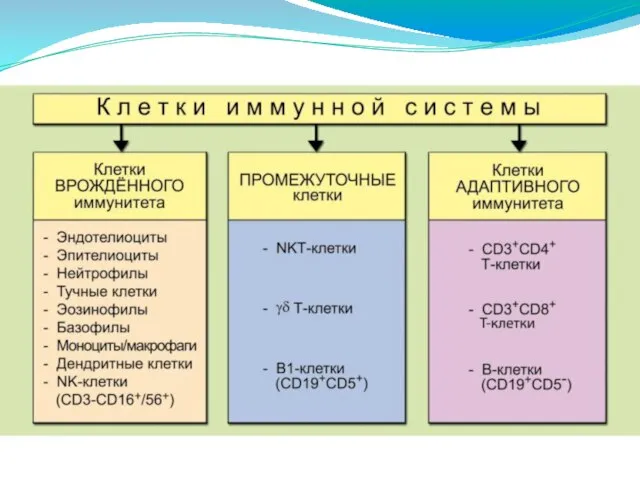
- Основы иммунологии

Содержание
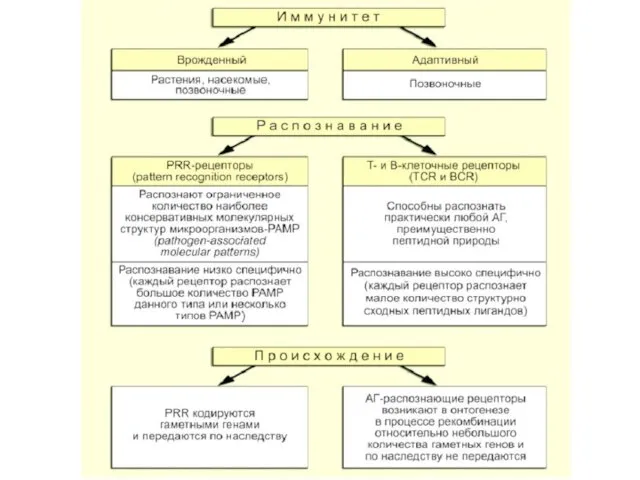
- 2. Иммунитет– это биологическое свойство многоклеточных организмов, обеспечивающее постоянство внутренней среды, по средствам: -распознавания -ослабления /деструкции -эллиминации
- 3. Врожденный Приобретенный Иммунитет Активный Пассивный Пассивный Активный Искусственный Естественный
- 5. Характеристика врожденного и приобретенного иммунитета
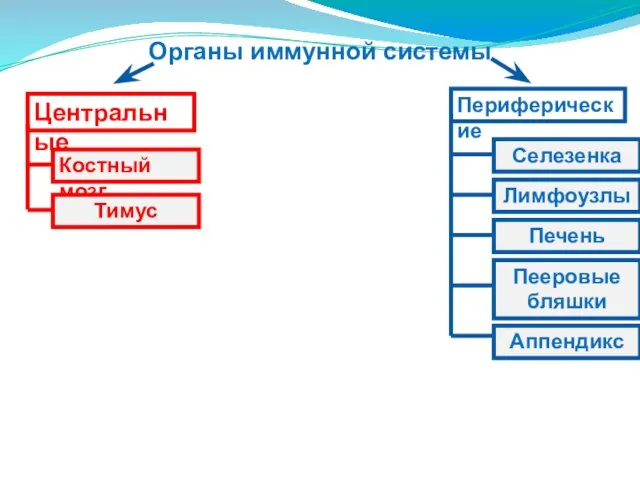
- 6. Органы иммунной системы Центральные Периферические Костный мозг Тимус Селезенка Аппендикс Пееровые бляшки Печень Лимфоузлы
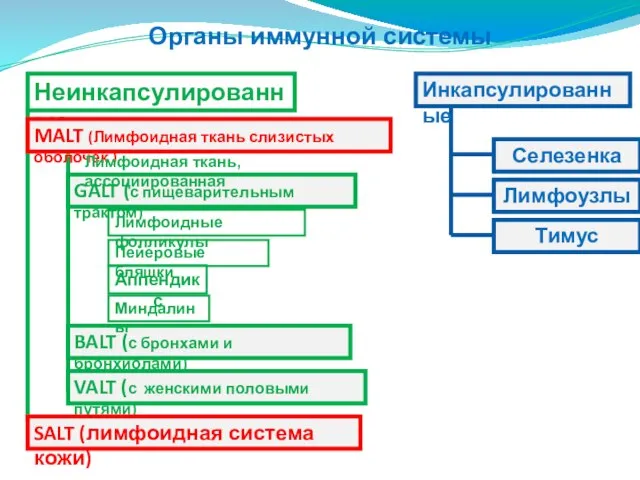
- 7. Органы иммунной системы Неинкапсулированные MALT (Лимфоидная ткань слизистых оболочек ) GALT (с пищеварительным трактом) Селезенка Тимус
- 9. Неспецифическая резистентность организма - устойчивость организма к любым патогенным воздействиям, независимо от их природы. 1.Механические барьеры
- 10. Эффекторные механизмы защиты организма 1.Фагоцитоз -Нейтрофиллы -Моноциты -Макрофаги 2.Экзоцитоз -эозинофилы -базофилы -мастоциты 3.Цитолиз -Комплемент -NK-клетки -ЦТЛ
- 11. Функции врожденного иммунитета
- 12. Основные факторы врожденного иммунитета Клеточные факторы
- 13. Основные защитные механизмы врожденного иммунитета
- 14. Фазы ответной реакции врожденного иммунитета
- 15. КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
- 16. КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА Нейтрофилы Тучные клетки Эозинофилы Базофилы Моноциты Дендритные клетки NK-клетки Эндотелиоциты Эпителиоциты Клетки
- 17. Развитие клеток крови
- 18. Гранулоциты
- 19. Фагоцитирующие клетки Моноциты Макрофаги Нейтрофилы
- 20. 1.Развитие нейтрофилов 2.Морфология клетки 3. Гранулы нейтрофилов и их свойства 4. Кластерные молекулы и их функции
- 21. Развитие нейтрофиллов Стволовая клетка крови Клетка предшественник миелопоэза Миелобласт Промиелоцит Миелоцит (Юный нейтрофил) Палочкоядерный нейтрофил Сегментоядерный
- 22. Развитие нейтрофилов В митотическую фазу (длится 7,5 дней) стволовая клетка дифференцируется в миелобласт. Далее следует стадия
- 23. Миелопероксидазу Эластазу Дефензины Азуроцидин Лизоцим Нейтральные протезы: катепсин G и протеазу 3 Промиелоцит содержит «первичные» азурофильные
- 24. Лактоферрин Лизоцим Фосфолипазу Ф2 Белок, повышающий проницаемость микробной стенки Белок NGAI Липокартин Кателицидин В12 связывающий протеин
- 25. В постмитотическую фазу (продолжительность около 6,5 дней) нейтрофил прекращает делиться, и начинается активная перестройка ядерного аппарата.
- 26. Основная часть морфологически зрелых нейтрофилов остаются в костном мозге, а меньшее их количество (около 3%) поступает
- 27. Морфология Нейтрофильные сегментоядерные лейкоциты - преобладающая популяция белых клеток крови. Клетки диаметром 9-12мкм, имеют сегментированное ядро
- 28. Гранулы нейтрофилов При дегрануляции азурофильные гранулы первыми сливаются с фагосомами, затем - специфические. Мембрана специфических и
- 29. Гранулы нейтрофилов Содержащиеся в цитоплазме нейтрофила гранулы принято делить на: 1-азурофильные или первичные гранулы; 2- специфические
- 30. Свойства и состав гранул нейтрофилов
- 31. Все указанные вещества обладают антимикробным действием Миелопероксидаза (MPO) составляет 2–4% массы нейтрофила, катализирует образование хлорноватистой кислоты
- 32. Катепсин G при нейтральном рН убивает грамположительные и грамотрицательные бактерии. Дефицит катепсина G и связанное с
- 33. СD14 – связывание комплекса LPS+LBP, фагоцитоз Гр- бактерий. CD15 – опосредует фагоцитоз и хемотаксис нейтрофилов. CD11c
- 34. Кластерные молекулы нейтрофилов
- 35. Кластерные молекулы нейтрофилов
- 36. Основные процессы, происходящие в нейтрофилах при их активации и фагоцитозе
- 37. На рисунке отражены участие НФ в фагоцитозе (миграция, поглощение, дегрануляция, внутриклеточный киллинг, деградация, экзоцитоз и апоптоз)
- 38. Участие нейтрофилов в асептическом воспалении
- 39. 1.Развитие моноцитов 2.Морфология моноцитов 3. Моноциты периферической крови 4. Развитие макрофагов 5. Морфология макрофагов 6.Кластерные молекулы
- 40. Моноцит Монобласт Моноцитарно-гранулоцитарный предшественник Плюрипотентная СК Моноцитарный предшественник Промоноцит Миелоидный полипотентный предшественник IL-1,3,6 IL-1,3 GM-CSF, M-CSF,
- 41. тонкая структура хроматина Моноциты представляют довольно крупные клетки диаметром 9–15 мкм ядром бобовидной формы Моноциты. Морфология
- 42. Моноциты периферической крови воспалительные (CD14++CD16-) резидентные (CD14+CD16+) В воспалительном очаге созревают в МФ. В регионарном лимфатическом
- 43. Основные рецепторы моноцитов
- 44. Гетерогенность клеток, берущих начало от моноцитов
- 45. Развитие макрофагов МФ, так же как ДК и остеокласты (ОК), происходят из стволовой клетки костного мозга
- 46. изменяется экспрессии некоторых мембранных молекул (ослабевает экспрессия CD13, CD14, CD15, β1-интегринов, FcγRI, усиливается экспрессия CD16). Это
- 47. Макрофаги значительно крупнее моноцитов (диаметр составляет 20–25 мкм). Имеют распластанную форму. В отличие от округлых моноцитов,
- 48. Кластерные молекулы макрофагов
- 49. Основные группы рецепторов макрофагов, распознающих чужеродные вещества
- 50. Моноциты периферической крови воспалительные (CD14++CD16-) резидентные (CD14+CD16+) В воспалительном очаге созревают в МФ. В регионарном лимфатическом
- 51. Моноциты воспалительные (CD14++CD16-) В первые часы острого воспаления моноциты для проникновения используют те же интегрины, что
- 52. Функции макрофагов Основная функция МФ - распознавать чужеродные агенты и осуществлять фагоцитоз
- 53. Активированный макрофаг
- 54. Различные типы активации макрофагов
- 55. Различные типы активации макрофагов В классическом виде процесс запускается ΙΚΝγ и TNF, индуцируемыми микроорганизмами и РАМР,
- 56. Активация макрофагов Во второй половине 90X гг. XX века в лаборатории Дэвида Моссера (Университет Мэриленда; США)
- 57. Начальные этапы фагоцитоза бактерий моноцитами/макрофагами
- 58. Фагоцитоз Явление фагоцитоза было открыто И.И. Мечниковым (1882). Он показал фундаментальную роль фагоцитоза как способа питания
- 59. Стадии фагоцитоза 1. Хемотаксис 2. Адгезия 3. Активация мембраны 4. Погружение; 5.Образование фагосомы; 6.Слияние фагосомы и
- 60. Стадии фагоцитоза
- 61. Поверхностные молекулы клеток, участвующие в фагоцитозе В миграции фагоцитов в очаг воспаления принимают участие три группы
- 62. Поверхностные молекулы клеток, участвующие в фагоцитозе Селектины (CD62Р, CD62E) - экспрессируются в небольших количествах на клетках
- 63. Поверхностные молекулы клеток, участвующие в фагоцитозе Интегрины являются гетеродимерами и включают четыре вида поверхностных молекул лейкоцитов.
- 64. Поверхностные молекулы клеток, участвующие в фагоцитозе Суперсемья иммуноглобулинов включает молекулы ICAM, которые экспрессируются на клетках эндотелия,
- 65. Движение нейтрофила. Хемотаксис. Хемотаксис - направленное движение клеток, определяемое градиентом химических факторов (хемоаттрактантов).При реализации врожденного иммунитета
- 66. Cтадии хемотаксиса 1. Стадия качения (3) 2. Стадия активации 3.Стадия прочной адгезии (4,5) 4.Стадия экстравазации(7) 1
- 67. Адгезия Обязательным условием адгезии фагоцита служит распознавание объекта фагоцитоза. Механизмы распознавания разнообразны и принципиально различаются в
- 68. Scavenger-рецепторы (рецепторы-мусорщики). Эти рецепторы экспрессированы на макрофагах и некоторых дендритных клетках. Лигандами для scavenger-рецепторов служат компоненты
- 69. Погружение Существуют различия в феноменологии процесса погружения частицы в зависимости от того, какие рецепторы участвуют в
- 70. Образование фаголизосомы Образование фаголизосомы Эффективность фагоцитоза обусловлена не столько поглощением патогена, сколько его разрушением внутри клетки.
- 71. Эозинофилы 1.Развитие эозинофилов 2. Морфология клетки 3. Гранулы эозинофилов и их свойства 4. Кластерные молекулы эозинофилов
- 72. ЭО (2) происходят из CD34+ предшественников костного мозга (1), где они созревают примерно в течение 5
- 73. МОРФОЛОГИЯ Эозинофилы составляют 1–5% лейкоцитов, циркулирующих в крови. Эозинофилы в течение нескольких дней после образования остаются
- 74. Гранулы эозинофилов
- 75. Гранулы эозинофилов
- 76. Белки MBP, ECP, EDN - токсичные для паразитов и клеток млекопитающих. Белки ECP, EDN - обладают
- 77. Кластерные молекулы эозинофилов CD9 и CD35 (рецептор для комплемента— CR1)- что позволяет отличить ихотнейтрофилов с помощью
- 78. 1. Повышение чувствительности рецепторов к иммуноглобулинам класса Е. За счет этого активизируется противопаразитарный иммунитет, и происходит
- 79. Функции Уничтожение паразитов. Эозинофилия возникает при многих паразитарных болезнях. Эозинофилы особенно активно уничтожают паразитов в местах
- 80. Побочные эффекты Секретируемые эозинофилом вещества могут повреждать нормальные ткани. Так, при постоянном высоком содержании эозинофилов в
- 81. Эозинофилы (ЭО) могут находиться в двух состояниях: неактивированном (1)и активированном (2). Главную роль в активации играют
- 82. Неактивированные эозинофилы Неактивированные (1) характеризуются наличием низкоаффинного рецептора FceRII (CD23), интегрина LFA-1, хемокинового рецептора CCR3 и
- 83. Направленная миграция обусловлена экспрессией на зрелых эозинофилах специфического для эо-таксина рецептора CCR3 (4), построенного аналогично рецептору
- 84. Эотаксины играют главную роль в направленной миграции ЭО в лёгкие (5), желудочно-кишечный (6) и урогенитальный (7)
- 85. Заболевания, связанные с эозинофилами
- 86. На верхней половине графика (1) представлены типы эозинофилии, на нижней (2) - заболевания, при которых наблюдается
- 87. Считается, что ЭО играют защитную роль при глистных инвазиях. В экспериментальных моделях in vivo можно наблюдать
- 88. ЭО участвуют в патогенезе бронхиальной астмы, при которой наблюдаются средний уровень эозинофилии и инфильтрация лёгких эозинофилами.
- 89. Особняком от этих заболеваний стоит идиопатический гиперэозинофильный синдром (HES), который характеризуется наличием в течение 6 мес
- 90. К гиперэозинофильным синдромам относится эозинофильный фибропластический эндокардит, характеризующийся стойкой эозинофилией в течение не менее 6 мес
- 91. 1.Развитие базофилов и тучных клеток 2. Гранулы базофилов и тучных клеток, их свойства Базофилы. Тучные клетки
- 92. Тучные клетки (мастоциты) и базофилы представляют тканевые клетки, содержащие в цитоплазме базофильные гранулы. Тучная клетка Базофил
- 93. Схема развития и миграции тучных клеток и базофилов. Рядом с кружками, обозначающими клетки, указаны их маркеры.
- 94. Оба типа клеток имеют костномозговое происхождение и принадлежат к миелоидному ряду. Предполагают, что у тучных клеток
- 95. CD13 - рецептор для распознавания ряда вирусов. CD33 - антиген (гликопротеин) миелоидной дифференцировки. CD34 - поверхностный
- 96. Гранулы базофилов и тучных клеток
- 97. Базофилы в норме представлены в кровяном русле. Их содержание в крови очень невелико — до 0,5%
- 98. CD11 - группа поверхностных рецепторов (ITGA, интегрины a), осуществляющих адгезию лейкоцитов между собой и к межклеточному
- 99. Спектр активных веществ, секретируемых базофилами, ограничен: лейкотриен C4, IL-4, IL-13, тромбоксаны простогландины, фактор хемотаксиса эозинофилов и
- 100. Активация базофилов IgE-зависимый путь активации базофилов IgE-независимый путь активации базофилов
- 101. IgE-зависимый путь активации базофилов Базофилы экспрессируют высокоаффинный рецептор FcsRI (1), который без участия АГ связывается с
- 102. IgE-независимый путь активации базофила Первый этап - индукция образования IL-4 фактором созревания базофилов цитокином IL-3. При
- 103. 1. На ранних этапах развития иммунного ответа базофилы являются главными индукторами образования Тh2-клеток. 2. Активированные базофилы
- 104. -регуляция процессов свертывания крови и проницаемости сосудов -базофилы, как и эозинофилы, участвуют в иммунных воспалительных реакциях
- 105. 1.Развитие базофилов и тучных клеток 2. Гранулы базофилов и тучных клеток, их свойства Базофилы. Тучные клетки
- 106. Диаметр тучных клеток варьирует от 10 до 20 мкм. Они имеют овальную форму с ворсинчатой поверхностью.
- 107. Характеристика тучных клеток 2-мембранносвязанный цитокин SCF , 3- c-kit -цитокин является лигандом для рецептора ТК, 4-FceRI
- 108. CD13 (GP150, поверхностный антиген лейкоцитов, аминопептидаза N-PEPN) - рецептор некоторых коронавирусов - возбудителей инфекционных заболеваний верхних
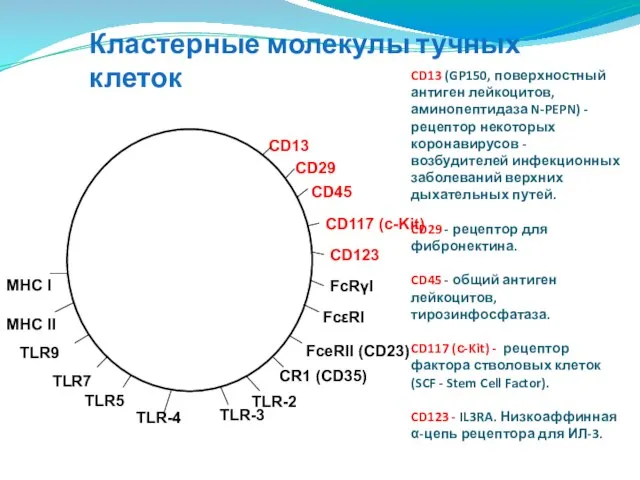
- 109. Кластерные молекулы тучных клеток FcRγI - CD64. FCGR1B. Высокоаффинный рецептор к IgG FcεRI - высокоаффинный рецептор
- 110. Кластерные молекулы тучных клеток Патогенраспознающие рецепторы - TLR - Toll-Like Receptors: TLR-2 (CD282) - опосредует ответ
- 111. Разновидности тучных клеток человека и их свойства
- 112. Разновидности тучных клеток человека и их свойства
- 113. 1 2 3 1 сигнальный путь – идет через Фосфолипазу С 2 сигнальный путь – идет
- 114. Дегрануляция тучных клеток Гистамин Химаза Триптаза Са 2+ Серотонин Гепарин Фактор хемотаксиса эозинофилов
- 115. Секреция эйкозаноидов Фосфолипиды клеточной стенки Фосфолипаза А2 Простогландины Лейкотриены Арахидоновая кислота ЦОГ ЛОГ Тромбоксаны Проницаемости и
- 116. 1) Участие в реакциях воспаления (гистамин, гепарин): повышение проницаемости сосудов, образование отёка; 2) Участие в аллергических
- 117. Функции тучных клеток
- 118. Дендритные клетки (ДК) Это специализированные антигенпрезентирующие лейкоциты, способные активировать как наивные Т-клетки, так и Т-клетки памяти
- 119. Места локализации Основные места локализации: барьерные органы, слизистые оболочки и регионарные лимфатические узлы.
- 120. Макрофаг Лимфойдный предшественник Моноциты Плазмацитоидный моноцит/лимфоцит Кровь Кровь Интерстициальная дентритная клетка Макрофаг Клетка Лангерганса Плазмацитойдная ДК
- 121. Миелоидные ДК и макрофаги Миелоидные ДК и макрофаги имеют общего костномозгового предшественника, несущего маркер CD34 (молекула
- 122. Плазмацитоидные ДК Плазмацитоидные ДК также происходят из CD34 позитивного предшественника. Внешне походи на плазматические клетки. У
- 123. Циркулирующие в крови предшественники Зрелые Дендритные клетки
- 125. Отличия зрелой от незрелой дендритной клетки
- 126. Активаторы ДК
- 127. Миграция ДК В зависимости от степени созревания, ДК клетки будут проявлять различную фагоцитарную активность. Незрелые ДК
- 128. Мембранные молекулы незрелых ДК
- 129. Мембранные молекулы зрелых ДК
- 130. Заболевания, связанные с нарушением дифференцировки и функционирования дендритных клеток
- 131. NK-клетки 1.Развитие NK-клеток 2.Морфология NK-клеток 3.Гранулы NK-клеток 4.Кластерные молекулы NK-клеток 5.Функции NK-клеток 6. Эффекторные функции NK-клеток.
- 132. Развитие NK-клеток
- 133. NK-клетки морфологически сходны с лимфоцитами. Однако крупнее их и содержат множество цитоплазматических вакуолей, содержат в цитоплазме
- 134. Гранулы Перфорин представляет собой белок с высокой молекулярной массой. На поверхности клетки мишени образуется гомополимер, повышающий
- 135. Гранулы Гранулизин - апсониноподобный положительно заряженный белок с молекулярной массой 9 kDa. Положительно заряженный гранулизин внедряется
- 136. Гранулы Гранзимы - протеазы химотрипсинового типа основной эффект гранзимов В активация протеазы – каспазы, нарушение целостности
- 137. NK CD16 CD8 CD2 CD11a/CD18 и CD11b/CD18 СD31 VLA4 VLA5 СD56 СD56 - обеспечивает взаимодействие NK-клеток
- 138. NK CD8 CD2 CD11a/CD18 и CD11b/CD18 СD31 VLA4 VLA5 СD56 CD16 CD16 - рецептор для FC-фрагмента
