- Плазмодесмы. Ультраструктура плазмодесм

Содержание
- 2. Что и у кого? Плазмодесмы – микроскопические каналы, пронизывающие клеточную стенку и обеспечивающие межклеточную коммуникацию. У
- 3. Первичные – формируются между сестринскими клетками, развиваются из канальцев в фрагмопласте во время цитокинеза. Вторичные –
- 4. Ультраструктура плазмодесм Спицеобразные линкерные белковые структуры (миозин + актин) разделяют цитоплазматическое кольцо на микроканалы. По ним
- 5. Через плазмодесмы возможен межклеточный транспорт растительных и вирусных белков Косвенное подтверждение Использование радиоактивной метки – 35S-метионин
- 6. Характеристики плазмодесм Плотность (= N плазмодесм / S поверхности КС) Пространственное распределение плазмодесм на поверхности клетки
- 7. Структура и работа плазмодесм
- 8. Надклеточная организация тела высших растений («supracellular organisms») Апопласт – непрерывный континуум клеточных стенок, межклеточных пластинок и
- 10. Гамалей Юрий Владимирович Транспортная система сосудистых растений.- СПб.: Изд-во С.-Петербургского университета, 2004. – 424 с. "Симпласт"
- 11. 2 транспортные сети Апопласт Симпласт Вакуом или Эндопласт Цитозоль не имеет серьёзного транспортного значения
- 12. Симпластное растение – загружает терминальную флоэму ассимилятами по эндоплазматической сети симпласта. Определяющими являются транспортные свойства тонопласта
- 13. Загрузка и разгрузка флоэмы
- 14. Плазмодесмы между клетками-спутницами и ситовидными элементами - точки интенсивного транспорта веществ Необходим транспорт белков в безъядерные
- 16. Симпластическая загрузка флоэмы Сосуд ксилемы Промежуточная клетка-спутница (Intermediary Cell) Плазмодесмы, контактирующие с мезофиллом Ситовидный элемент флоэмы
- 17. Сосуд ксилемы Трансферная клетка-спутница (Transfer Cell) Ситовидный элемент флоэмы Апопластическая загрузка флоэмы Клетка флоэмной паренхимы Плазмодесмы,
- 18. Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих факторов – жизненных форм, условий существования,
- 19. Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих факторов – жизненных форм, условий существования,
- 20. Эксперименты по выяснению пороговой концентрации флоэмных белков, необходимой для межклеточного транспорта Коинъекция постоянного количества FITC-декстранов 20kDa
- 22. Rice Phloem Protein 13kDa (RPP13-1) Высококонсервативный белок флоэмного экссудата, принадлежащий к тиоредоксиновому семейству Экспрессируется только в
- 23. Проводящий пучок стебеля риса, in situ гибридизация мРНК RPP13-1
- 24. In situ гибридизация мРНК RPP13-1 в проводящем пучке листа риса (СС – companion cell, STM -
- 25. Мутантный анализ RPP13-1 Выявлено 2 участка, играющих важную роль в эффективном взаимодействии с плазмодесмами: 5 аминокислотная
- 26. Транспорт вирусных белков: viral movement protein (MP) MP – неструктурные белки, кодируемые в вирусном геноме, основной
- 27. Модели распространения вирусной инфекции
- 28. Модель распространения инфекции клевера white clover mosaic virus - WClMV LC – linker complex (кодируется растением)
- 29. Эпидермальные клетки Arabidopsis thaliana экспрессирующие MP17-GFP (зеленый) Propidium iodide (окрашивание пектинов) маркирует клеточные стенки
- 30. Другой белок - TMV MP30-GFP – тоже зеленый. Видна локализация в плазмодесмах
- 31. Транспорт собственных белков: KN1 (KNOTTED1) – фактор транскрипции, индуцирует дифференцировку клеток Микроинъекция KN1 в клетки мезофилла
- 32. мРНК гена KN1 в апексе кукурузы Листовой примордий 1 Листовой примордий 2 Новый листовой примордий Темное
- 33. Обобщенная модель транспорта больших молекул
- 34. Частичное разворачивание белковой молекулы – необходимо для транслокации через плазмодесму Белки флоэмного экссудата различной массы (фракции
- 35. Cross-linking experiments (препятствие разворачиванию белковой молекулы) и использование с наночастиц золота различных размеров: 1.4, 6, 15
- 37. Плазмодесмы как ворота для инфекции Плазмодесмы используются инфекционными агентами для распространения по организму растения (транспорт вирусных
- 38. Биотический стресс → ↑ салицилаты → ↑транскрипция PDL5 (plasmodesmata-located protein 5) → накопление PDL5 в области
- 39. Вероятно плазмалемма выстилающая плазмодесму и мембрана ЭПР десмотрубочки отличаются по своим физико-химическим параметрам по сравнению с
- 40. Каллоза (β 1-3 глюкан) откладывается в области шейки. Содержание каллозы непостоянно и находится в обратной корреляции
- 41. Белки ассоциированные с плазмодесмами С центральной полостью ассоциированы: TMV MP (обеспечивает транспорт РНК ВТМ в виде
- 42. Недавно было показано, что целый ряд неклеточно-автономных сигнальных молекул, включая ARABIDOPSIS CRINKLY4 (ACR4), CLAVATA1 (CLV1), STRUBBELIG
- 44. Скачать презентацию
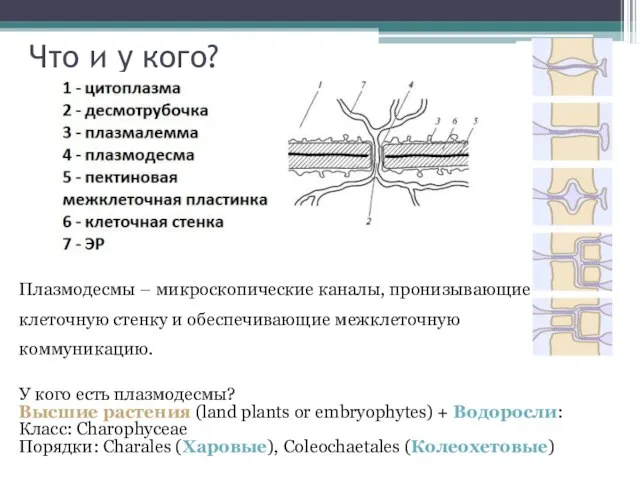
Что и у кого?
Плазмодесмы – микроскопические каналы, пронизывающие клеточную стенку и
Что и у кого?
Плазмодесмы – микроскопические каналы, пронизывающие клеточную стенку и

