- Клеточная мембрана

Содержание
- 2. Цитоплазматическая мембрана - физический полупроницаемый барьер, отграничивающий внутриклеточное пространство от внеклеточ-ной среды. Мембрана обеспечивает сохране-ние различий
- 3. Функции цитоплазматической мембраны 1. Барьерная функция; 2. Структурная функция - придание опреде- ленной формы клеткам; 3.
- 4. 7. Обозначение генетической идентичности данной клетки данному организму (поверх- ностные гликопротеины и полисахариды). Согласованная работа всех
- 5. Биомембрана - фосфолипидный бислой. Её основные составляющие: липиды и белки (% соотношение липиды : белки плазматиче-ской
- 6. Липидный компонент плазматических мембран
- 7. Три главных типа липидов плазматических мембран 1. ФОСФОЛИПИДЫ 1.А. ГЛИЦЕРОФОСФОЛИПИДЫ: Главные: ФХ, ФС, ФЭА Минорный: ФИ
- 8. Образование диацилглицерола и инозитолтрифосфата с участием фосфолипазы С Х
- 9. 1.Б. СФИНГОФОСФОЛИПИДЫ: главный представи- тель сфингомиелин. Располагается во внешнем монослое плазматической мембраны. Сфингомиелин и его метаболиты
- 10. 2. ГЛИКОЛИПИДЫ ЦЕРЕБРОЗИДЫ = гидрофобная часть молекулы: цера- мид + углеводная часть молекулы: моно- или олиго-сахаридный
- 11. Холестерол распределен между листками бислоя сравнительно равномерно. Его моле-кулы располагаются между остатками жир-ных кислот фосфолипидов. Холестерол,
- 12. Жидкостно-мозаичная (fluid mosaic) модель плазматической мембраны Сингера и Николсона (1972):
- 13. По Сингеру и Николсону, мембрана это: - Фосфолипидный бислой, образующий замкнутую сферу. В бислое «плавают» или
- 14. В настоящее время модель мембраны по Сингеру и Николсону существует с дополнени-ями Симонса и Ван Меера
- 15. Поперечная асимметрия липидного бислоя возможна благодаря селективным энергоза-висимым переносчикам липидов. К их числу относится семейство белков
- 16. Б. Флоппазы. Перемещают ФЛ в обратном направлении – из внутреннего монослоя во внешний. В. Скрэмблазы. Переносят
- 17. Катализируемый трансмембранный перенос
- 18. При физиологической То диффузия молекул липидов из одного монослоя в другой («флип-флоп»), посредством некатализируемой диф-фузии –
- 19. Некатализируемая трансмембраная диффузия
- 20. 2. Рафты и сигнальные платформы. Представления о рафтах в липидной фазе цитоплаз-матических мембран были сформированы Симонсом,
- 21. В составе рафтов типично присутствие рецепторов, обладающих собственной тирозин-киназной активно-стью и других белков, участвующих в передаче
- 22. В пределах образующейся сигнальной платформы в течение нескольких секунд происходит кластеризация рецепторов, что является эффективным способом
- 23. Образование сигнальных платформ из предсуществующих рафтов и кластери- зация рецепторов
- 24. Слияние рафтов с образованием сигнальных платформ и кластеризация рецепторов
- 25. КАВЕОЛЫ – разновидность рафтов Кавеола описана в 1955 г. - «сaveolae intracellulare» или просто «сaveolae». Это
- 26. Благодаря кавеолину (фор-мирование димеров) участок мембраны изгибается – роль в образовании инвагинации мембраны. ФУНКЦИИ КАВЕОЛ 1.
- 27. Белковый компонент плазматических мембран
- 28. В большинстве цитоплазматических мембран на долю липидов и белков приходится около 50% по массе. На долю
- 29. 2. Транспортная: формируют различные каналы (для диффузии молекул через мембрану), ионные насосы и специфические переносчики; 3.
- 30. Согласно Сингеру и Николсону, белки цитоплазмати-ческой мембраны делят на периферические, интег-ральные и амфитропные. 1. Периферические (поверхностные)
- 31. С помощью такого якоря на поверхности лимфо-цитов крепится белок Thy-1. Внеклеточное пространство Гликозилфосфатидилинозитольный якорь
- 32. б). Белки внутренней стороны мембраны: - ассоциированы с цитоскелетом, участвуют в форми- ровании силуэта клетки; Белок
- 33. цитоплазма Прикрепление белков Ras и Src к внутреннему монослою липидов цитоплазматической мембраны Ras и Src участвуют
- 34. 2. Интегральные (трансмембранные) белки. Белки имеют внеклеточный, внутриклеточный и трансмембранный домены. Трансмембранный домен гидрофобен и может
- 35. Трансмембранный гидрофобный домен интеграль-ного белка часто является α-спиралью. α-спиральные последовательности, каждая из кото-рых образует трансмебранный сегмент,
- 36. 3. Амфитропные белки. Могут находится в цитоплазме и обратимо связы-ваться с мембраной. Способы связывания с мембраной:
- 37. О латеральной диффузии мембранных белков Латеральная диффузия мембранных белков открыта L. Frye и M. Edidin в
- 38. Трансмембранная передача сигналов
- 39. Основополагающее свойство клетки, обеспечиваю-щее её полноценное функционирование – способ-ность получать сигналы из окружающей её среды (за
- 40. Сигнальная молекула (лиганд любой природы), связывается с рецептором слабой нековалентной связью: водородные связи, гидрофобные и электро-статические
- 41. Основные свойства систем, передающих внешний сигнал Специфичность передачи сигналов. Обеспечивается молекулярной комплементарно- стью сигнальной молекулы (первичного
- 42. 2. Высокая чувствительность молекул-посредников в передаче (трансдукции) сигнала. Она обеспечивается: а). Высоким сродством рецепторов к сигнальным
- 43. Особо о десенситизации рецептора. Это снижение (потеря) чувствительности рецептора к продолжительно действующей сигнальной молеку-ле. Чувствительность восстанавливается,
- 45. Скачать презентацию