- 139. NK CD16 CD2 CD11a/CD18 и CD11b/CD18 СD31 VLA4 VLA5 СD56 CD8 CD8 отвечают за цитотоксическую активность.
- 140. NK CD16 CD2 СD31 VLA4 VLA5 СD56 CD8 CD11a/CD18 и CD11b/CD18 CD11a/CD18 и CD11b/CD18 - присоединение
- 141. NK CD16 CD11a/CD18 и CD11b/CD18 СD31 VLA4 VLA5 СD56 CD8 CD2 CD2 - молекулярная адгезия к
- 142. NK CD16 CD2 CD11a/CD18 и CD11b/CD18 VLA4 VLA5 СD56 CD8 СD31 СD31-молекула гомофильного взаимодействия, отвечает за
- 143. NK CD16 CD2 CD11a/CD18 и CD11b/CD18 СD31 VLA5 СD56 CD8 VLA4 VLA4-присоединение к молекулам эндотелия VCAMI.
- 144. NK Активаторы Ингибиторы NKP (46, 44, 30) KIR (p50,1; p50,2; p50,3) NKG2D NKG2C, E KIR(p58,1; p58,2;
- 145. Регуляция процессов пролиферации, дифференцировки и элиминации стареющих соматических клеток организма Модуляция клеток врожденного иммунитета Супрессия или
- 146. Созревание предшественников CTL Генерация вирус-специфических CTL Созревание претимоцитов и тимоцитов Функции NK-клеток
- 147. Взаимодействие NK-клеток с клетками мишеньями
- 148. NK KIR p5a1 кл HLA-CW2 Активация NK кл NK Выход гранул в пресинаптическую щель кл поры
- 149. Основной механизм лизиса мишеней
- 150. NK кл FasL(CD178) Fas (CD95) NK кл FADD Сигнал передается на каспазы Запуск апоптоза Рецепторное включение
- 151. Опухолевые клетки (5). В сыворотке крови появляются в высоких титрах растворимые формы белков MIC. Это может
- 152. Растворимые формы М1С-белков (2) реагируют с NKG2D ΝΚ-клетки, в результате чего ΝΚ-клет-ка не распознаёт опухолевую клетку
- 153. Постоянный контакт ΝΚ-клетки с растворимыми формами MIC-белков приводит к анергии ΝΚ-клеток, проявляющейся в утрате или пониженной
- 154. Опухолевая клетка (5) синтезирует цитокин TGF-β, который подавляет экспрессию NKG2D у ΝΚ-клеток, вследствие чего она не
- 155. Участие естественных киллеров в вирусных заболеваниях
- 156. Отрицательный результат заключается в том, что поражённые клетки-мишени, не экспрессирующие или слабо экспрессирующие молекулы МНС I
- 157. Представлен пример лизиса клетки, поражённой ΝΚ-клетками. Литическая реакция стала возможной благодаря снижению экспрессии классических молекул МНС
- 158. NK-клетки выступают в роли в роли LGL клеток. Их функции: Регулируют инвазивный рост трофобласта; В трофобласт
- 159. ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
- 160. ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
- 161. ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА Гуморальная составляющая врожденного иммунитета представлена несколькими взаимосвязанными системами – системой комплемента цитокиновой
- 162. Компоненты системы комплемента
- 163. Система комплемента является важным компонентом врождённого иммунитета, играющим большую роль в защите и удалении из организма
- 164. Два пути активации комплемента Классический путь Альтернативный путь
- 165. Классический путь Альтернативный путь
- 166. Классический путь активации комплемента
- 167. А) После присоединения AT класса IgM или некоторых изотипов IgG к поверхности бактериальной клетки у молекулы
- 168. Альтернативный путь активации комплемента
- 169. Альтернативный путь происходит в отсутствие AT на поверхности микробной клетки, приводя к образованию СЗ-конвертазы. Для его
- 170. В сыворотке крови присутствует сложный белок, относящийся к семейству коллектинов, - маннозасвязывающий лектин (MBL - от
- 171. Белки острой фазы воспаления Белки (реактанты) острой фазы представляют группу протеинов, секретируемых гепатоцитами. При воспалении продукция
- 173. Пентраксины Наиболее полно проявляют свойства реактантов острой фазы белки семейства пентраксинов: в первые 2—3 сут развития
- 174. Пентраксины Короткие петраксины Длинные пентраксины С-реактивный белок Сывороточный амилоид Р РТХ3 (пентраксин 3) Белок PTX3 вырабатывают
- 175. Происхождение и функции пентраксинов
- 176. Распознавание и связывание РАМР (растворимые патогенраспознающие рецепторы ) Комплемент-активирующая(через Clq) и опсонизирующая функция пентраксинов делает их
- 177. Пентраксины. С-реактивный белок Строение Пять доменов, объединены нековалентными связями в кольцевую структуры и формируют молекулу С-реактивного
- 178. является предшественником Р-компонента амилоидных фибрилл - постоянной составной части амилоидных депозитов при всех формах амилоидоза в
- 179. Они играют роль переносчиков метаболитов, ионов металлов, физиологически активных факторов. К транспортным белкам относят: преальбумин альбумин
- 180. Протеазы и антипротеазы Другую группу образуют протеазы, активация которых необходима для формирования многих медиаторов воспаления, а
- 181. Цитокины Цитокины –это белковые или полипептидные факторы, лишенные специфичности в отношении антигенов, продуцируемые преимущественно активированными клетками
- 182. Интерфероны Общее свойство интерферонов - наличие у них противовирусной активности. В то же время, подобно другим
- 183. Функции основных интерферонов
- 184. Приобретенный иммунитет
- 186. Лимфоциты.Морфология Лимфоцит - округлая клетка, с диаметром 7 – 9мкм. Ядерно-цитоплазматическое соотношение в пользу ядра. Ядро
- 187. Дифференцировка лимфоцитов
- 188. Органы иммунной системы Центральные Периферические Костный мозг Тимус Селезенка Аппендикс Пееровые бляшки Печень Лимфоузлы
- 189. Органы иммунной системы Неинкапсулированные MALT (Лимфоидная ткань слизистых оболочек ) GALT (с пищеварительным трактом) Селезенка Тимус
- 190. Тимус (вилочковая железа) Тимус расположен за рукояткой грудины. У человека и большинства млекопитающих он состоит из
- 191. Тимус Тимус состоит из двух основных частей - коры и мозгового вещества. В коре выделяют наружный,
- 192. Тимус
- 193. 1 - корковое вещество; 2 - мозговое вещество; 3 - капсула; 4 - соединительнотканная перегородка (септа);
- 194. Клетки тимуса макрофаг В-лимфоцит Тучная клетка Дендритная клетка Фибробласт
- 195. Формирование и развитие эпителия тимуса
- 196. Возрастная инволюция тимуса Важная особенность тимуса - его возрастная инволюция. Максимальный относительный вес тимуса выявляют в
- 197. Возрастная инволюция тимуса Значительно уменьшается численность тимоцитов и секреторная активность эпителиальных клеток тимуса. Их способность размножаться
- 198. Лимфатические узлы
- 199. Лимфатические узлы - бобовидные образования величиной 0,2-1,0 см, образующие группы или цепочки. Они дренируют лимфу от
- 200. Лимфатические узлы
- 201. Селезенка служит иммунным барьером на путях гематогенного распространения патогенов и других чужеродных агентов. Ее функции более
- 202. Селезенка
- 203. Лимфоидная ткань слизистых оболочек MALT GALT (Лимфоидная ткань, ассоциированная с пищ.трактом)
- 204. Т-лимфоциты
- 205. Селекционная модель дифференцировки Селекционная модель дифференцировки Τ-клеток на субпопуляции CD4+- и CD8+- клеток предполагает спонтанную или
- 206. Дифференцировка Т-лимфоцитов Красный костный мозг ТИМУС 1 ЭТАП ПОЗИТИВНАЯ СЕЛЕКЦИЯ 2 ЭТАП НЕГАТИВНАЯ СЕЛЕКЦИЯ Есть связь
- 207. Дифференцировка Т-лимфоцитов Субкапсулярная зона Корковая часть Кортико- медулярная зона Мозговая часть Позитивная селекция Базальная мембрана Эндотелий
- 208. Полипотентность ранних тимических предшественников и роль дифференци-ровочных факторов в выборе пути развития их потомков
- 209. Кластерные молекулы Т-лмфоцитов Популяции Т- и В-клеток имеют клональную структуру: в процессе дифференцировки каждая клетка приобретает
- 210. Естественные субпопуляции Т-лимфоцитов и их дифференцировочные факторы
- 211. Типы эффекторных Т-клеток и их действие Действие всех 4 типов Т-клеток основано преимущественно на комбинации контактных
- 212. СD 4+ клетки
- 213. Жизнеспособность Т-клеток поддерживается благодаря получению 2 сигналов: при взаимодействии IL-7 с рецептором (IL7-R) на поверхности Т-клетки;
- 214. Субпопуляции CD4+ Т-лимфоцитов, их рецепторы и функции Тh1 несут рецепторы CD4 (как и Т- хелперы2) и
- 215. Тh2 имеют общий для хелперов CD4 и специфический CD28 рецепторы. обеспечивают пролиферацию и дифференцировку В- лимфоцитов
- 216. Маркерные цитокины и рецепторы Th1- и Th2-клеток
- 217. Взаимное ингибирование, опосредованное цитокинами
- 218. T-хелперы 17 – участвуют в реализации имунной защиты и в формировании иммунопатологий
- 219. Регуляторные Т-клетки (Treg) – их назначение состоит в предотвращении реакции на собственные АГ (естественные регуляторные Т-клетки)
- 220. Маркеры естественных регуляторных Т-клеток Действуют, в основном, через клеточные контакты с привлечением ДК.
- 221. Разновидности естественных и адаптивных регуляторных Т-клеток FOXP3 – внутриклеточный фактор (ген семейства FOX; необходим для реализации
- 222. Адаптивные субпопуляции CD4+ Т-клеток
- 223. Физиологические и патологические эффекты адаптивных субпопуляций CD4+Т-клеток Мф – макрофаг; Эо – эозинофил; В – В-лимфоцит;
- 224. Взаимоотношения адаптивных субпопуляций Т-хелперов
- 225. Связь форм иммунной защиты с локализацией патогена
- 226. Дифференцировка Т-хелперов Направление дифференцировки определяется характером антигенного стимула. Селекция осуществляется с помощью цитокинов дендритных клеток. Главное
- 227. Факторы, определяющие преобладание иммунного ответа Th1- или Th2-типа
- 229. Параллельное развитие эффекторных Т-клеток и Т-клеток памяти (вторичный иммунный ответ) В обоих случаях исходным событием является
- 230. СD 8+ клетки
- 231. Формирование ЦТЛ Наивные CD8+ Т-клетки являются предшественниками Tc-лимфоцитов. Они не проявляют цитотоксической активности, не экспрессируют IL-2R
- 232. Цитолиз клеток-мишеней Цитотоксическое действие Т-лимфоцитов на клетки-мишени (иммунный цитолиз) осуществляется в шесть этапов и принципиально подобно
- 233. Гранулы ЦТЛ и апоптоз клеток-мишеней В гранулах содержатся белки, участвующие в реализации цитолиза — перфорин, гранзимы,
- 234. При взаимодействии цитотоксического Т-лимфоцита (вверху) и клетки-мишени (внизу) образуется микрополость, в которую поступают молекулы перфорина и
- 235. Компонент комплемента С9 и перфорин являются гомологами и формируют поры на основе сходных физико-химических процессов. Пора,
- 236. Рецепторный апоптоз Цитолитическое действие СD8+Т-лимфоцитов, а также CD4--клеток на клетки-мишени наряду с перфориновым механизмом, включает механизм
- 237. Направленность действия Действие Tc-лимфоцитов направлено, главным образом, против вирус-инфицированных и опухолевых клеток. Пролиферация антигенспецифичных клонов является
- 238. Цитотоксический иммунный ответ Цитотоксический иммунный ответ складывается из четырёх этапов: Презентация дендритными клетками АГ CD8+ Т-клеткам,
- 239. Как на этапе индукции цитотоксических Т-лимфоцитов (CTL), так и при реализации их цитотоксического действия CD8+ Т-лимфоциты
- 240. Природа сигналов, генерируемых при взаимодействии В-клеток и Т-хелперов и необходимых для запуска активации и пролиферации В-клеток
- 241. Дифференцировка антителообразующих клеток Осуществляется преимущественно под влиянием цитокинов, секретируемых Th2-клетками
- 242. Параллельное развитие плазматических клеток-антителопродуцентов и В-клеток памяти (вторичный иммунный ответ)
- 243. Мукозальный иммунитет Слизистые оболочки располагаются в барьерных тканях с высокой антигенной нагрузкой. В слизистых имеются лимфоидные
- 244. Условия формирования толерантности и иммунитета в слизистых оболочках
- 246. В-лимфоциты представляют собой клетки адаптивного иммунитета – предшественники плазматических клеток, продуцирующих в организме антитела. Это неоднородная
- 247. Лимфоциты - вторые по количественному содержанию лейкоциты (20-35%). Бурсазависимые лимфоциты (В-лимфоциты) - впервые обнаружены в сумке
- 248. Рис. 1. В-лимфоциты: а - В-лимфоциты мыши с меченными 125I антителами (стрелки) против иммуноглобулинов кролика (по
- 249. По морфологическим признакам В- и Т-лимфоциты и их субпопуляции различать затруднительно (практически невозможно). Все лимфоциты имеют
- 251. Образование В-лимфоцитов начинается в эмбриональном периоде и продолжается в течение всей жизни. В развитии В-лимфоцитов, как
- 252. CD34+ CD10+ CD19+ CD34+ CD10+ CD19+ CD34+ CD10+ CD19+ IgM+ CD34+ CD10+ CD19+ cIgM+ CD34+ CD10+
- 253. CD34+ CD10+ CD19+ CD34+ CD10+ CD19+ CD34+ CD10+ CD19+ Костный мозг Селезенка Кровь CLP Про-В-клетки В-лимфоциты
- 254. CD34+ CD10+ CD19+ IgM+ Костный мозг Селезенка Кровь Большая пре-В-клетка Экспрессия иммуноглобулинов начинается на уровне пре-В-клетки:
- 255. CD34+ CD10+ CD19+ cIgM+ Костный мозг Селезенка Кровь Малая пре-В-клетка А затем цитоплазматического IgM (cIgM) CD34+
- 256. CD34+ CD10+ CD19+ cIgM+ CD34+ CD10+ CD19+ CD20+ CD40+ sIgM+ Костный мозг Селезенка Кровь Малая пре-В-клетка
- 257. CD34+ CD10+ CD19+ CD20+ CD40+ sIgM+ CD20+ CD21+ sIgM+ sIgD+ Костный мозг Селезенка Кровь Незрелая В-клетка
- 258. Родоначальник В-лимфопоэза
- 259. Негативная селекция и делеция клонов. Связывание мембранного антигена незрелой B-клеткой (экспрессирует IgM-BCR, но ещё отсутствует IgD-BCR)
- 261. Незрелая В-клетка КРОВЬ Т1 Транзиторная В-клетка Фенотип IgM+, IgD-, CD21-, CD23- Т2 СЕЛЕЗЕНКА Фенотип IgM+, IgD+,
- 265. В1 В2 В-клетки маргинальной зоны именно эти клетки мы привыкли называть собственно В-лимфоцитами (Marginal Zone B,
- 266. Предназначение B1-лимфоцитов - быстрый ответ на проникающие в организм широко распространённые патогены (преимущественно бактерии). Почти все
- 267. Предполагают, что основная функция B1a-клеток - секреция естественных антител. Считается, что B1b-лимфоциты участвуют в продукции антител
- 269. В2-клетки относятся к подсистеме адаптивного иммунитета, являясь основными клетками гуморального иммунного ответа.
- 271. Их основной задачей является ответ на Т-независимые антигены, находящиеся в крови. Поскольку для ответа на такие
- 274. Антитела Антигенраспознающие молекулы (АРМ) являются основными молекулами системы адаптивного иммунитета. Секретируемые иммуноглобулины (антитела) представляют собой единственный
- 275. История открытия Антитела были открыты в 1890 г., когда Э. Беринг и С. Китасато установили, что
- 276. Строение иммуноглобулинов Молекула Ig - мономер, состоит из двух лёгких (от light) и двух тяжёлых (от
- 277. Схема строения иммуноглобулина (на примере IgGl) L-цепь H-цепь
- 278. Молекула IgG Трёхмерная модель молекулы IgG, построенная на основе рентгеноструктурного анализа. Жёлтым и синим окрашены тяжёлые
- 279. Изотипия иммуноглобулинов Изотипические детерминанты специфичны для разновидностей Н- и L-цепей. Они локализуются в их С-доменах (в
- 280. Изотипы иммуноглобулинов
- 281. Аллотипия и идиотипия иммуноглобулинов Аллотипические детерминанты отражают генетический полиморфизм полипептидных цепей иммуноглобулинов, являясь аллельными продуктами полиморфных
- 282. Локализация антигенных детерминант, характеризующих изотипию, аллотипию и идиотипию иммуноглобулинов
- 283. Функции антител Проявлением эффекторной функции собственно антител является блокада АГ, приводящая к нейтрализации опасных молекул (например,
- 284. Основные механизмы реализации действия антител
- 285. Возрастная динамика уровней основных изотипов иммуноглобулинов человека
- 286. К моменту рождения успевает в значительной степени сформироваться только механизм выработки IgM-антител; полностью он стабилизируется к
- 287. Взаимодействие антигена с антителом В основе реакции антиген-антитело лежит взаимодействие между эпитопом антигена и активным центром
- 288. Нековалентные связи, обеспечивающие взаимодействие антигена с антителом Ионные связи - образуются противоположно заряженными химическими группами, чаще
- 289. Афинность антител Аффинность антител рассматривается как показатель специфичности антител к данному эпитопу. Это мера прочности связи
- 290. Авидность и валентность АТ Авидность (функциональная аффинность) - суммарное сродство. Это сила, с которой мультивалентное АТ
- 292. Скачать презентацию

