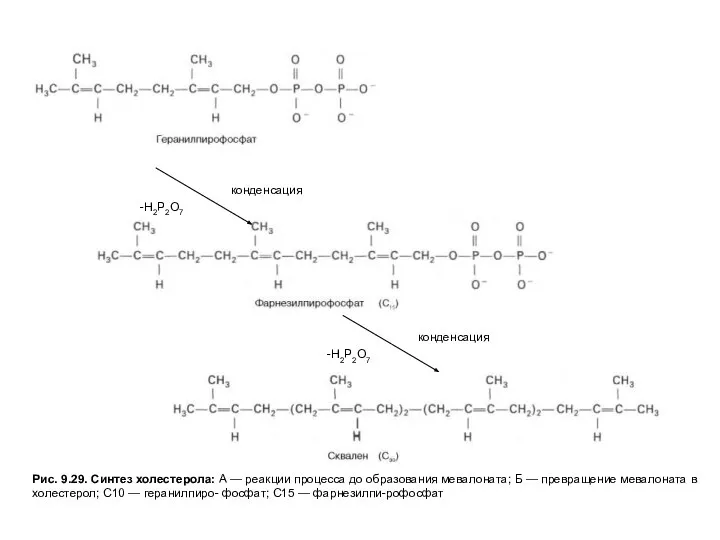
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
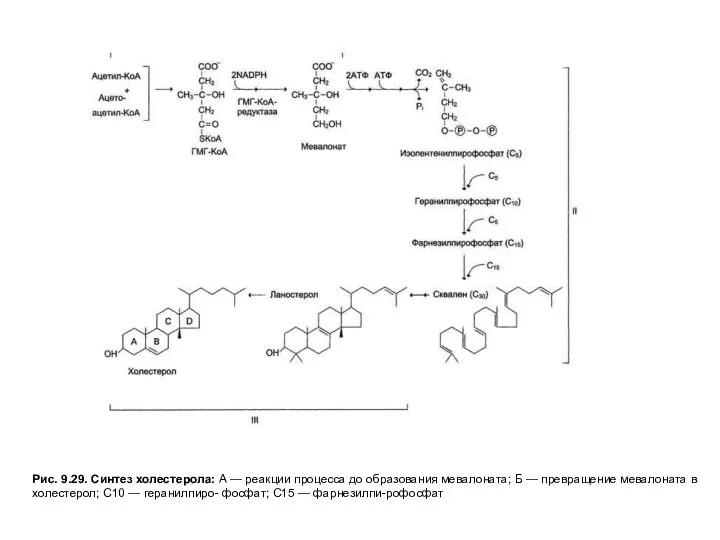
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
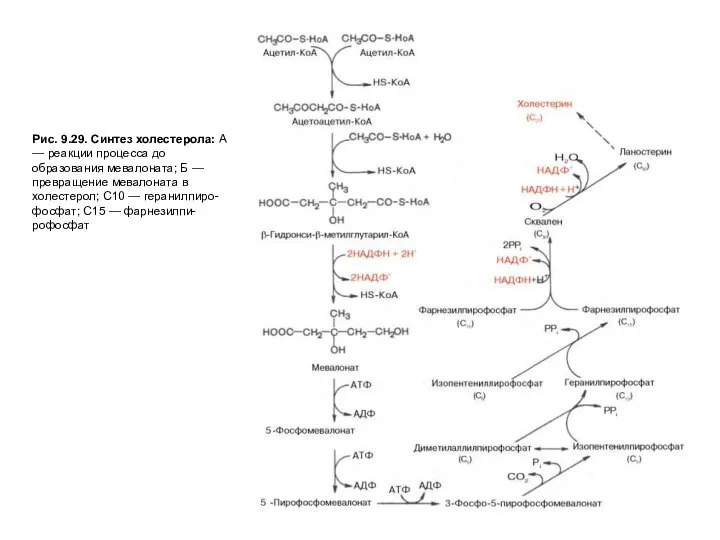
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;
Рис. 9.29. Синтез холестерола: А — реакции процесса до образования мевалоната;

