- Липидтердің метаболизмі

Содержание
- 2. ЛИПИДТЕРДІҢ БИОЛОГИЯЛЫҚ МАҢЫЗЫ Энергетикалық Мембрана компоненттері (құрылымдық) гормондардың алғы заты (стероидтар) Белгі бергіш молекулалар (простагландиндер) Қорғаныштық
- 3. ТРИАЦИЛГЛИЦЕРОЛДАР –ЭНЕРГИЯ ДЕПОСЫ Триацилглицерол (ТГ) және гликоген – энергия сақтаудың негізгі екі формасыТГ –энергия депосы, себебі:
- 4. Май қышқылдары және глицерол –ағзада қуат көзі. Май қышқылдары триацилглицеролдардан тұрады
- 5. Тағамдық липидтер: ∙ триацилглицерол (ТАГ) ∙ фосфолипид ∙ холестерол Жіңішке ішекте панкреатит ферменті липаза C1 және
- 6. Өт қышқылдарының тұздары липидтерд ыдыратуға қажет. Бауырда холестеролдан синтезделеді. Таурохол және гликохол қышқылдары – ең көп
- 7. ТАГ – суда ерімейтін, ал липаза – суда еритін. ТАГ ыдырауы липид-су шекарасында болады. ТАГ ыдырау
- 8. Май қышқылдарының тұздары липазаны белсендіреді.
- 9. Фосфолипидтер фосфолипазаның қатысында ыдырайды Фосфолипазалар асқазан асты сөлінде түзіледі Негізгі фосфолипаза – фосфолипаза A2 ( лизофосфоглицеридтердің
- 10. Лизофосфоглицеридтер детергенттер ретінде және жоғары концентрацияда жасушалық мембрананы бұзады. Лизофосфоглицеридтер жасушада қалыпты жағдайда төменгі концентацияда болады.
- 11. Тағам холестеролы Көптеген тағам холестерол бос жағдайда болады (этерифицирленбеген) Холестерол эфирлері жіңішке ішекте эстеразамен гидролизденеді. Бос
- 12. ЛИПИДТЕРДІҢ СІҢІРІЛУІ 2-моноацилглцеролдер, май қышқылдары, лизофосфоглицеролдар, бос холестерол өт қышқылдарының тұздарымен мицелла түзеді. Липидтердің сіңірілуі пассивті
- 13. Өт қышқылдарының тұздары бауырға вена арқылы белсенді тасымалданады. Өт қышқылдарының тұздары жіңішке ішек және бауыр арқылы
- 14. Бұл липидтер фосфолипидтермен және протеиндермен липопротеиндер түзеді. Құрылымы: Гидрофобты ядро: Гидрофильді беті Липидтердің тасымалдайтын формалары ТАГ,
- 15. Липопротеиндердің негізгі кластары Хиломикрондар. Өте төменгі тығыздықтағы липопротеиндер. Аралық тығыздықтағы липопротеиндер. Төменгі тығыздықтағы липопротеиндер. Жоғары тығыздықтағы
- 16. Липопротеиндер (180-500 нм ) 85 % ТАГ (ТАГ негізгі тасымалдаушы формасы). апопротеин B-48 (aпо B-48) –
- 17. өте төменгі тығыздықтағы липопротеиндер бауырда түзіледі ТАГ 50 % және 22 % холестерол екі апопротеин —
- 18. Төменгі тығыздықтағы липопротиндер ЛНП қанда ЛПП және бауырда ЛПП түзіледі (фермент – бауыр липазасы) ЛНП холестерол
- 19. Жоғары тығыздықтағы липопротеиндер бауырда және аз мөлшерде жіңішке ішекте түзіледі Құрамында көп мөлшерде ақуыз (40 %)
- 20. Липидтерді тасымалдау формалары
- 21. ХОЛЕСТЕРОЛ МЕТАБОЛИЗМІ
- 22. Холестеринің биосинтезі
- 23. Холестеролдың функциясы стероидты гормондардың алғы заты (прогестерон, тестостерон, эстрадиол, кортизол) өт қышқылдары тұздарының алғы заты D
- 24. Холестеролдың көзі Диета, бауырда (көпшілігінде), ішекте холестерол жасушаға липопротеиндермен тасымалданады
- 25. холестерол синтезі холестерол синтезінің үш сатысы 1. изопентенил пирофосфаттың синтезі, 2. алты молекула изопентенил пирофосфатының сквален
- 26. Холестеролдың барлық көміртегі атомы цитозольдегі ацетил CoA түзіледі (митохондриядан цитратты тасымалдану жүйесімен түзіледі) A. 1 саты
- 27. Келесіде үш молекла ацетил CoA конденсацияланады Екі молекула ацетил CoA ацетоацетил CoA түзе конденсирленеді. Фермент –
- 28. Ацетоацетил CoA ацетил CoA мен судың қатысында 3-гидрокси-3-метилглютарил CoA түзеді. Фермент: гидроксиметилглютарил CoA синтаза
- 29. Цитоплазмада 3-гидрокси-3-метилглютарил CoA мевалонатқа дейін тотықсызданады. Митохондрияда 3-гидрокси-3-метилглютарил CoA кейіннен ацетил CoA және ацетоацетатқа дейін ыдырайды.
- 30. ГМГ-CoA редуктаза - эндоплазмалық ретикулумдегі интегральді мембраналық ақуыз холестерол синтезінің негізгі айналымын құрайды Холестерол- статиндерді төмендетеді
- 31. Мевалонат кейіннен 3-изопентенил пирофосфатқа айналады, онда ATФ және декарбоксилдену реакциясы жүреді.
- 32. 2 Саты : изопентенил пирофосфаттың скваленге айналуы Изопентенил пирофосфат изомерленіп диметилалил пирофосфат түзеді.
- 33. C. 3 Саты: скваленнің холестеролға айналуы
- 34. Ланостерол холестеролға айналады
- 35. ХОЛЕСТЕРОЛ БИОСИНТЕЗІНІҢ РЕГУЛЯЦИЯСЫ Регуляторлық фермент - 3-гидрокси-3-метилглютарил CoA редуктаза. Тетрамерлі фермент. НАДФН - кофермент
- 36. холестерол метаболизмінің өнімдері
- 37. Қан тамырларын бұзады. ХХ және ХХІ ғасырдағы негізгі медициналық мәселе Биохимилық негізі - гиперхолестеролемия Қан плазмасындағы
- 38. ЛИПИДТЕРДІҢ МЕТАБОЛИЗМІ : КЕТОНДЫҚ ДЕНЕЛЕР
- 39. ацетил CoA- нің Кребс цикліне қатысуы оксалоацетатақа байланысты. Оксалоацетаттың концентрациясы көмірсу түспесе кемиді (аштық) немесе метаболизденеді
- 40. Аштықта немесе диабетте глюконеогенез және оксалоацетат белсендіріледі. Май қышқылдары тотығады, нәтижесінде ацетил CoA артық мөлшері түзіледі
- 41. A. Кетонды денелер синтезі Екі молекула ацетил СоА ацетоацетил CoA түзе конденсирленеді. Фермент – тиолаза.
- 42. Ацетоацетил CoA кейіннен ацетил CoA және сумен 3-гидрокси-3-метилглутарил CoA (HMG-CoA) және CoA. Фермент: HMG-CoA синтаза
- 43. 3-Гидрокси-3-метилглутарил CoA кейіннен ацетил CoA және ацетоацетата түзе ыдырайды. Фермент: HMG-CoA лиаза.
- 44. 3-Гидроксибутират ацетоацетаттың 3-гидроксибутират-дегидрогеназ қатысында түзіледі. Ацетоацетат ацетонға дейін декарбоксилденеді. Ацетона иісі ацетоацетаттың қанның құрамындағы артық концентрациясымен
- 45. B. Кетонды денелер – кейбір ұлпалар үшін «жанармай»көзі Кетондық денелер бауыр митохондриясынан қанға өт перифериялық ұлпаларға
- 46. 3-Гидроксибутират тотығып ацетоацетата және НАДН түзеді.
- 48. Скачать презентацию