Иммунитет– это биологическое свойство многоклеточных организмов, обеспечивающее постоянство внутренней среды, по
Иммунитет– это биологическое свойство многоклеточных организмов, обеспечивающее постоянство внутренней среды, по
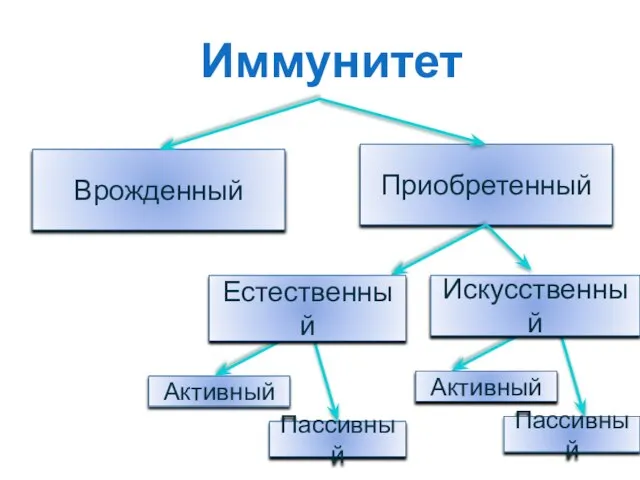
Врожденный
Приобретенный
Иммунитет
Активный
Пассивный
Пассивный
Активный
Искусственный
Естественный
Врожденный
Приобретенный
Иммунитет
Активный
Пассивный
Пассивный
Активный
Искусственный
Естественный
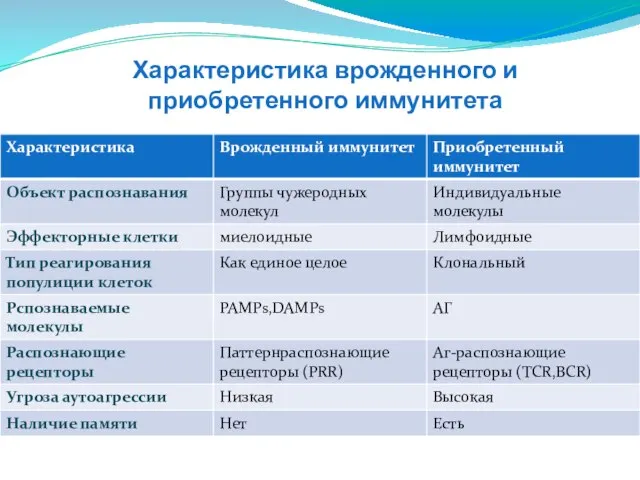
Характеристика врожденного и приобретенного иммунитета
Характеристика врожденного и приобретенного иммунитета
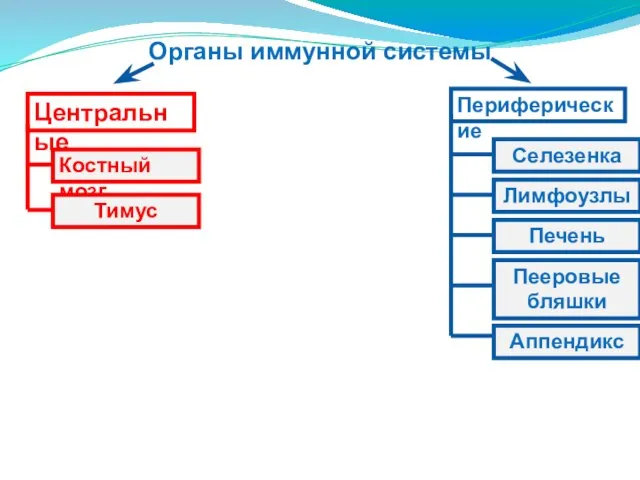
Органы иммунной системы
Центральные
Периферические
Костный мозг
Тимус
Селезенка
Аппендикс
Пееровые бляшки
Печень
Лимфоузлы
Органы иммунной системы
Центральные
Периферические
Костный мозг
Тимус
Селезенка
Аппендикс
Пееровые бляшки
Печень
Лимфоузлы
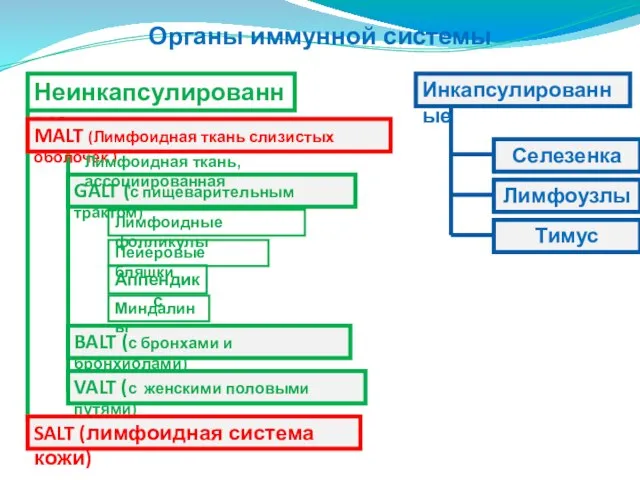
Органы иммунной системы
Неинкапсулированные
MALT (Лимфоидная ткань слизистых оболочек )
GALT (с пищеварительным трактом)
Селезенка
Тимус
Лимфоузлы
Инкапсулированные
BALT
Органы иммунной системы
Неинкапсулированные
MALT (Лимфоидная ткань слизистых оболочек )
GALT (с пищеварительным трактом)
Селезенка
Тимус
Лимфоузлы
Инкапсулированные
BALT


Неспецифическая резистентность организма - устойчивость организма к любым
патогенным воздействиям, независимо
Неспецифическая резистентность организма - устойчивость организма к любым
патогенным воздействиям, независимо

Эффекторные механизмы защиты организма
1.Фагоцитоз
-Нейтрофиллы
-Моноциты
-Макрофаги
2.Экзоцитоз
-эозинофилы
-базофилы
-мастоциты
3.Цитолиз
-Комплемент
-NK-клетки
-ЦТЛ
4.Нейтрализация (антитоксические АТ)
5.Образование гранулем (макрофаги,
Эффекторные механизмы защиты организма
1.Фагоцитоз
-Нейтрофиллы
-Моноциты
-Макрофаги
2.Экзоцитоз
-эозинофилы
-базофилы
-мастоциты
3.Цитолиз
-Комплемент
-NK-клетки
-ЦТЛ
4.Нейтрализация (антитоксические АТ)
5.Образование гранулем (макрофаги,
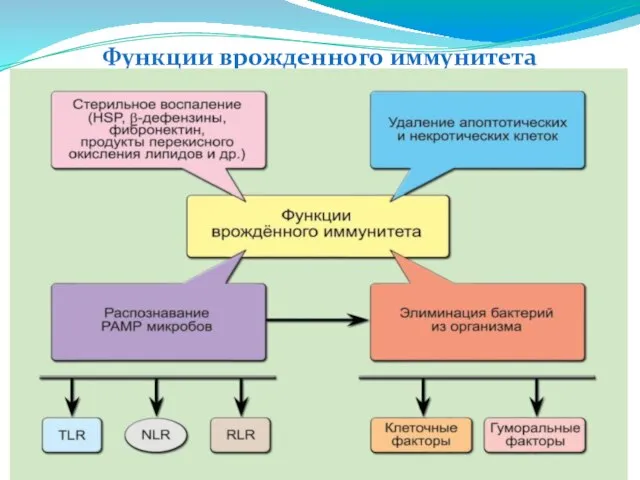
Функции врожденного иммунитета
Функции врожденного иммунитета