Первичные – формируются между сестринскими клетками, развиваются из канальцев в фрагмопласте
Первичные – формируются между сестринскими клетками, развиваются из канальцев в фрагмопласте
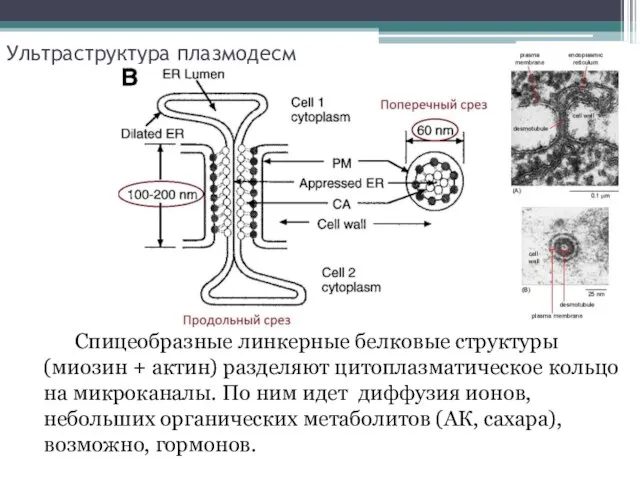
Ультраструктура плазмодесм
Спицеобразные линкерные белковые структуры (миозин + актин) разделяют цитоплазматическое
Ультраструктура плазмодесм
Спицеобразные линкерные белковые структуры (миозин + актин) разделяют цитоплазматическое

