- Деление клетки

Содержание
- 2. Схема 1. Фазы митоза
- 3. 0.Интерфаза - G1 После деления - общее содержание белков и РНК вдвое меньше, чем в исходной
- 4. S-период удвоение количества ДНК на ядро удваивается число хромосом. В разных клетках, находящихся в S-периоде, можно
- 5. Постсинтетическая (G2) фаза называется также премитотической. синтез иРНК, синтезируется рРНК. Среди белков - тубулины - белки
- 6. В растущих тканях растений и животных всегда есть клетки, которые находятся как бы вне цикла. Такие
- 7. Митоз - кариокинез, или непрямое деление универсальный способ деления любых эукариотических клеток конденсированные и уже редуплицированные
- 8. Морфология митотических хромосом Фибриллы хроматина в митотической хромосоме образуют многочисленные розетковидные петлевые домены (хромомеры), которые при
- 10. Морфология митотических хромосом Хромосомы представляют собой палочковидные структуры разной длины с довольно постоянной толщиной Плечи хромосом
- 11. Морфология митотических хромосом В области первичной перетяжки расположен кинетохор - сложная белковая структура, имеющая форму овальной
- 12. Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из концов хромосомы и отделяющие маленький
- 14. Динамика митоза 1.Профаза Двойственность хромосом в начинающей делиться клетке - 4 n Конденсации хромосом исчезновение ядрышек
- 15. 1.Профаза уменьшение количества гранулярного ЭПС, распадается на цистерны и вакуоли. количество рибосом падает (до 25 %)
- 16. 1.Профаза центриоли начинают расхождение - полюса веретена. К каждому полюсу - двойная центриоль, диплосома. начинают формироваться
- 17. Метафаза заканчивается образование веретена деления хромосомы выстраиваются в экваториальной плоскости веретена - метафазная пластинка хромосом, или
- 18. 3.Анафаза Хромосомы одновременно теряют связь в области центромер синхронно удаляются к полюсам. обособление двух идентичных наборов
- 19. 4.Телофаза Телофаза начинается с остановки разошедшихся диплоидных (2 п) наборов хромосом (ранняя телофаза) кончается началом реконструкции
- 20. 4.Телофаза В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки - к полюсу, теломерные -
- 21. 4.Телофаза Цитотомия у клеток животных -образование перетяжки, впячивание плазматической мембраны внутрь клетки в кортикальном, подмембранном слое
- 22. Интерфаза Предмейотическая интерфаза отличается от обычной тем, что процесс репликации ДНК не закончен: примерно 0,2...0,4 %
- 23. Первое деление мейоза Сущность - в уменьшение числа хромосом в два раза: из исходной диплоидной клетки
- 25. Профаза 1 стадии: Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в виде клубка тонких
- 26. Зиготена (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом При конъюгации образуются биваленты - комплексы из одной
- 27. Пахитена (стадия толстых нитей). Хромосомы спирализуются. Завершается репликация ДНК Завершается кроссинговер – перекрест хромосом, в результате
- 29. Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных
- 30. Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра. Количество бивалентов – n.
- 31. Метафаза I (метафаза первого деления) Ядерная оболочка разрушается Формируется веретено деления Биваленты перемещаются в экваториальную плоскость
- 32. Анафаза I (анафаза первого деления) Гомологичные хромосомы, (биваленты), разъединяются каждая хромосома движется в сторону ближайшего полюса
- 33. Телофаза I (телофаза первого деления) Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки дочерняя клетка получает
- 34. Интеркинез короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы не происходит репликации ДНК, удвоения хромосом
- 35. Второе деление мейоза (эквационное деление, или мейоз II) В ходе мейоза II уменьшения числа хромосом не
- 36. Профаза II Не отличается существенно от профазы митоза Хромосомы видны в световой микроскоп в виде тонких
- 37. Метафаза II Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток
- 38. Анафаза II Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы перемещаются к полюсам клеток.
- 40. Скачать презентацию
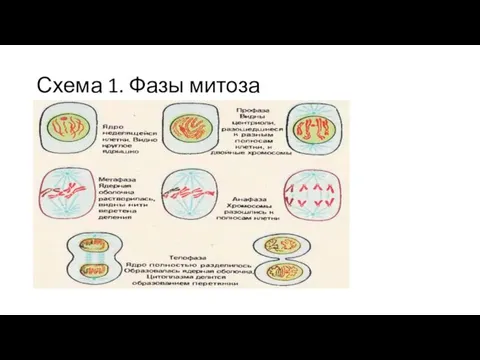
Схема 1. Фазы митоза
Схема 1. Фазы митоза

