- Презентация на тему "Регуляторные системы клетки" - скачать презентации по Медицине

Содержание
- 2. В процессе жизнедеятельности клетка постоянно получает различные сигналы из вне: в результате чего она изменяет свой
- 3. В клетке существует довольно ограниченное количество таких универсальных регуляторов. К ним относятся: циклические нуклеотиды - цАМФ,
- 4. Широко распространенной и наиболее универсальной регуляторной системой является система, связанная с Са²+ Функции Са2+ в клетке:
- 5. Общая схема работы Са2+ в клетке В состоянии покоя концентрация Са²⁺ в клетке всего 10 ¯7
- 6. Пути поступления Са2+ в клетку I Прямой Хемочуствительные каналы. Открываются в результате специфического взаимодействия поверхностных рецепторов
- 7. IP3-путь
- 8. Пути удаления Са2+ из цитоплазмы клетки Системы, удаляющие Са2+ из клетки встроены в мембраны. Эти каналы
- 9. Работа Са²⁺-АТФ-азы Связывание 2-х молекул Ca2+ с центром связывания Ca – АТФ – азы. Связывание одной
- 10. Са-связывающие белки СаСБ имеют в своей структуре специальные участки связывания с Са2+. В зависимости от того,
- 11. Роль кальмодулина в клетке Кальмодулин широко распространен и встречается в клетках животных, растений и грибов. Взаимодействуя
- 12. Гормональная регуляция содержания Са2+ в организме Гормональная регуляция обмена Са2+ в организме осуществляется гормоном щитовидной железы
- 14. Скачать презентацию

В процессе жизнедеятельности клетка постоянно получает различные сигналы из вне:
В процессе жизнедеятельности клетка постоянно получает различные сигналы из вне:

В клетке существует довольно ограниченное количество таких универсальных регуляторов. К
В клетке существует довольно ограниченное количество таких универсальных регуляторов. К

Широко распространенной и наиболее универсальной регуляторной системой является система, связанная с
Широко распространенной и наиболее универсальной регуляторной системой является система, связанная с

Общая схема работы Са2+ в клетке
В состоянии покоя концентрация Са²⁺ в
Общая схема работы Са2+ в клетке
В состоянии покоя концентрация Са²⁺ в

Пути поступления Са2+ в клетку
I Прямой
Хемочуствительные каналы. Открываются в результате специфического
Пути поступления Са2+ в клетку
I Прямой
Хемочуствительные каналы. Открываются в результате специфического
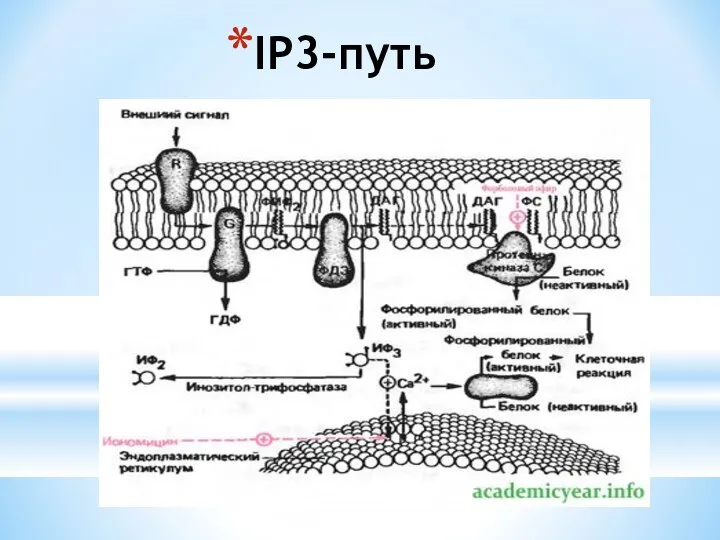
IP3-путь
IP3-путь

Пути удаления Са2+ из цитоплазмы клетки
Системы, удаляющие Са2+ из клетки
Пути удаления Са2+ из цитоплазмы клетки
Системы, удаляющие Са2+ из клетки

Работа Са²⁺-АТФ-азы
Связывание 2-х молекул Ca2+ с центром связывания Ca – АТФ
Работа Са²⁺-АТФ-азы
Связывание 2-х молекул Ca2+ с центром связывания Ca – АТФ

Са-связывающие белки
СаСБ имеют в своей структуре специальные участки связывания с Са2+.
Са-связывающие белки
СаСБ имеют в своей структуре специальные участки связывания с Са2+.