ЛИПИДТЕРДІҢ БИОЛОГИЯЛЫҚ МАҢЫЗЫ
Энергетикалық
Мембрана компоненттері (құрылымдық)
гормондардың алғы заты
ЛИПИДТЕРДІҢ БИОЛОГИЯЛЫҚ МАҢЫЗЫ
Энергетикалық
Мембрана компоненттері (құрылымдық)
гормондардың алғы заты

ТРИАЦИЛГЛИЦЕРОЛДАР –ЭНЕРГИЯ ДЕПОСЫ
Триацилглицерол (ТГ) және гликоген – энергия сақтаудың негізгі екі
ТРИАЦИЛГЛИЦЕРОЛДАР –ЭНЕРГИЯ ДЕПОСЫ
Триацилглицерол (ТГ) және гликоген – энергия сақтаудың негізгі екі
Май қышқылдары және глицерол –ағзада қуат көзі.
Май қышқылдары триацилглицеролдардан тұрады
Май қышқылдары және глицерол –ағзада қуат көзі.
Май қышқылдары триацилглицеролдардан тұрады
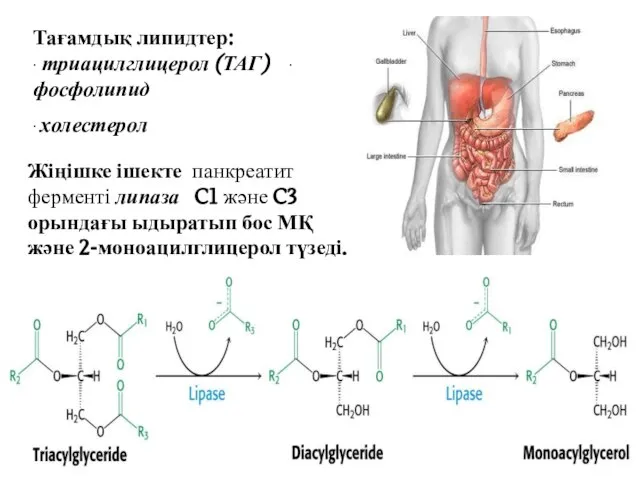
Тағамдық липидтер: ∙ триацилглицерол (ТАГ) ∙ фосфолипид
∙ холестерол
Жіңішке ішекте панкреатит
Тағамдық липидтер: ∙ триацилглицерол (ТАГ) ∙ фосфолипид
∙ холестерол
Жіңішке ішекте панкреатит

Өт қышқылдарының тұздары липидтерд ыдыратуға қажет. Бауырда холестеролдан синтезделеді.
Таурохол және
Өт қышқылдарының тұздары липидтерд ыдыратуға қажет. Бауырда холестеролдан синтезделеді.
Таурохол және

ТАГ – суда ерімейтін, ал липаза – суда еритін.
ТАГ ыдырауы
ТАГ – суда ерімейтін, ал липаза – суда еритін.
ТАГ ыдырауы
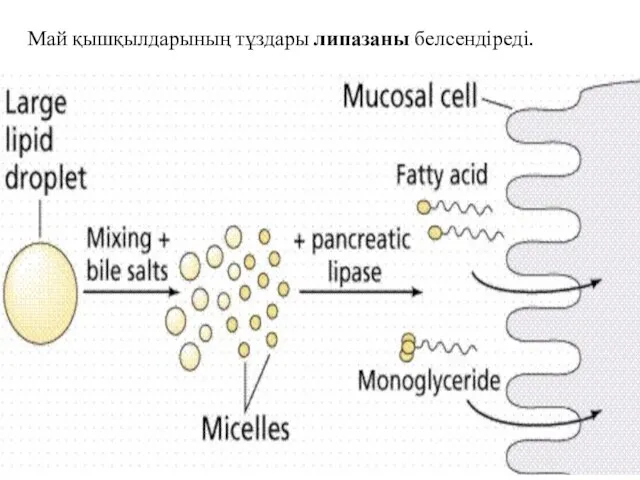
Май қышқылдарының тұздары липазаны белсендіреді.
Май қышқылдарының тұздары липазаны белсендіреді.
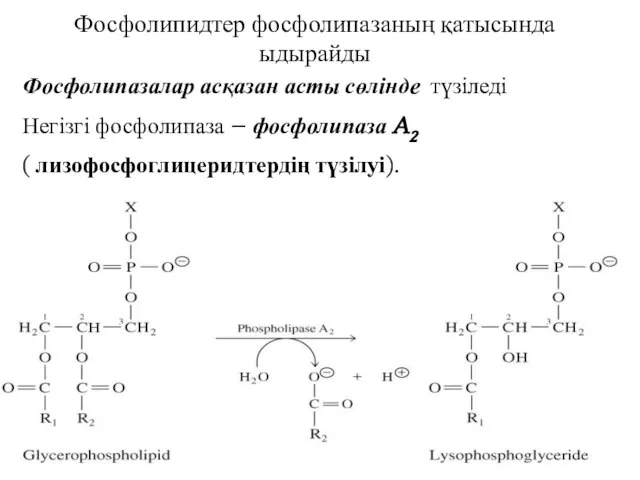
Фосфолипидтер фосфолипазаның қатысында ыдырайды
Фосфолипазалар асқазан асты сөлінде түзіледі
Негізгі фосфолипаза – фосфолипаза
Фосфолипидтер фосфолипазаның қатысында ыдырайды
Фосфолипазалар асқазан асты сөлінде түзіледі
Негізгі фосфолипаза – фосфолипаза