Основные факторы врожденного
иммунитета
Клеточные факторы
Основные факторы врожденного
иммунитета
Клеточные факторы
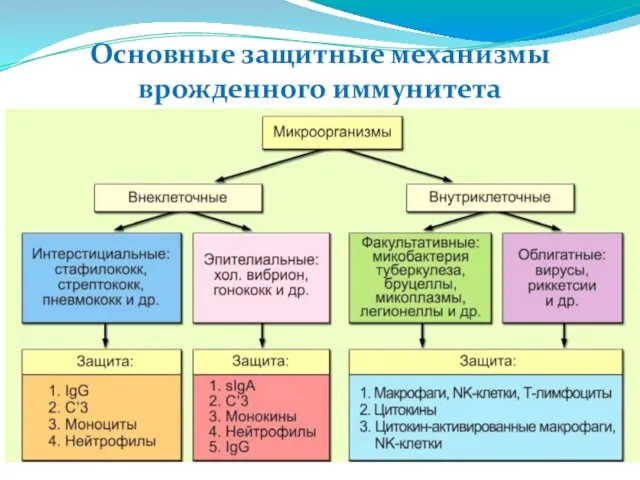
Основные защитные механизмы
врожденного иммунитета
Основные защитные механизмы
врожденного иммунитета

Фазы ответной реакции
врожденного иммунитета
Фазы ответной реакции
врожденного иммунитета

КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
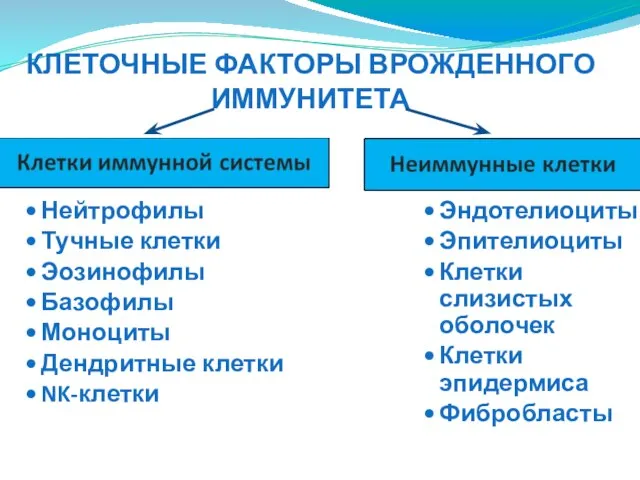
КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
Нейтрофилы
Тучные клетки
Эозинофилы
Базофилы
Моноциты
Дендритные клетки
NK-клетки
Эндотелиоциты
Эпителиоциты
Клетки слизистых оболочек
Клетки эпидермиса
Фибробласты
КЛЕТОЧНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
Нейтрофилы
Тучные клетки
Эозинофилы
Базофилы
Моноциты
Дендритные клетки
NK-клетки
Эндотелиоциты
Эпителиоциты
Клетки слизистых оболочек
Клетки эпидермиса
Фибробласты
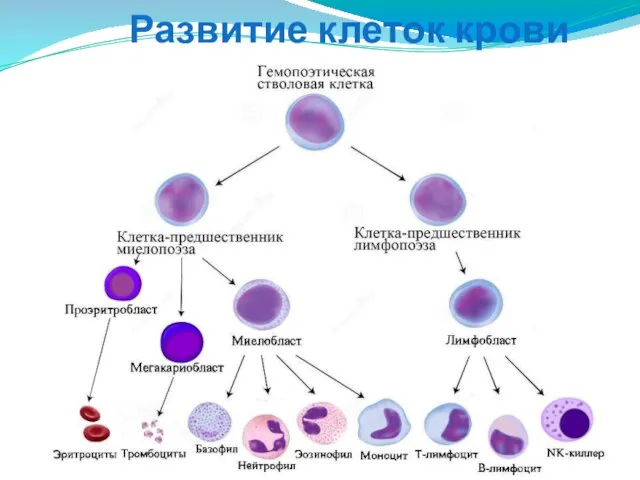
Развитие клеток крови
Развитие клеток крови
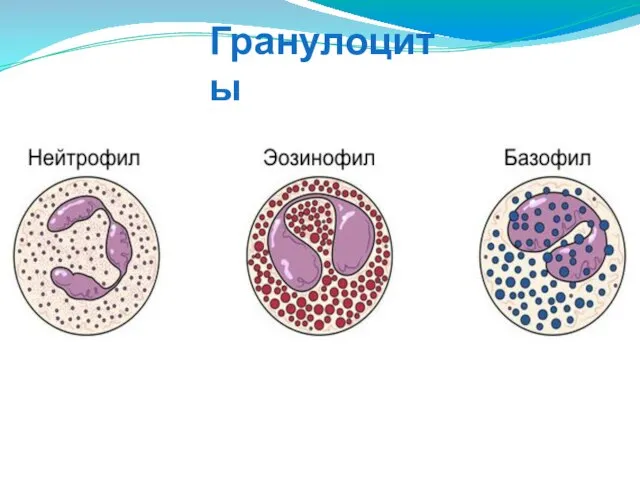
Гранулоциты
Гранулоциты
Фагоцитирующие клетки
Моноциты
Макрофаги
Нейтрофилы
Фагоцитирующие клетки
Моноциты
Макрофаги
Нейтрофилы
1.Развитие нейтрофилов
2.Морфология клетки
3. Гранулы нейтрофилов и их свойства
4. Кластерные молекулы
1.Развитие нейтрофилов
2.Морфология клетки
3. Гранулы нейтрофилов и их свойства
4. Кластерные молекулы
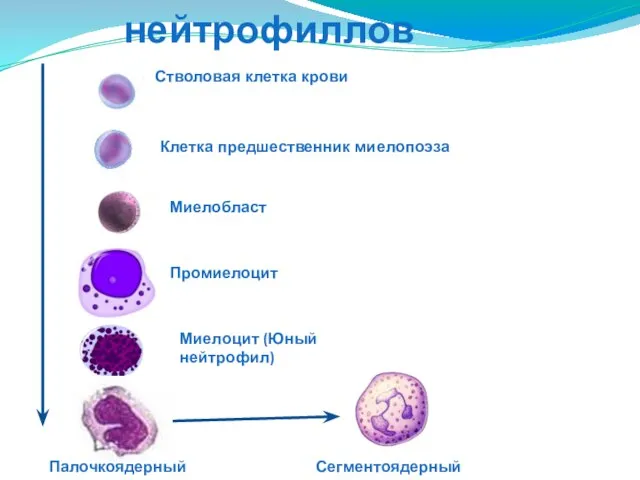
Развитие нейтрофиллов
Стволовая клетка крови
Клетка предшественник миелопоэза
Миелобласт
Промиелоцит
Миелоцит (Юный нейтрофил)
Палочкоядерный
Развитие нейтрофиллов
Стволовая клетка крови
Клетка предшественник миелопоэза
Миелобласт
Промиелоцит
Миелоцит (Юный нейтрофил)
Палочкоядерный
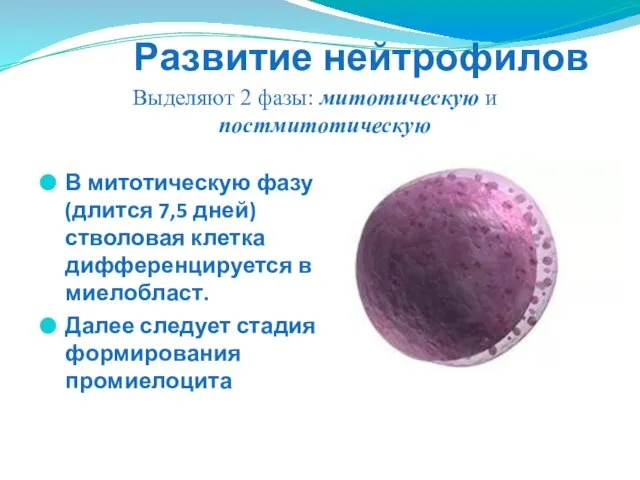
Развитие нейтрофилов
В митотическую фазу (длится 7,5 дней) стволовая клетка дифференцируется в
Развитие нейтрофилов
В митотическую фазу (длится 7,5 дней) стволовая клетка дифференцируется в
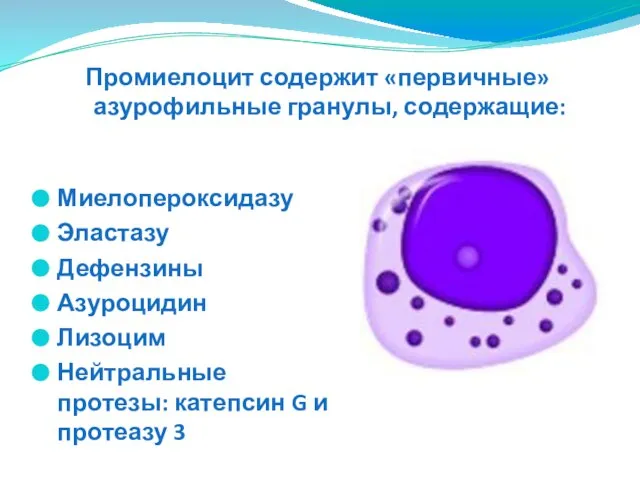
Миелопероксидазу
Эластазу
Дефензины
Азуроцидин
Лизоцим
Нейтральные протезы: катепсин G и протеазу 3
Промиелоцит содержит «первичные» азурофильные
Эластазу
Дефензины
Азуроцидин
Лизоцим
Нейтральные протезы: катепсин G и протеазу 3
Промиелоцит содержит «первичные» азурофильные

Лактоферрин
Лизоцим
Фосфолипазу Ф2
Белок, повышающий проницаемость микробной стенки
Белок NGAI
Липокартин
Кателицидин
В12 связывающий протеин
На стадии миелоцита
Лизоцим
Фосфолипазу Ф2
Белок, повышающий проницаемость микробной стенки
Белок NGAI
Липокартин
Кателицидин
В12 связывающий протеин
На стадии миелоцита
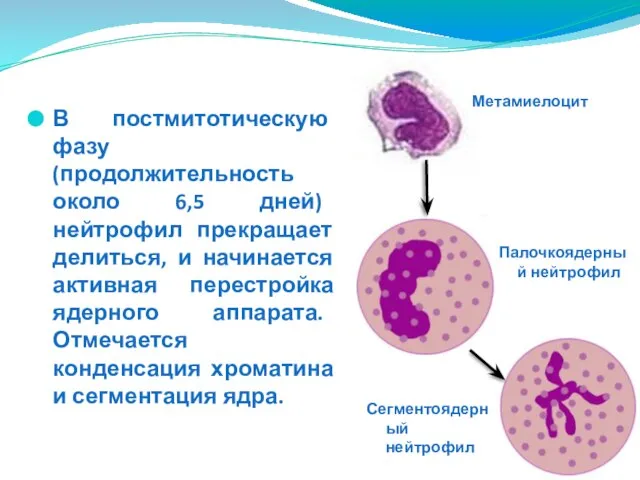
В постмитотическую фазу (продолжительность около 6,5 дней) нейтрофил прекращает делиться, и
В постмитотическую фазу (продолжительность около 6,5 дней) нейтрофил прекращает делиться, и
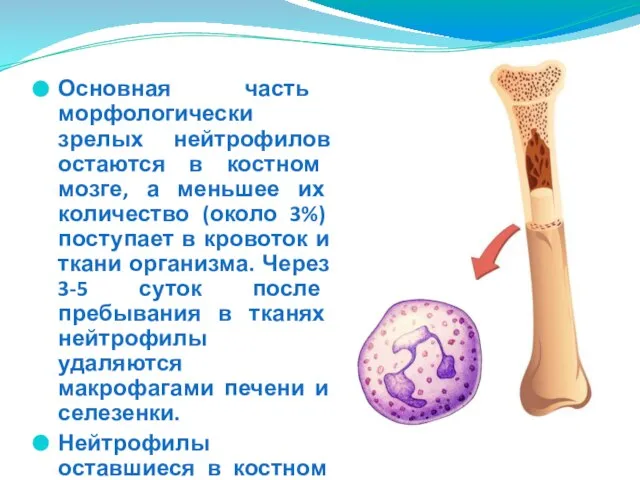
Основная часть морфологически зрелых нейтрофилов остаются в костном мозге, а меньшее
Основная часть морфологически зрелых нейтрофилов остаются в костном мозге, а меньшее

Морфология
Нейтрофильные сегментоядерные
лейкоциты - преобладающая
популяция белых клеток крови.
Клетки диаметром
Морфология
Нейтрофильные сегментоядерные
лейкоциты - преобладающая
популяция белых клеток крови.
Клетки диаметром

Гранулы нейтрофилов
При дегрануляции азурофильные гранулы первыми сливаются с фагосомами, затем -
Гранулы нейтрофилов
При дегрануляции азурофильные гранулы первыми сливаются с фагосомами, затем -
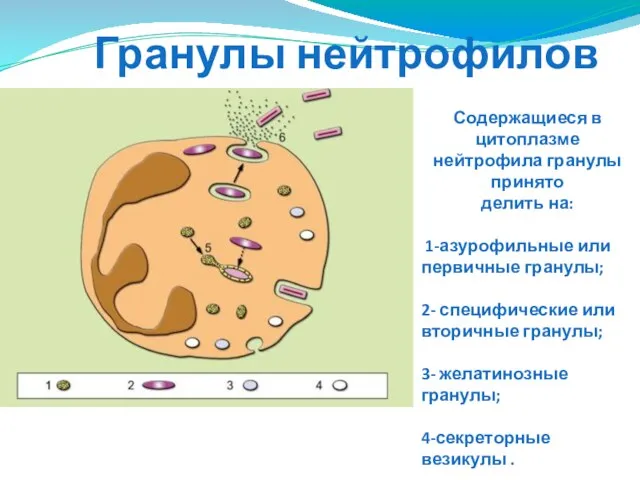
Гранулы нейтрофилов
Содержащиеся в цитоплазме нейтрофила гранулы принято
делить на:
1-азурофильные или
Гранулы нейтрофилов
Содержащиеся в цитоплазме нейтрофила гранулы принято
делить на:
1-азурофильные или

Свойства и состав гранул нейтрофилов
Свойства и состав гранул нейтрофилов

Все указанные вещества обладают антимикробным действием
Миелопероксидаза (MPO) составляет 2–4% массы нейтрофила, катализирует образование
Все указанные вещества обладают антимикробным действием
Миелопероксидаза (MPO) составляет 2–4% массы нейтрофила, катализирует образование