Роль кальмодулина в клетке
Кальмодулин широко распространен и встречается в клетках животных,
Роль кальмодулина в клетке
Кальмодулин широко распространен и встречается в клетках животных,

Гормональная регуляция содержания Са2+ в организме
Гормональная регуляция обмена Са2+ в
Гормональная регуляция содержания Са2+ в организме
Гормональная регуляция обмена Са2+ в
 Мониторинг рН желудка и пищевода в оценке антисекреторного действия лекарственных средств к.мед.н. Просоленко К.А. Институт терап
Мониторинг рН желудка и пищевода в оценке антисекреторного действия лекарственных средств к.мед.н. Просоленко К.А. Институт терап Глава 3 Спрос, предложение и рыночное равновесие
Глава 3 Спрос, предложение и рыночное равновесие  Роль музыки в жизни человека Авторы: Яровая Анна Тишенина Влада ученицы 10 «А» класса МОУСОШ №7 Руководитель проекта: Шматько На
Роль музыки в жизни человека Авторы: Яровая Анна Тишенина Влада ученицы 10 «А» класса МОУСОШ №7 Руководитель проекта: Шматько На Агбис
Агбис Внешнее трение
Внешнее трение ТренажёрСложение в пределах 20 - презентация для начальной школы
ТренажёрСложение в пределах 20 - презентация для начальной школы Рубежно – зачетная работа по системам компьютерной графики
Рубежно – зачетная работа по системам компьютерной графики  Қазақ кәдесілер
Қазақ кәдесілер Микросхемы с программируемой структурой
Микросхемы с программируемой структурой Технология совершенствования двигательных действий лыжников-спринтеров
Технология совершенствования двигательных действий лыжников-спринтеров Основные инструменты формирования служебно-профессиональной морали и нравственности культуры государственных гражданских служ
Основные инструменты формирования служебно-профессиональной морали и нравственности культуры государственных гражданских служ Правописание -ЧИК - ЩИК (с упражнениями)
Правописание -ЧИК - ЩИК (с упражнениями)  Мотострелковый (танковый) взвод в различный видах боя
Мотострелковый (танковый) взвод в различный видах боя Архитектурные достижения Древнего Рима
Архитектурные достижения Древнего Рима Андрей Николаев Художник, иллюстрирующий всемирно известное произведение литературы, должен быть готов умереть в писателе.
Андрей Николаев Художник, иллюстрирующий всемирно известное произведение литературы, должен быть готов умереть в писателе. Цифровые видеокамеры
Цифровые видеокамеры Презентация Аутсорсинг
Презентация Аутсорсинг Урок в 7 классе по теме: Решение задач с помощью уравнений. Учитель: Барышенская Елена Ивановна
Урок в 7 классе по теме: Решение задач с помощью уравнений. Учитель: Барышенская Елена Ивановна  Глобальные проблемы человечества
Глобальные проблемы человечества Функциональные элементы САР ЭПС: исполнительные и управляющие элементы
Функциональные элементы САР ЭПС: исполнительные и управляющие элементы Білорусь та її національні страви
Білорусь та її національні страви Тайна одной семьи: на примере жизненного пути Салихова Губайдуллы Зарифовича
Тайна одной семьи: на примере жизненного пути Салихова Губайдуллы Зарифовича Как пишутся иконы
Как пишутся иконы Качество жизни и качество медицинской помощи по мнению населения геронтологической группы Туребеков Думан Кажибаевич к.м.н., д
Качество жизни и качество медицинской помощи по мнению населения геронтологической группы Туребеков Думан Кажибаевич к.м.н., д Презентация "Модели жизненных циклов организации" - скачать презентации по Экономике
Презентация "Модели жизненных циклов организации" - скачать презентации по Экономике Программирование на языке Си. Переменные. Ввод и вывод. Ветвления. Сложные условия. Циклы с условием
Программирование на языке Си. Переменные. Ввод и вывод. Ветвления. Сложные условия. Циклы с условием Хуже грозного царя только междуцарствие - презентация для начальной школы_
Хуже грозного царя только междуцарствие - презентация для начальной школы_ Задачи в стихах Часть 1
Задачи в стихах Часть 1