0.Интерфаза - G1
После деления - общее содержание белков и РНК вдвое
0.Интерфаза - G1
После деления - общее содержание белков и РНК вдвое

S-период
удвоение количества ДНК на ядро
удваивается число хромосом.
В разных клетках,
S-период
удвоение количества ДНК на ядро
удваивается число хромосом.
В разных клетках,

Постсинтетическая (G2) фаза
называется также премитотической.
синтез иРНК,
синтезируется рРНК.
Постсинтетическая (G2) фаза
называется также премитотической.
синтез иРНК,
синтезируется рРНК.

В растущих тканях растений и животных всегда есть клетки, которые находятся
В растущих тканях растений и животных всегда есть клетки, которые находятся

Митоз - кариокинез, или непрямое деление
универсальный способ деления любых эукариотических
Митоз - кариокинез, или непрямое деление
универсальный способ деления любых эукариотических

Морфология митотических хромосом
Фибриллы хроматина в митотической хромосоме образуют многочисленные розетковидные
Морфология митотических хромосом
Фибриллы хроматина в митотической хромосоме образуют многочисленные розетковидные
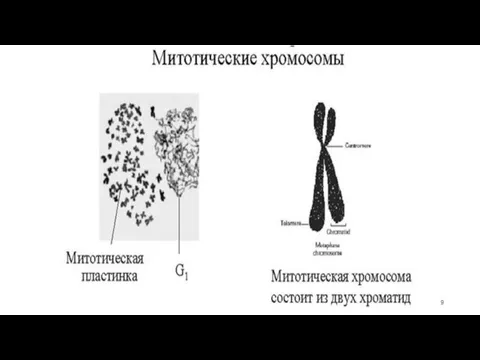

Морфология митотических хромосом
Хромосомы представляют собой палочковидные структуры разной длины с довольно
Морфология митотических хромосом
Хромосомы представляют собой палочковидные структуры разной длины с довольно

Морфология митотических хромосом
В области первичной перетяжки расположен кинетохор - сложная белковая
Морфология митотических хромосом
В области первичной перетяжки расположен кинетохор - сложная белковая

Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из
Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из
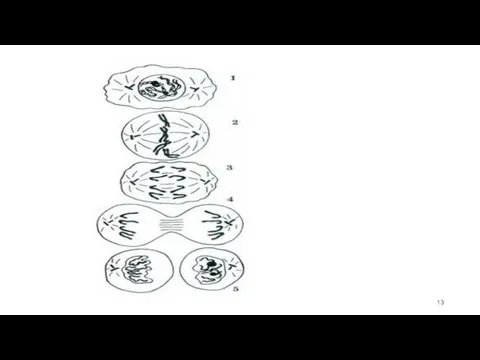

Динамика митоза
1.Профаза
Двойственность хромосом в начинающей делиться клетке - 4 n
Динамика митоза
1.Профаза
Двойственность хромосом в начинающей делиться клетке - 4 n

1.Профаза
уменьшение количества гранулярного ЭПС, распадается на цистерны и вакуоли.
количество рибосом падает
1.Профаза
уменьшение количества гранулярного ЭПС, распадается на цистерны и вакуоли.
количество рибосом падает

1.Профаза
центриоли начинают расхождение - полюса веретена.
К каждому полюсу - двойная
1.Профаза
центриоли начинают расхождение - полюса веретена.
К каждому полюсу - двойная

Метафаза
заканчивается образование веретена деления
хромосомы выстраиваются в экваториальной плоскости веретена
Метафаза
заканчивается образование веретена деления
хромосомы выстраиваются в экваториальной плоскости веретена

3.Анафаза
Хромосомы одновременно теряют связь в области центромер
синхронно удаляются к полюсам.
3.Анафаза
Хромосомы одновременно теряют связь в области центромер
синхронно удаляются к полюсам.