Катепсин G при нейтральном рН убивает грамположительные и грамотрицательные бактерии. Дефицит катепсина G и связанное
Катепсин G при нейтральном рН убивает грамположительные и грамотрицательные бактерии. Дефицит катепсина G и связанное
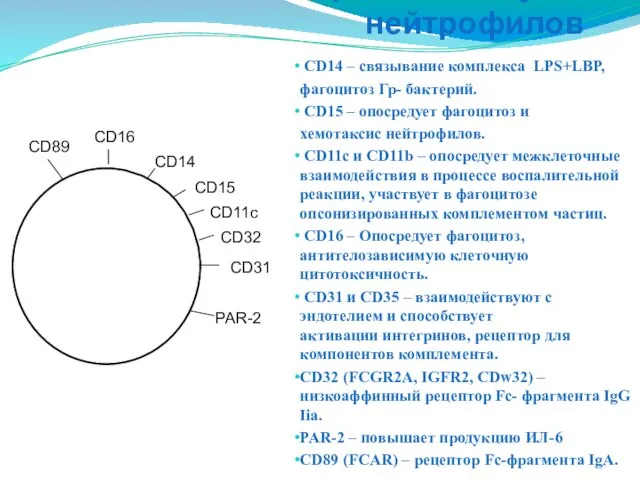
СD14 – связывание комплекса LPS+LBP,
фагоцитоз Гр- бактерий.
CD15 –
СD14 – связывание комплекса LPS+LBP,
фагоцитоз Гр- бактерий.
CD15 –
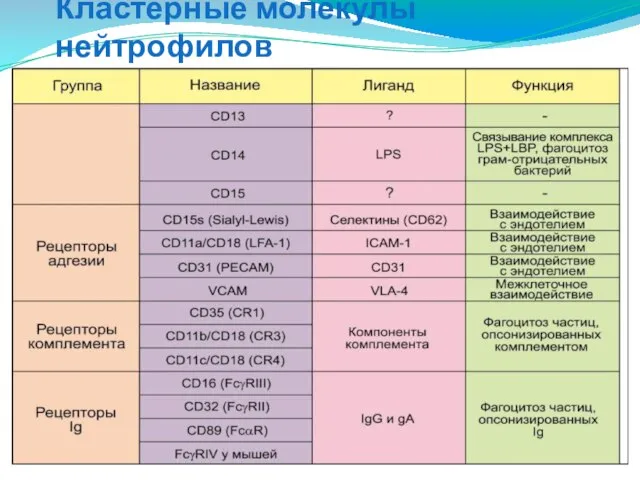
Кластерные молекулы нейтрофилов
Кластерные молекулы нейтрофилов
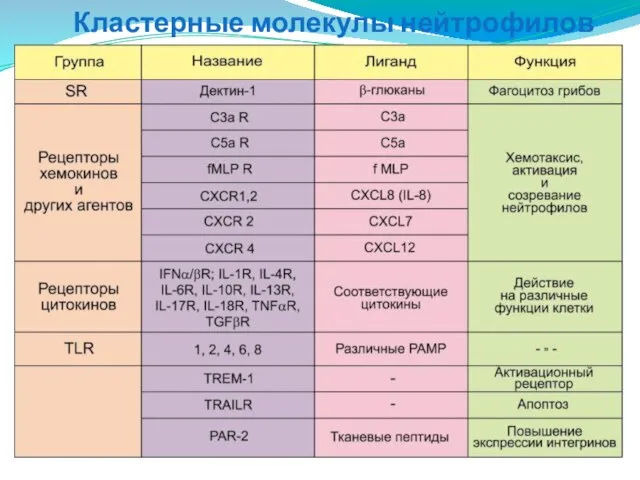
Кластерные молекулы нейтрофилов
Кластерные молекулы нейтрофилов
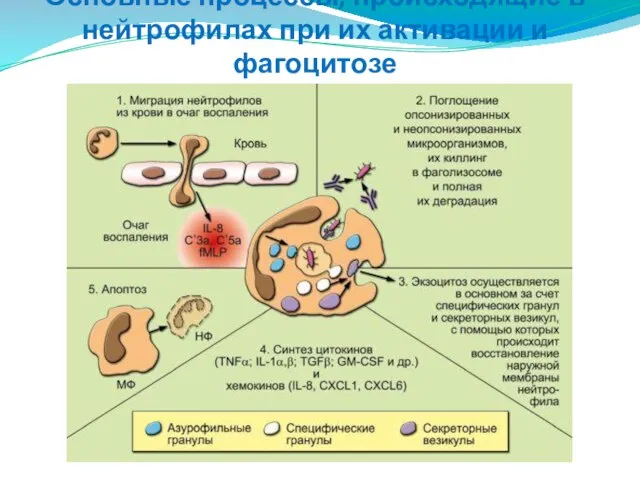
Основные процессы, происходящие в нейтрофилах при их активации и фагоцитозе
Основные процессы, происходящие в нейтрофилах при их активации и фагоцитозе

На рисунке отражены участие НФ в фагоцитозе (миграция, поглощение, дегрануляция, внутриклеточный
На рисунке отражены участие НФ в фагоцитозе (миграция, поглощение, дегрануляция, внутриклеточный
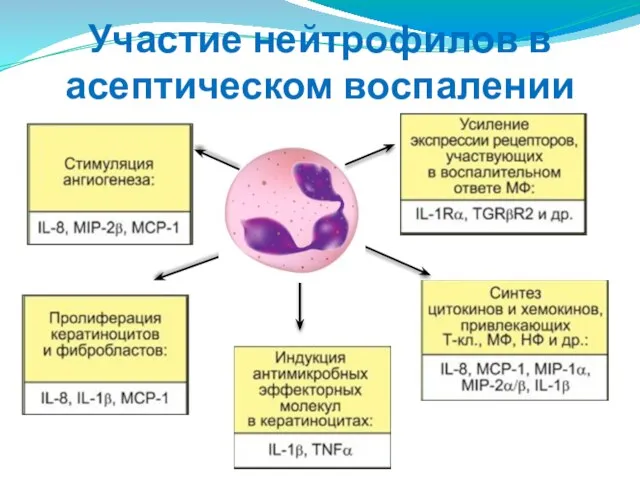
Участие нейтрофилов в асептическом воспалении
Участие нейтрофилов в асептическом воспалении

1.Развитие моноцитов
2.Морфология моноцитов
3. Моноциты периферической крови
4. Развитие макрофагов
5. Морфология макрофагов
6.Кластерные
1.Развитие моноцитов
2.Морфология моноцитов
3. Моноциты периферической крови
4. Развитие макрофагов
5. Морфология макрофагов
6.Кластерные
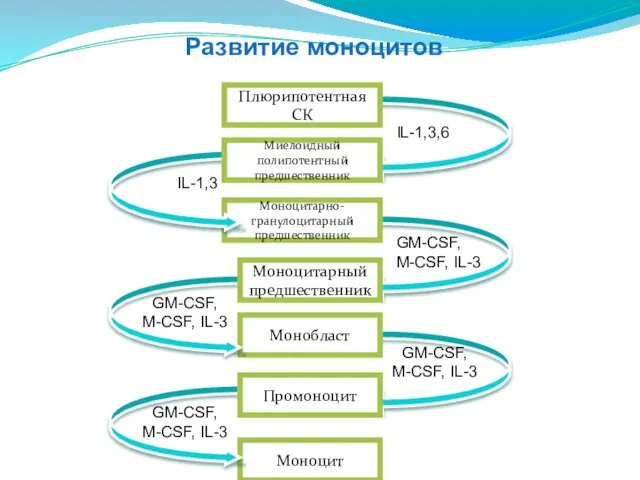
Моноцит
Монобласт
Моноцитарно-гранулоцитарный предшественник
Плюрипотентная СК
Моноцитарный предшественник
Промоноцит
Миелоидный полипотентный предшественник
IL-1,3,6
IL-1,3
GM-CSF, M-CSF, IL-3
GM-CSF, M-CSF, IL-3
GM-CSF, M-CSF,
Моноцит
Монобласт
Моноцитарно-гранулоцитарный предшественник
Плюрипотентная СК
Моноцитарный предшественник
Промоноцит
Миелоидный полипотентный предшественник
IL-1,3,6
IL-1,3
GM-CSF, M-CSF, IL-3
GM-CSF, M-CSF, IL-3
GM-CSF, M-CSF,
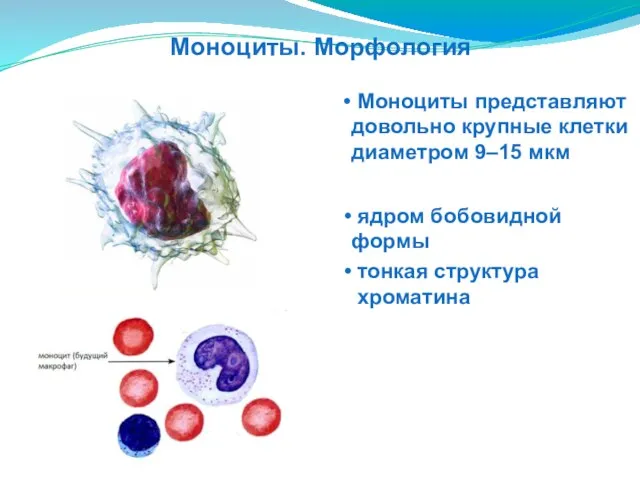
тонкая структура
хроматина
Моноциты представляют довольно крупные клетки
тонкая структура
хроматина
Моноциты представляют довольно крупные клетки
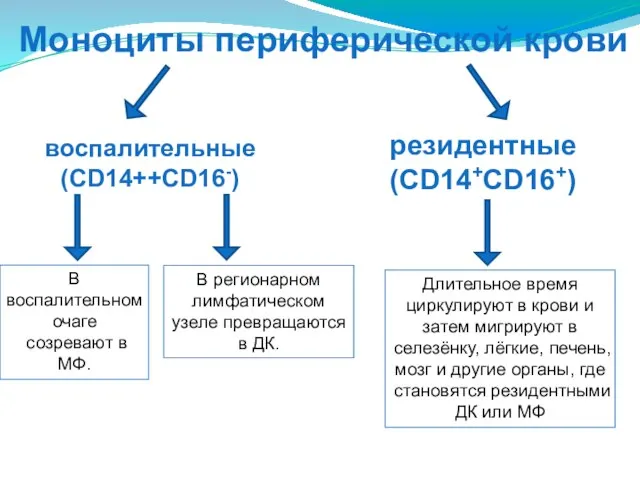
Моноциты периферической крови
воспалительные
(CD14++CD16-)
резидентные
(CD14+CD16+)
В воспалительном очаге
созревают в МФ.
В регионарном
Моноциты периферической крови
воспалительные
(CD14++CD16-)
резидентные
(CD14+CD16+)
В воспалительном очаге
созревают в МФ.
В регионарном
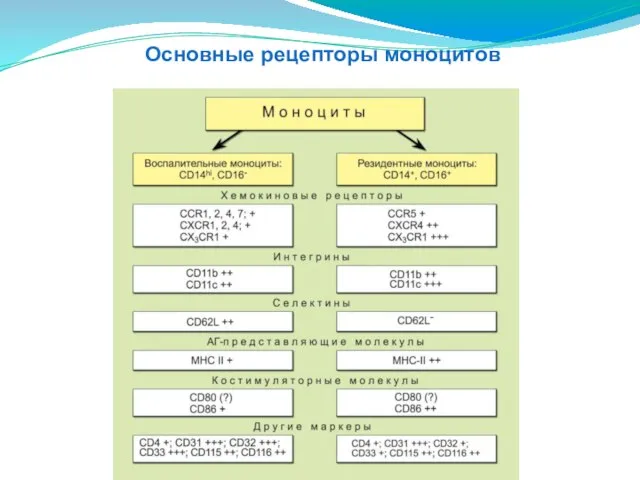
Основные рецепторы моноцитов
Основные рецепторы моноцитов
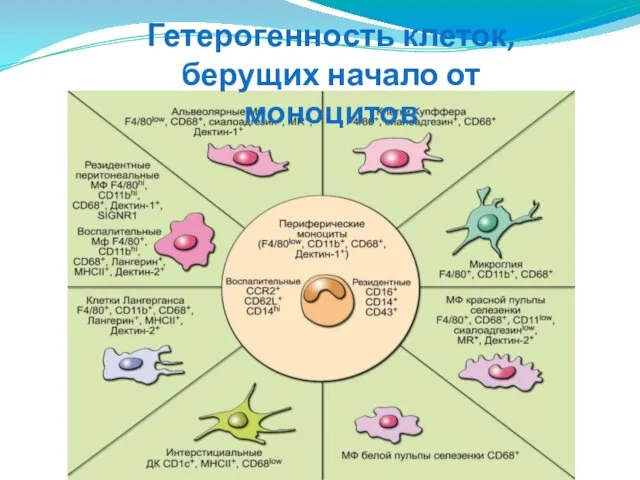
Гетерогенность клеток,
берущих начало от моноцитов
Гетерогенность клеток,
берущих начало от моноцитов
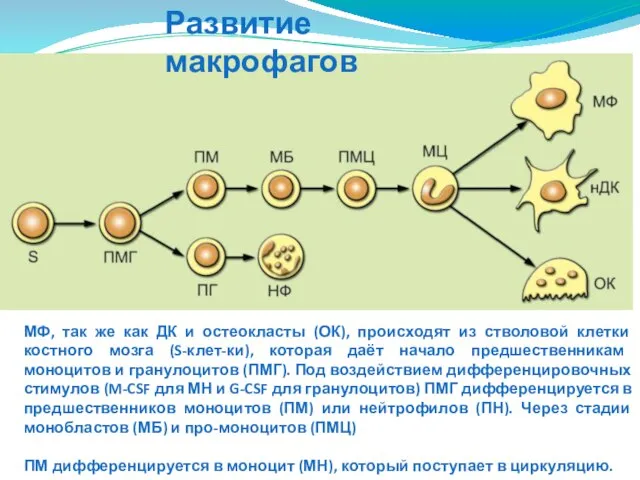
Развитие макрофагов
МФ, так же как ДК и остеокласты (ОК), происходят из
Развитие макрофагов
МФ, так же как ДК и остеокласты (ОК), происходят из

изменяется экспрессии некоторых мембранных молекул (ослабевает экспрессия CD13, CD14, CD15,
изменяется экспрессии некоторых мембранных молекул (ослабевает экспрессия CD13, CD14, CD15,
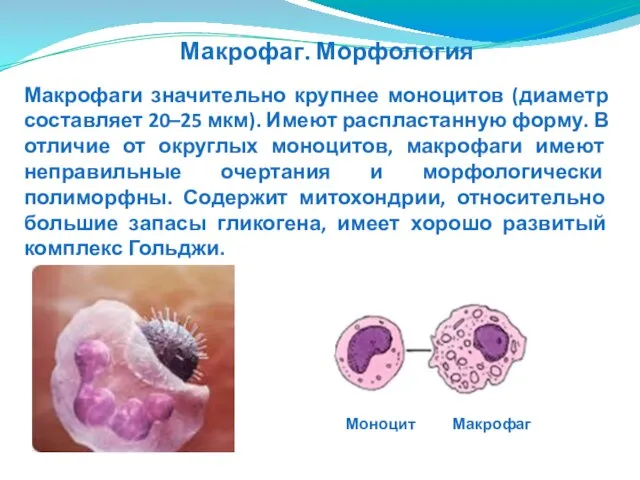
Макрофаги значительно крупнее моноцитов (диаметр составляет 20–25 мкм). Имеют распластанную форму.
Макрофаги значительно крупнее моноцитов (диаметр составляет 20–25 мкм). Имеют распластанную форму.
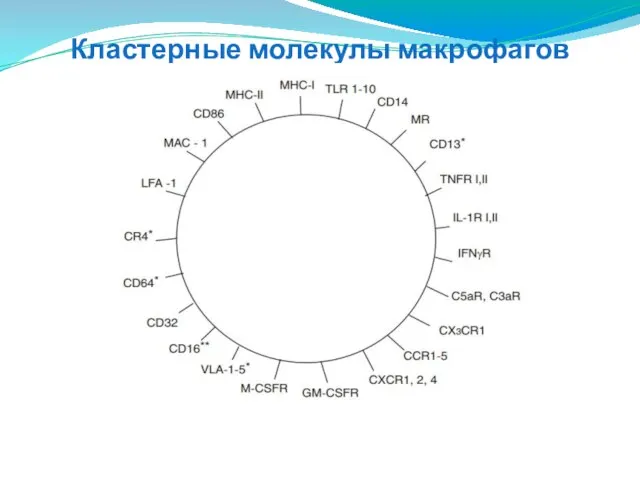
Кластерные молекулы макрофагов
Кластерные молекулы макрофагов
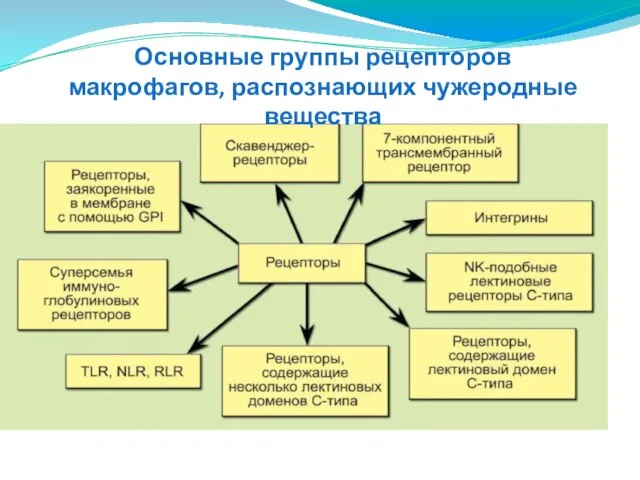
Основные группы рецепторов макрофагов, распознающих чужеродные вещества
Основные группы рецепторов макрофагов, распознающих чужеродные вещества
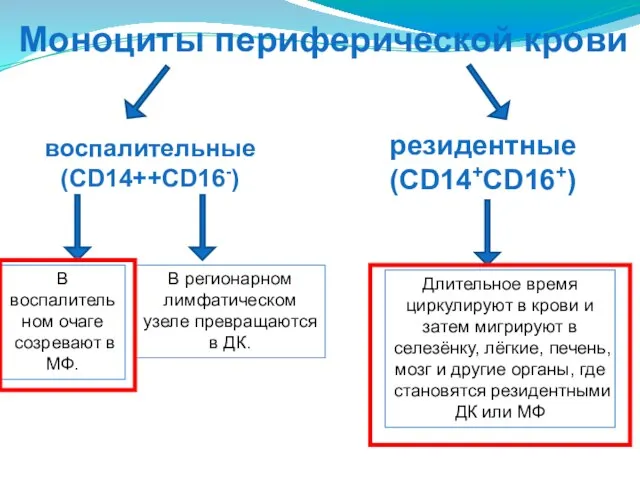
Моноциты периферической крови
воспалительные
(CD14++CD16-)
резидентные
(CD14+CD16+)
В воспалительном очаге
созревают в МФ.
В регионарном
Моноциты периферической крови
воспалительные
(CD14++CD16-)
резидентные
(CD14+CD16+)
В воспалительном очаге
созревают в МФ.
В регионарном

