- Клеточный цикл и его регуляция

Содержание
- 2. Клеточный цикл и его регуляция G0 G1 S G2 M интерфаза профаза метафаза анафаза телофаза ЦИТОКИНЕЗ
- 3. G0 G1 G2 S M характеризуется экспрессией генов и синтезом белков. Клетка становится способной к росту
- 4. прохождение цикла обеспечивается последовательной сменой активированных cyclin-зависимых киназ, для активации которых необходимо присутствие соответствующей активаторной субъединицы
- 5. G0 G1 S M cyclin E Cdk2 пресинтетическая постсинтетическая синтез ДНК митоз cyclin В Cdс2 Точки
- 6. В регуляции активности checkpoint задействованы три группы генов, кодирующих: 1) белки-сенсоры, которые контролируют завершение событий клеточного
- 7. Три гена отслеживают повреждение ДНК в checkpoint: регулирует активность E2F-DP, фактора транскрипции cyclin A, E и
- 8. Клеточный цикл регулируется не только внутриклеточ- ными, но также и внешними факторами. ФР и состояние внеклеточного
- 9. КАНЦЕРОГЕНЕЗ многоступенчатый процесс накопления мутаций и других генетических изменений, приводящих к нарушениям регуляции клеточного цикла (гиперактивация
- 10. химические канцерогены: бензпирен и диметилбензантрацен, нитрозамины; многие инсектициды и гербициды, полициклические ароматические углеводороды; цикламаты и сахарин,
- 11. вирусы - индуцируют опухоли (возникновение лейкозов связана с РНК -содержащими вирусами, а другие опухоли вызываются ДНК-содержащими).
- 12. ТЕОРИИ КАНЦЕРОГЕНЕЗА Мутационная теория канцерогенеза Развитие опухолевого процесса - следствие мутации. Химические канцерогенные вещества, излучения вызывают
- 13. Процесс непосредственного действия канцерогена на клетки, запускающий их трансформацию, называется инициацией опухолевого роста Опухолевая трансформация -
- 14. Трансформированные клетки характеризуются: нарушением дифференцировки клоны клеток, как бы "заморожены" на той или иной стадии созревания,
- 15. ослаблением адгезивных свойств нарушение клеточной адгезии и подвижности играет фундаментальную роль в инициации канцерогенеза. Нарушения адгезионных
- 16. дефектами апоптоза для опухолевых клеток характерно ослабление обоих путей индукции апоптоза. Обнаруживаются: - потеря экспрессии на
- 17. Иммунологический атипизм - появление в опухолях белков, имеющих АГ значение для организма - хозяина Атипизм регуляции
- 18. ПРОМОЦИЯ Мутации могут проявляться при воздействии опухолевого промотора даже спустя большой промежуток времени. Промотор стимулирует деление
- 19. ОПУХОЛЕВАЯ ПРОГРЕССИЯ процесс малигнизации опухоли, в основе которого лежит дальнейшая трансформация генетического материала клеток Изначально опухоль
- 20. Генетическая нестабильность популяций опухолевых клеток складывается из основных типов нарушений: уменьшения точности репликации ДНК и сегрегации
- 21. качественные изменения в ходе опухолевого роста представляют микроэволюционный процесс, в котором генетическая нестабильность клетки обусловливает гено-
- 22. ПРОТИВООПУХОЛЕВЫЙ ИММУНИТЕТ Уничтожение трансформированных клеток с помощью клеточных и иммунных реакций называется иммунным надзором за клетками
- 23. Эффекторные механизмы противоопухолевого иммунитета Неспецифические ФНОα, ИФγ, ИЛ-2 Тh1 CD4+ NK CD56/CD57 К.-М. перфорины перфорины АЗКЦ
- 24. МНСI АГ ИЛ-1, 6, 12; ФНОα ИЛ-2; ИФ-γ АПК Т (CD4) Тh1 перфорины К.-М. МФ
- 25. Специфические МНСI АГ К.-М. АПК Т (CD4) Тk(CD8) перфорины цитотоксические гуморальные C5b6789 МАК комплементарный К.-М. комплемент
- 26. ВЛИЯНИЕ ОПУХОЛИ НА ОРГАНИЗМ Местные обусловлены характером роста опухоли экспансивный рост
- 27. инфильтративный рост аппозиционный рост происходит за счет неопластической трансформации нормальных клеток в опухолевые, что наблюдается в
- 28. экзофитный рост эндофитный рост
- 29. метастазирование проникновение перенос остановка выход рост
- 30. ПАРАНЕОПЛАСТИЧЕСКИЙ СИНДРОМ дистанционное (системное) воздействие новообразования, не связанное с прямой инвазией или компрессией опухоли и/или метастазами
- 32. Скачать презентацию
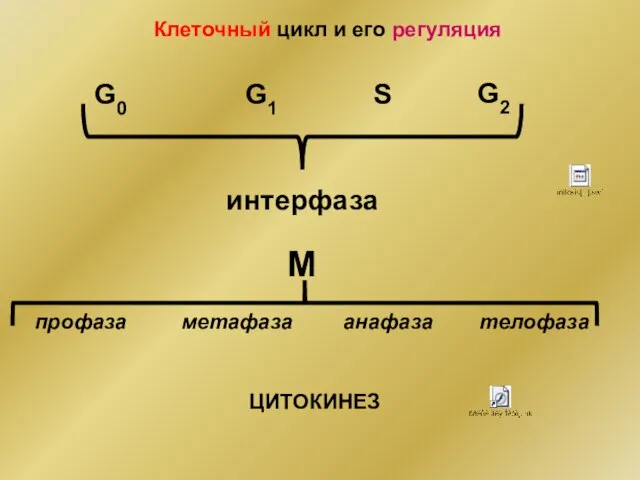
Клеточный цикл и его регуляция
G0
G1
S
G2
M
интерфаза
профаза
метафаза
анафаза
телофаза
ЦИТОКИНЕЗ
Клеточный цикл и его регуляция
G0
G1
S
G2
M
интерфаза
профаза
метафаза
анафаза
телофаза
ЦИТОКИНЕЗ
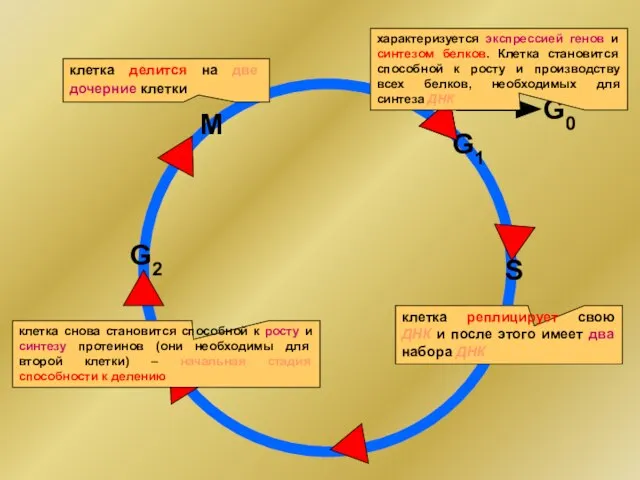
G0
G1
G2
S
M
характеризуется экспрессией генов и синтезом белков. Клетка становится способной к росту
G0
G1
G2
S
M
характеризуется экспрессией генов и синтезом белков. Клетка становится способной к росту