Лизофосфоглицеридтер детергенттер ретінде және жоғары концентрацияда жасушалық мембрананы бұзады.
Лизофосфоглицеридтер жасушада
Лизофосфоглицеридтер детергенттер ретінде және жоғары концентрацияда жасушалық мембрананы бұзады.
Лизофосфоглицеридтер жасушада

Тағам холестеролы
Көптеген тағам холестерол бос жағдайда болады (этерифицирленбеген)
Холестерол эфирлері жіңішке ішекте
Тағам холестеролы
Көптеген тағам холестерол бос жағдайда болады (этерифицирленбеген)
Холестерол эфирлері жіңішке ішекте
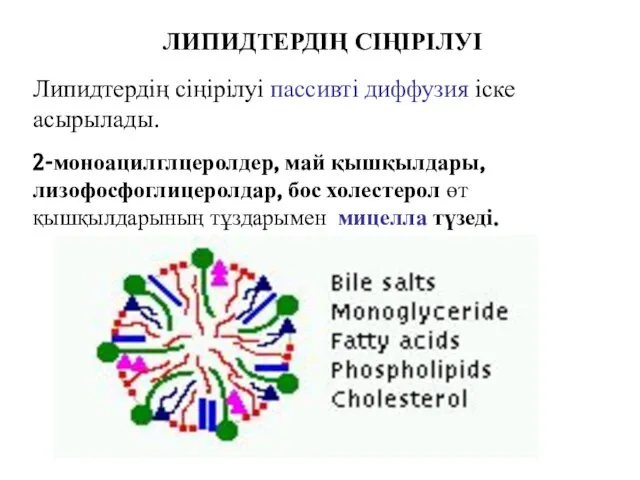
ЛИПИДТЕРДІҢ СІҢІРІЛУІ
2-моноацилглцеролдер, май қышқылдары, лизофосфоглицеролдар, бос холестерол өт қышқылдарының тұздарымен мицелла
ЛИПИДТЕРДІҢ СІҢІРІЛУІ
2-моноацилглцеролдер, май қышқылдары, лизофосфоглицеролдар, бос холестерол өт қышқылдарының тұздарымен мицелла
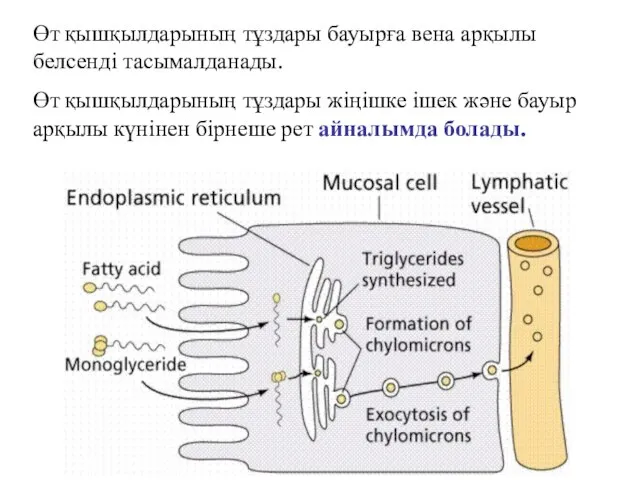
Өт қышқылдарының тұздары бауырға вена арқылы белсенді тасымалданады.
Өт қышқылдарының тұздары жіңішке
Өт қышқылдарының тұздары бауырға вена арқылы белсенді тасымалданады.
Өт қышқылдарының тұздары жіңішке
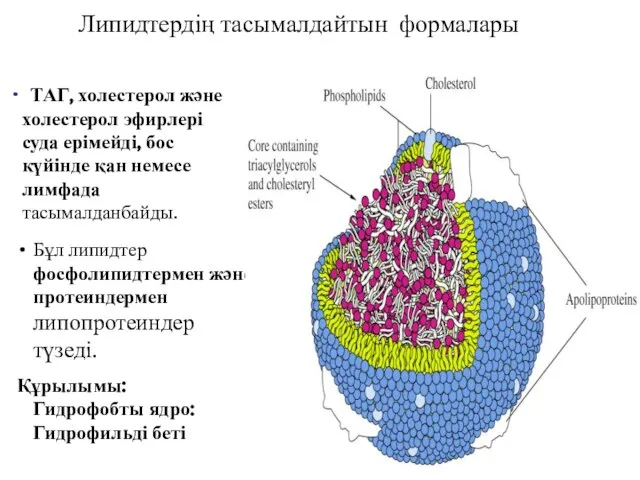
Бұл липидтер фосфолипидтермен және протеиндермен липопротеиндер түзеді.
Құрылымы: Гидрофобты ядро:
Гидрофильді беті
Липидтердің
Бұл липидтер фосфолипидтермен және протеиндермен липопротеиндер түзеді.
Құрылымы: Гидрофобты ядро:
Гидрофильді беті
Липидтердің

