- The molecular basis of inheritance. (Chapter 16)

Содержание
- 2. Overview: Life’s Operating Instructions In 1953, James Watson and Francis Crick introduced an elegant double-helical model
- 3. Figure 16.1
- 4. Concept 16.1: DNA is the genetic material Early in the 20th century, the identification of the
- 5. The Search for the Genetic Material: Scientific Inquiry When T. H. Morgan’s group showed that genes
- 6. Evidence That DNA Can Transform Bacteria The discovery of the genetic role of DNA began with
- 7. When he mixed heat-killed remains of the pathogenic strain with living cells of the harmless strain,
- 8. Living S cells (control) Living R cells (control) Heat-killed S cells (control) Mixture of heat-killed S
- 9. In 1944, Oswald Avery, Maclyn McCarty, and Colin MacLeod announced that the transforming substance was DNA
- 10. Evidence That Viral DNA Can Program Cells More evidence for DNA as the genetic material came
- 11. Animation: Phage T2 Reproductive Cycle Right-click slide / select “Play”
- 12. Figure 16.3 Phage head Tail sheath Tail fiber DNA Bacterial cell 100 nm
- 13. In 1952, Alfred Hershey and Martha Chase performed experiments showing that DNA is the genetic material
- 14. Animation: Hershey-Chase Experiment Right-click slide / select “Play”
- 15. Figure 16.4-1 Bacterial cell Phage Batch 1: Radioactive sulfur (35S) DNA Batch 2: Radioactive phosphorus (32P)
- 16. Figure 16.4-2 Bacterial cell Phage Batch 1: Radioactive sulfur (35S) Radioactive protein DNA Batch 2: Radioactive
- 17. Figure 16.4-3 Bacterial cell Phage Batch 1: Radioactive sulfur (35S) Radioactive protein DNA Batch 2: Radioactive
- 18. Additional Evidence That DNA Is the Genetic Material It was known that DNA is a polymer
- 19. Animation: DNA and RNA Structure Right-click slide / select “Play”
- 20. Two findings became known as Chargaff’s rules The base composition of DNA varies between species In
- 21. Figure 16.5 Sugar–phosphate backbone Nitrogenous bases Thymine (T) Adenine (A) Cytosine (C) Guanine (G) Nitrogenous base
- 22. Building a Structural Model of DNA: Scientific Inquiry After DNA was accepted as the genetic material,
- 23. Figure 16.6 (a) Rosalind Franklin
- 24. Figure 16.6a (a) Rosalind Franklin
- 25. Figure 16.6b
- 26. Franklin’s X-ray crystallographic images of DNA enabled Watson to deduce that DNA was helical The X-ray
- 27. Animation: DNA Double Helix Right-click slide / select “Play”
- 28. Figure 16.7 3.4 nm 1 nm 0.34 nm Hydrogen bond (b) Partial chemical structure 3′ end
- 29. 3.4 nm 1 nm 0.34 nm Hydrogen bond (b) Partial chemical structure 3′ end 5′ end
- 30. Figure 16.7b (c) Space-filling model
- 31. Watson and Crick built models of a double helix to conform to the X-rays and chemistry
- 32. At first, Watson and Crick thought the bases paired like with like (A with A, and
- 33. Figure 16.UN01 Purine + purine: too wide Pyrimidine + pyrimidine: too narrow Purine + pyrimidine: width
- 34. Watson and Crick reasoned that the pairing was more specific, dictated by the base structures They
- 35. Figure 16.8 Sugar Sugar Sugar Sugar Adenine (A) Thymine (T) Guanine (G) Cytosine (C)
- 36. Concept 16.2: Many proteins work together in DNA replication and repair The relationship between structure and
- 37. The Basic Principle: Base Pairing to a Template Strand Since the two strands of DNA are
- 38. Animation: DNA Replication Overview Right-click slide / select “Play”
- 39. Figure 16.9-1 (a) Parent molecule A A A T T T C C G G
- 40. Figure 16.9-2 (a) Parent molecule A A A A A A T T T T T
- 41. Figure 16.9-3 (a) Parent molecule A A A A A A A A A A A
- 42. Watson and Crick’s semiconservative model of replication predicts that when a double helix replicates, each daughter
- 43. Figure 16.10 (c) Dispersive model Parent cell First replication Second replication
- 44. Experiments by Matthew Meselson and Franklin Stahl supported the semiconservative model They labeled the nucleotides of
- 45. The first replication produced a band of hybrid DNA, eliminating the conservative model A second replication
- 46. Figure 16.11 Bacteria cultured in medium with 15N (heavy isotope) Bacteria transferred to medium with 14N
- 47. Figure 16.11a Bacteria cultured in medium with 15N (heavy isotope) Bacteria transferred to medium with 14N
- 48. Figure 16.11b Predictions: First replication Second replication Conservative model Semiconservative model Dispersive model CONCLUSION
- 49. DNA Replication: A Closer Look The copying of DNA is remarkable in its speed and accuracy
- 50. Getting Started Replication begins at particular sites called origins of replication, where the two DNA strands
- 51. Animation: Origins of Replication Right-click slide / select “Play”
- 52. Figure 16.12 (a) Origin of replication in an E. coli cell (b) Origins of replication in
- 53. Figure 16.12a (a) Origin of replication in an E. coli cell Origin of replication Parental (template)
- 54. Figure 16.12b (b) Origins of replication in a eukaryotic cell Origin of replication Double-stranded DNA molecule
- 55. Figure 16.12c 0.5 μm
- 56. Figure 16.12d 0.25 μm
- 57. At the end of each replication bubble is a replication fork, a Y-shaped region where new
- 58. Figure 16.13 Topoisomerase Primase RNA primer Helicase Single-strand binding proteins 5′ 3′ 5′ 5′ 3′ 3′
- 59. DNA polymerases cannot initiate synthesis of a polynucleotide; they can only add nucleotides to the 3′
- 60. An enzyme called primase can start an RNA chain from scratch and adds RNA nucleotides one
- 61. Synthesizing a New DNA Strand Enzymes called DNA polymerases catalyze the elongation of new DNA at
- 62. Each nucleotide that is added to a growing DNA strand is a nucleoside triphosphate dATP supplies
- 63. Figure 16.14 New strand Template strand Sugar Phosphate Base Nucleoside triphosphate DNA polymerase Pyrophosphate 5′ 5′
- 64. Antiparallel Elongation The antiparallel structure of the double helix affects replication DNA polymerases add nucleotides only
- 65. Along one template strand of DNA, the DNA polymerase synthesizes a leading strand continuously, moving toward
- 66. Animation: Leading Strand Right-click slide / select “Play”
- 67. Figure 16.15 Leading strand Lagging strand Overview Origin of replication Lagging strand Leading strand Primer Overall
- 68. Figure 16.15a Leading strand Lagging strand Overview Origin of replication Lagging strand Leading strand Primer Overall
- 69. Origin of replication RNA primer Sliding clamp DNA pol III Parental DNA 3′ 5′ 5′ 3′
- 70. To elongate the other new strand, called the lagging strand, DNA polymerase must work in the
- 71. Animation: Lagging Strand Right-click slide / select “Play”
- 72. Origin of replication Overview Leading strand Leading strand Lagging strand Lagging strand Overall directions of replication
- 73. Figure 16.16a Origin of replication Overview Leading strand Leading strand Lagging strand Lagging strand Overall directions
- 74. Figure 16.16b-1 Template strand 3′ 3′ 5′ 5′
- 75. Figure 16.16b-2 Template strand RNA primer for fragment 1 3′ 3′ 3′ 3′ 5′ 5′ 5′
- 76. Figure 16.16b-3 Template strand RNA primer for fragment 1 Okazaki fragment 1 3′ 3′ 3′ 3′
- 77. Figure 16.16b-4 Template strand RNA primer for fragment 1 Okazaki fragment 1 RNA primer for fragment
- 78. Figure 16.16b-5 Template strand RNA primer for fragment 1 Okazaki fragment 1 RNA primer for fragment
- 79. Figure 16.16b-6 Template strand RNA primer for fragment 1 Okazaki fragment 1 RNA primer for fragment
- 80. Figure 16.17 Overview Leading strand Origin of replication Lagging strand Leading strand Lagging strand Overall directions
- 81. Figure 16.17a Overview Leading strand Origin of replication Lagging strand Leading strand Lagging strand Overall directions
- 82. Overview Leading strand Origin of replication Lagging strand Leading strand Lagging strand Overall directions of replication
- 83. The DNA Replication Complex The proteins that participate in DNA replication form a large complex, a
- 84. Animation: DNA Replication Review Right-click slide / select “Play”
- 85. Figure 16.18 Parental DNA DNA pol III Leading strand Connecting protein Helicase Lagging strand DNA pol
- 86. Proofreading and Repairing DNA DNA polymerases proofread newly made DNA, replacing any incorrect nucleotides In mismatch
- 87. Figure 16.19 Nuclease DNA polymerase DNA ligase 5′ 5′ 5′ 5′ 5′ 5′ 5′ 5′ 3′
- 88. Evolutionary Significance of Altered DNA Nucleotides Error rate after proofreading repair is low but not zero
- 89. Replicating the Ends of DNA Molecules Limitations of DNA polymerase create problems for the linear DNA
- 90. Figure 16.20 Ends of parental DNA strands Leading strand Lagging strand Last fragment Next-to-last fragment Lagging
- 91. Figure 16.20a Ends of parental DNA strands Leading strand Lagging strand Last fragment Next-to-last fragment Lagging
- 92. Figure 16.20b Second round of replication Further rounds of replication New leading strand New lagging strand
- 93. Eukaryotic chromosomal DNA molecules have special nucleotide sequences at their ends called telomeres Telomeres do not
- 94. Figure 16.21 1 μm
- 95. If chromosomes of germ cells became shorter in every cell cycle, essential genes would eventually be
- 96. The shortening of telomeres might protect cells from cancerous growth by limiting the number of cell
- 97. Concept 16.3 A chromosome consists of a DNA molecule packed together with proteins The bacterial chromosome
- 98. Chromatin, a complex of DNA and protein, is found in the nucleus of eukaryotic cells Chromosomes
- 99. Animation: DNA Packing Right-click slide / select “Play”
- 100. Figure 16.22a DNA double helix (2 nm in diameter) DNA, the double helix Nucleosome (10 nm
- 101. Figure 16.22b 30-nm fiber 30-nm fiber Loops Scaffold 300-nm fiber Chromatid (700 nm) Replicated chromosome (1,400
- 102. Figure 16.22c DNA double helix (2 nm in diameter)
- 103. Figure 16.22d Nucleosome (10 nm in diameter)
- 104. Figure 16.22e 30-nm fiber
- 105. Figure 16.22f Loops Scaffold
- 106. Figure 16.22g Chromatid (700 nm)
- 107. Chromatin undergoes changes in packing during the cell cycle At interphase, some chromatin is organized into
- 108. Figure 16.23 5 μm
- 109. Figure 16.23a
- 110. Figure 16.23b
- 111. Figure 16.23c 5 μm
- 112. Most chromatin is loosely packed in the nucleus during interphase and condenses prior to mitosis Loosely
- 113. Histones can undergo chemical modifications that result in changes in chromatin organization
- 114. Figure 16.UN02 Sugar-phosphate backbone Nitrogenous bases Hydrogen bond G G G G C C C C
- 115. Figure 16.UN03 DNA pol III synthesizes leading strand continuously Parental DNA DNA pol III starts DNA
- 116. Figure 16.UN04
- 117. Figure 16.UN05
- 118. Figure 16.UN06
- 120. Скачать презентацию

Overview: Life’s Operating Instructions
In 1953, James Watson and Francis Crick introduced
Overview: Life’s Operating Instructions
In 1953, James Watson and Francis Crick introduced

Figure 16.1
Figure 16.1

Concept 16.1: DNA is the genetic material
Early in the 20th century,
Concept 16.1: DNA is the genetic material
Early in the 20th century,

The Search for the Genetic Material: Scientific Inquiry
When T. H. Morgan’s
The Search for the Genetic Material: Scientific Inquiry
When T. H. Morgan’s

Evidence That DNA Can Transform Bacteria
The discovery of the genetic role
Evidence That DNA Can Transform Bacteria
The discovery of the genetic role