прохождение цикла обеспечивается последовательной сменой активированных cyclin-зависимых киназ, для активации которых
прохождение цикла обеспечивается последовательной сменой активированных cyclin-зависимых киназ, для активации которых
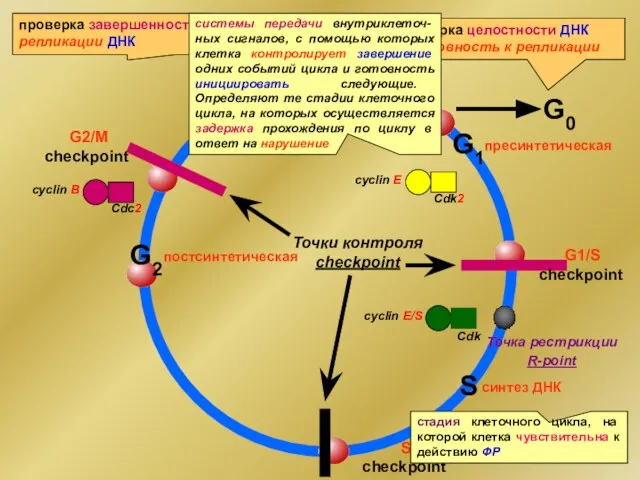
G0
G1
S
M
cyclin E
Cdk2
пресинтетическая
постсинтетическая
синтез ДНК
митоз
cyclin В
Cdс2
Точки контроля checkpoint
G2
G0
G1
S
M
cyclin E
Cdk2
пресинтетическая
постсинтетическая
синтез ДНК
митоз
cyclin В
Cdс2
Точки контроля checkpoint
G2