Липопротеиндердің негізгі кластары
Хиломикрондар. Өте төменгі тығыздықтағы липопротеиндер.
Аралық тығыздықтағы липопротеиндер. Төменгі
Липопротеиндердің негізгі кластары
Хиломикрондар. Өте төменгі тығыздықтағы липопротеиндер.
Аралық тығыздықтағы липопротеиндер. Төменгі
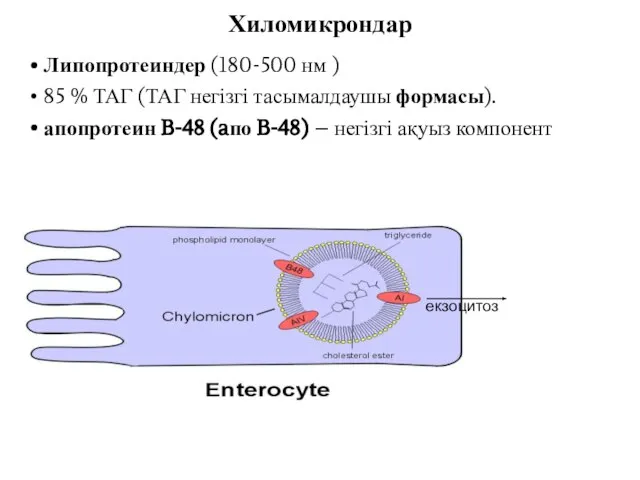
Липопротеиндер (180-500 нм )
85 % ТАГ (ТАГ негізгі тасымалдаушы формасы).
апопротеин B-48
Липопротеиндер (180-500 нм )
85 % ТАГ (ТАГ негізгі тасымалдаушы формасы).
апопротеин B-48
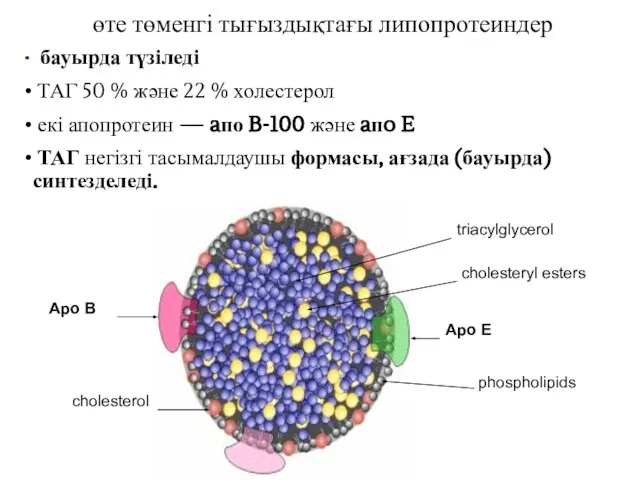
өте төменгі тығыздықтағы липопротеиндер
бауырда түзіледі
ТАГ 50 % және 22
өте төменгі тығыздықтағы липопротеиндер
бауырда түзіледі
ТАГ 50 % және 22
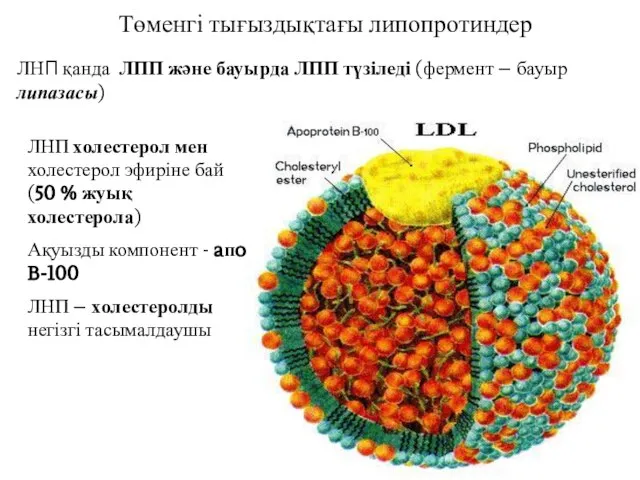
Төменгі тығыздықтағы липопротиндер
ЛНП қанда ЛПП және бауырда ЛПП түзіледі (фермент –
Төменгі тығыздықтағы липопротиндер
ЛНП қанда ЛПП және бауырда ЛПП түзіледі (фермент –

Жоғары тығыздықтағы липопротеиндер
бауырда және аз мөлшерде жіңішке ішекте түзіледі
Құрамында
Жоғары тығыздықтағы липопротеиндер
бауырда және аз мөлшерде жіңішке ішекте түзіледі
Құрамында
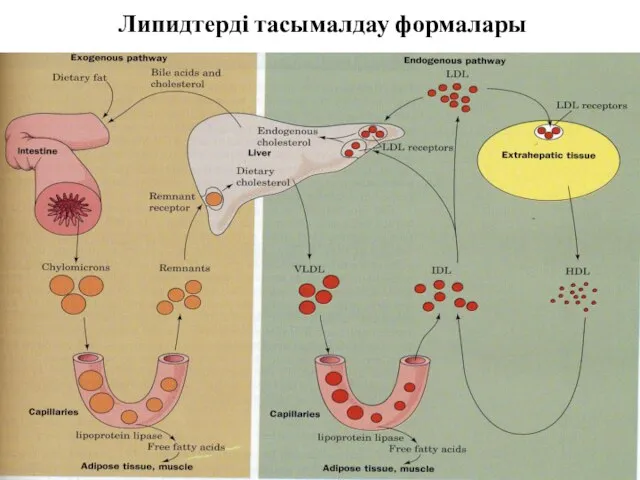
Липидтерді тасымалдау формалары
Липидтерді тасымалдау формалары
ХОЛЕСТЕРОЛ МЕТАБОЛИЗМІ
ХОЛЕСТЕРОЛ МЕТАБОЛИЗМІ
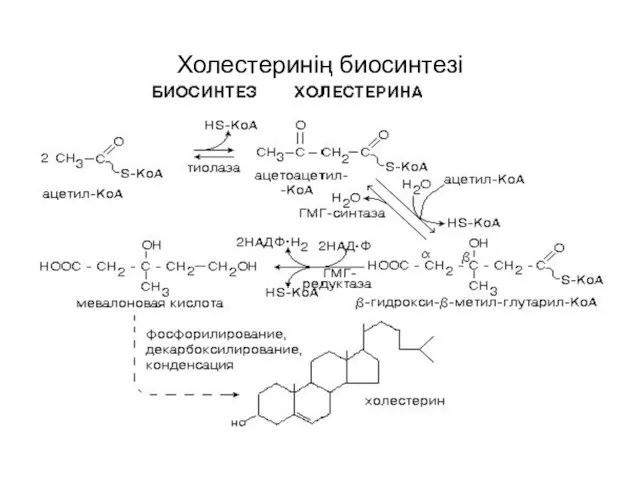
Холестеринің биосинтезі
Холестеринің биосинтезі

Холестеролдың функциясы
стероидты гормондардың алғы заты (прогестерон, тестостерон, эстрадиол, кортизол)
Холестеролдың функциясы
стероидты гормондардың алғы заты (прогестерон, тестостерон, эстрадиол, кортизол)
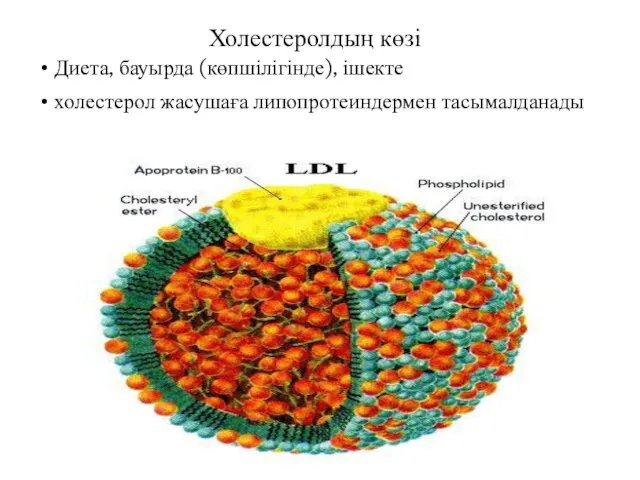
Холестеролдың көзі
Диета, бауырда (көпшілігінде), ішекте
холестерол жасушаға липопротеиндермен тасымалданады
Холестеролдың көзі
Диета, бауырда (көпшілігінде), ішекте
холестерол жасушаға липопротеиндермен тасымалданады