When he mixed heat-killed remains of the pathogenic strain with living
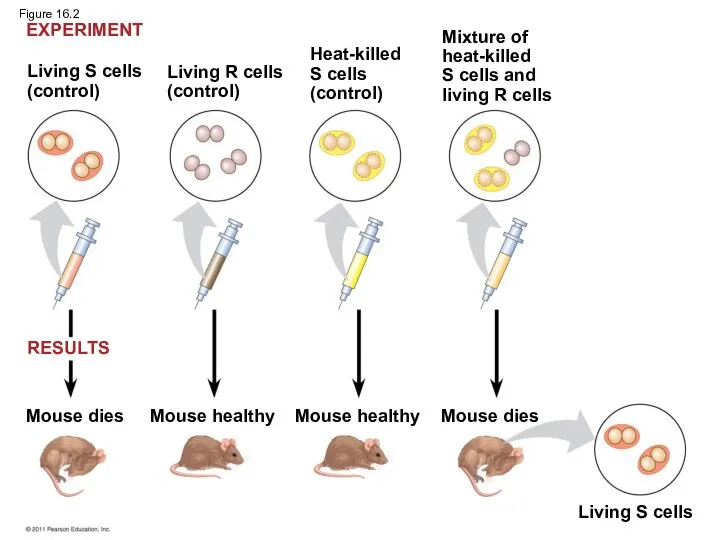
Living S cells
(control)
Living R cells
(control)
Heat-killed
S cells
(control)
Mixture of
heat-killed
S cells and
living R cells
Mouse
Living S cells
(control)
Living R cells
(control)
Heat-killed
S cells
(control)
Mixture of
heat-killed
S cells and
living R cells
Mouse

In 1944, Oswald Avery, Maclyn McCarty, and Colin MacLeod announced that
In 1944, Oswald Avery, Maclyn McCarty, and Colin MacLeod announced that

Evidence That Viral DNA Can Program Cells
More evidence for DNA as
Evidence That Viral DNA Can Program Cells
More evidence for DNA as

Animation: Phage T2 Reproductive Cycle Right-click slide / select “Play”
Animation: Phage T2 Reproductive Cycle Right-click slide / select “Play”
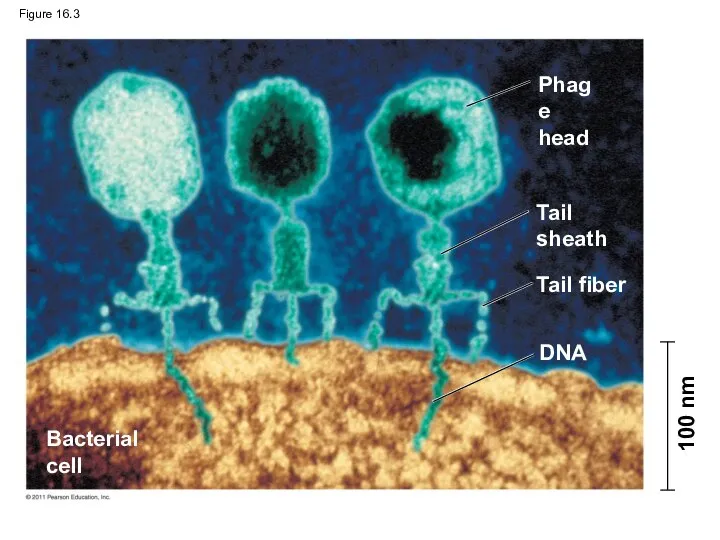
Figure 16.3
Phage
head
Tail
sheath
Tail fiber
DNA
Bacterial
cell
100 nm
Figure 16.3
Phage
head
Tail
sheath
Tail fiber
DNA
Bacterial
cell
100 nm

In 1952, Alfred Hershey and Martha Chase performed experiments showing that
In 1952, Alfred Hershey and Martha Chase performed experiments showing that

Animation: Hershey-Chase Experiment
Right-click slide / select “Play”
Animation: Hershey-Chase Experiment
Right-click slide / select “Play”
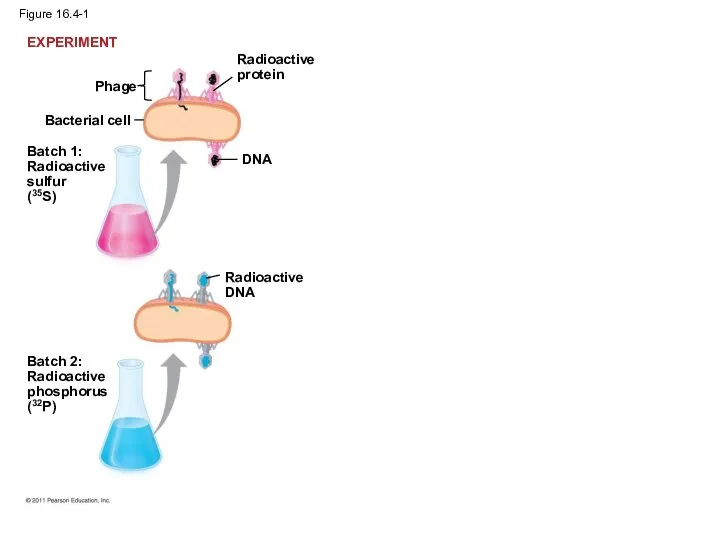
Figure 16.4-1
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
EXPERIMENT
Radioactive
protein
Figure 16.4-1
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
EXPERIMENT
Radioactive
protein
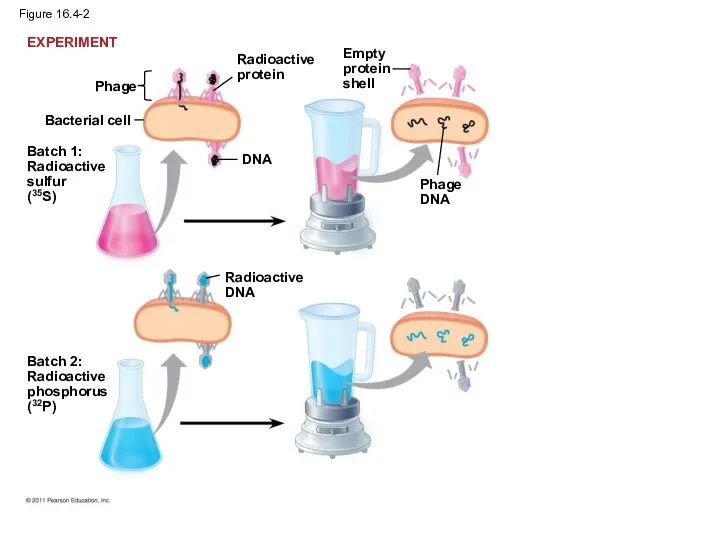
Figure 16.4-2
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
Radioactive
protein
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
Empty
protein
shell
Phage
DNA
EXPERIMENT
Figure 16.4-2
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
Radioactive
protein
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
Empty
protein
shell
Phage
DNA
EXPERIMENT
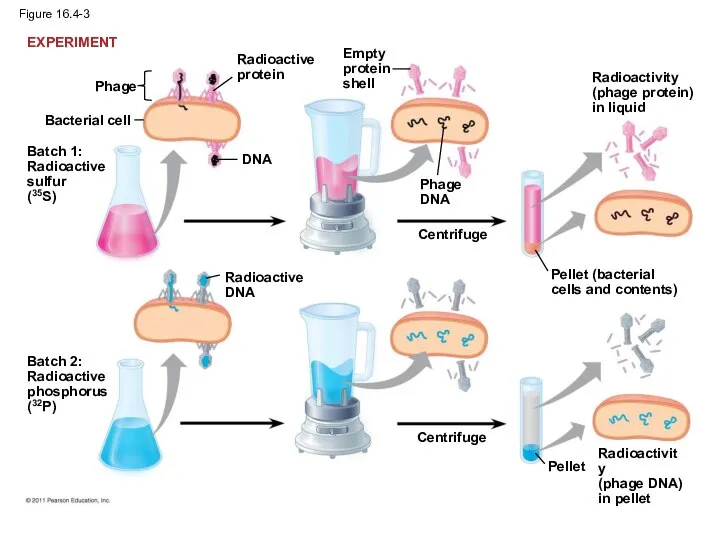
Figure 16.4-3
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
Radioactive
protein
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
Empty
protein
shell
Phage
DNA
Centrifuge
Centrifuge
Radioactivity
(phage protein)
in liquid
Pellet (bacterial
cells and contents)
Pellet
Radioactivity
(phage DNA)
in
Figure 16.4-3
Bacterial cell
Phage
Batch 1:
Radioactive
sulfur
(35S)
Radioactive
protein
DNA
Batch 2:
Radioactive
phosphorus
(32P)
Radioactive
DNA
Empty
protein
shell
Phage
DNA
Centrifuge
Centrifuge
Radioactivity
(phage protein)
in liquid
Pellet (bacterial
cells and contents)
Pellet
Radioactivity (phage DNA) in

Additional Evidence That DNA Is the Genetic Material
It was known that
Additional Evidence That DNA Is the Genetic Material
It was known that
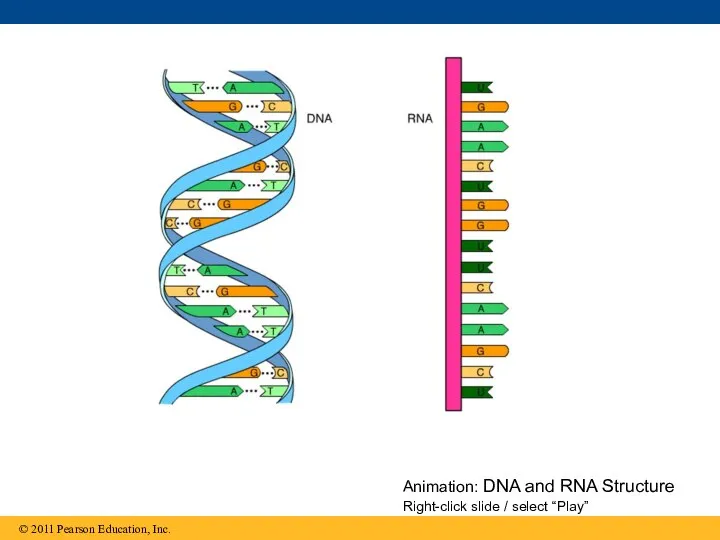
Animation: DNA and RNA Structure Right-click slide / select “Play”
Animation: DNA and RNA Structure Right-click slide / select “Play”

Two findings became known as Chargaff’s rules
The base composition of DNA
Two findings became known as Chargaff’s rules
The base composition of DNA
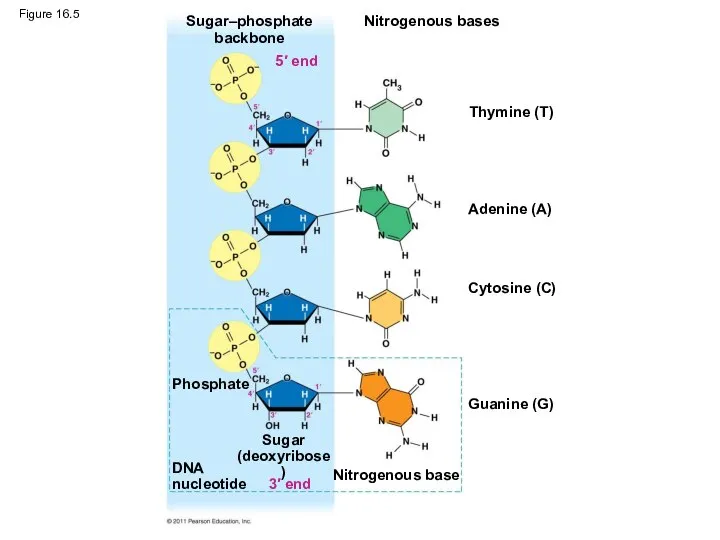
Figure 16.5
Sugar–phosphate
backbone
Nitrogenous bases
Thymine (T)
Adenine (A)
Cytosine (C)
Guanine (G)
Nitrogenous base
Phosphate
DNA
nucleotide
Sugar
(deoxyribose)
3′ end
5′ end
Figure 16.5
Sugar–phosphate
backbone
Nitrogenous bases
Thymine (T)
Adenine (A)
Cytosine (C)
Guanine (G)
Nitrogenous base
Phosphate
DNA
nucleotide
Sugar
(deoxyribose)
3′ end
5′ end

Building a Structural Model of DNA: Scientific Inquiry
After DNA was accepted
Building a Structural Model of DNA: Scientific Inquiry
After DNA was accepted