4.Телофаза
Телофаза начинается с остановки разошедшихся диплоидных (2 п) наборов хромосом (ранняя
4.Телофаза
Телофаза начинается с остановки разошедшихся диплоидных (2 п) наборов хромосом (ранняя

4.Телофаза
В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки -
4.Телофаза
В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки -

4.Телофаза
Цитотомия у клеток животных -образование перетяжки, впячивание плазматической мембраны внутрь клетки
4.Телофаза
Цитотомия у клеток животных -образование перетяжки, впячивание плазматической мембраны внутрь клетки

Интерфаза
Предмейотическая интерфаза отличается от обычной тем, что процесс репликации ДНК
Интерфаза
Предмейотическая интерфаза отличается от обычной тем, что процесс репликации ДНК

Первое деление мейоза
Сущность - в уменьшение числа хромосом в два
Первое деление мейоза
Сущность - в уменьшение числа хромосом в два
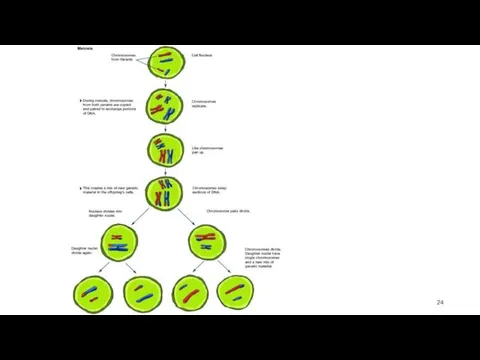

Профаза 1
стадии:
Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в
Профаза 1
стадии:
Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в

Зиготена (стадия сливающихся нитей).
Происходит конъюгация гомологичных хромосом
При конъюгации образуются биваленты -
Зиготена (стадия сливающихся нитей).
Происходит конъюгация гомологичных хромосом
При конъюгации образуются биваленты -

Пахитена (стадия толстых нитей).
Хромосомы спирализуются.
Завершается репликация ДНК
Завершается кроссинговер – перекрест
Пахитена (стадия толстых нитей).
Хромосомы спирализуются.
Завершается репликация ДНК
Завершается кроссинговер – перекрест


Диплотена (стадия двойных нитей).
Гомологичные хромосомы в бивалентах отталкиваются друг от друга.
Диплотена (стадия двойных нитей).
Гомологичные хромосомы в бивалентах отталкиваются друг от друга.

Диакинез (стадия расхождения бивалентов).
Отдельные биваленты располагаются на периферии ядра. Количество бивалентов
Диакинез (стадия расхождения бивалентов).
Отдельные биваленты располагаются на периферии ядра. Количество бивалентов

Метафаза I (метафаза первого деления)
Ядерная оболочка разрушается
Формируется веретено деления
Биваленты перемещаются
Метафаза I (метафаза первого деления)
Ядерная оболочка разрушается
Формируется веретено деления
Биваленты перемещаются

Анафаза I (анафаза первого деления)
Гомологичные хромосомы, (биваленты), разъединяются
каждая хромосома движется в
Анафаза I (анафаза первого деления)
Гомологичные хромосомы, (биваленты), разъединяются
каждая хромосома движется в

Телофаза I (телофаза первого деления)
Гомологичные двухроматидные хромосомы полностью расходятся к полюсам
Телофаза I (телофаза первого деления)
Гомологичные двухроматидные хромосомы полностью расходятся к полюсам

Интеркинез
короткий промежуток между двумя мейотическими делениями.
Отличается от интерфазы
не
Интеркинез
короткий промежуток между двумя мейотическими делениями.
Отличается от интерфазы
не