В регуляции активности checkpoint задействованы три группы генов, кодирующих:
1) белки-сенсоры, которые
В регуляции активности checkpoint задействованы три группы генов, кодирующих:
1) белки-сенсоры, которые

Три гена отслеживают повреждение ДНК в checkpoint:
регулирует активность E2F-DP, фактора транскрипции
Три гена отслеживают повреждение ДНК в checkpoint:
регулирует активность E2F-DP, фактора транскрипции

Клеточный цикл регулируется не только внутриклеточ- ными, но также и внешними
Клеточный цикл регулируется не только внутриклеточ- ными, но также и внешними

КАНЦЕРОГЕНЕЗ
многоступенчатый процесс накопления мутаций и других генетических изменений, приводящих к
КАНЦЕРОГЕНЕЗ
многоступенчатый процесс накопления мутаций и других генетических изменений, приводящих к

химические канцерогены: бензпирен и диметилбензантрацен, нитрозамины; многие инсектициды и гербициды, полициклические
химические канцерогены: бензпирен и диметилбензантрацен, нитрозамины; многие инсектициды и гербициды, полициклические

вирусы - индуцируют опухоли (возникновение лейкозов связана с РНК -содержащими вирусами,
вирусы - индуцируют опухоли (возникновение лейкозов связана с РНК -содержащими вирусами,

ТЕОРИИ КАНЦЕРОГЕНЕЗА
Мутационная теория канцерогенеза
Развитие опухолевого процесса - следствие мутации.
ТЕОРИИ КАНЦЕРОГЕНЕЗА
Мутационная теория канцерогенеза
Развитие опухолевого процесса - следствие мутации.

Процесс непосредственного действия канцерогена на клетки, запускающий их трансформацию, называется инициацией
Процесс непосредственного действия канцерогена на клетки, запускающий их трансформацию, называется инициацией

Трансформированные клетки характеризуются:
нарушением дифференцировки
клоны клеток, как бы "заморожены" на той или
Трансформированные клетки характеризуются:
нарушением дифференцировки
клоны клеток, как бы "заморожены" на той или

ослаблением адгезивных свойств
нарушение клеточной адгезии и подвижности играет фундаментальную роль в
ослаблением адгезивных свойств
нарушение клеточной адгезии и подвижности играет фундаментальную роль в

дефектами апоптоза
для опухолевых клеток характерно ослабление обоих путей индукции апоптоза. Обнаруживаются:
дефектами апоптоза
для опухолевых клеток характерно ослабление обоих путей индукции апоптоза. Обнаруживаются:

Иммунологический атипизм - появление в опухолях белков, имеющих АГ значение для
Иммунологический атипизм - появление в опухолях белков, имеющих АГ значение для

ПРОМОЦИЯ
Мутации могут проявляться при воздействии опухолевого промотора даже спустя большой
ПРОМОЦИЯ
Мутации могут проявляться при воздействии опухолевого промотора даже спустя большой

ОПУХОЛЕВАЯ ПРОГРЕССИЯ
процесс малигнизации опухоли, в основе которого лежит дальнейшая трансформация
ОПУХОЛЕВАЯ ПРОГРЕССИЯ
процесс малигнизации опухоли, в основе которого лежит дальнейшая трансформация

Генетическая нестабильность популяций опухолевых клеток складывается из основных типов нарушений:
уменьшения
Генетическая нестабильность популяций опухолевых клеток складывается из основных типов нарушений:
уменьшения

качественные изменения в ходе опухолевого роста представляют микроэволюционный процесс, в котором
качественные изменения в ходе опухолевого роста представляют микроэволюционный процесс, в котором