Figure 16.6
(a) Rosalind Franklin
Figure 16.6
(a) Rosalind Franklin

Figure 16.6a
(a) Rosalind Franklin
Figure 16.6a
(a) Rosalind Franklin
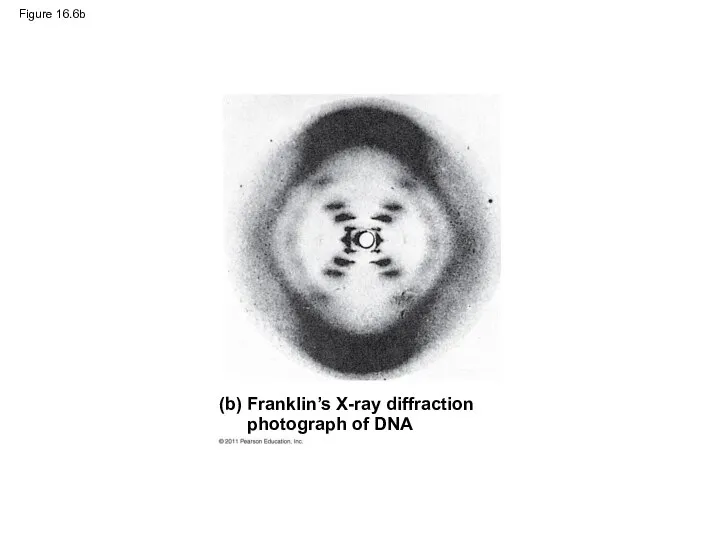
Figure 16.6b
Figure 16.6b

Franklin’s X-ray crystallographic images of DNA enabled Watson to deduce that
Franklin’s X-ray crystallographic images of DNA enabled Watson to deduce that

Animation: DNA Double Helix Right-click slide / select “Play”
Animation: DNA Double Helix Right-click slide / select “Play”
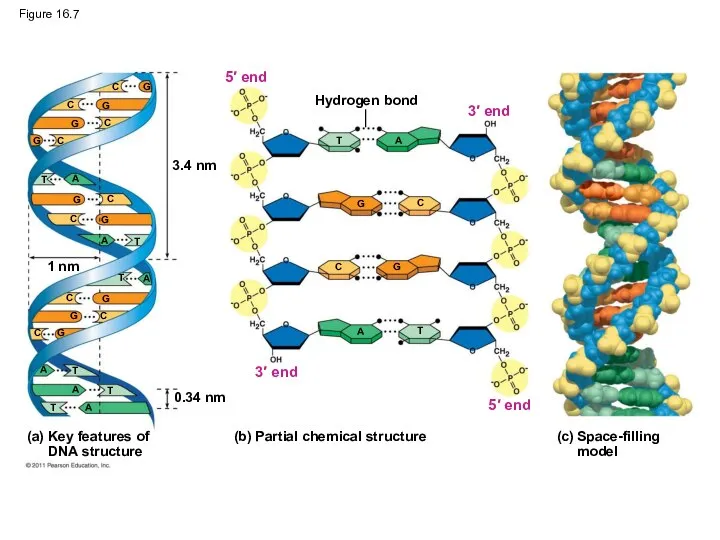
Figure 16.7
3.4 nm
1 nm
0.34 nm
Hydrogen bond
(b) Partial chemical structure
3′ end
5′ end
3′
Figure 16.7
3.4 nm
1 nm
0.34 nm
Hydrogen bond
(b) Partial chemical structure
3′ end
5′ end
3′
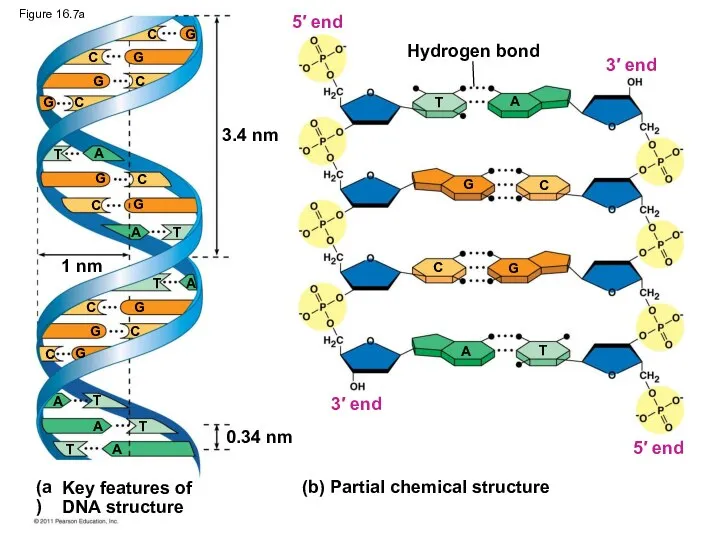
3.4 nm
1 nm
0.34 nm
Hydrogen bond
(b) Partial chemical structure
3′ end
5′ end
3′ end
5′
3.4 nm
1 nm
0.34 nm
Hydrogen bond
(b) Partial chemical structure
3′ end
5′ end
3′ end
5′
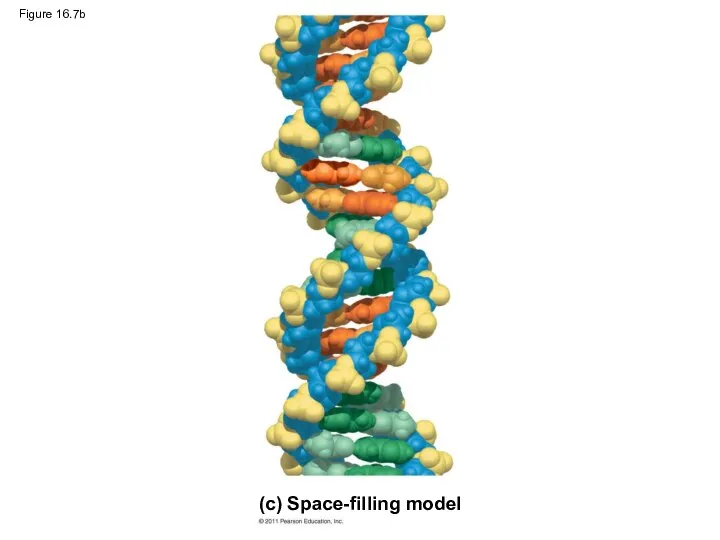
Figure 16.7b
(c) Space-filling model
Figure 16.7b
(c) Space-filling model

Watson and Crick built models of a double helix to conform
Watson and Crick built models of a double helix to conform

At first, Watson and Crick thought the bases paired like with
At first, Watson and Crick thought the bases paired like with
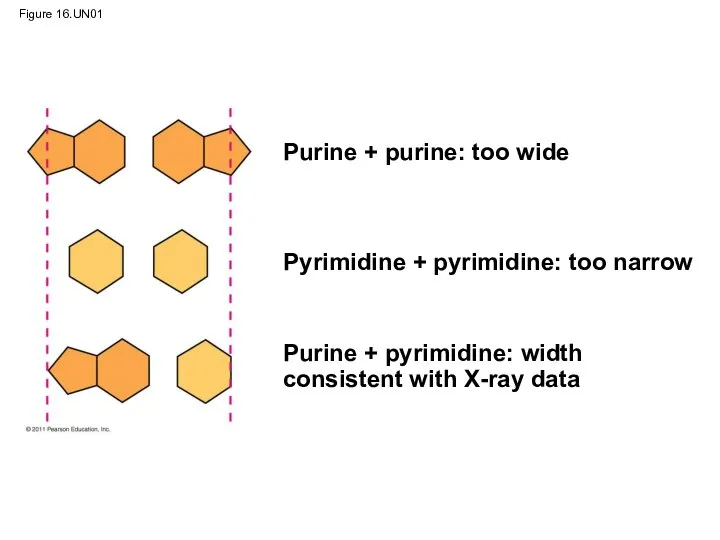
Figure 16.UN01
Purine + purine: too wide
Pyrimidine + pyrimidine: too narrow
Purine +
Figure 16.UN01
Purine + purine: too wide
Pyrimidine + pyrimidine: too narrow
Purine +

Watson and Crick reasoned that the pairing was more specific, dictated
Watson and Crick reasoned that the pairing was more specific, dictated
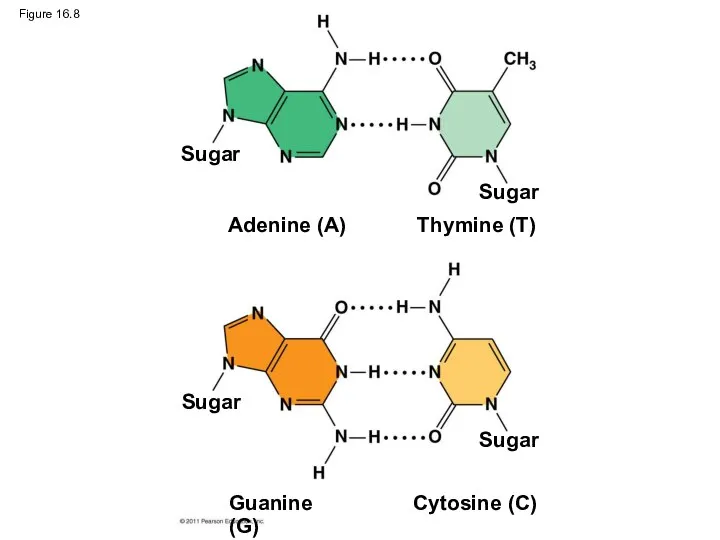
Figure 16.8
Sugar
Sugar
Sugar
Sugar
Adenine (A)
Thymine (T)
Guanine (G)
Cytosine (C)
Figure 16.8
Sugar
Sugar
Sugar
Sugar
Adenine (A)
Thymine (T)
Guanine (G)
Cytosine (C)

Concept 16.2: Many proteins work together in DNA replication and repair
The
Concept 16.2: Many proteins work together in DNA replication and repair
The

The Basic Principle: Base Pairing to a Template Strand
Since the two
The Basic Principle: Base Pairing to a Template Strand
Since the two
Animation: DNA Replication Overview
Right-click slide / select “Play”
Animation: DNA Replication Overview
Right-click slide / select “Play”
Figure 16.9-1
(a) Parent molecule
A
A
A
T
T
T
C
C
G
G
Figure 16.9-1
(a) Parent molecule
A
A
A
T
T
T
C
C
G
G
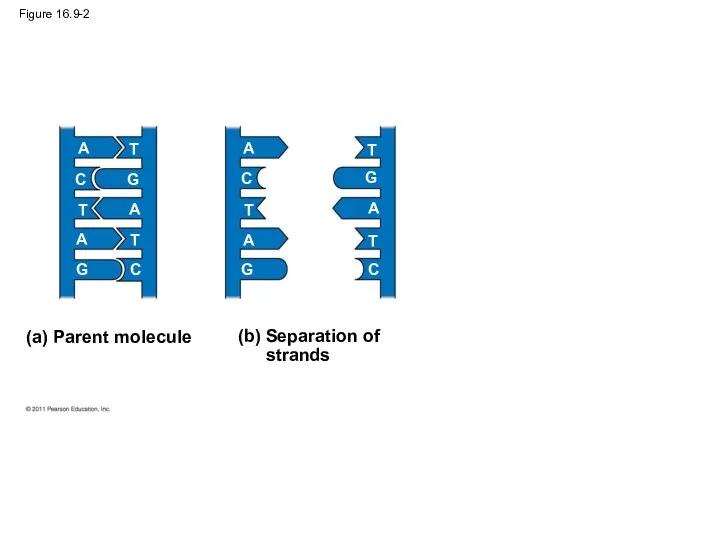
Figure 16.9-2
(a) Parent molecule
A
A
A
A
A
A
T
T
T
T
T
T
C
C
C
C
G
G
G
G
Figure 16.9-2
(a) Parent molecule
A
A
A
A
A
A
T
T
T
T
T
T
C
C
C
C
G
G
G
G
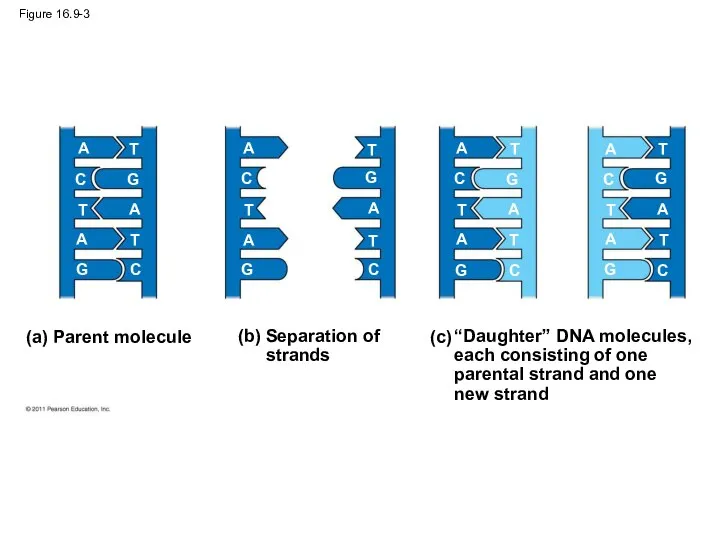
Figure 16.9-3
(a) Parent molecule
A
A
A
A
A
A
A
A
A
A
A
A
T
T
T
T
T
T
T
T
T
T
T
T
C
C
C
C
C
C
C
C
G
G
G
G
G
G
G
G
Figure 16.9-3
(a) Parent molecule
A
A
A
A
A
A
A
A
A
A
A
A
T
T
T
T
T
T
T
T
T
T
T
T
C
C
C
C
C
C
C
C
G
G
G
G
G
G
G
G