холестерол синтезі
холестерол синтезінің үш сатысы
1. изопентенил пирофосфаттың синтезі,
2. алты молекула
холестерол синтезі
холестерол синтезінің үш сатысы
1. изопентенил пирофосфаттың синтезі,
2. алты молекула
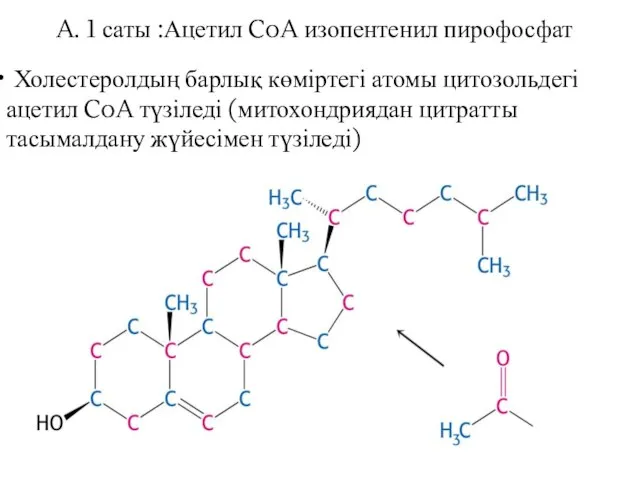
Холестеролдың барлық көміртегі атомы цитозольдегі ацетил CoA түзіледі (митохондриядан цитратты
Холестеролдың барлық көміртегі атомы цитозольдегі ацетил CoA түзіледі (митохондриядан цитратты
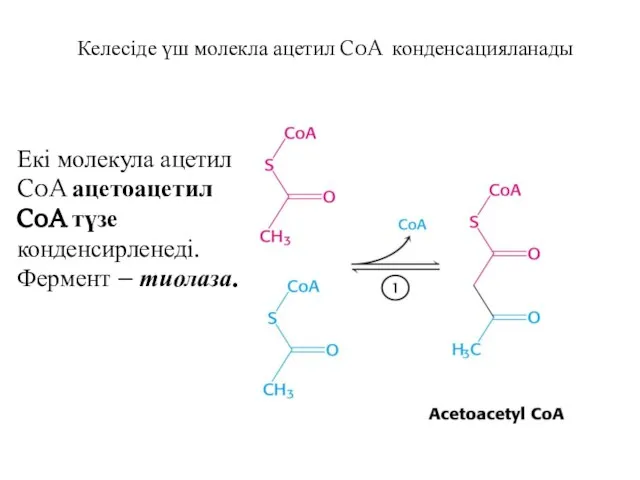
Келесіде үш молекла ацетил CoA конденсацияланады
Екі молекула ацетил CoA ацетоацетил CoA
Келесіде үш молекла ацетил CoA конденсацияланады
Екі молекула ацетил CoA ацетоацетил CoA
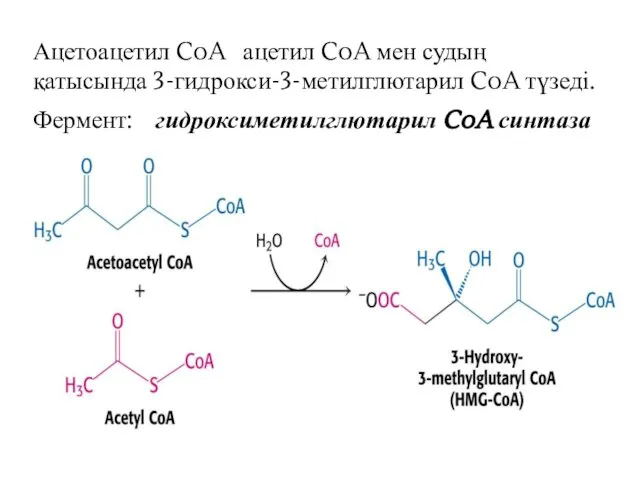
Ацетоацетил CoA ацетил CoA мен судың қатысында 3-гидрокси-3-метилглютарил CoA түзеді.
Фермент: гидроксиметилглютарил
Ацетоацетил CoA ацетил CoA мен судың қатысында 3-гидрокси-3-метилглютарил CoA түзеді.
Фермент: гидроксиметилглютарил
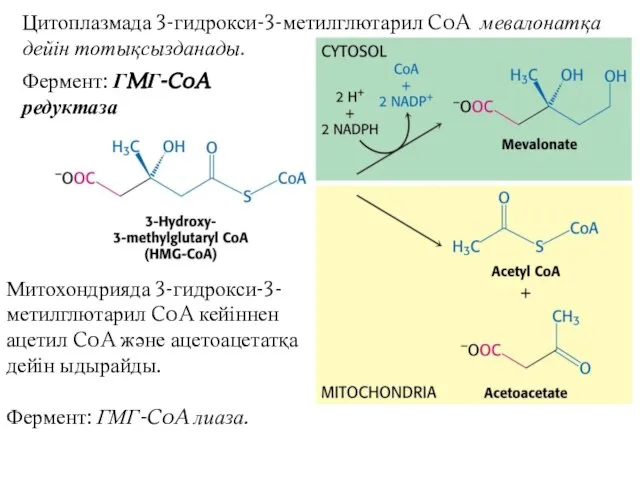
Цитоплазмада 3-гидрокси-3-метилглютарил CoA мевалонатқа дейін тотықсызданады.
Митохондрияда 3-гидрокси-3-метилглютарил CoA кейіннен ацетил
Цитоплазмада 3-гидрокси-3-метилглютарил CoA мевалонатқа дейін тотықсызданады.
Митохондрияда 3-гидрокси-3-метилглютарил CoA кейіннен ацетил
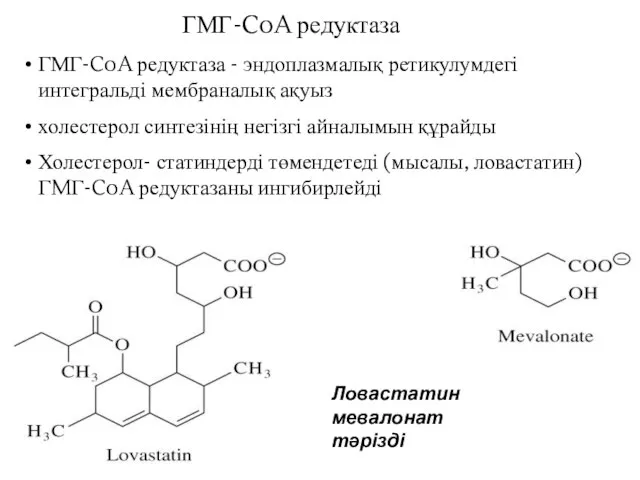
ГМГ-CoA редуктаза - эндоплазмалық ретикулумдегі интегральді мембраналық ақуыз
холестерол синтезінің негізгі айналымын
ГМГ-CoA редуктаза - эндоплазмалық ретикулумдегі интегральді мембраналық ақуыз
холестерол синтезінің негізгі айналымын
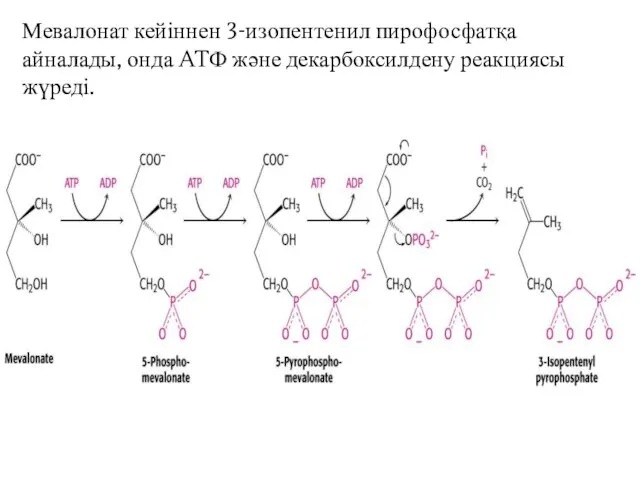
Мевалонат кейіннен 3-изопентенил пирофосфатқа айналады, онда ATФ және декарбоксилдену реакциясы жүреді.
Мевалонат кейіннен 3-изопентенил пирофосфатқа айналады, онда ATФ және декарбоксилдену реакциясы жүреді.
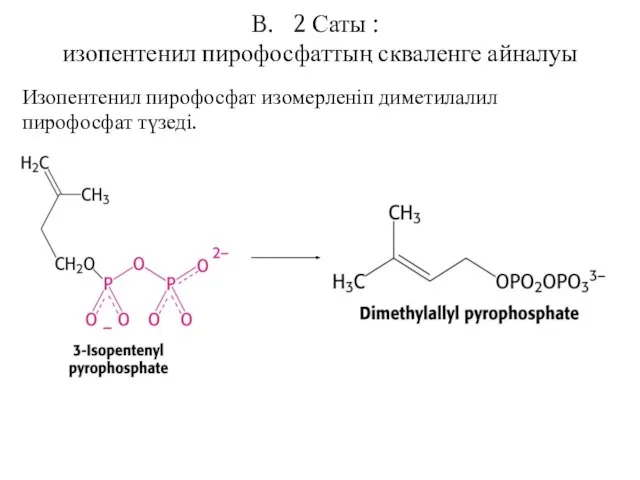
2 Саты :
изопентенил пирофосфаттың скваленге айналуы
Изопентенил пирофосфат изомерленіп диметилалил пирофосфат
2 Саты :
изопентенил пирофосфаттың скваленге айналуы
Изопентенил пирофосфат изомерленіп диметилалил пирофосфат
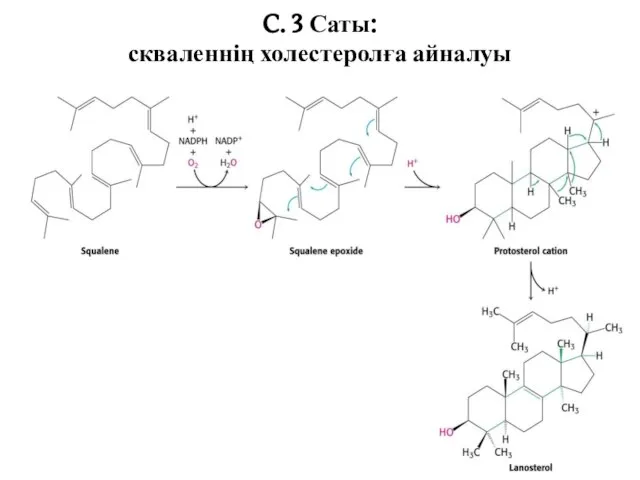
C. 3 Саты:
скваленнің холестеролға айналуы
C. 3 Саты:
скваленнің холестеролға айналуы
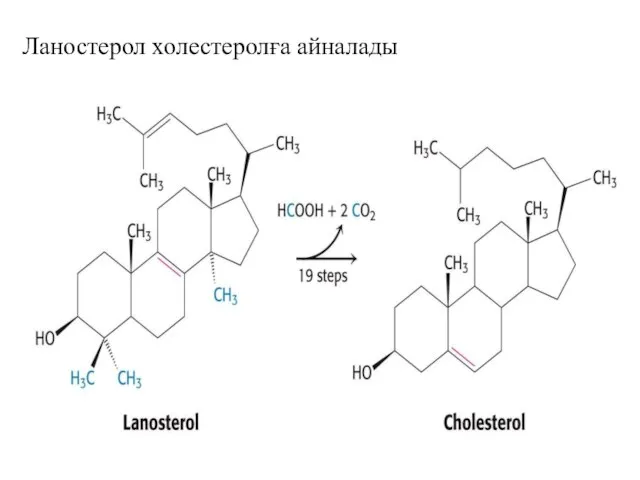
Ланостерол холестеролға айналады
Ланостерол холестеролға айналады