Через плазмодесмы возможен межклеточный транспорт растительных и вирусных белков
Косвенное подтверждение
Через плазмодесмы возможен межклеточный транспорт растительных и вирусных белков
Косвенное подтверждение

Характеристики плазмодесм
Плотность (= N плазмодесм / S поверхности КС)
Пространственное распределение плазмодесм
Характеристики плазмодесм
Плотность (= N плазмодесм / S поверхности КС)
Пространственное распределение плазмодесм
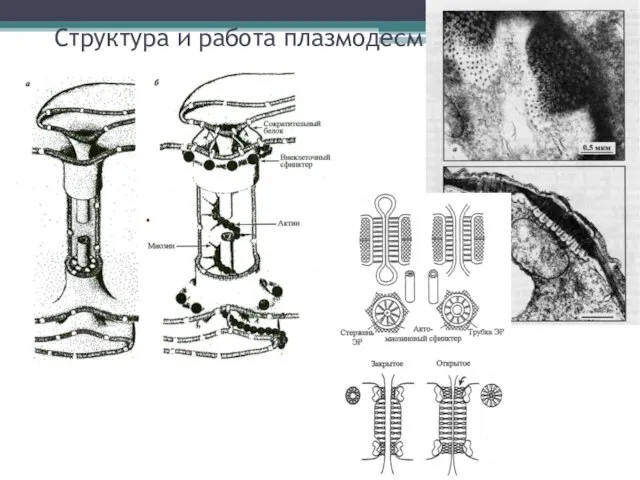
Структура и работа плазмодесм
Структура и работа плазмодесм

Надклеточная организация тела высших растений («supracellular organisms»)
Апопласт – непрерывный континуум
Надклеточная организация тела высших растений («supracellular organisms»)
Апопласт – непрерывный континуум


Гамалей Юрий Владимирович
Транспортная система сосудистых растений.- СПб.: Изд-во С.-Петербургского университета, 2004.
Гамалей Юрий Владимирович Транспортная система сосудистых растений.- СПб.: Изд-во С.-Петербургского университета, 2004.

2 транспортные сети
Апопласт
Симпласт
Вакуом
или Эндопласт
Цитозоль
не имеет серьёзного транспортного значения
2 транспортные сети
Апопласт
Симпласт
Вакуом
или Эндопласт
Цитозоль
не имеет серьёзного транспортного значения

Симпластное растение – загружает терминальную флоэму ассимилятами по эндоплазматической сети симпласта.
Симпластное растение – загружает терминальную флоэму ассимилятами по эндоплазматической сети симпласта.
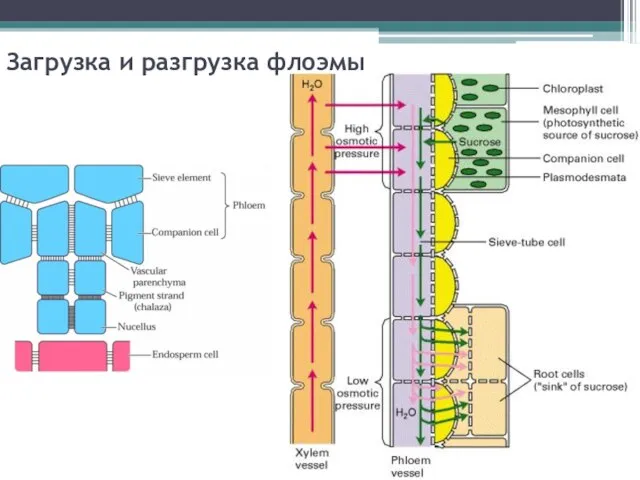
Загрузка и разгрузка флоэмы
Загрузка и разгрузка флоэмы

Плазмодесмы между клетками-спутницами и ситовидными элементами - точки интенсивного транспорта веществ
Необходим
Плазмодесмы между клетками-спутницами и ситовидными элементами - точки интенсивного транспорта веществ
Необходим