Обмен холестерола. Синтез холестерола de novo
Ключевой регуляторный фермент —ГМГ-КоА-редуктаза,активность которого в печени
Обмен холестерола. Синтез холестерола de novo
Ключевой регуляторный фермент —ГМГ-КоА-редуктаза,активность которого в печени
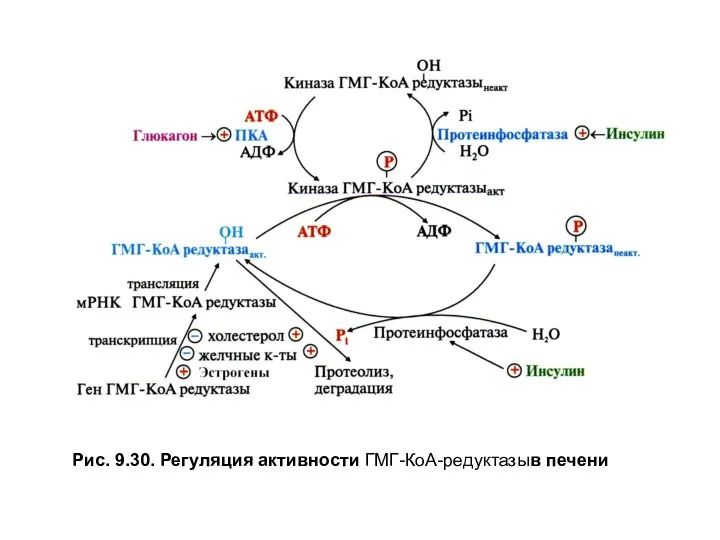
Рис. 9.30. Регуляция активности ГМГ-КоА-редуктазыв печени
Рис. 9.30. Регуляция активности ГМГ-КоА-редуктазыв печени

Обмен холестерола.
Синтез и функции желчных кислот
В печени ежесуточно около 0,5
Обмен холестерола.
Синтез и функции желчных кислот
В печени ежесуточно около 0,5
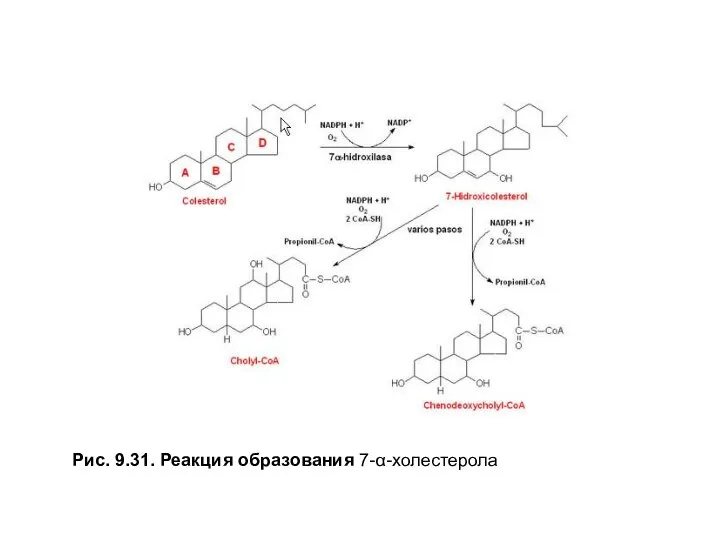
Рис. 9.31. Реакция образования 7-α-холестерола
Рис. 9.31. Реакция образования 7-α-холестерола

Обмен холестерола. Синтез и функции желчных кислот
Затем происходит восстановление двойной связи В-кольцаи
Обмен холестерола. Синтез и функции желчных кислот
Затем происходит восстановление двойной связи В-кольцаи
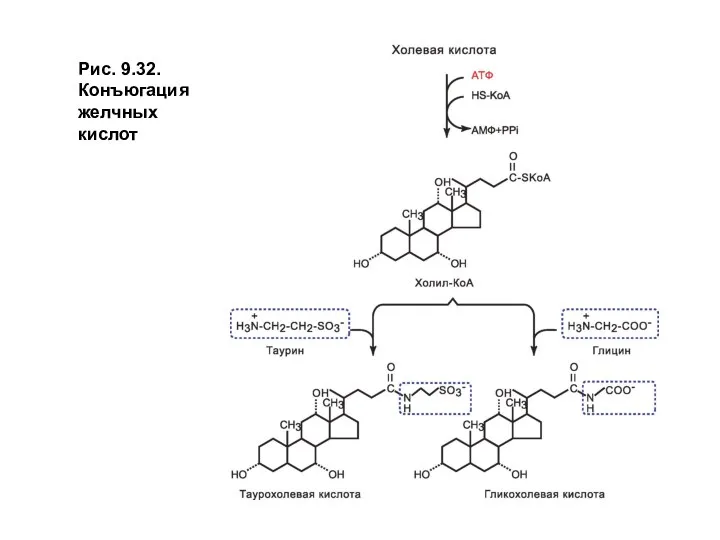
Рис. 9.32. Конъюгация желчных кислот
Рис. 9.32. Конъюгация желчных кислот