ХОЛЕСТЕРОЛ БИОСИНТЕЗІНІҢ РЕГУЛЯЦИЯСЫ
Регуляторлық фермент - 3-гидрокси-3-метилглютарил CoA редуктаза.
Тетрамерлі фермент.
НАДФН - кофермент
ХОЛЕСТЕРОЛ БИОСИНТЕЗІНІҢ РЕГУЛЯЦИЯСЫ
Регуляторлық фермент - 3-гидрокси-3-метилглютарил CoA редуктаза.
Тетрамерлі фермент.
НАДФН - кофермент
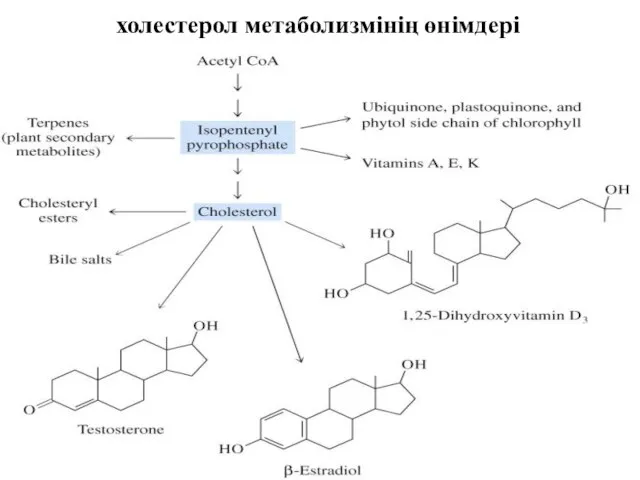
холестерол метаболизмінің өнімдері
холестерол метаболизмінің өнімдері