Цитоплазматическая мембрана - физический полупроницаемый барьер, отграничивающий внутриклеточное пространство от
Цитоплазматическая мембрана - физический полупроницаемый барьер, отграничивающий внутриклеточное пространство от

Функции цитоплазматической мембраны
1. Барьерная функция;
2. Структурная функция - придание опреде-
Функции цитоплазматической мембраны
1. Барьерная функция;
2. Структурная функция - придание опреде-

7. Обозначение генетической идентичности
данной клетки данному организму (поверх-
ностные гликопротеины
7. Обозначение генетической идентичности
данной клетки данному организму (поверх-
ностные гликопротеины

Биомембрана - фосфолипидный бислой.
Её основные составляющие: липиды и белки
Биомембрана - фосфолипидный бислой.
Её основные составляющие: липиды и белки

Липидный компонент плазматических мембран
Липидный компонент плазматических мембран

Три главных типа липидов плазматических
мембран
1. ФОСФОЛИПИДЫ
1.А. ГЛИЦЕРОФОСФОЛИПИДЫ:
Главные: ФХ, ФС,
Три главных типа липидов плазматических
мембран
1. ФОСФОЛИПИДЫ
1.А. ГЛИЦЕРОФОСФОЛИПИДЫ:
Главные: ФХ, ФС,
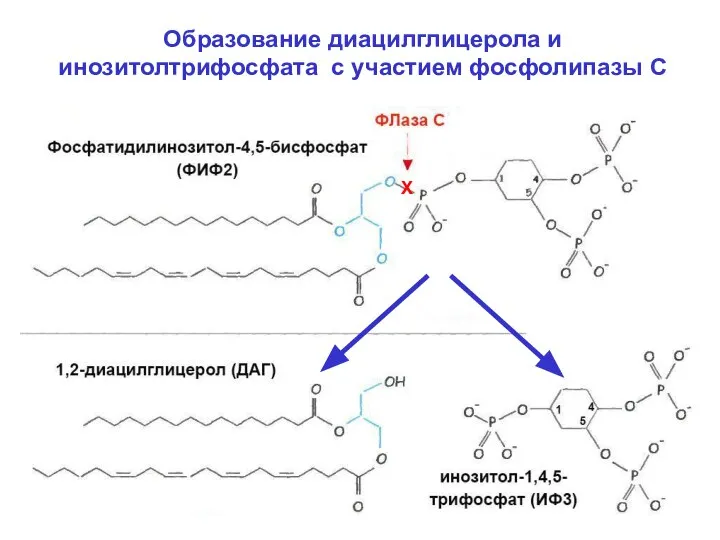
Образование диацилглицерола и инозитолтрифосфата с участием фосфолипазы С
Х
Образование диацилглицерола и инозитолтрифосфата с участием фосфолипазы С
Х
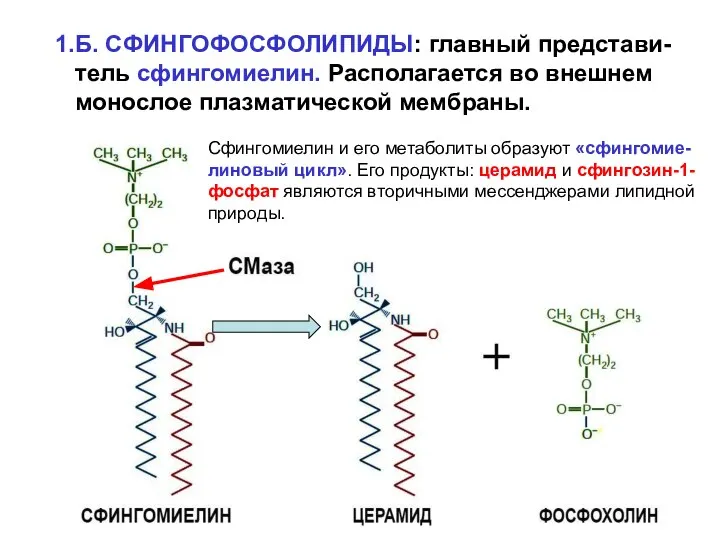
1.Б. СФИНГОФОСФОЛИПИДЫ: главный представи-
тель сфингомиелин. Располагается во внешнем
монослое плазматической
1.Б. СФИНГОФОСФОЛИПИДЫ: главный представи-
тель сфингомиелин. Располагается во внешнем
монослое плазматической

2. ГЛИКОЛИПИДЫ
ЦЕРЕБРОЗИДЫ = гидрофобная часть молекулы: цера-
мид + углеводная часть
2. ГЛИКОЛИПИДЫ
ЦЕРЕБРОЗИДЫ = гидрофобная часть молекулы: цера-
мид + углеводная часть

Холестерол распределен между листками бислоя сравнительно равномерно. Его моле-кулы располагаются
Холестерол распределен между листками бислоя сравнительно равномерно. Его моле-кулы располагаются
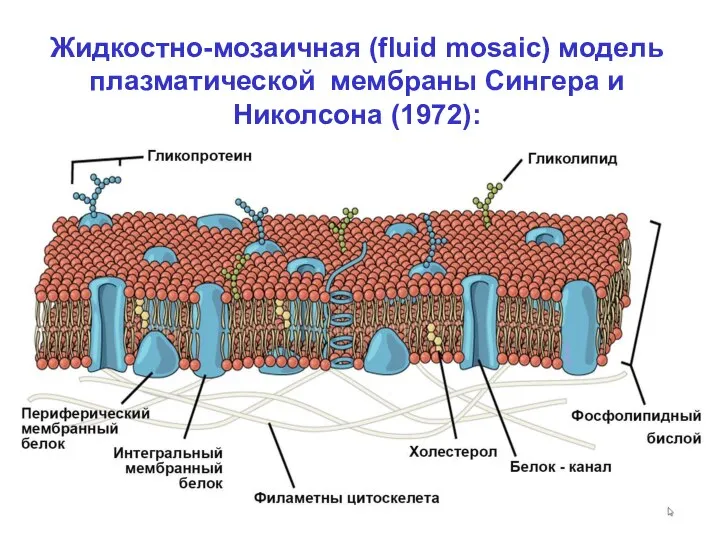
Жидкостно-мозаичная (fluid mosaic) модель
плазматической мембраны Сингера и
Николсона (1972):
Жидкостно-мозаичная (fluid mosaic) модель
плазматической мембраны Сингера и
Николсона (1972):