Watson and Crick’s semiconservative model of replication predicts that when a
Watson and Crick’s semiconservative model of replication predicts that when a
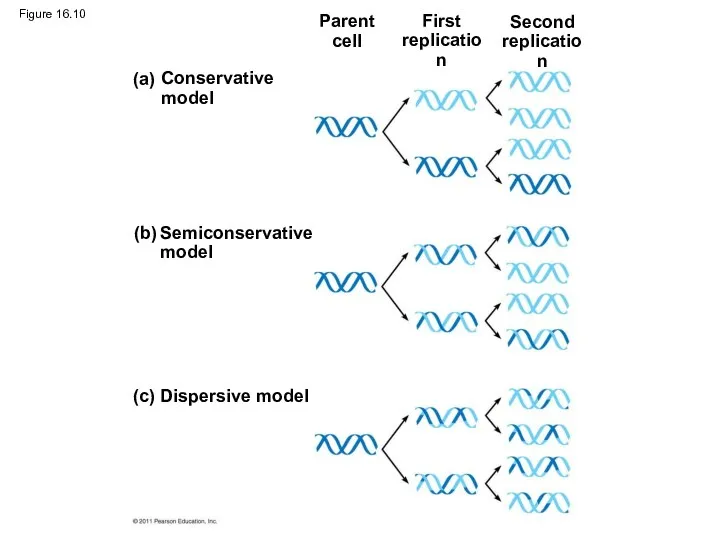
Figure 16.10
(c) Dispersive model
Parent
cell
First
replication
Second
replication
Figure 16.10
(c) Dispersive model
Parent
cell
First
replication
Second
replication

Experiments by Matthew Meselson and Franklin Stahl supported the semiconservative model
Experiments by Matthew Meselson and Franklin Stahl supported the semiconservative model

The first replication produced a band of hybrid DNA, eliminating the
The first replication produced a band of hybrid DNA, eliminating the
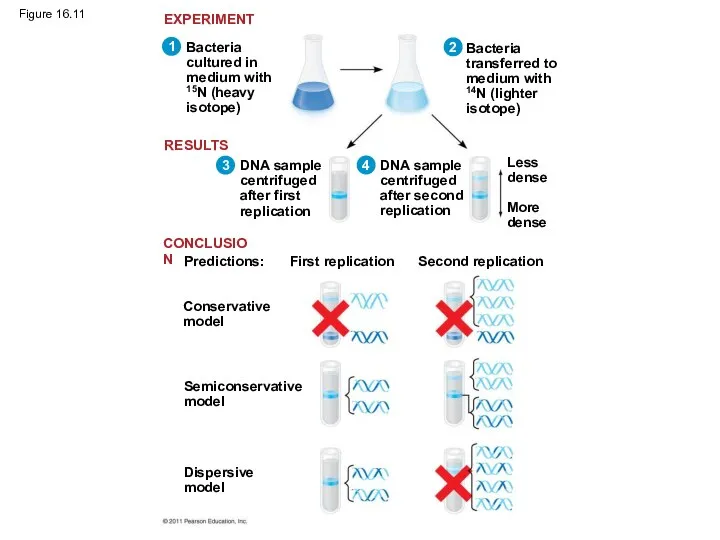
Figure 16.11
Bacteria
cultured in
medium with
15N (heavy
isotope)
Bacteria
transferred to
medium with
14N (lighter
isotope)
DNA sample
centrifuged
after first
replication
DNA sample
centrifuged
after
Figure 16.11
Bacteria
cultured in
medium with
15N (heavy
isotope)
Bacteria
transferred to
medium with
14N (lighter
isotope)
DNA sample
centrifuged
after first
replication
DNA sample centrifuged after
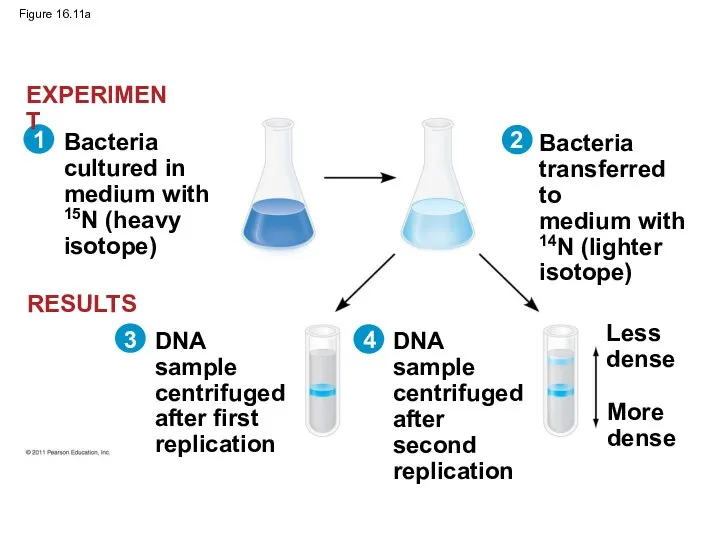
Figure 16.11a
Bacteria
cultured in
medium with
15N (heavy
isotope)
Bacteria
transferred to
medium with
14N (lighter
isotope)
DNA sample
centrifuged
after first
replication
DNA sample
centrifuged
after
Figure 16.11a
Bacteria
cultured in
medium with
15N (heavy
isotope)
Bacteria
transferred to
medium with
14N (lighter
isotope)
DNA sample
centrifuged
after first
replication
DNA sample centrifuged after
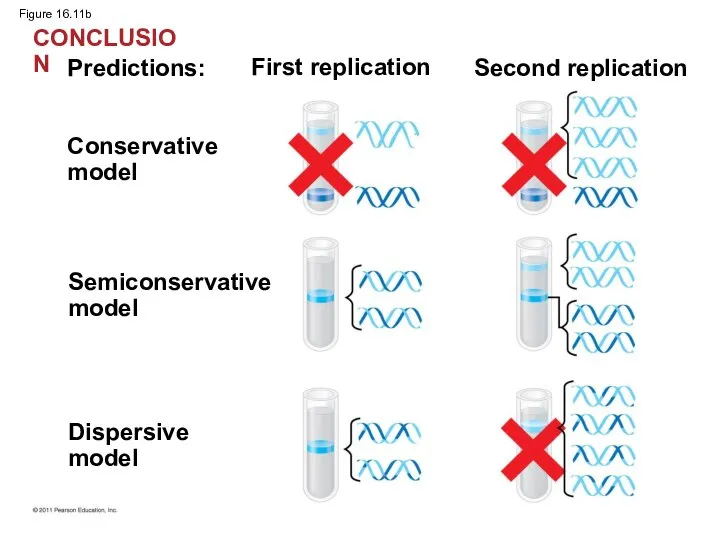
Figure 16.11b
Predictions:
First replication
Second replication
Conservative
model
Semiconservative
model
Dispersive
model
CONCLUSION
Figure 16.11b
Predictions:
First replication
Second replication
Conservative
model
Semiconservative
model
Dispersive
model
CONCLUSION

DNA Replication: A Closer Look
The copying of DNA is remarkable in
DNA Replication: A Closer Look
The copying of DNA is remarkable in

Getting Started
Replication begins at particular sites called origins of replication, where
Getting Started
Replication begins at particular sites called origins of replication, where

Animation: Origins of Replication
Right-click slide / select “Play”
Animation: Origins of Replication
Right-click slide / select “Play”
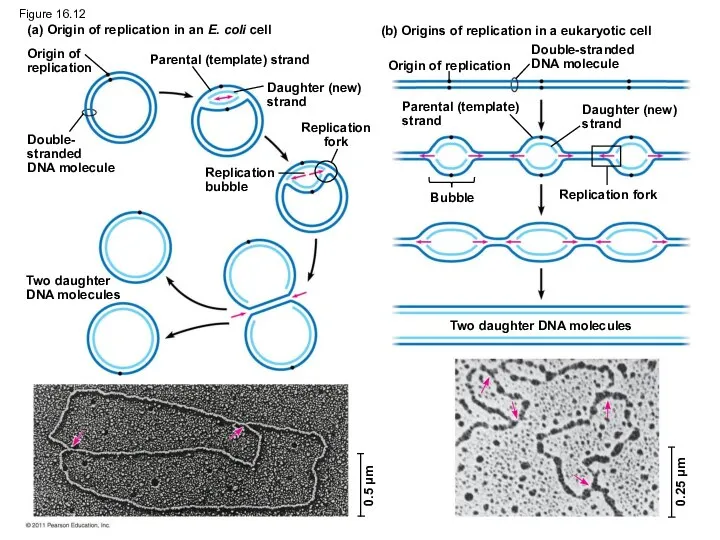
Figure 16.12
(a) Origin of replication in an E. coli cell
(b) Origins
Figure 16.12
(a) Origin of replication in an E. coli cell
(b) Origins

Figure 16.12a
(a) Origin of replication in an E. coli cell
Origin of
replication
Parental
Figure 16.12a
(a) Origin of replication in an E. coli cell
Origin of
replication
Parental
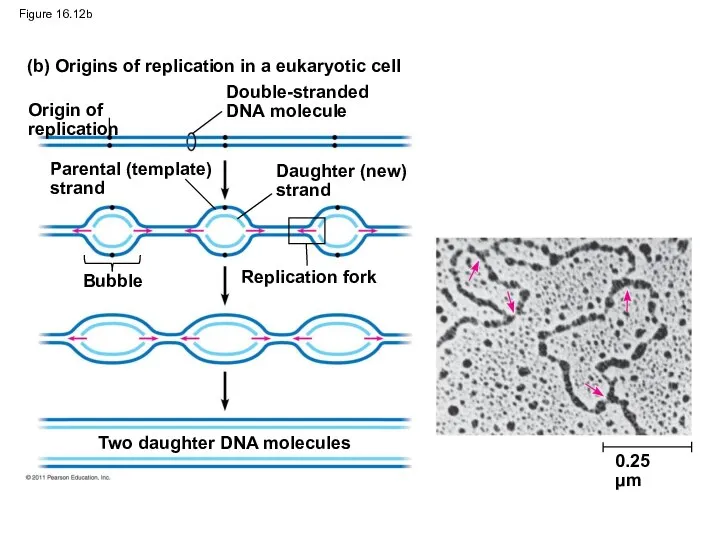
Figure 16.12b
(b) Origins of replication in a eukaryotic cell
Origin of replication
Double-stranded
DNA
Figure 16.12b
(b) Origins of replication in a eukaryotic cell
Origin of replication
Double-stranded DNA

Figure 16.12c
0.5 μm
Figure 16.12c
0.5 μm
Figure 16.12d
0.25 μm
Figure 16.12d
0.25 μm

At the end of each replication bubble is a replication fork,
At the end of each replication bubble is a replication fork,
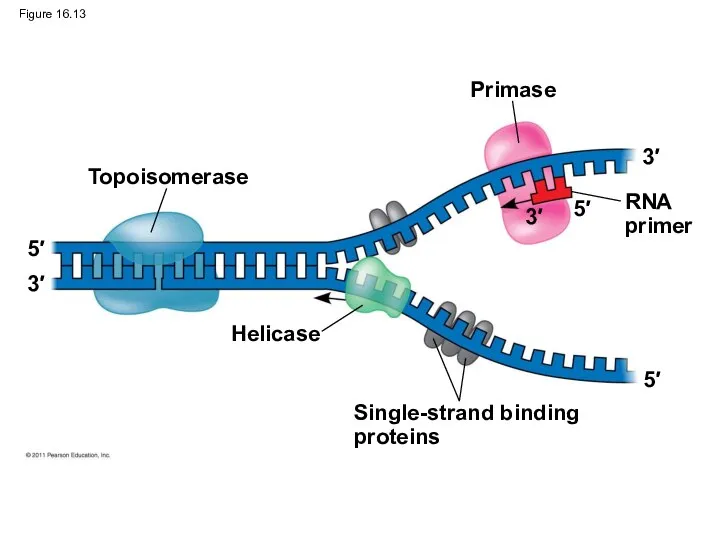
Figure 16.13
Topoisomerase
Primase
RNA
primer
Helicase
Single-strand binding
proteins
5′
3′
5′
5′
3′
3′
Figure 16.13
Topoisomerase
Primase
RNA
primer
Helicase
Single-strand binding
proteins
5′
3′
5′
5′
3′
3′