Қан тамырларын бұзады.
ХХ және ХХІ ғасырдағы негізгі медициналық мәселе
Биохимилық негізі -
Қан тамырларын бұзады.
ХХ және ХХІ ғасырдағы негізгі медициналық мәселе
Биохимилық негізі -

ЛИПИДТЕРДІҢ МЕТАБОЛИЗМІ : КЕТОНДЫҚ ДЕНЕЛЕР
ЛИПИДТЕРДІҢ МЕТАБОЛИЗМІ : КЕТОНДЫҚ ДЕНЕЛЕР
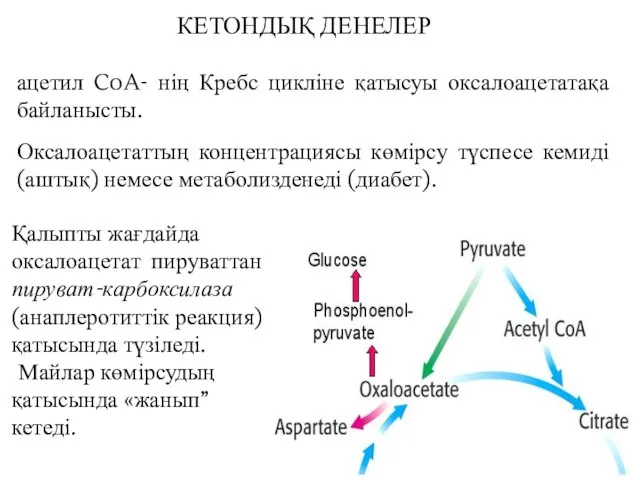
ацетил CoA- нің Кребс цикліне қатысуы оксалоацетатақа байланысты.
Оксалоацетаттың концентрациясы көмірсу түспесе
ацетил CoA- нің Кребс цикліне қатысуы оксалоацетатақа байланысты.
Оксалоацетаттың концентрациясы көмірсу түспесе
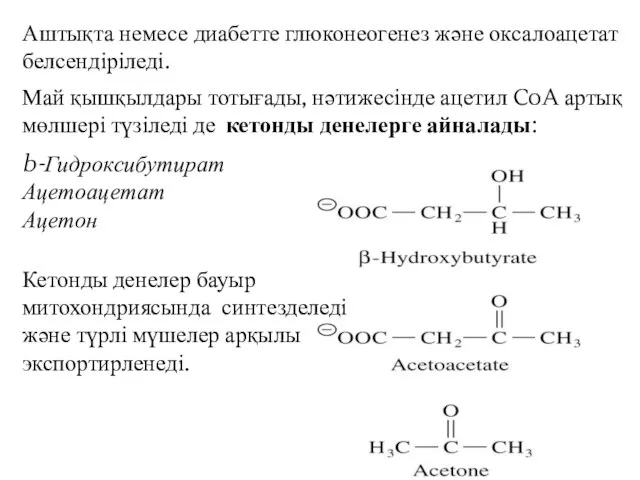
Аштықта немесе диабетте глюконеогенез және оксалоацетат белсендіріледі.
Май қышқылдары тотығады, нәтижесінде ацетил
Аштықта немесе диабетте глюконеогенез және оксалоацетат белсендіріледі.
Май қышқылдары тотығады, нәтижесінде ацетил
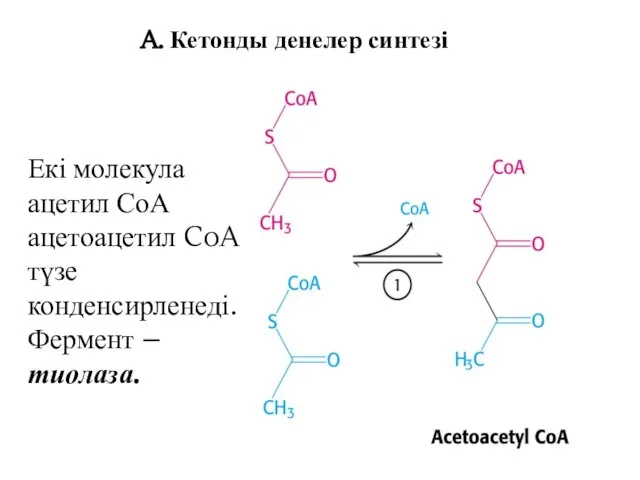
A. Кетонды денелер синтезі
Екі молекула ацетил СоА ацетоацетил CoA түзе
конденсирленеді.
Фермент
A. Кетонды денелер синтезі
Екі молекула ацетил СоА ацетоацетил CoA түзе
конденсирленеді.
Фермент
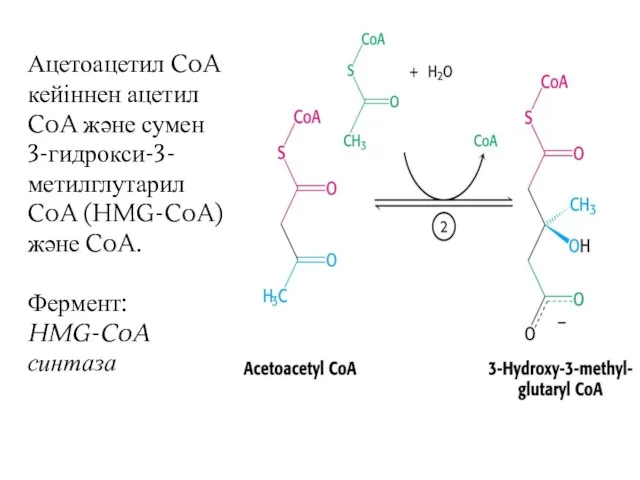
Ацетоацетил CoA кейіннен ацетил CoA және сумен 3-гидрокси-3-метилглутарил CoA (HMG-CoA) және
Ацетоацетил CoA кейіннен ацетил CoA және сумен 3-гидрокси-3-метилглутарил CoA (HMG-CoA) және
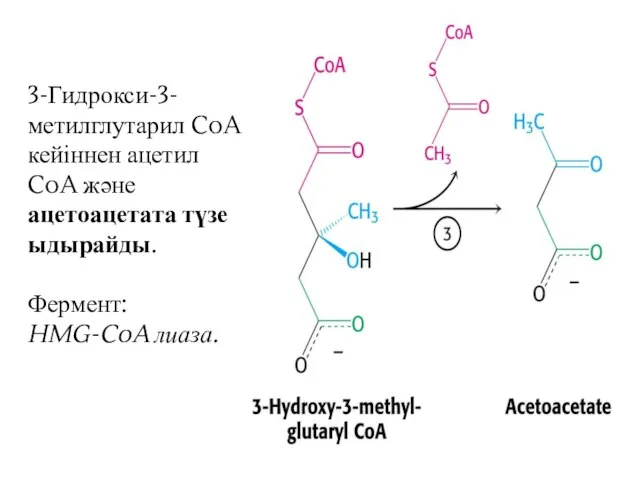
3-Гидрокси-3-метилглутарил CoA кейіннен ацетил CoA және ацетоацетата түзе ыдырайды.
Фермент: HMG-CoA
3-Гидрокси-3-метилглутарил CoA кейіннен ацетил CoA және ацетоацетата түзе ыдырайды.
Фермент: HMG-CoA
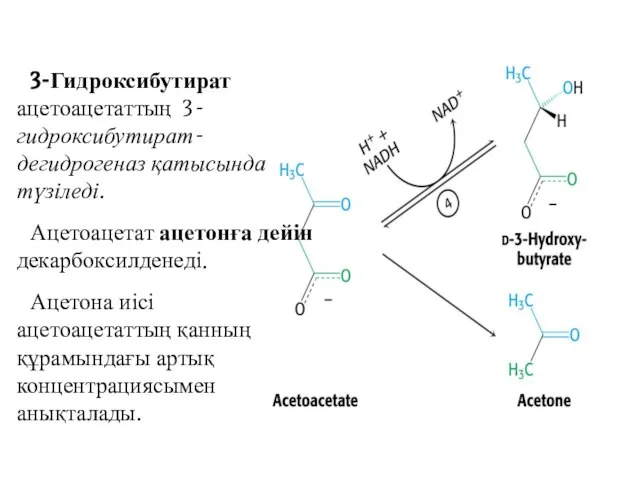
3-Гидроксибутират ацетоацетаттың 3-гидроксибутират-дегидрогеназ қатысында түзіледі.
Ацетоацетат ацетонға дейін декарбоксилденеді.
Ацетона иісі
3-Гидроксибутират ацетоацетаттың 3-гидроксибутират-дегидрогеназ қатысында түзіледі.
Ацетоацетат ацетонға дейін декарбоксилденеді.
Ацетона иісі