По Сингеру и Николсону, мембрана это:
- Фосфолипидный бислой, образующий
По Сингеру и Николсону, мембрана это:
- Фосфолипидный бислой, образующий
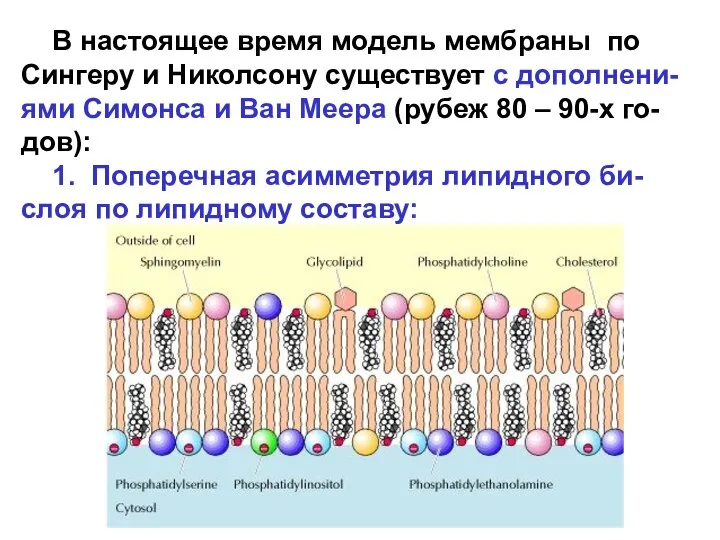
В настоящее время модель мембраны по Сингеру и Николсону существует
В настоящее время модель мембраны по Сингеру и Николсону существует

Поперечная асимметрия липидного бислоя возможна благодаря селективным энергоза-висимым переносчикам липидов.
Поперечная асимметрия липидного бислоя возможна благодаря селективным энергоза-висимым переносчикам липидов.

Б. Флоппазы. Перемещают ФЛ в обратном направлении – из внутреннего
Б. Флоппазы. Перемещают ФЛ в обратном направлении – из внутреннего
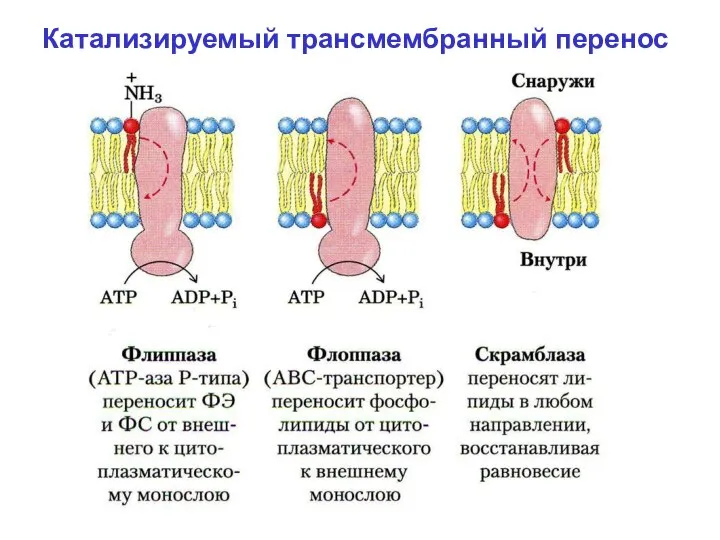
Катализируемый трансмембранный перенос
Катализируемый трансмембранный перенос

При физиологической То диффузия молекул липидов из одного монослоя
При физиологической То диффузия молекул липидов из одного монослоя
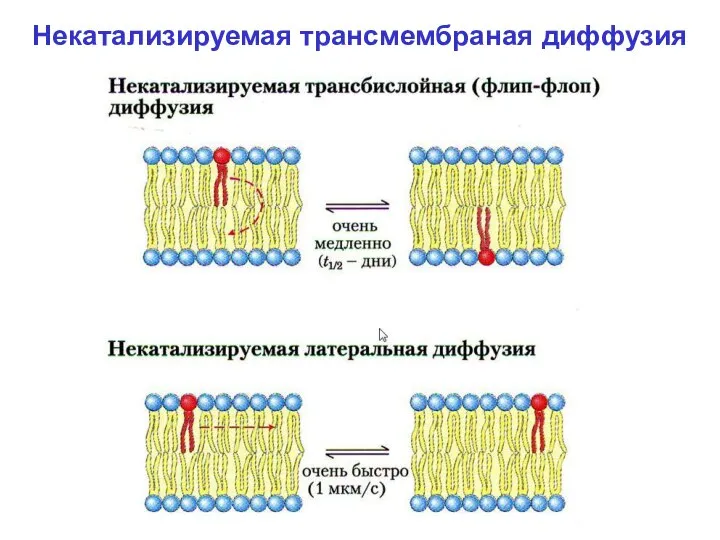
Некатализируемая трансмембраная диффузия
Некатализируемая трансмембраная диффузия