Моноциты воспалительные
(CD14++CD16-)
В первые часы острого воспаления моноциты для проникновения используют
Моноциты воспалительные
(CD14++CD16-)
В первые часы острого воспаления моноциты для проникновения используют
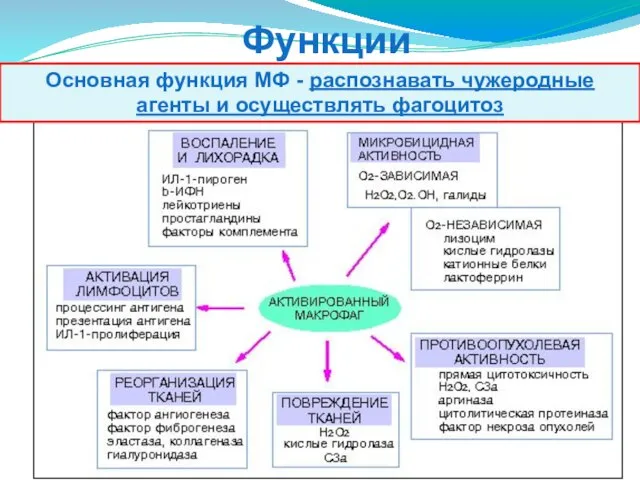
Функции макрофагов
Основная функция МФ - распознавать чужеродные агенты и осуществлять фагоцитоз
Функции макрофагов
Основная функция МФ - распознавать чужеродные агенты и осуществлять фагоцитоз
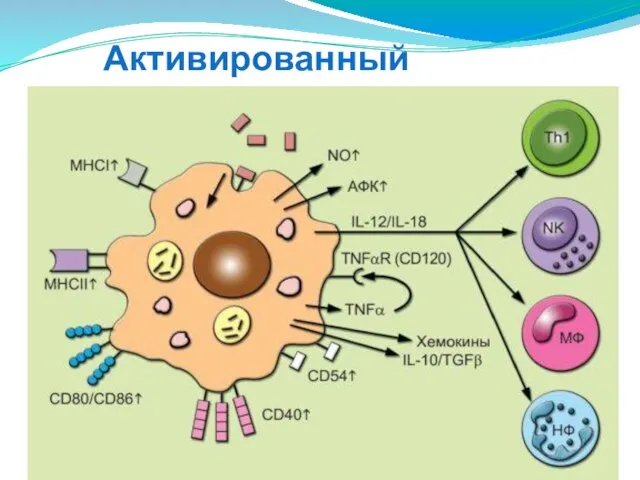
Активированный макрофаг
Активированный макрофаг
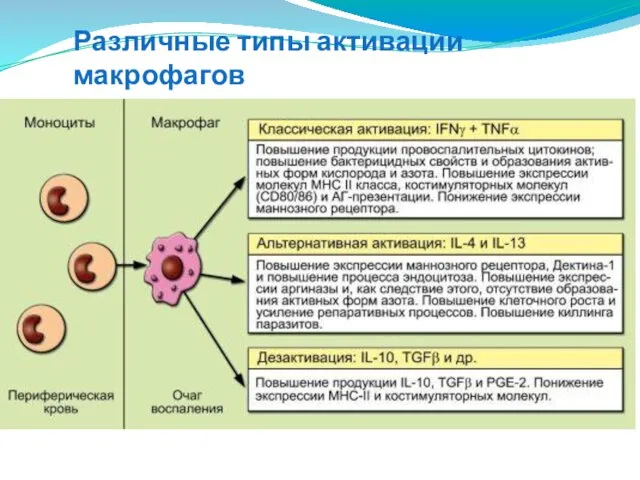
Различные типы активации макрофагов
Различные типы активации макрофагов

Различные типы активации макрофагов
В классическом виде процесс запускается ΙΚΝγ и TNF,
Различные типы активации макрофагов
В классическом виде процесс запускается ΙΚΝγ и TNF,
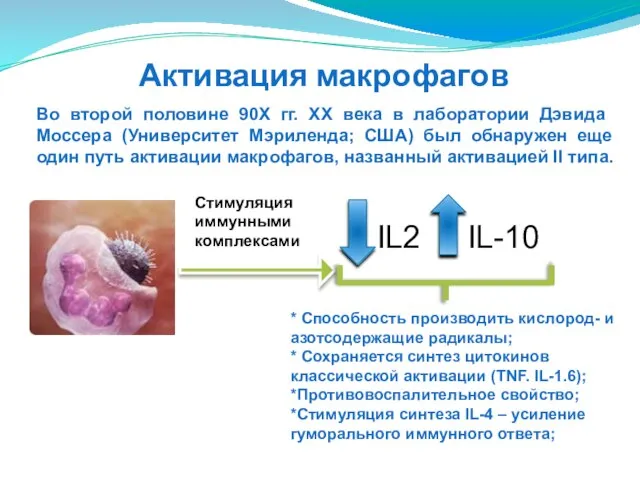
Активация макрофагов
Во второй половине 90X гг. XX века в лаборатории Дэвида
Активация макрофагов
Во второй половине 90X гг. XX века в лаборатории Дэвида
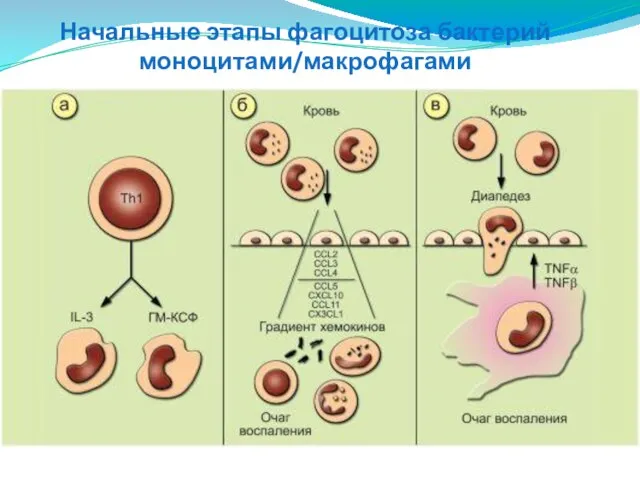
Начальные этапы фагоцитоза бактерий моноцитами/макрофагами
Начальные этапы фагоцитоза бактерий моноцитами/макрофагами

Фагоцитоз
Явление фагоцитоза было открыто И.И. Мечниковым (1882). Он показал фундаментальную роль
Фагоцитоз
Явление фагоцитоза было открыто И.И. Мечниковым (1882). Он показал фундаментальную роль

Стадии фагоцитоза
1. Хемотаксис
2. Адгезия
3. Активация мембраны
4. Погружение;
5.Образование фагосомы;
6.Слияние фагосомы и лизосомы;
7.Киллинг
Стадии фагоцитоза
1. Хемотаксис
2. Адгезия
3. Активация мембраны
4. Погружение;
5.Образование фагосомы;
6.Слияние фагосомы и лизосомы;
7.Киллинг
Стадии фагоцитоза
Стадии фагоцитоза
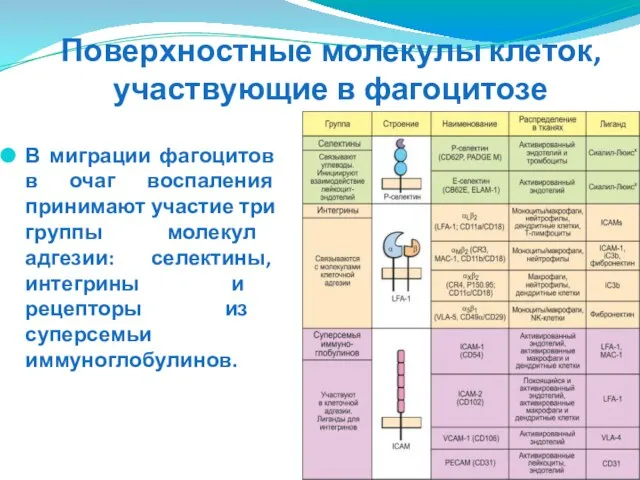
Поверхностные молекулы клеток, участвующие в фагоцитозе
В миграции фагоцитов в очаг воспаления
Поверхностные молекулы клеток, участвующие в фагоцитозе
В миграции фагоцитов в очаг воспаления
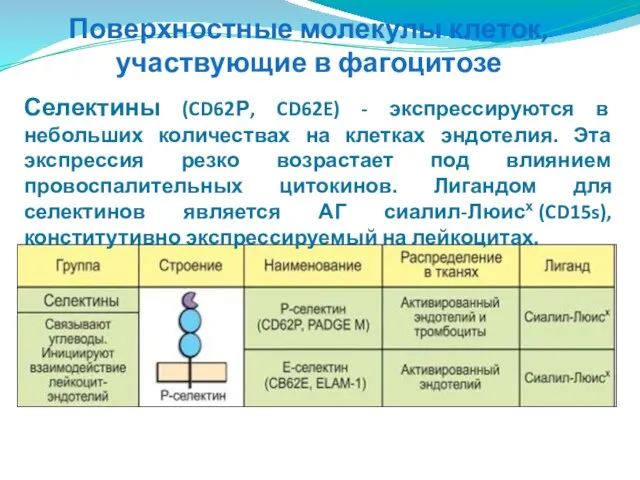
Поверхностные молекулы клеток, участвующие в фагоцитозе
Селектины (CD62Р, CD62E) - экспрессируются в
Поверхностные молекулы клеток, участвующие в фагоцитозе
Селектины (CD62Р, CD62E) - экспрессируются в
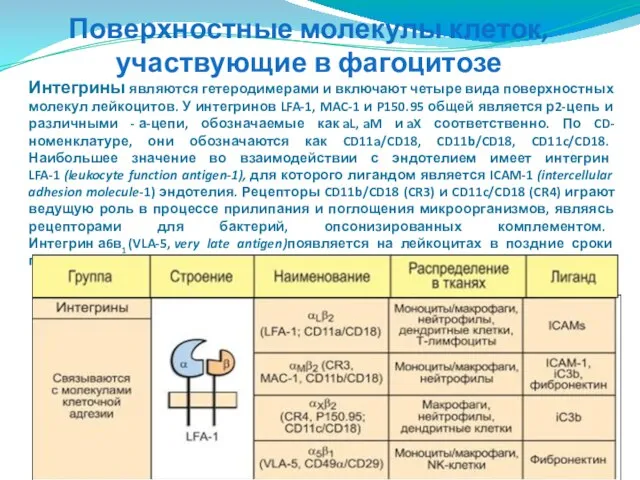
Поверхностные молекулы клеток, участвующие в фагоцитозе
Интегрины являются гетеродимерами и включают четыре
Поверхностные молекулы клеток, участвующие в фагоцитозе
Интегрины являются гетеродимерами и включают четыре
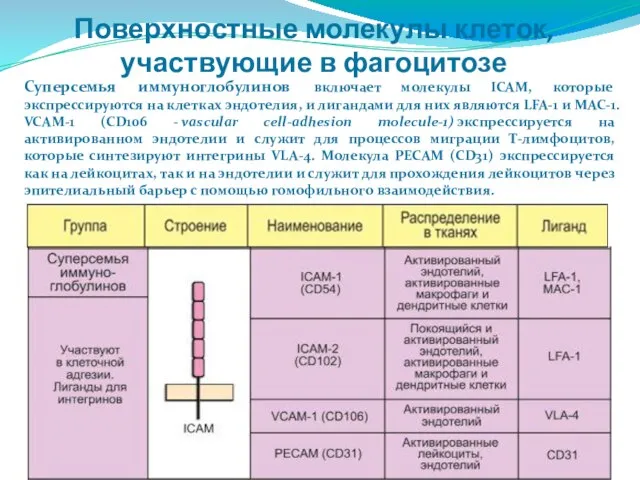
Поверхностные молекулы клеток, участвующие в фагоцитозе
Суперсемья иммуноглобулинов включает молекулы ICAM, которые
Поверхностные молекулы клеток, участвующие в фагоцитозе
Суперсемья иммуноглобулинов включает молекулы ICAM, которые
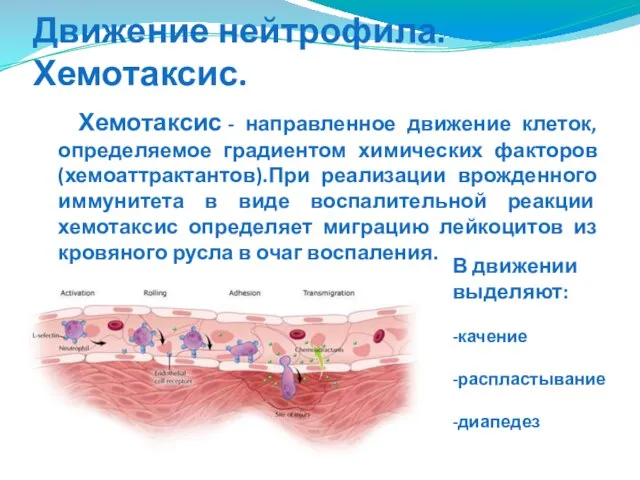
Движение нейтрофила. Хемотаксис.
Хемотаксис - направленное движение клеток, определяемое градиентом химических факторов
Движение нейтрофила. Хемотаксис.
Хемотаксис - направленное движение клеток, определяемое градиентом химических факторов
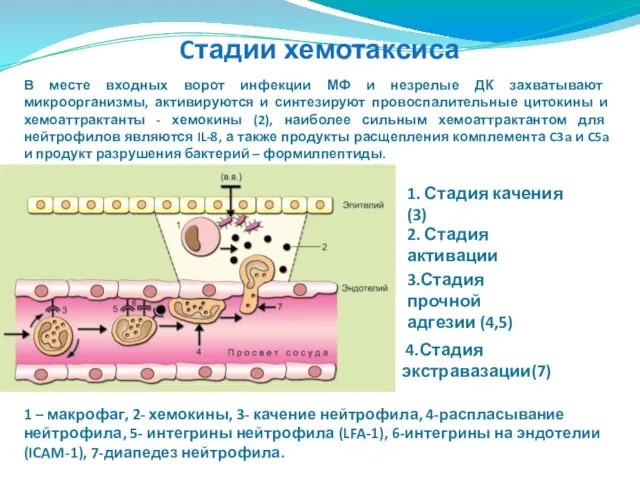
Cтадии хемотаксиса
1. Стадия качения (3)
2. Стадия активации
3.Стадия прочной
адгезии
Cтадии хемотаксиса
1. Стадия качения (3)
2. Стадия активации
3.Стадия прочной
адгезии