Обмен холестерола. Синтез и функции желчных кислот
Свободные и парные или конъюгированные
Обмен холестерола. Синтез и функции желчных кислот
Свободные и парные или конъюгированные
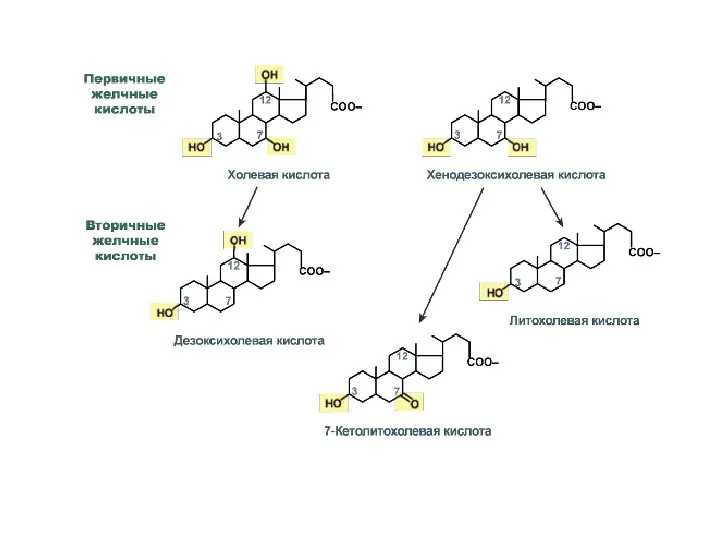
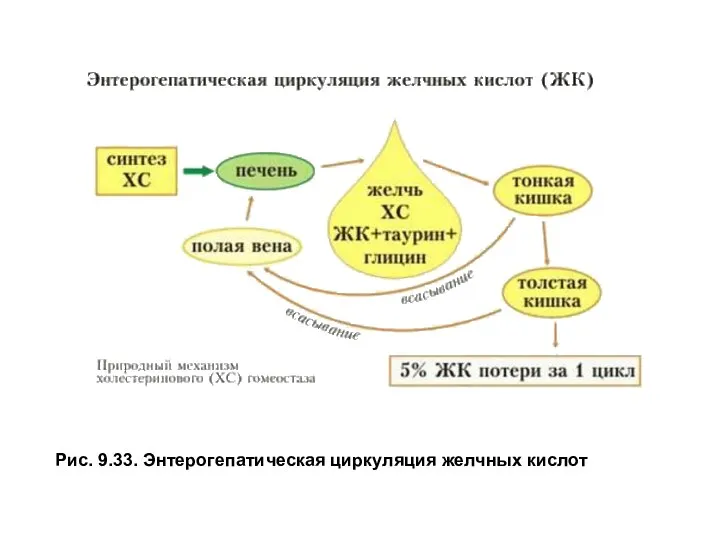
Рис. 9.33. Энтерогепатическая циркуляция желчных кислот
Рис. 9.33. Энтерогепатическая циркуляция желчных кислот

Обмен холестерола.
Транспорт холестерола по крови в составе липопротеинов
В транспорте холестерола
Обмен холестерола.
Транспорт холестерола по крови в составе липопротеинов
В транспорте холестерола

Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Важную роль в
Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Важную роль в
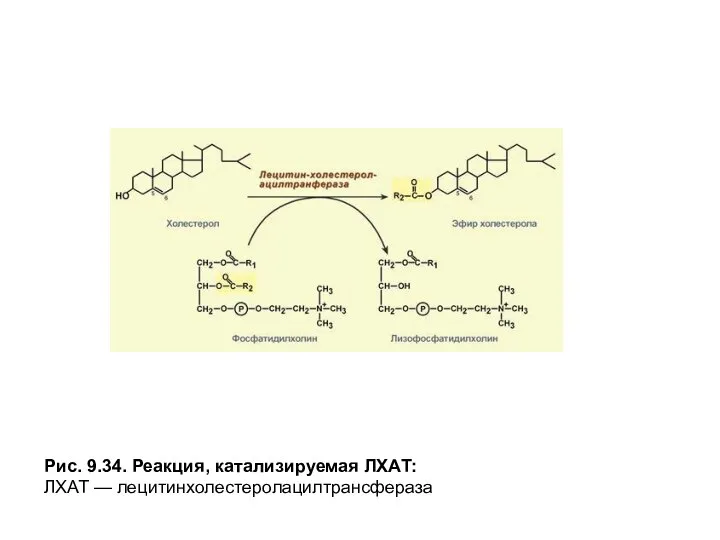
Рис. 9.34. Реакция, катализируемая ЛХАТ:
ЛХАТ — лецитинхолестеролацилтрансфераза
Рис. 9.34. Реакция, катализируемая ЛХАТ:
ЛХАТ — лецитинхолестеролацилтрансфераза

Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Второй продукт реакции
Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Второй продукт реакции

Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Ворганизме ХМ и
Обмен холестерола. Транспорт холестерола по крови в составе липопротеинов
Ворганизме ХМ и
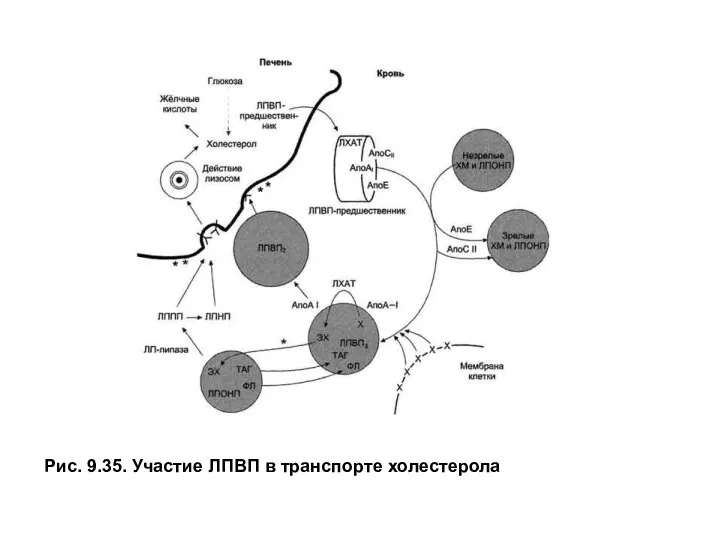
Рис. 9.35. Участие ЛПВП в транспорте холестерола
Рис. 9.35. Участие ЛПВП в транспорте холестерола

Обмен холестерола.
Гиперлипидемии и атеросклероз
В норме общая концентрация холестерола ( свободный Хс
Обмен холестерола.
Гиперлипидемии и атеросклероз
В норме общая концентрация холестерола ( свободный Хс
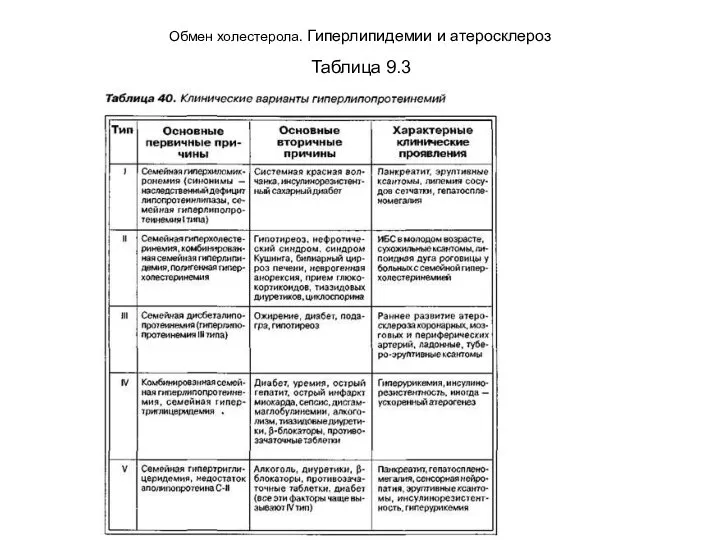
Обмен холестерола. Гиперлипидемии и атеросклероз
Таблица 9.3
Обмен холестерола. Гиперлипидемии и атеросклероз
Таблица 9.3
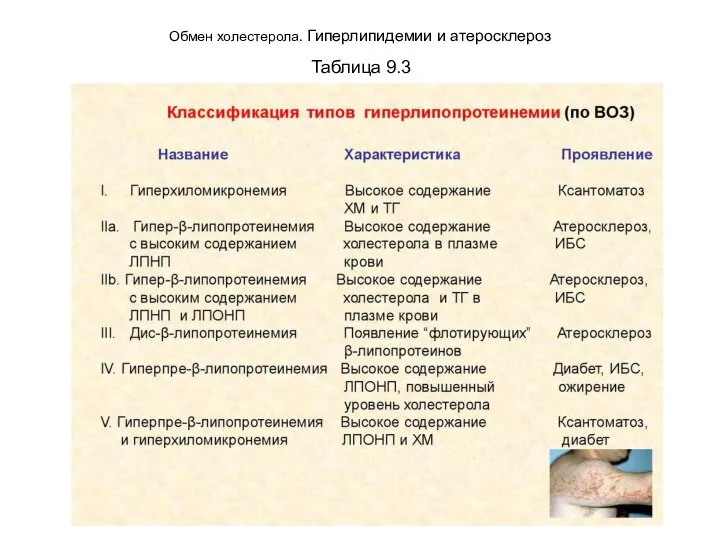
Обмен холестерола. Гиперлипидемии и атеросклероз
Таблица 9.3
Обмен холестерола. Гиперлипидемии и атеросклероз
Таблица 9.3