2. Рафты и сигнальные платформы.
Представления о рафтах в
2. Рафты и сигнальные платформы.
Представления о рафтах в

В составе рафтов типично присутствие рецепторов, обладающих собственной тирозин-киназной активно-стью
В составе рафтов типично присутствие рецепторов, обладающих собственной тирозин-киназной активно-стью

В пределах образующейся сигнальной платформы в течение нескольких секунд происходит
В пределах образующейся сигнальной платформы в течение нескольких секунд происходит
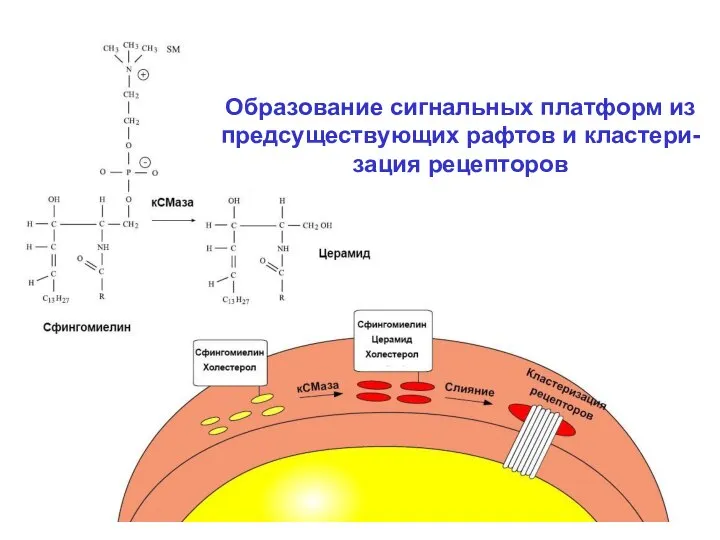
Образование сигнальных платформ из
предсуществующих рафтов и кластери-
зация рецепторов
Образование сигнальных платформ из
предсуществующих рафтов и кластери-
зация рецепторов
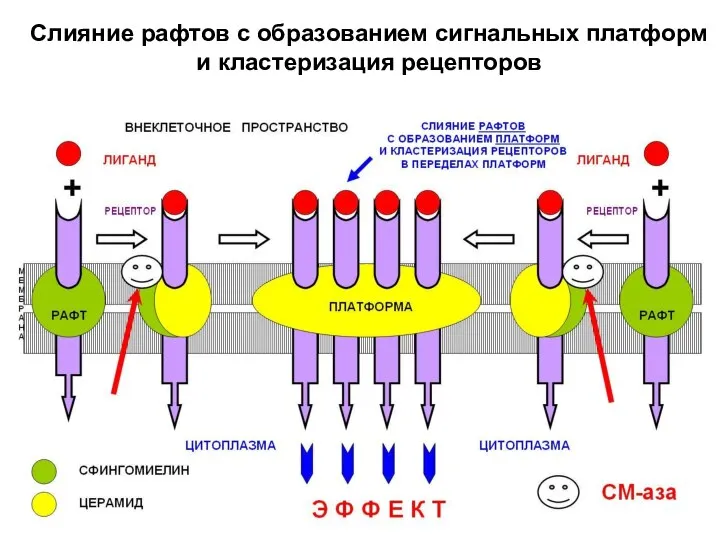
Слияние рафтов с образованием сигнальных платформ
и кластеризация рецепторов
Слияние рафтов с образованием сигнальных платформ
и кластеризация рецепторов