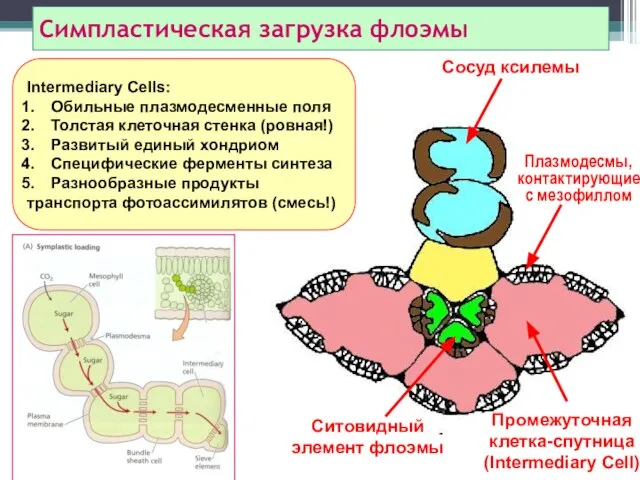
Симпластическая загрузка флоэмы
Сосуд ксилемы
Промежуточная клетка-спутница
(Intermediary Cell)
Плазмодесмы, контактирующие
с мезофиллом
Ситовидный элемент флоэмы
Intermediary
Симпластическая загрузка флоэмы
Сосуд ксилемы
Промежуточная клетка-спутница
(Intermediary Cell)
Плазмодесмы, контактирующие
с мезофиллом
Ситовидный элемент флоэмы
Intermediary
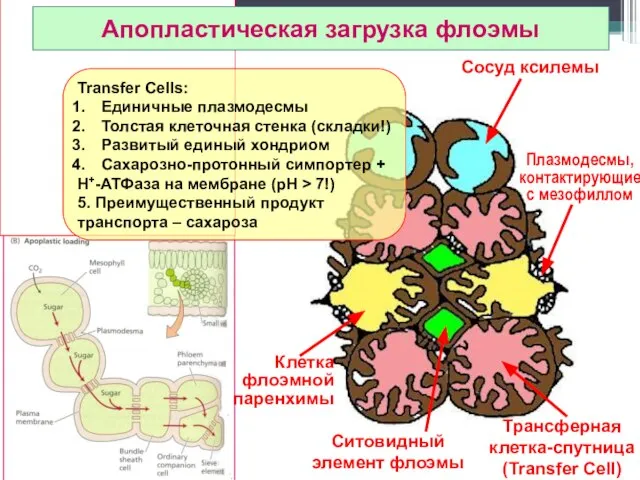
Сосуд ксилемы
Трансферная клетка-спутница
(Transfer Cell)
Ситовидный элемент флоэмы
Апопластическая загрузка флоэмы
Клетка
флоэмной
паренхимы
Плазмодесмы, контактирующие
с
Сосуд ксилемы
Трансферная клетка-спутница
(Transfer Cell)
Ситовидный элемент флоэмы
Апопластическая загрузка флоэмы
Клетка
флоэмной
паренхимы
Плазмодесмы, контактирующие
с

Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих
факторов
Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих
факторов
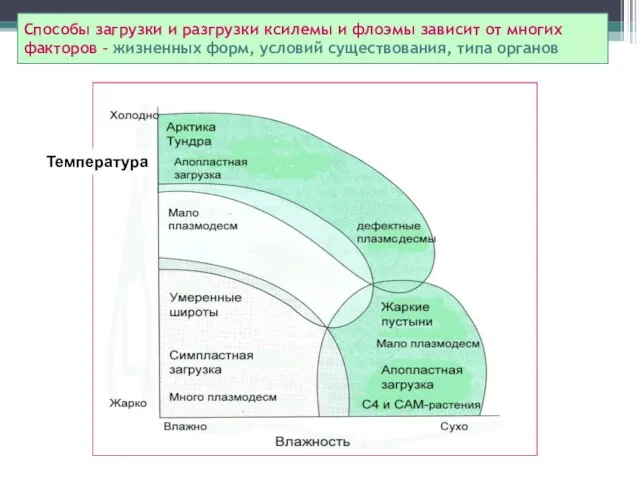
Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих
факторов
Способы загрузки и разгрузки ксилемы и флоэмы зависит от многих факторов