DNA polymerases cannot initiate synthesis of a polynucleotide; they can only
DNA polymerases cannot initiate synthesis of a polynucleotide; they can only

An enzyme called primase can start an RNA chain from scratch
An enzyme called primase can start an RNA chain from scratch

Synthesizing a New DNA Strand
Enzymes called DNA polymerases catalyze the elongation
Synthesizing a New DNA Strand
Enzymes called DNA polymerases catalyze the elongation

Each nucleotide that is added to a growing DNA strand is
Each nucleotide that is added to a growing DNA strand is
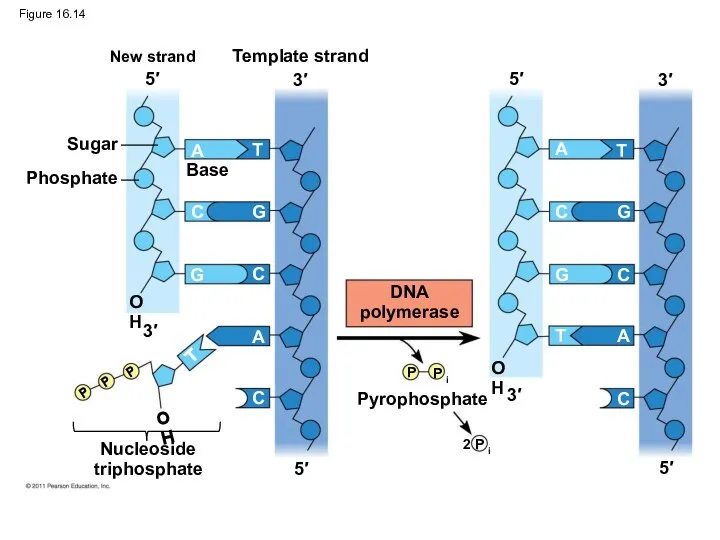
Figure 16.14
New strand
Template strand
Sugar
Phosphate
Base
Nucleoside
triphosphate
DNA
polymerase
Pyrophosphate
5′
5′
5′
5′
3′
3′
3′
3′
OH
OH
OH
P
P i
2 P i
P
P
P
A
A
A
A
T
T
T
T
C
C
C
C
C
C
G
G
G
G
Figure 16.14
New strand
Template strand
Sugar
Phosphate
Base
Nucleoside
triphosphate
DNA
polymerase
Pyrophosphate
5′
5′
5′
5′
3′
3′
3′
3′
OH
OH
OH
P
P i
2 P i
P
P
P
A
A
A
A
T
T
T
T
C
C
C
C
C
C
G
G
G
G

Antiparallel Elongation
The antiparallel structure of the double helix affects replication
DNA polymerases
Antiparallel Elongation
The antiparallel structure of the double helix affects replication
DNA polymerases

Along one template strand of DNA, the DNA polymerase synthesizes a
Along one template strand of DNA, the DNA polymerase synthesizes a

Animation: Leading Strand Right-click slide / select “Play”
Animation: Leading Strand Right-click slide / select “Play”
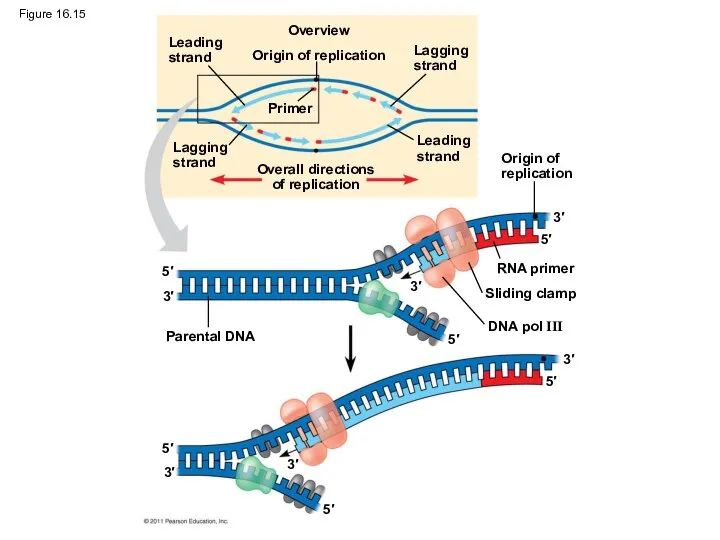
Figure 16.15
Leading
strand
Lagging
strand
Overview
Origin of replication
Lagging
strand
Leading
strand
Primer
Overall directions
of replication
Origin of
replication
RNA primer
Sliding clamp
DNA pol
Figure 16.15
Leading
strand
Lagging
strand
Overview
Origin of replication
Lagging
strand
Leading
strand
Primer
Overall directions
of replication
Origin of
replication
RNA primer
Sliding clamp
DNA pol
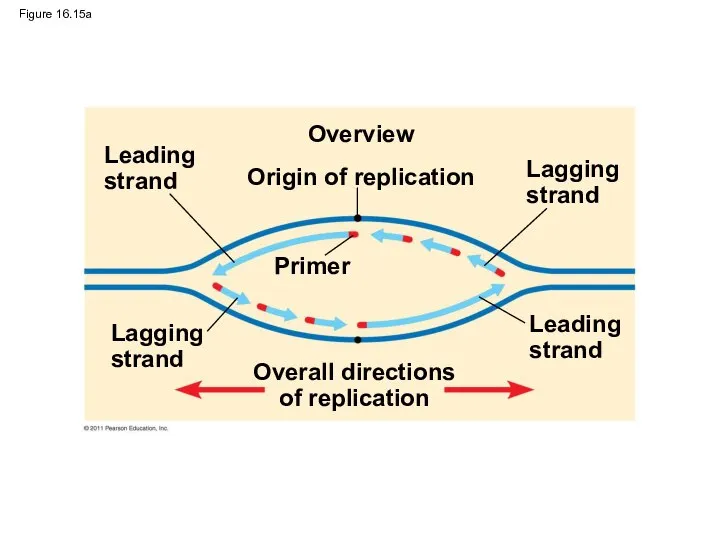
Figure 16.15a
Leading
strand
Lagging
strand
Overview
Origin of replication
Lagging
strand
Leading
strand
Primer
Overall directions
of replication
Figure 16.15a
Leading
strand
Lagging
strand
Overview
Origin of replication
Lagging
strand
Leading
strand
Primer
Overall directions
of replication
Origin of
replication
RNA primer
Sliding clamp
DNA pol III
Parental DNA
3′
5′
5′
3′
3′
5′
3′
5′
3′
5′
3′
5′
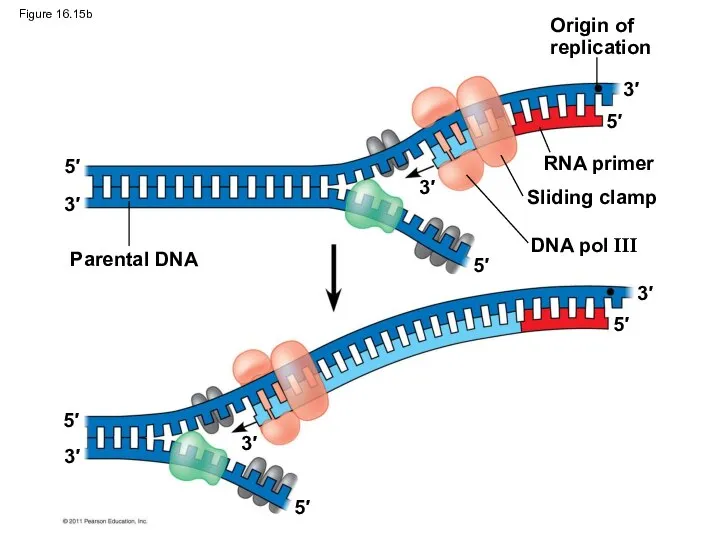
Figure 16.15b
Origin of
replication
RNA primer
Sliding clamp
DNA pol III
Parental DNA
3′
5′
5′
3′
3′
5′
3′
5′
3′
5′
3′
5′
Figure 16.15b

To elongate the other new strand, called the lagging strand, DNA
To elongate the other new strand, called the lagging strand, DNA

Animation: Lagging Strand Right-click slide / select “Play”
Animation: Lagging Strand Right-click slide / select “Play”
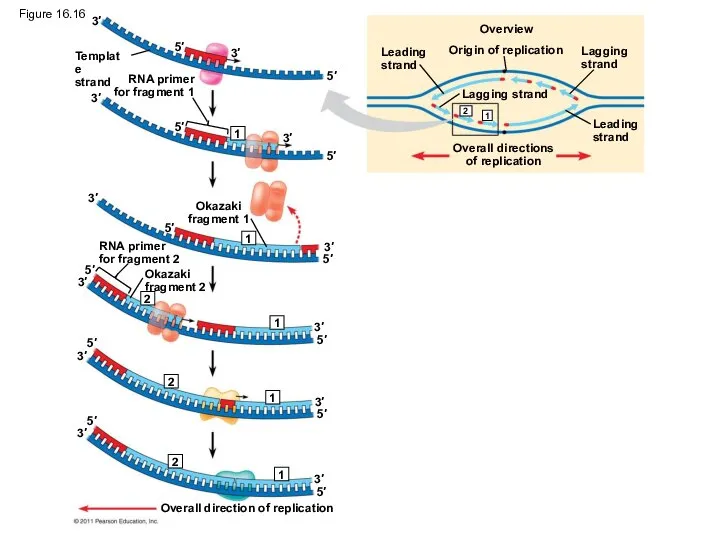
Origin of replication
Overview
Leading
strand
Leading
strand
Lagging
strand
Lagging strand
Overall directions
of replication
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for
Origin of replication
Overview
Leading
strand
Leading
strand
Lagging
strand
Lagging strand
Overall directions
of replication
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer for
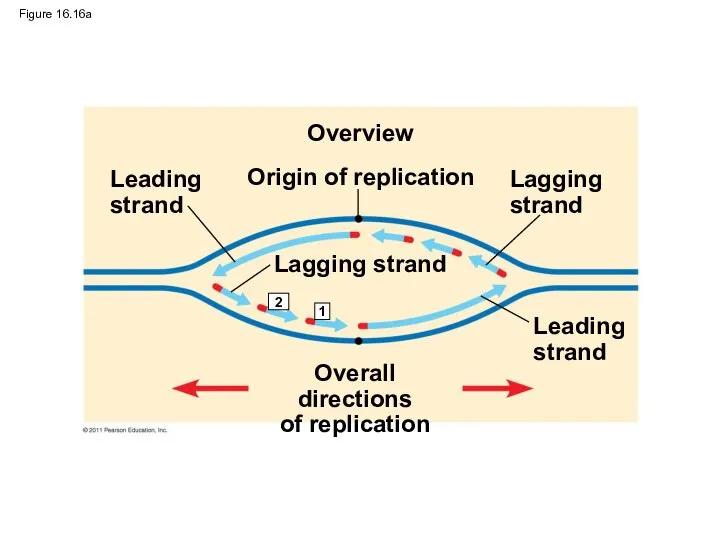
Figure 16.16a
Origin of replication
Overview
Leading
strand
Leading
strand
Lagging
strand
Lagging strand
Overall directions
of replication
1
2
Figure 16.16a
Origin of replication
Overview
Leading
strand
Leading
strand
Lagging
strand
Lagging strand
Overall directions
of replication
1
2

Figure 16.16b-1
Template
strand
3′
3′
5′
5′
Figure 16.16b-1
Template
strand
3′
3′
5′
5′