ПРОТИВООПУХОЛЕВЫЙ ИММУНИТЕТ
Уничтожение трансформированных клеток с помощью клеточных и иммунных реакций называется
ПРОТИВООПУХОЛЕВЫЙ ИММУНИТЕТ
Уничтожение трансформированных клеток с помощью клеточных и иммунных реакций называется
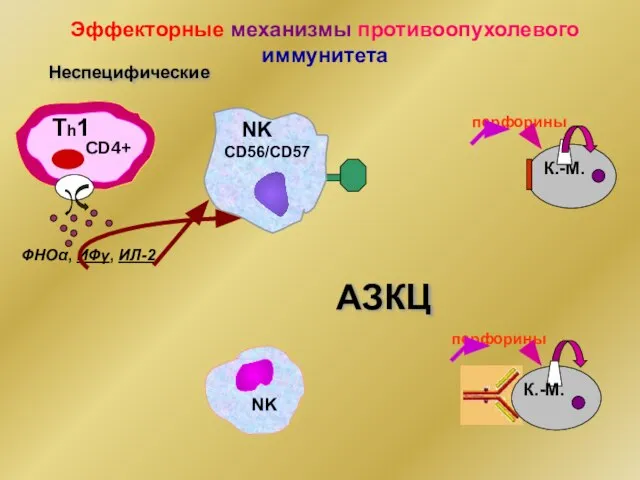
Эффекторные механизмы противоопухолевого иммунитета
Неспецифические
ФНОα, ИФγ, ИЛ-2
Тh1
CD4+
NK
CD56/CD57
К.-М.
перфорины
перфорины
АЗКЦ
NK
К.-М.
Эффекторные механизмы противоопухолевого иммунитета
Неспецифические
ФНОα, ИФγ, ИЛ-2
Тh1
CD4+
NK
CD56/CD57
К.-М.
перфорины
перфорины
АЗКЦ
NK
К.-М.
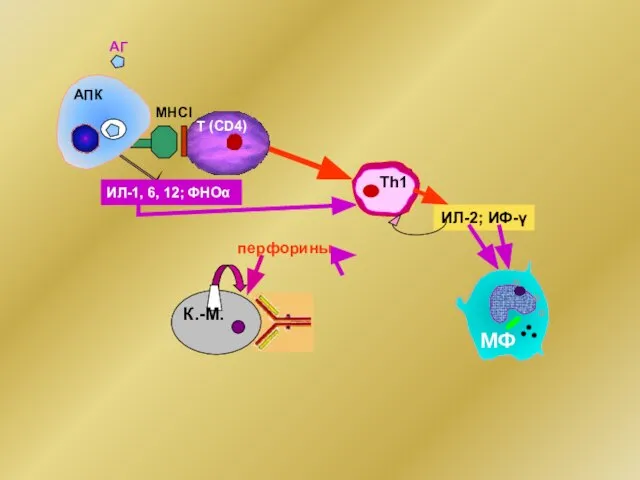
МНСI
АГ
ИЛ-1, 6, 12; ФНОα
ИЛ-2; ИФ-γ
АПК
Т (CD4)
Тh1
перфорины
К.-М.
МФ
МНСI
АГ
ИЛ-1, 6, 12; ФНОα
ИЛ-2; ИФ-γ
АПК
Т (CD4)
Тh1
перфорины
К.-М.
МФ
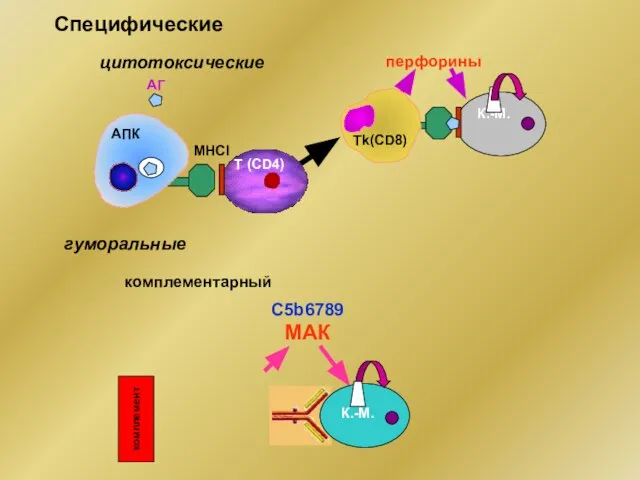
Специфические
МНСI
АГ
К.-М.
АПК
Т (CD4)
Тk(CD8)
перфорины
цитотоксические
гуморальные
C5b6789 МАК
комплементарный
К.-М.
комплемент
Специфические
МНСI
АГ
К.-М.
АПК
Т (CD4)
Тk(CD8)
перфорины
цитотоксические
гуморальные
C5b6789 МАК
комплементарный
К.-М.
комплемент

ВЛИЯНИЕ ОПУХОЛИ НА ОРГАНИЗМ
Местные
обусловлены характером роста опухоли
экспансивный рост
ВЛИЯНИЕ ОПУХОЛИ НА ОРГАНИЗМ
Местные
обусловлены характером роста опухоли
экспансивный рост