Эксперименты по выяснению пороговой концентрации флоэмных белков, необходимой для межклеточного транспорта
Коинъекция
Эксперименты по выяснению пороговой концентрации флоэмных белков, необходимой для межклеточного транспорта
Коинъекция


Rice Phloem Protein 13kDa (RPP13-1)
Высококонсервативный белок флоэмного экссудата, принадлежащий к тиоредоксиновому
Rice Phloem Protein 13kDa (RPP13-1)
Высококонсервативный белок флоэмного экссудата, принадлежащий к тиоредоксиновому
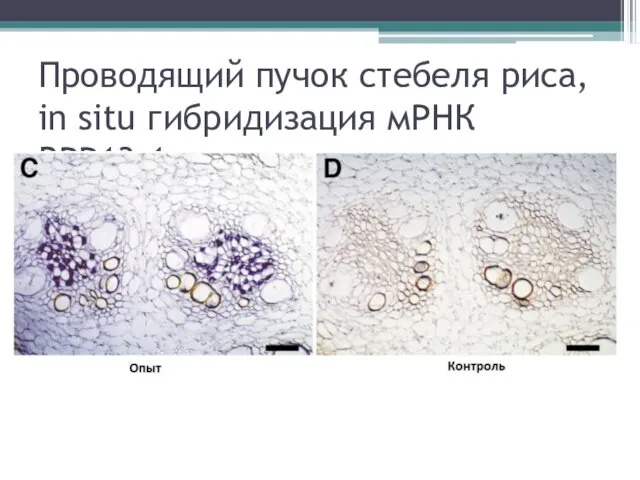
Проводящий пучок стебеля риса, in situ гибридизация мРНК RPP13-1
Проводящий пучок стебеля риса, in situ гибридизация мРНК RPP13-1

In situ гибридизация мРНК RPP13-1 в проводящем пучке листа риса (СС
In situ гибридизация мРНК RPP13-1 в проводящем пучке листа риса (СС

Мутантный анализ RPP13-1
Выявлено 2 участка, играющих важную роль в эффективном взаимодействии
Мутантный анализ RPP13-1
Выявлено 2 участка, играющих важную роль в эффективном взаимодействии

Транспорт вирусных белков: viral movement protein (MP)
MP – неструктурные белки, кодируемые
Транспорт вирусных белков: viral movement protein (MP)
MP – неструктурные белки, кодируемые

Модели распространения вирусной инфекции
Модели распространения вирусной инфекции