B. Кетонды денелер – кейбір ұлпалар үшін «жанармай»көзі
Кетондық денелер бауыр митохондриясынан
B. Кетонды денелер – кейбір ұлпалар үшін «жанармай»көзі
Кетондық денелер бауыр митохондриясынан
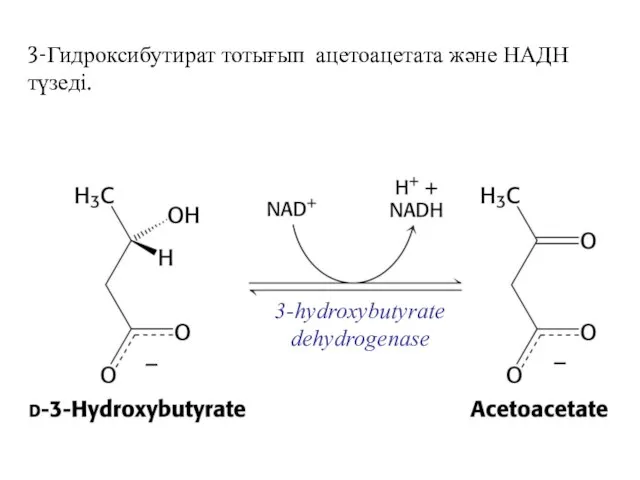
3-Гидроксибутират тотығып ацетоацетата және НАДН түзеді.
3-Гидроксибутират тотығып ацетоацетата және НАДН түзеді.
 Информационные технологии на уроках химии
Информационные технологии на уроках химии Способы переработки нефти
Способы переработки нефти Пластические массы
Пластические массы Правила безпечного використання засобів побутової хімії
Правила безпечного використання засобів побутової хімії  Теория электролитической диссоциации
Теория электролитической диссоциации Тема: «Самородные металлы».
Тема: «Самородные металлы». Энергетика химических процессов
Энергетика химических процессов Вплив фосфатної побутової хімії на організм людини.
Вплив фосфатної побутової хімії на організм людини.  Решение задач по теме «Растворы»
Решение задач по теме «Растворы» Химическая термодинамика и кинетика
Химическая термодинамика и кинетика соединения АЗОТА Материал для повторения и подготовки к ГИА Учитель химии МОУ «Гимназия №1»г. Саратова Шишкина И.Ю.
соединения АЗОТА Материал для повторения и подготовки к ГИА Учитель химии МОУ «Гимназия №1»г. Саратова Шишкина И.Ю. Основные свойства кристаллов
Основные свойства кристаллов Презентация по Химии "Етери та естери" - скачать смотреть бесплатно
Презентация по Химии "Етери та естери" - скачать смотреть бесплатно Поліетелен. Застосування поліетилену
Поліетелен. Застосування поліетилену Гипер-гипофосфатемия
Гипер-гипофосфатемия Ультразвук к в химической технологии. Лекция 5
Ультразвук к в химической технологии. Лекция 5 Класифікація пластмас
Класифікація пластмас  Дисциплина: Химия. Лекция 1. Растворы
Дисциплина: Химия. Лекция 1. Растворы Подготовка к ГИА. Тренажер по химии
Подготовка к ГИА. Тренажер по химии Ферменттер. Зерттелу тарихы
Ферменттер. Зерттелу тарихы Важнейшие кислоты и кислотные остатки. (Леция 1)
Важнейшие кислоты и кислотные остатки. (Леция 1) Углеводы. Строение углеводов
Углеводы. Строение углеводов Презентация по Химии "Горение кислорода" - скачать смотреть
Презентация по Химии "Горение кислорода" - скачать смотреть  Застосування нових сучасних матеріалів з оригінльними властивостями
Застосування нових сучасних матеріалів з оригінльними властивостями  Непредельные углероды. Алкены
Непредельные углероды. Алкены Синтетические моющие средства
Синтетические моющие средства Конденсация. Капельная и пленочная конденсация
Конденсация. Капельная и пленочная конденсация Імунопатологія. Імунопрофілактика і імунотерапія інфекційних захворювань. Засоби імунокорекції
Імунопатологія. Імунопрофілактика і імунотерапія інфекційних захворювань. Засоби імунокорекції