Figure 16.16b-2
Template
strand
RNA primer
for fragment 1
3′
3′
3′
3′
5′
5′
5′
5′
1
Figure 16.16b-2
Template
strand
RNA primer
for fragment 1
3′
3′
3′
3′
5′
5′
5′
5′
1
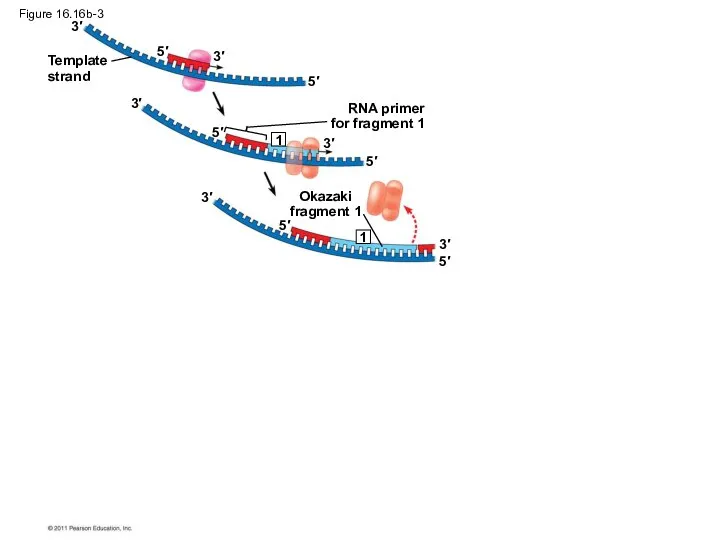
Figure 16.16b-3
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
1
1
Figure 16.16b-3
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
1
1
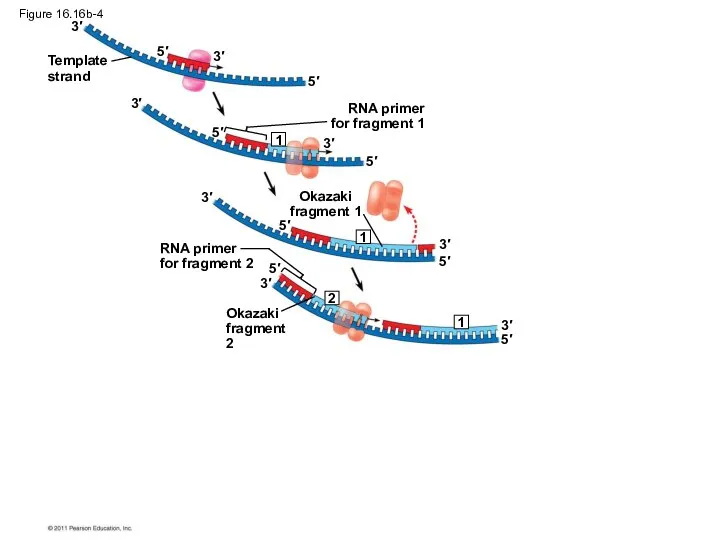
Figure 16.16b-4
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
3′
3′
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
5′
5′
2
1
1
1
Figure 16.16b-4
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
3′
3′
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
5′
5′
2
1
1
1
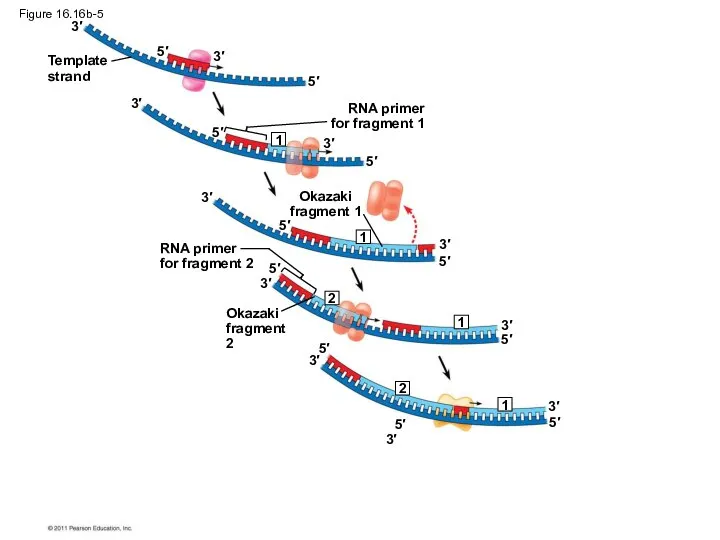
Figure 16.16b-5
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
3′
3′
3′
3′
3′
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
5′
5′
5′
5′
5′
2
2
1
1
1
1
Figure 16.16b-5
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
3′
3′
3′
3′
3′
3′
3′
3′
3′
3′
3′
5′
5′
5′
5′
5′
5′
5′
5′
5′
5′
5′
2
2
1
1
1
1
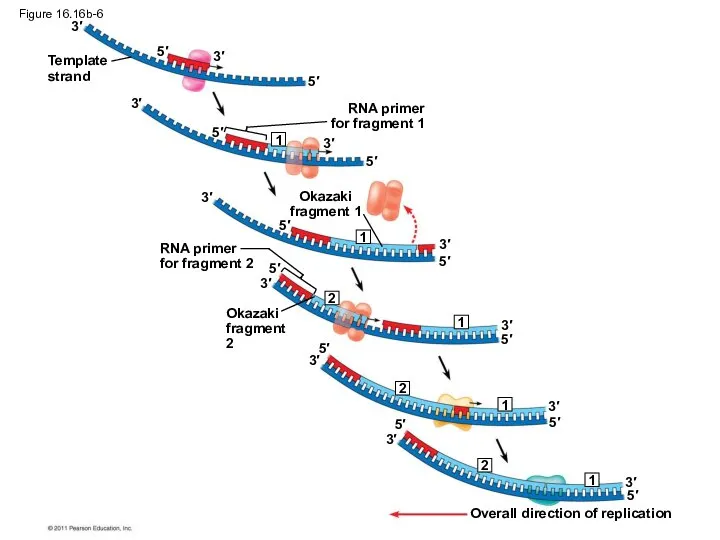
Figure 16.16b-6
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
Overall direction
Figure 16.16b-6
Template
strand
RNA primer
for fragment 1
Okazaki
fragment 1
RNA primer
for fragment 2
Okazaki
fragment 2
Overall direction
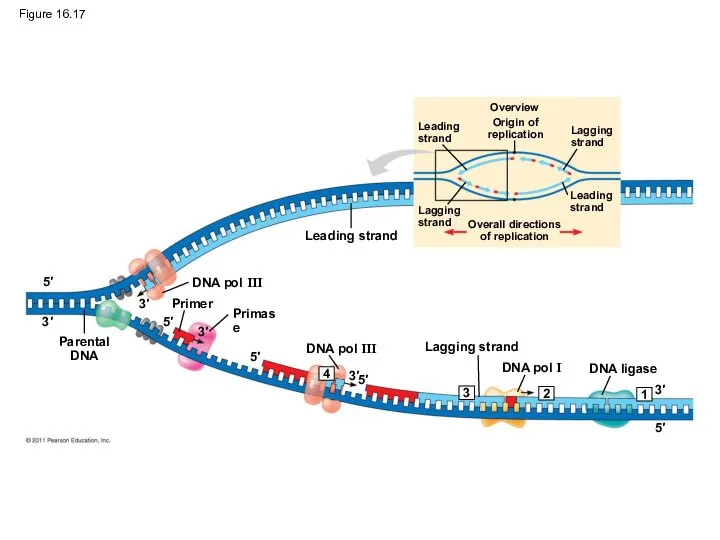
Figure 16.17
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
DNA pol III
DNA pol III
Lagging
Figure 16.17
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
DNA pol III
DNA pol III
Lagging
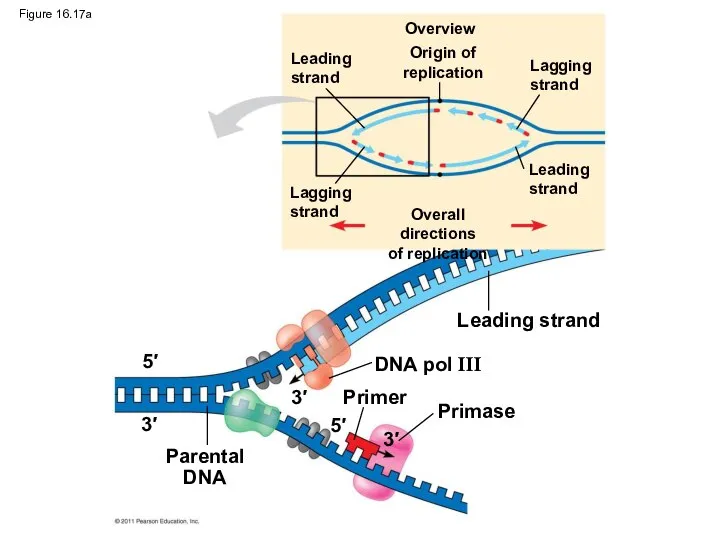
Figure 16.17a
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
DNA pol III
Primer
Primase
Parental
DNA
5′
5′
3′
3′
3′
Figure 16.17a
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
DNA pol III
Primer
Primase
Parental
DNA
5′
5′
3′
3′
3′
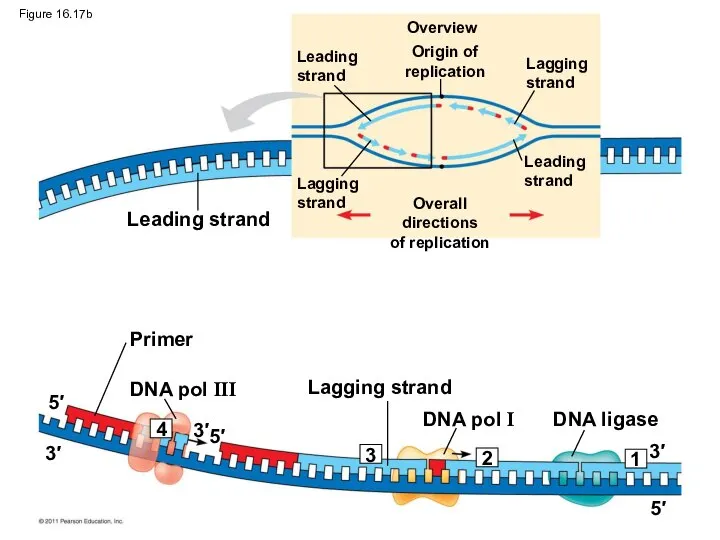
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
Primer
DNA pol III
DNA pol I
Lagging strand
DNA
Overview
Leading
strand
Origin of
replication
Lagging
strand
Leading
strand
Lagging
strand
Overall directions
of replication
Leading strand
Primer
DNA pol III
DNA pol I
Lagging strand
DNA

The DNA Replication Complex
The proteins that participate in DNA replication form
The DNA Replication Complex
The proteins that participate in DNA replication form

Animation: DNA Replication Review
Right-click slide / select “Play”
Animation: DNA Replication Review
Right-click slide / select “Play”
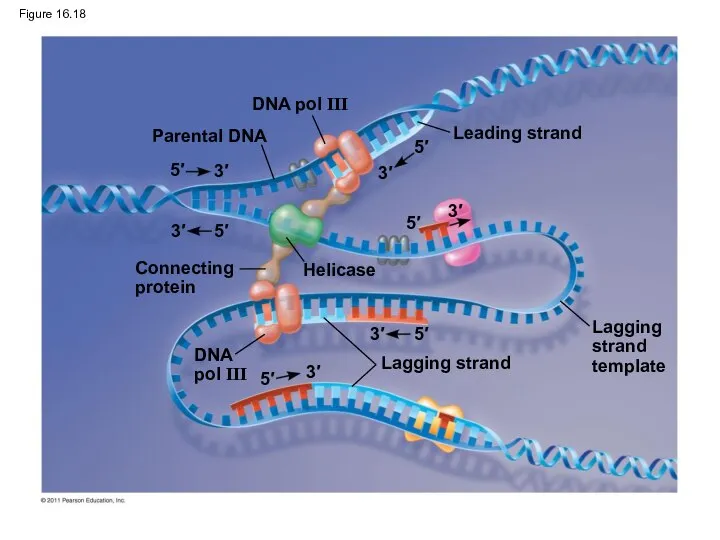
Figure 16.18
Parental DNA
DNA pol III
Leading strand
Connecting
protein
Helicase
Lagging strand
DNA
pol III
Lagging
strand
template
5′
5′
5′
5′
5′
5′
3′
3′
3′
3′
3′
3′
Figure 16.18
Parental DNA
DNA pol III
Leading strand
Connecting
protein
Helicase
Lagging strand
DNA
pol III
Lagging
strand
template
5′
5′
5′
5′
5′
5′
3′
3′
3′
3′
3′
3′