Модель распространения инфекции клевера white clover mosaic virus - WClMV
LC –
Модель распространения инфекции клевера white clover mosaic virus - WClMV
LC –
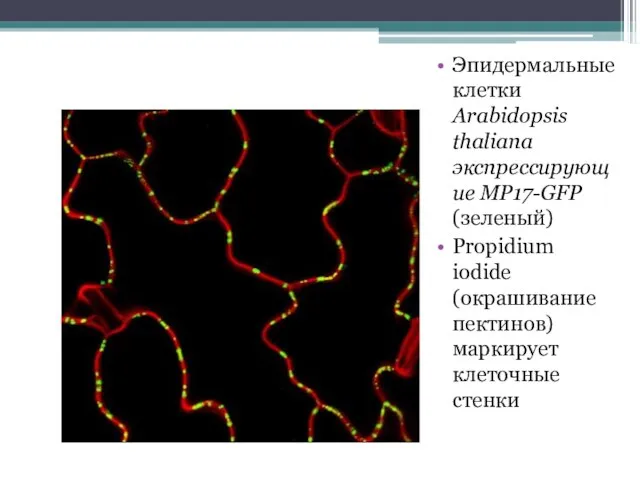
Эпидермальные клетки Arabidopsis thaliana экспрессирующие MP17-GFP (зеленый)
Propidium iodide (окрашивание пектинов) маркирует
Эпидермальные клетки Arabidopsis thaliana экспрессирующие MP17-GFP (зеленый)
Propidium iodide (окрашивание пектинов) маркирует
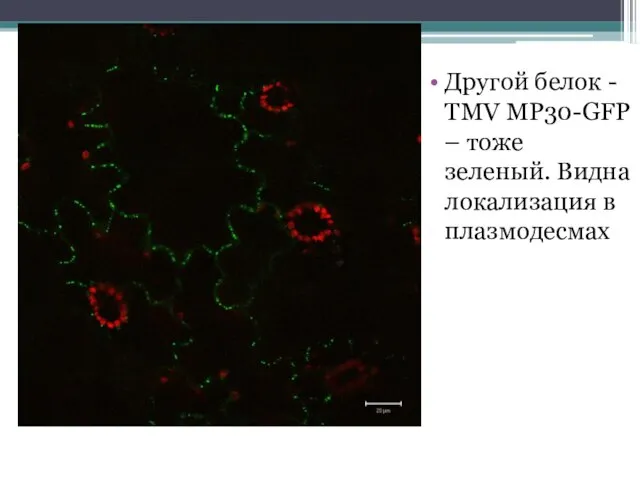
Другой белок - TMV MP30-GFP – тоже зеленый. Видна локализация в
Другой белок - TMV MP30-GFP – тоже зеленый. Видна локализация в

Транспорт собственных белков: KN1 (KNOTTED1)
– фактор транскрипции, индуцирует дифференцировку клеток
Микроинъекция KN1
Транспорт собственных белков: KN1 (KNOTTED1)
– фактор транскрипции, индуцирует дифференцировку клеток
Микроинъекция KN1
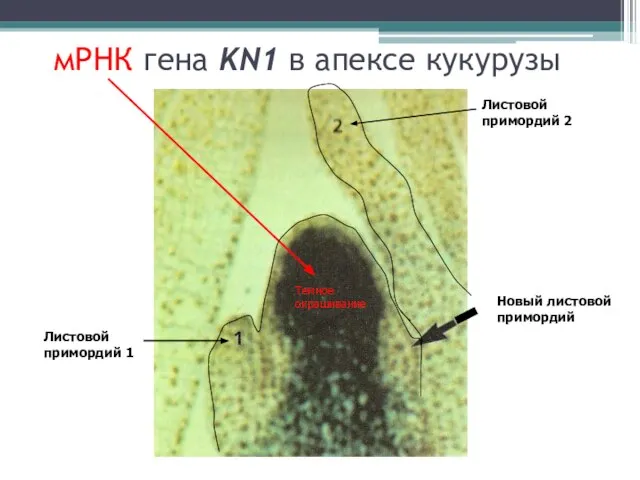
мРНК гена KN1 в апексе кукурузы
Листовой примордий 1
Листовой примордий 2
Новый листовой
мРНК гена KN1 в апексе кукурузы
Листовой примордий 1
Листовой примордий 2
Новый листовой

Обобщенная модель транспорта больших молекул
Обобщенная модель транспорта больших молекул

Частичное разворачивание белковой молекулы – необходимо для транслокации через плазмодесму
Белки флоэмного
Частичное разворачивание белковой молекулы – необходимо для транслокации через плазмодесму Белки флоэмного

Cross-linking experiments (препятствие разворачиванию белковой молекулы) и использование с наночастиц золота
Cross-linking experiments (препятствие разворачиванию белковой молекулы) и использование с наночастиц золота


Плазмодесмы как ворота для инфекции
Плазмодесмы используются инфекционными агентами для распространения по
Плазмодесмы как ворота для инфекции
Плазмодесмы используются инфекционными агентами для распространения по

Биотический стресс → ↑ салицилаты → ↑транскрипция PDL5 (plasmodesmata-located protein 5)
Биотический стресс → ↑ салицилаты → ↑транскрипция PDL5 (plasmodesmata-located protein 5)