Второе деление мейоза
(эквационное деление, или мейоз II)
В ходе мейоза II
Второе деление мейоза
(эквационное деление, или мейоз II)
В ходе мейоза II

Профаза II
Не отличается существенно от профазы митоза
Хромосомы видны в световой микроскоп
Профаза II
Не отличается существенно от профазы митоза
Хромосомы видны в световой микроскоп

Метафаза II
Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток
Метафаза II
Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток

Анафаза II
Хромосомы разделяются на хроматиды (как при митозе).
Получившиеся однохроматидные хромосомы
Анафаза II
Хромосомы разделяются на хроматиды (как при митозе).
Получившиеся однохроматидные хромосомы
 Птицеводство. Систематика
Птицеводство. Систематика Морфофункциональные особенности системы пищеварения рыб
Морфофункциональные особенности системы пищеварения рыб Презентация на тему Пища для ума
Презентация на тему Пища для ума Игра ДА-НЕТ
Игра ДА-НЕТ Презентация на тему "Признаки живых организмов" - скачать бесплатно презентации по Биологии
Презентация на тему "Признаки живых организмов" - скачать бесплатно презентации по Биологии Природа и наша безопасность. Тест №40
Природа и наша безопасность. Тест №40 Опорно-двигательная система
Опорно-двигательная система Питание бактерий и грибов
Питание бактерий и грибов Загадка человека
Загадка человека ОРГАНЫ ЧУВСТВ ЧЕЛОВЕКА
ОРГАНЫ ЧУВСТВ ЧЕЛОВЕКА ДИСТ. ЮНЫЕ ДРУЗЬЯ ПРИРОДЫ СШ №11
ДИСТ. ЮНЫЕ ДРУЗЬЯ ПРИРОДЫ СШ №11 Клиническая анатомия и физиология среднего уха
Клиническая анатомия и физиология среднего уха Строение клетки. 5 класс
Строение клетки. 5 класс Презентация на тему "Отряд Крокодилы" - скачать презентации по Биологии
Презентация на тему "Отряд Крокодилы" - скачать презентации по Биологии Организм, как единое целое. Клетки, ткани, органы, системы. Регуляция функций
Организм, как единое целое. Клетки, ткани, органы, системы. Регуляция функций Роль углеводов в жизни человека
Роль углеводов в жизни человека Зрение человека! Выполнили учащиеся 4 г класса МОУ СОШ № 70 Будруев Иван, Макридин Максим, Коньков Алексей Закунов Михаил, рук
Зрение человека! Выполнили учащиеся 4 г класса МОУ СОШ № 70 Будруев Иван, Макридин Максим, Коньков Алексей Закунов Михаил, рук Энзимодиагностика
Энзимодиагностика Видоутворення Еволюційний процес виникнення нових видів.
Видоутворення Еволюційний процес виникнення нових видів.  Охотники и собиратели. Эволюция человека
Охотники и собиратели. Эволюция человека Бесполое размножение организмов – формы и значение в природе Автор презентации учитель биологии высшей категории МОУ «Погорел
Бесполое размножение организмов – формы и значение в природе Автор презентации учитель биологии высшей категории МОУ «Погорел Пищеварительная система человека
Пищеварительная система человека Знакомство с осами
Знакомство с осами Спілкування тварин
Спілкування тварин Класс Птицы: особенности строения, жизнедеятельности как высокоорганизованных позвоночных. АННОТАЦИЯ
Класс Птицы: особенности строения, жизнедеятельности как высокоорганизованных позвоночных. АННОТАЦИЯ Цель: ознакомление с общей характеристикой класса, многообразием и особенностями отрядов акул, скатов, химерообразных
Цель: ознакомление с общей характеристикой класса, многообразием и особенностями отрядов акул, скатов, химерообразных  Игра Интеллект. Вопросы о животных
Игра Интеллект. Вопросы о животных Прикоснись к природе сердцем
Прикоснись к природе сердцем