Обмен холестерола. Гиперлипидемии и атеросклероз
в норме у взрослого человека этот показатель
Обмен холестерола. Гиперлипидемии и атеросклероз
в норме у взрослого человека этот показатель
 Органические и неорганические вещества. Нуклеиновые кислоты
Органические и неорганические вещества. Нуклеиновые кислоты ОМЕГА 3 и ОМЕГА 6 полиненасыщенные жирные кислоты (ПНЖК) Все более популярными в ветеринарии становятся ОМЕГА 3 и ОМЕГА 6 полиненасы
ОМЕГА 3 и ОМЕГА 6 полиненасыщенные жирные кислоты (ПНЖК) Все более популярными в ветеринарии становятся ОМЕГА 3 и ОМЕГА 6 полиненасы Тест: Азот
Тест: Азот Витамины и их производные-коферменты ферментов
Витамины и их производные-коферменты ферментов Получение гидроксида никеля (II)
Получение гидроксида никеля (II) Розчини електролітів
Розчини електролітів Вода – растворитель. Растворы. Значение воды
Вода – растворитель. Растворы. Значение воды Використання кислот
Використання кислот Гелий
Гелий Индол. Методы синтеза. Химические свойства. Биологически активные производные. (Лекция 4)
Индол. Методы синтеза. Химические свойства. Биологически активные производные. (Лекция 4) Карбон қышқылдары
Карбон қышқылдары Вступ в гетерогенний катализ. Загальні положення каталізу
Вступ в гетерогенний катализ. Загальні положення каталізу Синтез и свойства кукурбитурилов
Синтез и свойства кукурбитурилов Жиры. 9 класс
Жиры. 9 класс ОКСИДЫ АЗОТА.
ОКСИДЫ АЗОТА.  ВПР Окружающий мир 4 класс
ВПР Окружающий мир 4 класс  Урок по теме: «Жиры. Техническая переработка жиров» Пермякова Ирина Александровна Учитель химии ГБОУ СОШ № 982 г. Москва Индиф
Урок по теме: «Жиры. Техническая переработка жиров» Пермякова Ирина Александровна Учитель химии ГБОУ СОШ № 982 г. Москва Индиф Условия хранения и подготовка скоропортящихся грузов к перевозкам
Условия хранения и подготовка скоропортящихся грузов к перевозкам Инертные газы
Инертные газы Агрегатные состояния вещества
Агрегатные состояния вещества Нанопористые материалы
Нанопористые материалы Предельные углеводороды. Химические свойства. 10 класс
Предельные углеводороды. Химические свойства. 10 класс Природные и синтетические красители и ароматизаторы
Природные и синтетические красители и ароматизаторы Химические свойства фурана, пиррола, тиофена. Биологически активные производные ряда фурана, пиррола, тиофена. (Лекция 3)
Химические свойства фурана, пиррола, тиофена. Биологически активные производные ряда фурана, пиррола, тиофена. (Лекция 3) МОУ «Каратунская СОШ с углубленным изучением отдельных предметов» Выполнила : учитель химии второй квалификационной категор
МОУ «Каратунская СОШ с углубленным изучением отдельных предметов» Выполнила : учитель химии второй квалификационной категор Элективный курс "Решение задач по химии повышенного уровня сложности"
Элективный курс "Решение задач по химии повышенного уровня сложности" Сложные эфиры R-COOR'
Сложные эфиры R-COOR' Оценка опасности взрыва горючих газов
Оценка опасности взрыва горючих газов