Вероятно плазмалемма выстилающая плазмодесму и мембрана ЭПР десмотрубочки отличаются по своим
Вероятно плазмалемма выстилающая плазмодесму и мембрана ЭПР десмотрубочки отличаются по своим

Каллоза (β 1-3 глюкан) откладывается в области шейки. Содержание каллозы непостоянно
Каллоза (β 1-3 глюкан) откладывается в области шейки. Содержание каллозы непостоянно

Белки ассоциированные с плазмодесмами
С центральной полостью ассоциированы: TMV MP (обеспечивает транспорт
Белки ассоциированные с плазмодесмами
С центральной полостью ассоциированы: TMV MP (обеспечивает транспорт

Недавно было показано, что целый ряд неклеточно-автономных сигнальных молекул, включая ARABIDOPSIS
Недавно было показано, что целый ряд неклеточно-автономных сигнальных молекул, включая ARABIDOPSIS
 Эмбриологические доказательства
Эмбриологические доказательства Нервная система. Головной мозг
Нервная система. Головной мозг Урок-путешествие Козлова И.А. МАОУСОШ №8 г.Старая Русса Новгородской обл.
Урок-путешествие Козлова И.А. МАОУСОШ №8 г.Старая Русса Новгородской обл. Полосатый рейс онлайн - квиз
Полосатый рейс онлайн - квиз Введение в обмен веществ
Введение в обмен веществ Презентация на тему Экологическая роль и медицинское значение бактерий
Презентация на тему Экологическая роль и медицинское значение бактерий Корень как орган растения. Значение и разнообразие корней
Корень как орган растения. Значение и разнообразие корней Многообразие бабочек города Волгореченска. Автор: Жочкина А. учащаяся 9а класса МОУ СОШ №3 Руководитель: Звёздочкина С.А. Учитель
Многообразие бабочек города Волгореченска. Автор: Жочкина А. учащаяся 9а класса МОУ СОШ №3 Руководитель: Звёздочкина С.А. Учитель  Какое дерево самое большое
Какое дерево самое большое Нервная система: строение и функционирование. Подготовка к ОГЭ/ ЕГЭ
Нервная система: строение и функционирование. Подготовка к ОГЭ/ ЕГЭ Питание растений
Питание растений Среда обитания
Среда обитания Синквейн
Синквейн Организация огорода в детском саду (из опыта работы)
Организация огорода в детском саду (из опыта работы) Evolution: Artificial Selection
Evolution: Artificial Selection Решение задач на моногибридное скрещивание 1 и 2 законы Г. Менделя
Решение задач на моногибридное скрещивание 1 и 2 законы Г. Менделя Распространение и роль водорослей в природе. Экологические группы водорослей. (Лекция 7)
Распространение и роль водорослей в природе. Экологические группы водорослей. (Лекция 7) Клещи
Клещи Презентация на тему "Детям о вакцинации" - скачать бесплатно презентации по Биологии
Презентация на тему "Детям о вакцинации" - скачать бесплатно презентации по Биологии Систематическое положение человека
Систематическое положение человека Проверка домашнего задания: Укажите какие утверждения верны. Все микробы- бактерии. Все заболевания человека вызываются бакт
Проверка домашнего задания: Укажите какие утверждения верны. Все микробы- бактерии. Все заболевания человека вызываются бакт Членистоногие. Строение паукообразных, насекомых, ракообразных
Членистоногие. Строение паукообразных, насекомых, ракообразных Форменные элементы крови
Форменные элементы крови Половой цикл КРС и его основные фазы
Половой цикл КРС и его основные фазы Презентация на тему "Путешествие в мир Простейших" - скачать презентации по Биологии
Презентация на тему "Путешествие в мир Простейших" - скачать презентации по Биологии Сорокопуты
Сорокопуты Презентация на тему "Птицы Забота о потомстве" - скачать презентации по Биологии
Презентация на тему "Птицы Забота о потомстве" - скачать презентации по Биологии Невидимые нити. Окружающий мир 2 класс
Невидимые нити. Окружающий мир 2 класс