КАВЕОЛЫ – разновидность рафтов
Кавеола описана в 1955 г. - «сaveolae
КАВЕОЛЫ – разновидность рафтов
Кавеола описана в 1955 г. - «сaveolae
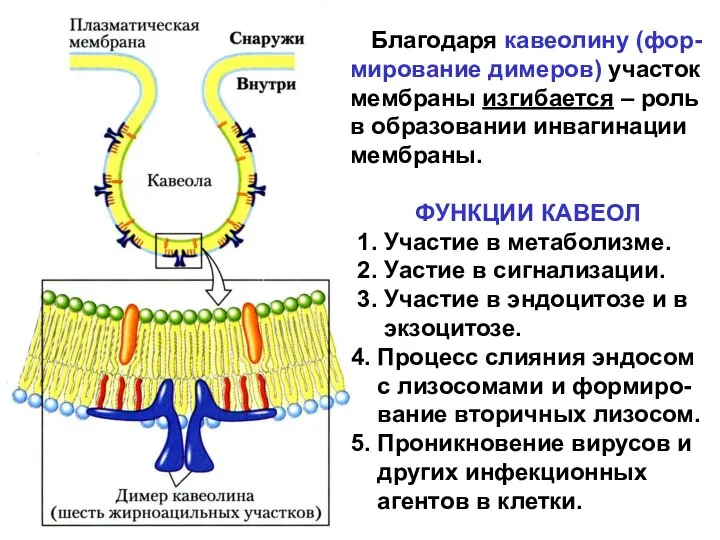
Благодаря кавеолину (фор-мирование димеров) участок мембраны изгибается – роль в
Благодаря кавеолину (фор-мирование димеров) участок мембраны изгибается – роль в

Белковый компонент плазматических мембран
Белковый компонент плазматических мембран

В большинстве цитоплазматических мембран на долю липидов и белков приходится
В большинстве цитоплазматических мембран на долю липидов и белков приходится

2. Транспортная: формируют различные каналы (для диффузии молекул через мембрану), ионные
2. Транспортная: формируют различные каналы (для диффузии молекул через мембрану), ионные

Согласно Сингеру и Николсону, белки цитоплазмати-ческой мембраны делят на периферические,
Согласно Сингеру и Николсону, белки цитоплазмати-ческой мембраны делят на периферические,
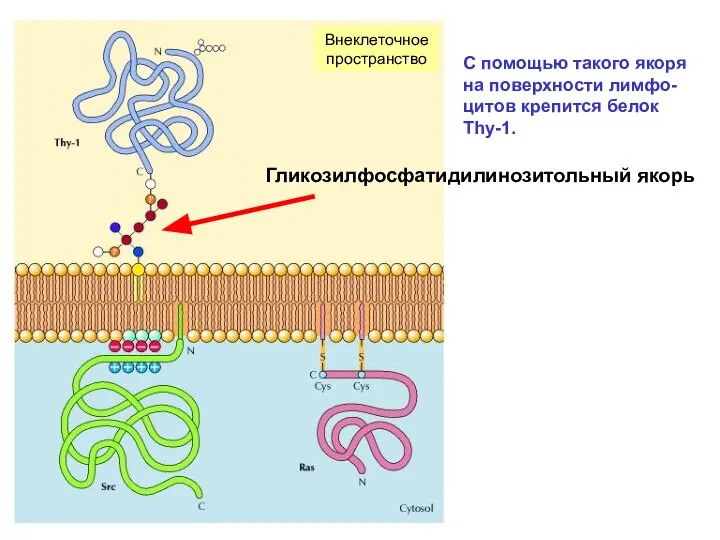
С помощью такого якоря на поверхности лимфо-цитов крепится белок Thy-1.
Внеклеточное
С помощью такого якоря на поверхности лимфо-цитов крепится белок Thy-1.
Внеклеточное