Proofreading and Repairing DNA
DNA polymerases proofread newly made DNA, replacing any
Proofreading and Repairing DNA
DNA polymerases proofread newly made DNA, replacing any
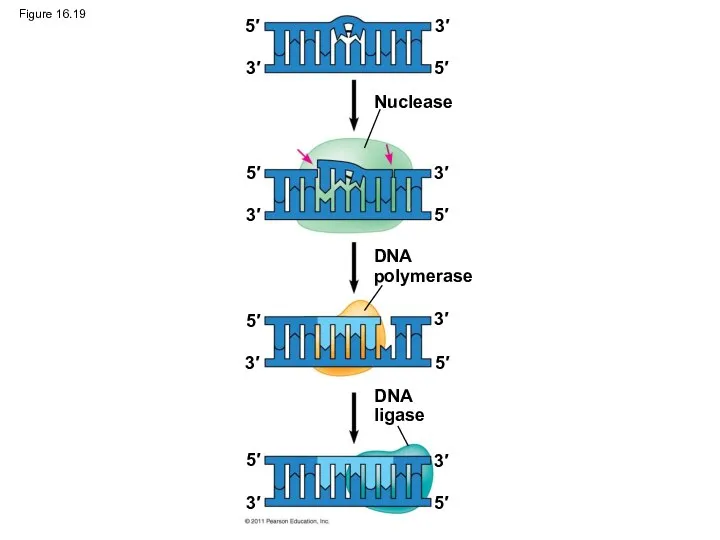
Figure 16.19
Nuclease
DNA
polymerase
DNA
ligase
5′
5′
5′
5′
5′
5′
5′
5′
3′
3′
3′
3′
3′
3′
3′
3′
Figure 16.19
Nuclease
DNA
polymerase
DNA
ligase
5′
5′
5′
5′
5′
5′
5′
5′
3′
3′
3′
3′
3′
3′
3′
3′

Evolutionary Significance of Altered DNA Nucleotides
Error rate after proofreading repair is
Evolutionary Significance of Altered DNA Nucleotides
Error rate after proofreading repair is

Replicating the Ends of DNA Molecules
Limitations of DNA polymerase create problems
Replicating the Ends of DNA Molecules
Limitations of DNA polymerase create problems
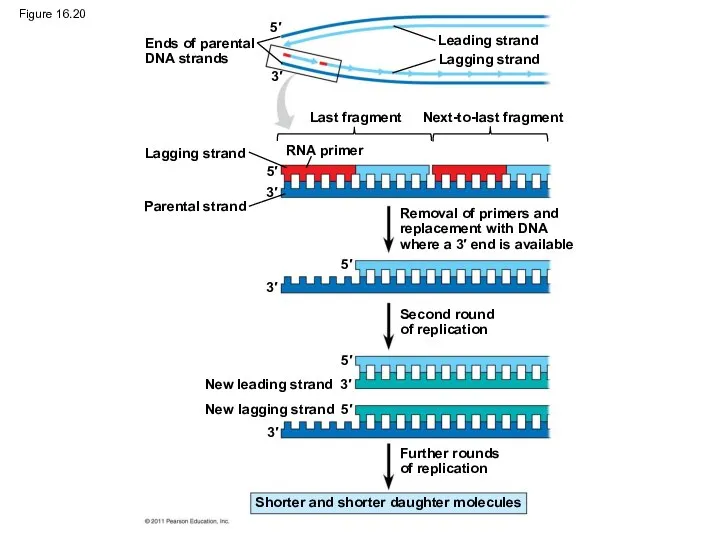
Figure 16.20
Ends of parental
DNA strands
Leading strand
Lagging strand
Last fragment
Next-to-last fragment
Lagging strand
RNA primer
Parental
Figure 16.20
Ends of parental
DNA strands
Leading strand
Lagging strand
Last fragment
Next-to-last fragment
Lagging strand
RNA primer
Parental
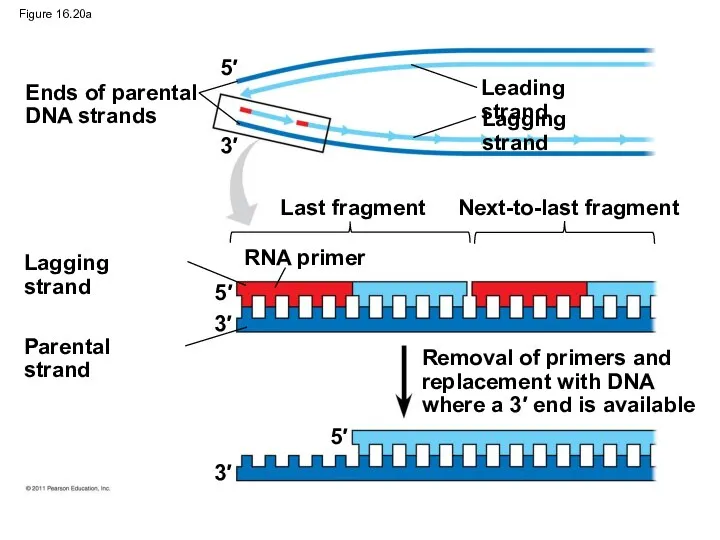
Figure 16.20a
Ends of parental
DNA strands
Leading strand
Lagging strand
Last fragment
Next-to-last fragment
Lagging strand
RNA primer
Parental
Figure 16.20a
Ends of parental
DNA strands
Leading strand
Lagging strand
Last fragment
Next-to-last fragment
Lagging strand
RNA primer
Parental
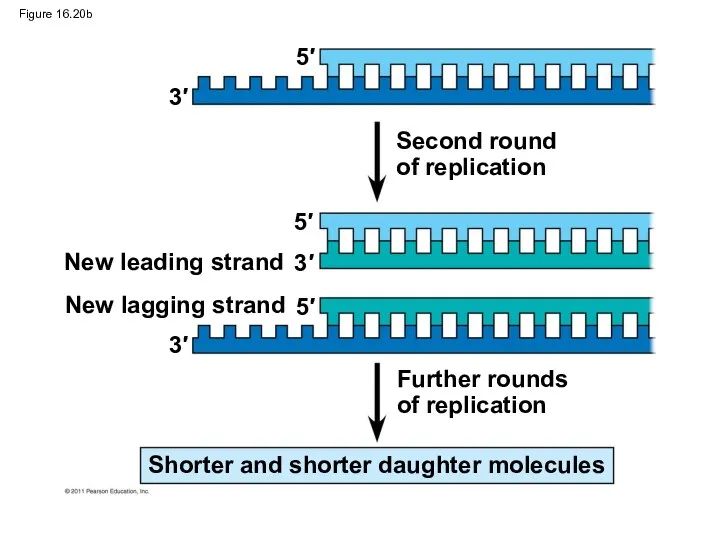
Figure 16.20b
Second round
of replication
Further rounds
of replication
New leading strand
New lagging strand
Shorter and
Figure 16.20b
Second round
of replication
Further rounds
of replication
New leading strand
New lagging strand
Shorter and

Eukaryotic chromosomal DNA molecules have special nucleotide sequences at their ends
Eukaryotic chromosomal DNA molecules have special nucleotide sequences at their ends

Figure 16.21
1 μm
Figure 16.21
1 μm

If chromosomes of germ cells became shorter in every cell cycle,
If chromosomes of germ cells became shorter in every cell cycle,

The shortening of telomeres might protect cells from cancerous growth by
The shortening of telomeres might protect cells from cancerous growth by

Concept 16.3 A chromosome consists of a DNA molecule packed together
Concept 16.3 A chromosome consists of a DNA molecule packed together

Chromatin, a complex of DNA and protein, is found in the
Chromatin, a complex of DNA and protein, is found in the
Animation: DNA Packing Right-click slide / select “Play”
Animation: DNA Packing Right-click slide / select “Play”
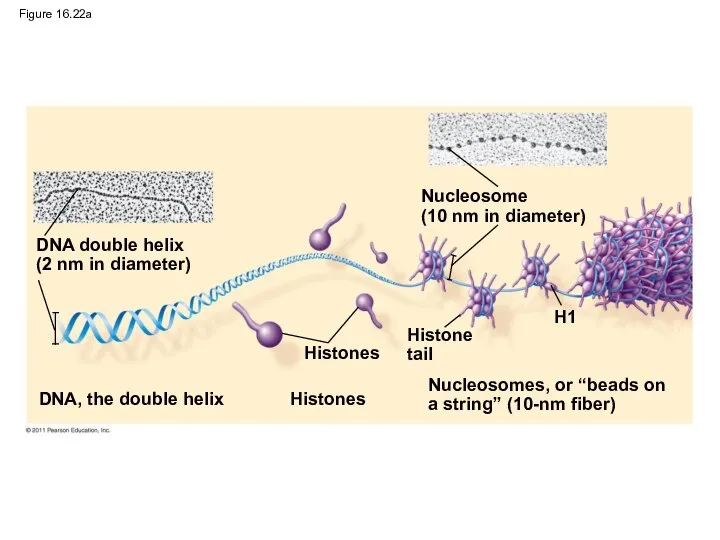
Figure 16.22a
DNA double helix
(2 nm in diameter)
DNA, the double helix
Nucleosome
(10 nm
Figure 16.22a
DNA double helix
(2 nm in diameter)
DNA, the double helix
Nucleosome (10 nm
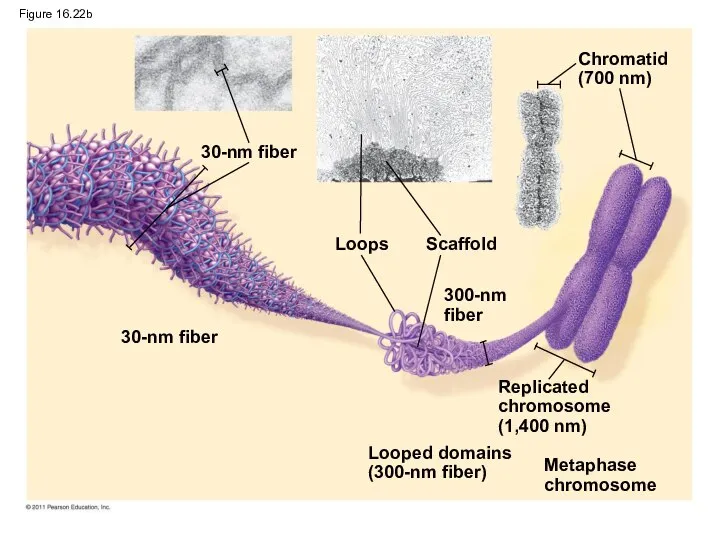
Figure 16.22b
30-nm fiber
30-nm fiber
Loops
Scaffold
300-nm fiber
Chromatid
(700 nm)
Replicated
chromosome
(1,400 nm)
Looped domains
(300-nm fiber)
Metaphase
chromosome
Figure 16.22b
30-nm fiber
30-nm fiber
Loops
Scaffold
300-nm fiber
Chromatid
(700 nm)
Replicated
chromosome
(1,400 nm)
Looped domains
(300-nm fiber)
Metaphase
chromosome
Figure 16.22c
DNA double helix (2 nm in diameter)
Figure 16.22c
DNA double helix (2 nm in diameter)

Figure 16.22d
Nucleosome (10 nm in diameter)
Figure 16.22d
Nucleosome (10 nm in diameter)

Figure 16.22e
30-nm fiber
Figure 16.22e
30-nm fiber
Figure 16.22f
Loops
Scaffold
Figure 16.22f
Loops
Scaffold
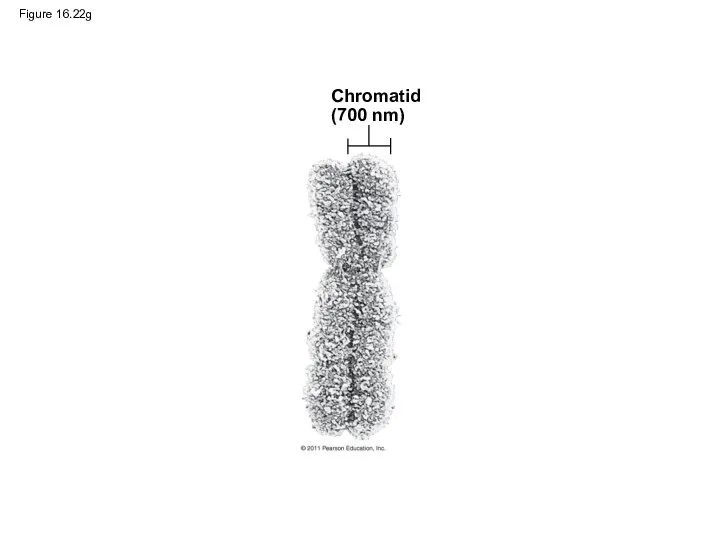
Figure 16.22g
Chromatid
(700 nm)
Figure 16.22g
Chromatid
(700 nm)