Адгезия
Обязательным условием адгезии фагоцита служит распознавание объекта фагоцитоза. Механизмы распознавания разнообразны
Адгезия
Обязательным условием адгезии фагоцита служит распознавание объекта фагоцитоза. Механизмы распознавания разнообразны

Scavenger-рецепторы (рецепторы-мусорщики).
Эти рецепторы экспрессированы на макрофагах и некоторых дендритных
Scavenger-рецепторы (рецепторы-мусорщики).
Эти рецепторы экспрессированы на макрофагах и некоторых дендритных
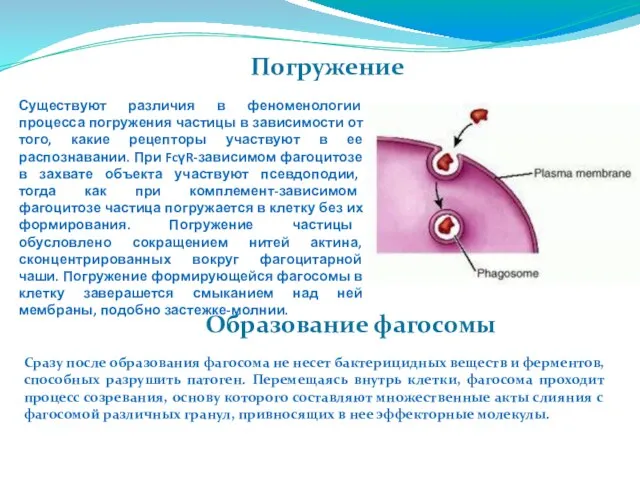
Погружение
Существуют различия в феноменологии процесса погружения частицы в зависимости от того,
Погружение
Существуют различия в феноменологии процесса погружения частицы в зависимости от того,
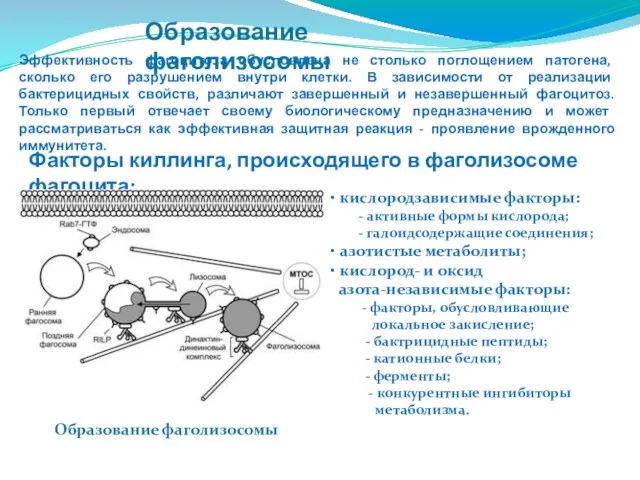
Образование фаголизосомы
Образование фаголизосомы
Эффективность фагоцитоза обусловлена не столько поглощением патогена, сколько его
Образование фаголизосомы
Образование фаголизосомы
Эффективность фагоцитоза обусловлена не столько поглощением патогена, сколько его

Эозинофилы
1.Развитие эозинофилов
2. Морфология клетки
3. Гранулы эозинофилов и их свойства
4. Кластерные молекулы
Эозинофилы
1.Развитие эозинофилов
2. Морфология клетки
3. Гранулы эозинофилов и их свойства
4. Кластерные молекулы
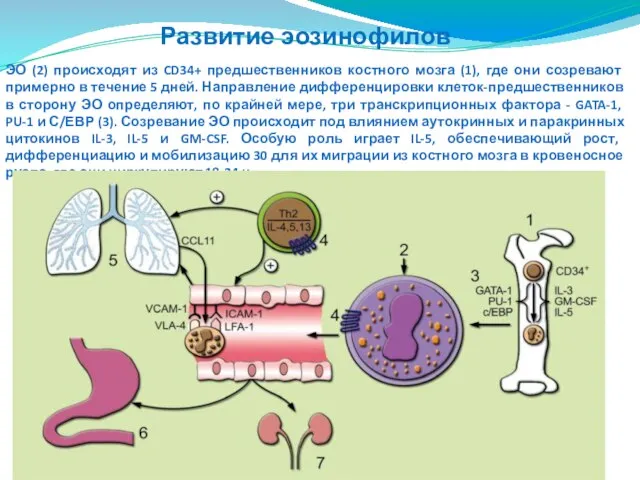
ЭО (2) происходят из CD34+ предшественников костного мозга (1), где они
ЭО (2) происходят из CD34+ предшественников костного мозга (1), где они

МОРФОЛОГИЯ
Эозинофилы составляют 1–5% лейкоцитов, циркулирующих в крови. Эозинофилы в течение нескольких
МОРФОЛОГИЯ
Эозинофилы составляют 1–5% лейкоцитов, циркулирующих в крови. Эозинофилы в течение нескольких

Гранулы эозинофилов
Гранулы эозинофилов
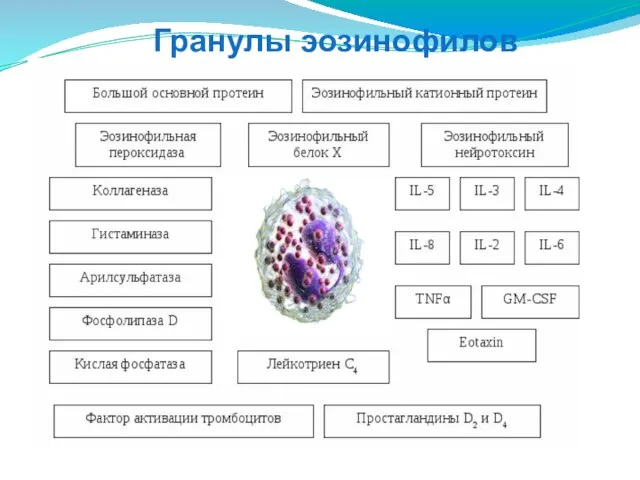
Гранулы эозинофилов
Гранулы эозинофилов
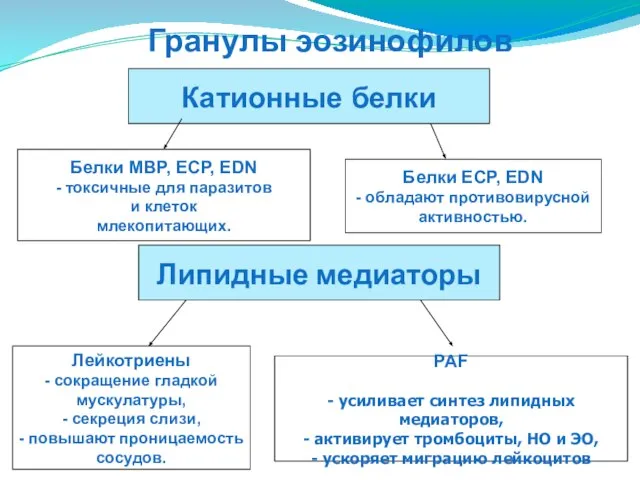
Белки MBP, ECP, EDN
- токсичные для паразитов
и клеток
млекопитающих.
Белки ECP, EDN
-
Белки MBP, ECP, EDN
- токсичные для паразитов
и клеток
млекопитающих.
Белки ECP, EDN -
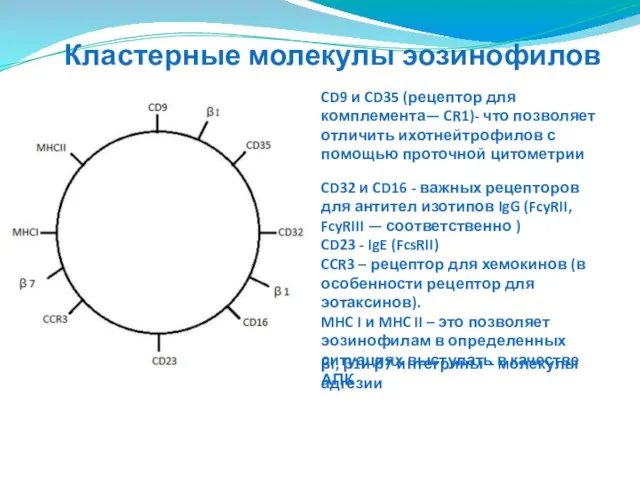
Кластерные молекулы эозинофилов
CD9 и CD35 (рецептор для комплемента— CR1)- что позволяет отличить
Кластерные молекулы эозинофилов
CD9 и CD35 (рецептор для комплемента— CR1)- что позволяет отличить

1. Повышение чувствительности рецепторов к иммуноглобулинам класса Е. За счет этого
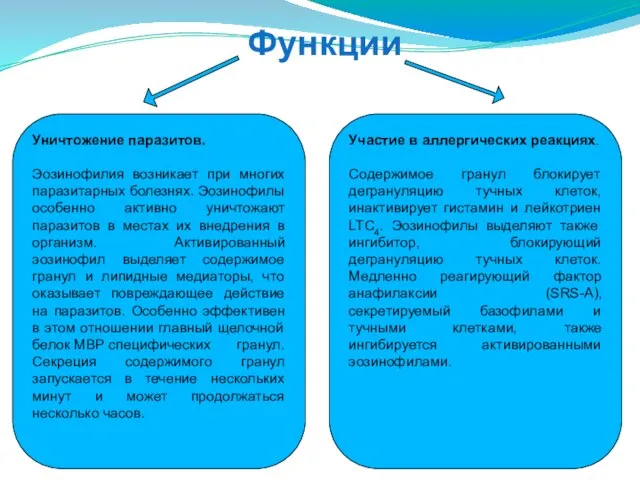
Функции
Уничтожение паразитов.
Эозинофилия возникает при многих паразитарных болезнях. Эозинофилы особенно активно уничтожают
Функции
Уничтожение паразитов.
Эозинофилия возникает при многих паразитарных болезнях. Эозинофилы особенно активно уничтожают

Побочные эффекты
Секретируемые эозинофилом вещества могут повреждать нормальные ткани. Так, при постоянном
Побочные эффекты
Секретируемые эозинофилом вещества могут повреждать нормальные ткани. Так, при постоянном
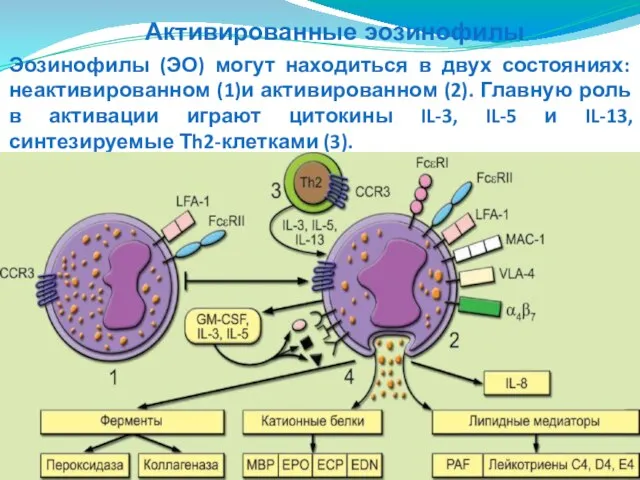
Эозинофилы (ЭО) могут находиться в двух состояниях: неактивированном (1)и активированном (2).
Эозинофилы (ЭО) могут находиться в двух состояниях: неактивированном (1)и активированном (2).
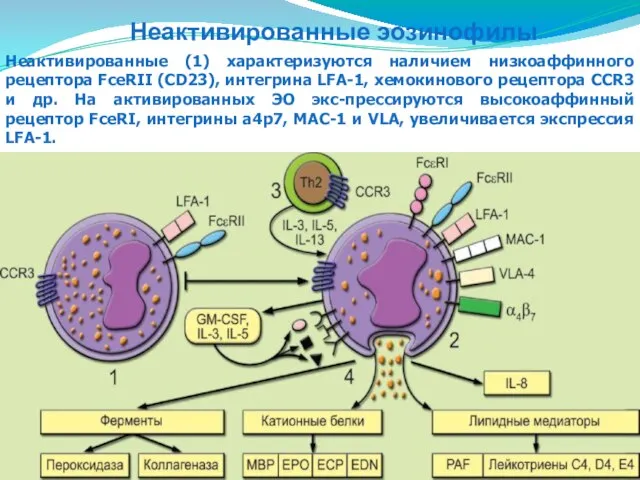
Неактивированные эозинофилы
Неактивированные (1) характеризуются наличием низкоаффинного рецептора FceRII (CD23), интегрина LFA-1,
Неактивированные эозинофилы
Неактивированные (1) характеризуются наличием низкоаффинного рецептора FceRII (CD23), интегрина LFA-1,
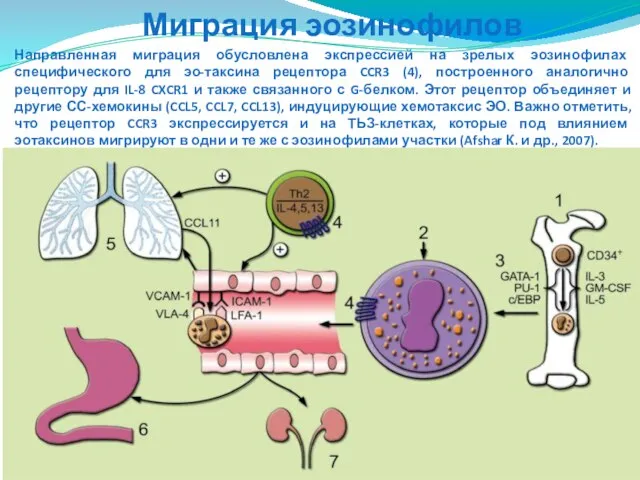
Направленная миграция обусловлена экспрессией на зрелых эозинофилах специфического для эо-таксина рецептора
Направленная миграция обусловлена экспрессией на зрелых эозинофилах специфического для эо-таксина рецептора
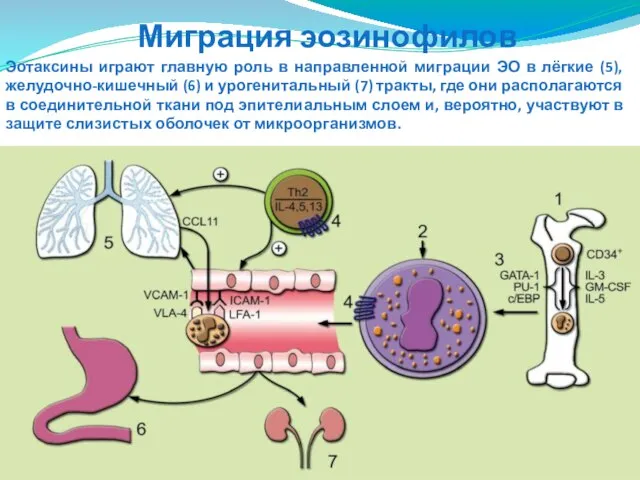
Эотаксины играют главную роль в направленной миграции ЭО в лёгкие (5),
Эотаксины играют главную роль в направленной миграции ЭО в лёгкие (5),
Заболевания, связанные с
эозинофилами
Заболевания, связанные с
эозинофилами
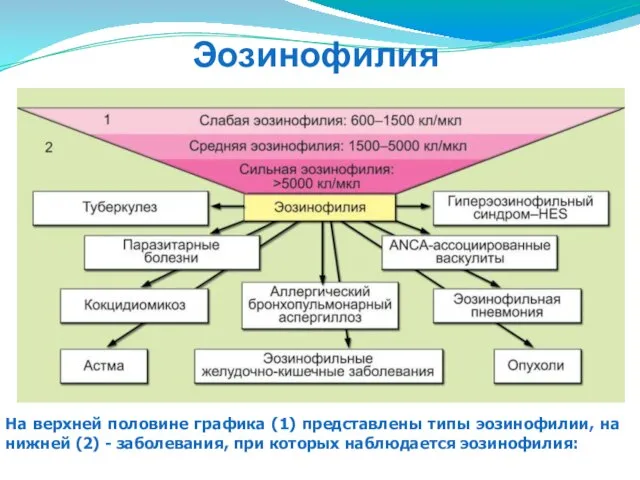
На верхней половине графика (1) представлены типы эозинофилии, на нижней (2)
На верхней половине графика (1) представлены типы эозинофилии, на нижней (2)
Считается, что ЭО играют защитную роль при глистных инвазиях. В экспериментальных
Считается, что ЭО играют защитную роль при глистных инвазиях. В экспериментальных
ЭО участвуют в патогенезе бронхиальной астмы, при которой наблюдаются средний уровень
ЭО участвуют в патогенезе бронхиальной астмы, при которой наблюдаются средний уровень
Особняком от этих заболеваний стоит идиопатический гиперэозинофильный синдром (HES), который характеризуется
Особняком от этих заболеваний стоит идиопатический гиперэозинофильный синдром (HES), который характеризуется