б). Белки внутренней стороны мембраны:
- ассоциированы с цитоскелетом, участвуют в
б). Белки внутренней стороны мембраны:
- ассоциированы с цитоскелетом, участвуют в
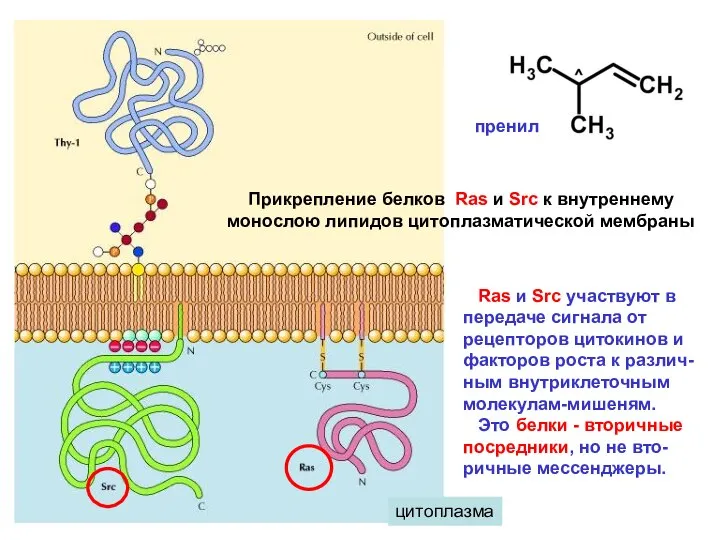
цитоплазма
Прикрепление белков Ras и Src к внутреннему
монослою липидов цитоплазматической мембраны
Ras
цитоплазма
Прикрепление белков Ras и Src к внутреннему
монослою липидов цитоплазматической мембраны
Ras
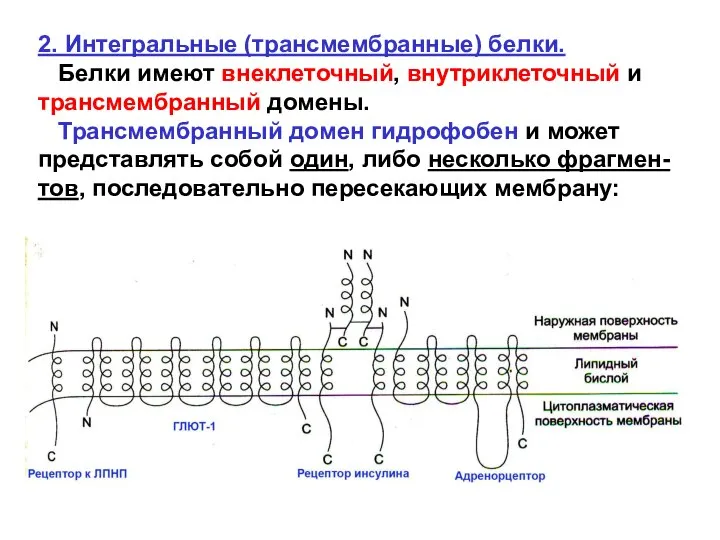
2. Интегральные (трансмембранные) белки.
Белки имеют внеклеточный, внутриклеточный и трансмембранный домены.
2. Интегральные (трансмембранные) белки.
Белки имеют внеклеточный, внутриклеточный и трансмембранный домены.

Трансмембранный гидрофобный домен интеграль-ного белка часто является α-спиралью.
α-спиральные последовательности,
Трансмембранный гидрофобный домен интеграль-ного белка часто является α-спиралью.
α-спиральные последовательности,

3. Амфитропные белки.
Могут находится в цитоплазме и обратимо связы-ваться с
3. Амфитропные белки.
Могут находится в цитоплазме и обратимо связы-ваться с

О латеральной диффузии мембранных белков
Латеральная диффузия мембранных белков открыта L.
О латеральной диффузии мембранных белков
Латеральная диффузия мембранных белков открыта L.

Трансмембранная передача сигналов
Трансмембранная передача сигналов

Основополагающее свойство клетки, обеспечиваю-щее её полноценное функционирование – способ-ность получать
Основополагающее свойство клетки, обеспечиваю-щее её полноценное функционирование – способ-ность получать

Сигнальная молекула (лиганд любой природы), связывается с рецептором слабой нековалентной связью:
Сигнальная молекула (лиганд любой природы), связывается с рецептором слабой нековалентной связью:

Основные свойства систем, передающих внешний
сигнал
Специфичность передачи сигналов.
Обеспечивается молекулярной
Основные свойства систем, передающих внешний
сигнал
Специфичность передачи сигналов.
Обеспечивается молекулярной

2. Высокая чувствительность молекул-посредников в
передаче (трансдукции) сигнала.
Она обеспечивается:
а).
2. Высокая чувствительность молекул-посредников в
передаче (трансдукции) сигнала.
Она обеспечивается:
а).