Chromatin undergoes changes in packing during the cell cycle
At interphase, some
Chromatin undergoes changes in packing during the cell cycle
At interphase, some
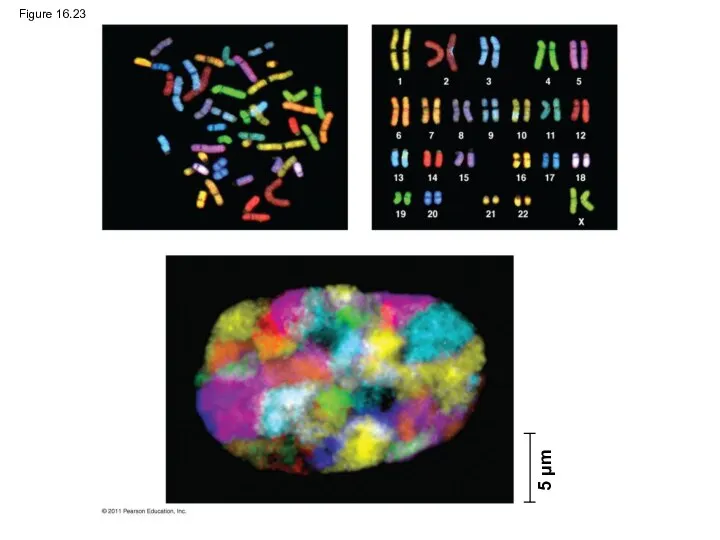
Figure 16.23
5 μm
Figure 16.23
5 μm

Figure 16.23a
Figure 16.23a

Figure 16.23b
Figure 16.23b
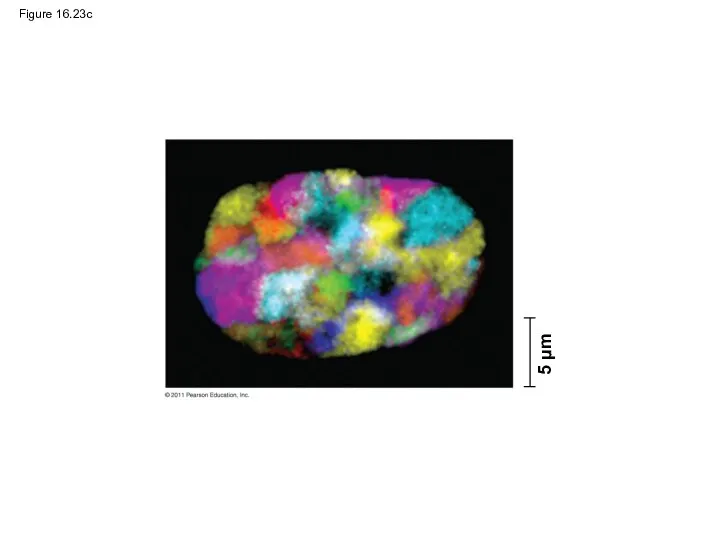
Figure 16.23c
5 μm
Figure 16.23c
5 μm

Most chromatin is loosely packed in the nucleus during interphase and
Most chromatin is loosely packed in the nucleus during interphase and

Histones can undergo chemical modifications that result in changes in chromatin
Histones can undergo chemical modifications that result in changes in chromatin
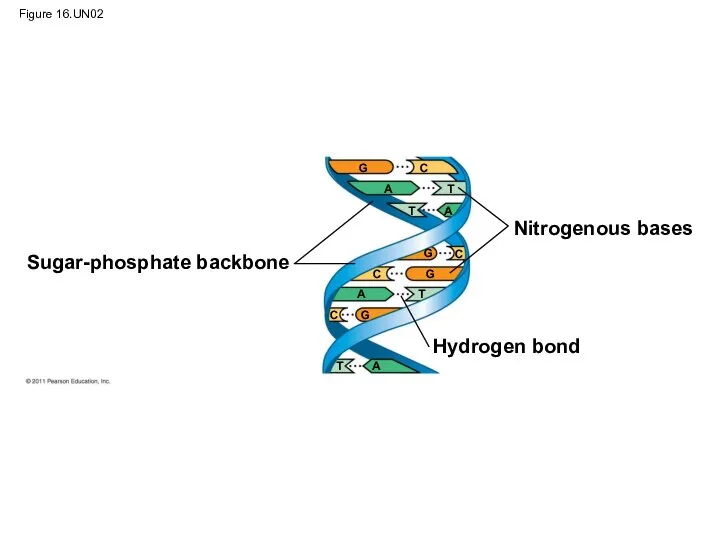
Figure 16.UN02
Sugar-phosphate backbone
Nitrogenous bases
Hydrogen bond
G
G
G
G
C
C
C
C
A
A
A
A
T
T
T
T
Figure 16.UN02
Sugar-phosphate backbone
Nitrogenous bases
Hydrogen bond
G
G
G
G
C
C
C
C
A
A
A
A
T
T
T
T
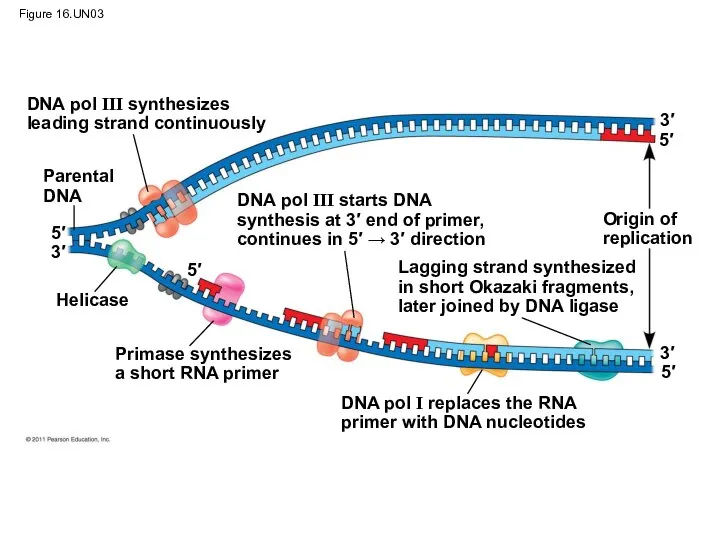
Figure 16.UN03
DNA pol III synthesizes
leading strand continuously
Parental
DNA
DNA pol III starts DNA
synthesis
Figure 16.UN03
DNA pol III synthesizes
leading strand continuously
Parental
DNA
DNA pol III starts DNA synthesis
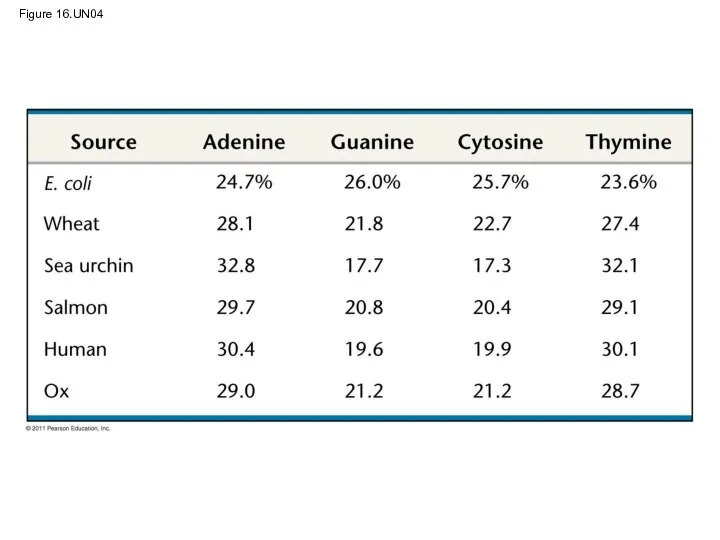
Figure 16.UN04
Figure 16.UN04
Figure 16.UN05
Figure 16.UN05
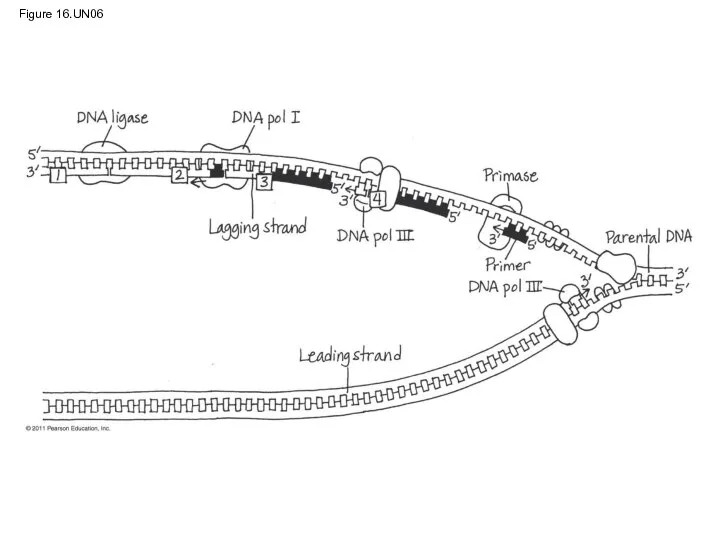
Figure 16.UN06
Figure 16.UN06
 Вода. Классы неорганических соединений. 8 класс
Вода. Классы неорганических соединений. 8 класс Вакуумное фильтрование
Вакуумное фильтрование Битумы природного происхождения
Битумы природного происхождения Растворение. Растворы
Растворение. Растворы Вещества
Вещества Пропан, C3H8
Пропан, C3H8 Презентация по Химии "Соединения" - скачать смотреть бесплатно
Презентация по Химии "Соединения" - скачать смотреть бесплатно Минералы и Близнецы
Минералы и Близнецы Озоновый слой. Механизмы образования и разрушения
Озоновый слой. Механизмы образования и разрушения Химически опасные объекты (ХОО)
Химически опасные объекты (ХОО) Химические реакции (11 класс)
Химические реакции (11 класс) Диазо- и азосоединения
Диазо- и азосоединения Структура кристаллических полимеров
Структура кристаллических полимеров Положение металлов в Периодической системе Д.И. Менделеева. Особенности строения атомов, свойства
Положение металлов в Периодической системе Д.И. Менделеева. Особенности строения атомов, свойства Железо. Физические и химические свойства
Железо. Физические и химические свойства Презентация по Химии "ЖИРНЫЕ КИСЛОТЫ СОСТАВНЫЕ ЧАСТИ ЛИПИДОВ" - скачать смотреть бесплатно
Презентация по Химии "ЖИРНЫЕ КИСЛОТЫ СОСТАВНЫЕ ЧАСТИ ЛИПИДОВ" - скачать смотреть бесплатно Работу выполнила: Ученица 9 класса ГОУ Лицея № 1524 Г.Москвы Себко Екатерина Научный руководитель: Учитель химии ГОУ Лицея № 1524 К
Работу выполнила: Ученица 9 класса ГОУ Лицея № 1524 Г.Москвы Себко Екатерина Научный руководитель: Учитель химии ГОУ Лицея № 1524 К Методы светорассеяния для исследования растворов (био) полимеров и наночастиц
Методы светорассеяния для исследования растворов (био) полимеров и наночастиц Презентация по Химии "Семь доисторических металлов" - скачать смотреть
Презентация по Химии "Семь доисторических металлов" - скачать смотреть  Окислительно-восстановительные реакции
Окислительно-восстановительные реакции Shapes of molecules
Shapes of molecules Над проектом работали ученики 6 класса: Над проектом работали ученики 6 класса: Крючков Слава Орлов Слава Старова Катя Пугачев Се
Над проектом работали ученики 6 класса: Над проектом работали ученики 6 класса: Крючков Слава Орлов Слава Старова Катя Пугачев Се Статья двумя способами. Приготовление катализатора
Статья двумя способами. Приготовление катализатора Кислотно-основные взаимодействия. Принцип ЖМКО Пирсона
Кислотно-основные взаимодействия. Принцип ЖМКО Пирсона Оксиды азота
Оксиды азота Тест по теме «Альдегиды и кетоны»
Тест по теме «Альдегиды и кетоны» Тұздар. Құрамы және химиялық қасиеттері
Тұздар. Құрамы және химиялық қасиеттері Химическая очистка воды
Химическая очистка воды