инфильтративный рост
аппозиционный рост
происходит за счет неопластической трансформации нормальных клеток
инфильтративный рост
аппозиционный рост
происходит за счет неопластической трансформации нормальных клеток
экзофитный рост
эндофитный рост
экзофитный рост
эндофитный рост
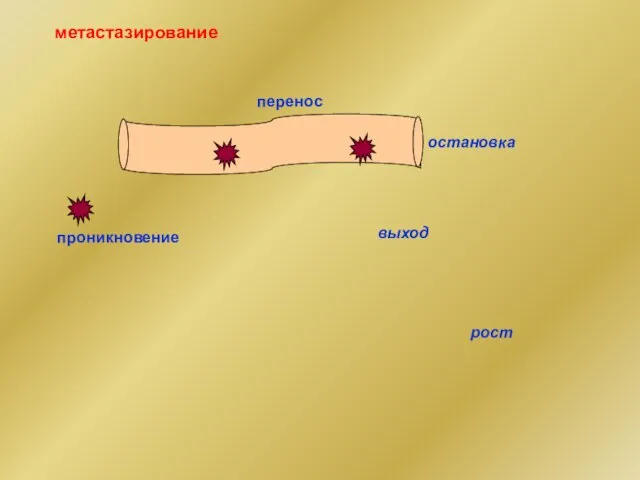
метастазирование
проникновение
перенос
остановка
выход
рост
метастазирование
проникновение
перенос
остановка
выход
рост

ПАРАНЕОПЛАСТИЧЕСКИЙ СИНДРОМ
дистанционное (системное) воздействие новообразования, не связанное с прямой инвазией
ПАРАНЕОПЛАСТИЧЕСКИЙ СИНДРОМ
дистанционное (системное) воздействие новообразования, не связанное с прямой инвазией
 Фармакогностическое изучение черёмухи обыкновенной
Фармакогностическое изучение черёмухи обыкновенной Предлежание плаценты. ПОНРП
Предлежание плаценты. ПОНРП Пропедевтика. Расспрос и осмотр больных с заболеваниями печени и желчевыводящих путей. Перкуссия и пальпация печени и селезенки
Пропедевтика. Расспрос и осмотр больных с заболеваниями печени и желчевыводящих путей. Перкуссия и пальпация печени и селезенки Общественное здоровье и здравоохранение как наука, предмет преподавания и область практической деятельности
Общественное здоровье и здравоохранение как наука, предмет преподавания и область практической деятельности Cиндром красного глаза
Cиндром красного глаза Ультразвук и его применение в медицине
Ультразвук и его применение в медицине Стилистика PR-жанров
Стилистика PR-жанров Профилактика некариозных поражений: флоороза, гипоплазия эмали. Факторы риска возникновения флюороза
Профилактика некариозных поражений: флоороза, гипоплазия эмали. Факторы риска возникновения флюороза Система здравоохранения в странах СНГ
Система здравоохранения в странах СНГ ВИЧ и СПИД
ВИЧ и СПИД Ревизия органов брюшной полости при ранениях и повреждениях
Ревизия органов брюшной полости при ранениях и повреждениях GCP стандарт. Инновационный результат
GCP стандарт. Инновационный результат Теория "зеркального Я" Чарльза Кули
Теория "зеркального Я" Чарльза Кули Диагностика во фтизиатрии
Диагностика во фтизиатрии Оппортунистические инфекции
Оппортунистические инфекции Диаскинтест. Показания
Диаскинтест. Показания Основы ультразвуковой диагностики
Основы ультразвуковой диагностики Рак предстательной железы
Рак предстательной железы Проблема туберкулеза в акушерско - гинекологической практике
Проблема туберкулеза в акушерско - гинекологической практике Профилактическая медицина. Концепция здоровья. Сестринские технологии в профилактической медицине
Профилактическая медицина. Концепция здоровья. Сестринские технологии в профилактической медицине Гипербилирубинемии у новорожденных. Классификация. Дифференциальная диагностика. Клиника, лечение. профилактика
Гипербилирубинемии у новорожденных. Классификация. Дифференциальная диагностика. Клиника, лечение. профилактика Заболевание органов пищеварения
Заболевание органов пищеварения Проект механизма управления здравоохранением Челябинской области
Проект механизма управления здравоохранением Челябинской области Асқазанның жара ауруын зерттеу әдістері және сәулелік семиотикасы
Асқазанның жара ауруын зерттеу әдістері және сәулелік семиотикасы Акушерлік стационардағы санитарлық – гигиеналық режимнің ерекшеліктері
Акушерлік стационардағы санитарлық – гигиеналық режимнің ерекшеліктері Этапы хирургического лечения
Этапы хирургического лечения Слизистая оболочка полости рта
Слизистая оболочка полости рта Система подхода к обеспечению сестринского ухода за пациентом с сахарным диабетом, осложненным диабетической стопой
Система подхода к обеспечению сестринского ухода за пациентом с сахарным диабетом, осложненным диабетической стопой