Особо о десенситизации рецептора.
Это снижение (потеря) чувствительности рецептора
Особо о десенситизации рецептора.
Это снижение (потеря) чувствительности рецептора
 Учебный план 1 класса на 2010-2011учебный год ГОУ СОШ с УИОП г.Белой Холуницы Кировской области
Учебный план 1 класса на 2010-2011учебный год ГОУ СОШ с УИОП г.Белой Холуницы Кировской области Психология восприятия 2
Психология восприятия 2 Презентация на тему "Велосипедист - Водитель транспортного средства" - скачать презентации по Педагогике
Презентация на тему "Велосипедист - Водитель транспортного средства" - скачать презентации по Педагогике ОБРАЗОВАНИЕ ДЕТЕЙ С НАРУШЕНИЯМИ ЗРЕНИЯ
ОБРАЗОВАНИЕ ДЕТЕЙ С НАРУШЕНИЯМИ ЗРЕНИЯ Понятие административно-правовых отношений, их структура
Понятие административно-правовых отношений, их структура Основные свойства темперамента
Основные свойства темперамента  Николай Михайлович Пржевальский
Николай Михайлович Пржевальский Содержание права собственности на землю Выполнили студентки группы Ю-103 Пустовая Анастасия и Мятлик Елена.
Содержание права собственности на землю Выполнили студентки группы Ю-103 Пустовая Анастасия и Мятлик Елена. Словарные слова - презентация для начальной школы
Словарные слова - презентация для начальной школы Генерация идей. Оценка возможностей. Работа с бизнес-гипотезой (идеей)
Генерация идей. Оценка возможностей. Работа с бизнес-гипотезой (идеей) Поняття циклу. Оператори циклу. Виклик функції
Поняття циклу. Оператори циклу. Виклик функції . Вступление в венерологию, сифилис (история)
. Вступление в венерологию, сифилис (история) Типовые примеры сборки узлов и панелей клепанной конструкции
Типовые примеры сборки узлов и панелей клепанной конструкции Презентация "Выборы в моей жизни: от теории к практике" - скачать презентации по Экономике
Презентация "Выборы в моей жизни: от теории к практике" - скачать презентации по Экономике Практикум № 1. Точка, прямая, плоскость на комплексном чертеже
Практикум № 1. Точка, прямая, плоскость на комплексном чертеже Микропроцессорные системы
Микропроцессорные системы Комп`ютер у моєму житті
Комп`ютер у моєму житті печень
печень Қылмыстық жауапкершіліктен және жазадан босату
Қылмыстық жауапкершіліктен және жазадан босату Презентация Интерактивная карта Северной Америки
Презентация Интерактивная карта Северной Америки Селектор адреса
Селектор адреса Религия и культура: конфликты и взаимодействие в современных условиях (на примере г. Москвы)
Религия и культура: конфликты и взаимодействие в современных условиях (на примере г. Москвы) Понятие правовой статистики как отрасли социальной статистики
Понятие правовой статистики как отрасли социальной статистики Аттестационная работа. Исследование произведений А.С.Грина, выявление значимых для писателя мест в г. Севастополе
Аттестационная работа. Исследование произведений А.С.Грина, выявление значимых для писателя мест в г. Севастополе Викторина «Стили в архитектуре»
Викторина «Стили в архитектуре» АТРЕЗИЯ ЛЕГОЧНОЙ АРТЕРИИ
АТРЕЗИЯ ЛЕГОЧНОЙ АРТЕРИИ Оценивание в процессе изучения общественных дисциплин
Оценивание в процессе изучения общественных дисциплин ТОГУ «Уваровская специальная (коррекционная) общеобразовательная школа-интернат» Индивидуальное коррекционное занятие Учите
ТОГУ «Уваровская специальная (коррекционная) общеобразовательная школа-интернат» Индивидуальное коррекционное занятие Учите