К гиперэозинофильным синдромам относится эозинофильный фибропластический эндокардит, характеризующийся стойкой эозинофилией
К гиперэозинофильным синдромам относится эозинофильный фибропластический эндокардит, характеризующийся стойкой эозинофилией

1.Развитие базофилов и тучных клеток
2. Гранулы базофилов и тучных клеток, их
1.Развитие базофилов и тучных клеток
2. Гранулы базофилов и тучных клеток, их
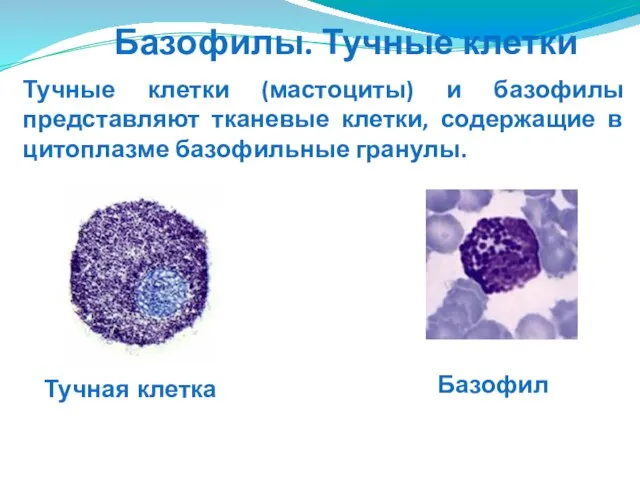
Тучные клетки (мастоциты) и базофилы представляют тканевые клетки, содержащие в цитоплазме
Тучные клетки (мастоциты) и базофилы представляют тканевые клетки, содержащие в цитоплазме
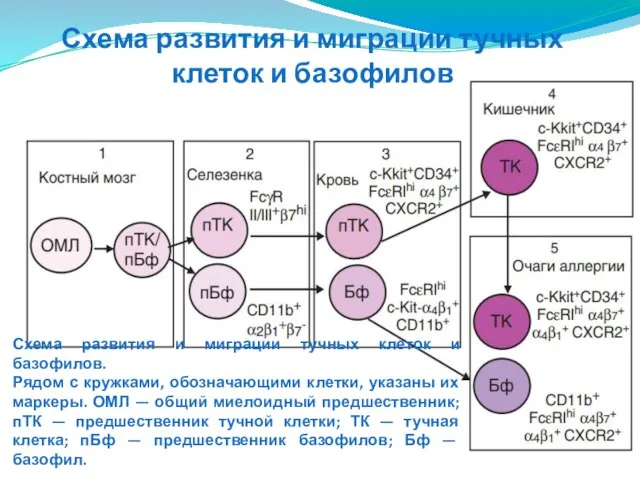
Схема развития и миграции тучных клеток и базофилов.
Рядом с кружками,
Схема развития и миграции тучных клеток и базофилов. Рядом с кружками,

Оба типа клеток имеют костномозговое происхождение и принадлежат к миелоидному ряду.
Оба типа клеток имеют костномозговое происхождение и принадлежат к миелоидному ряду.
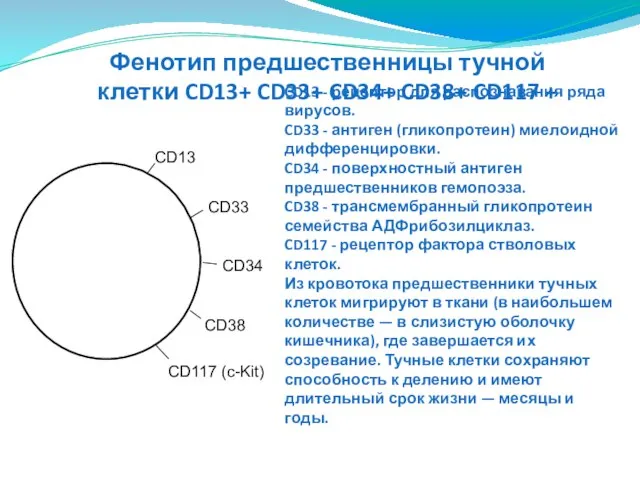
CD13 - рецептор для распознавания ряда вирусов.
CD33 - антиген (гликопротеин) миелоидной
CD13 - рецептор для распознавания ряда вирусов. CD33 - антиген (гликопротеин) миелоидной
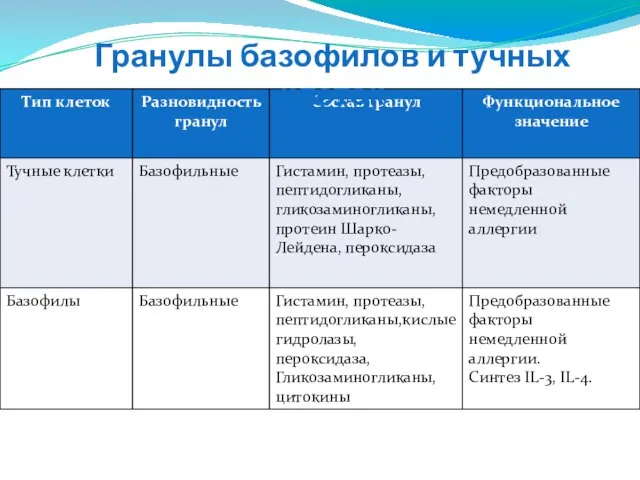
Гранулы базофилов и тучных клеток
Гранулы базофилов и тучных клеток
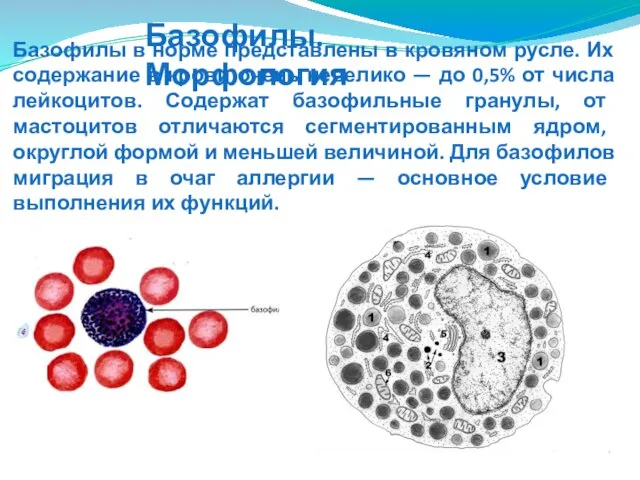
Базофилы в норме представлены в кровяном русле. Их содержание в крови
Базофилы в норме представлены в кровяном русле. Их содержание в крови
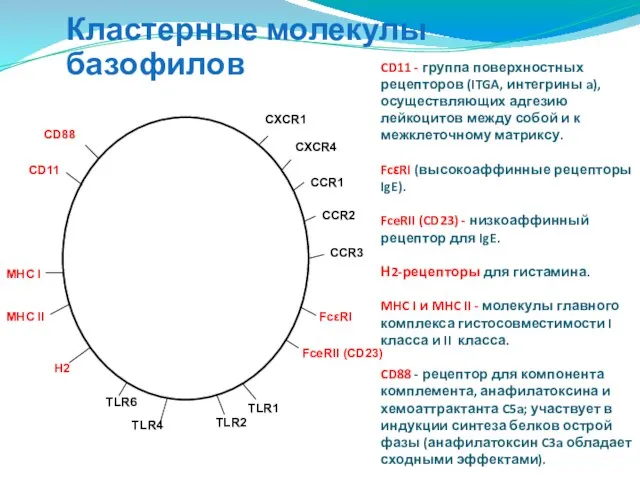
CD11 - группа поверхностных рецепторов (ITGA, интегрины a), осуществляющих адгезию лейкоцитов
CD11 - группа поверхностных рецепторов (ITGA, интегрины a), осуществляющих адгезию лейкоцитов

Спектр активных веществ, секретируемых базофилами, ограничен:
лейкотриен C4, IL-4, IL-13,
тромбоксаны
простогландины,
фактор
Спектр активных веществ, секретируемых базофилами, ограничен: лейкотриен C4, IL-4, IL-13, тромбоксаны простогландины, фактор
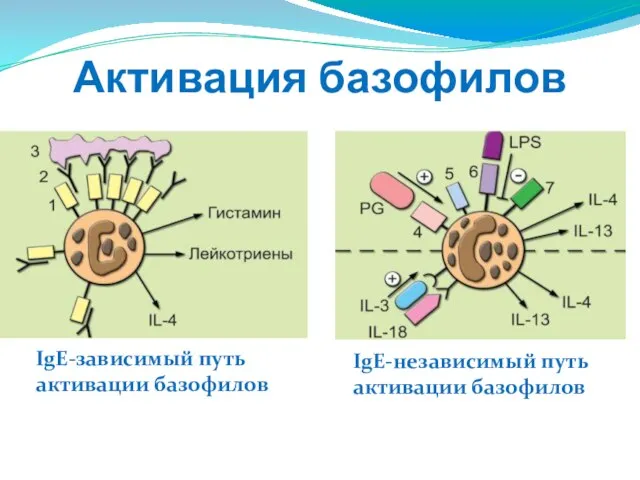
Активация базофилов
IgE-зависимый путь активации базофилов
IgE-независимый путь активации базофилов
Активация базофилов
IgE-зависимый путь активации базофилов
IgE-независимый путь активации базофилов

IgE-зависимый путь активации базофилов
Базофилы экспрессируют высокоаффинный рецептор FcsRI (1), который
IgE-зависимый путь активации базофилов
Базофилы экспрессируют высокоаффинный рецептор FcsRI (1), который

IgE-независимый путь активации базофила
Первый этап - индукция образования IL-4
IgE-независимый путь активации базофила Первый этап - индукция образования IL-4
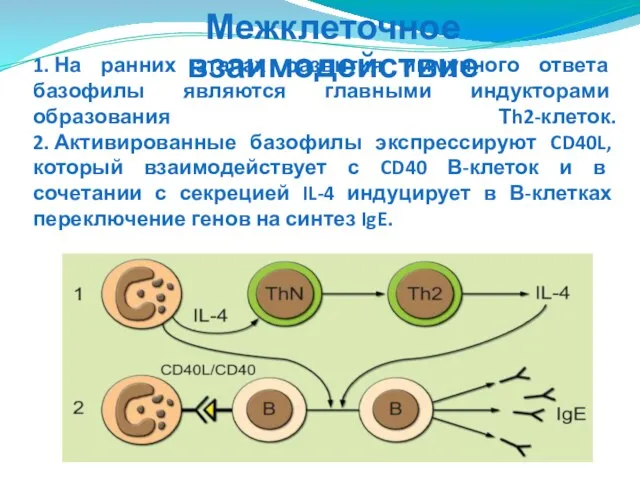
1. На ранних этапах развития иммунного ответа базофилы являются главными индукторами образования
1. На ранних этапах развития иммунного ответа базофилы являются главными индукторами образования

-регуляция процессов свертывания крови и проницаемости сосудов
-базофилы, как и эозинофилы, участвуют
-регуляция процессов свертывания крови и проницаемости сосудов -базофилы, как и эозинофилы, участвуют

1.Развитие базофилов и тучных клеток
2. Гранулы базофилов и тучных клеток, их
1.Развитие базофилов и тучных клеток
2. Гранулы базофилов и тучных клеток, их
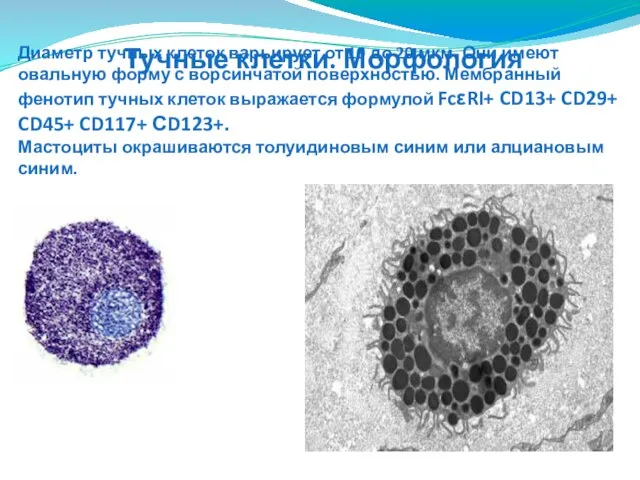
Диаметр тучных клеток варьирует от 10 до 20 мкм. Они имеют
Диаметр тучных клеток варьирует от 10 до 20 мкм. Они имеют
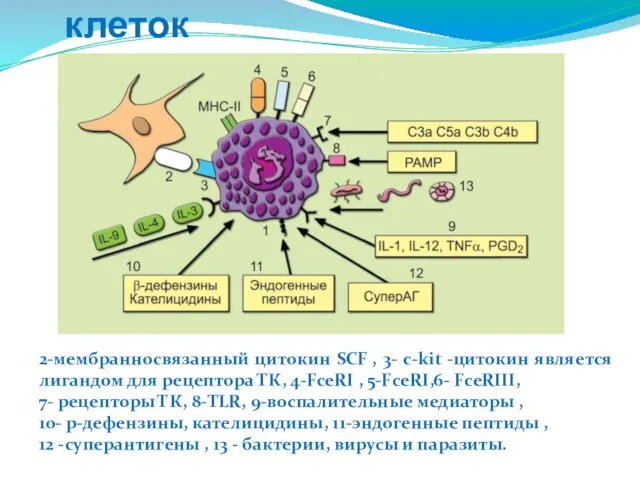
Характеристика тучных клеток
2-мембранносвязанный цитокин SCF , 3- c-kit -цитокин является лигандом для
Характеристика тучных клеток
2-мембранносвязанный цитокин SCF , 3- c-kit -цитокин является лигандом для
CD13 (GP150, поверхностный антиген лейкоцитов, аминопептидаза N-PEPN) - рецептор некоторых коронавирусов
CD13 (GP150, поверхностный антиген лейкоцитов, аминопептидаза N-PEPN) - рецептор некоторых коронавирусов
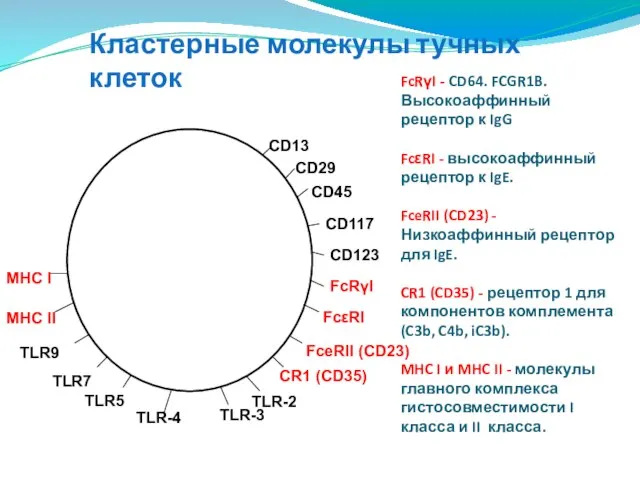
Кластерные молекулы тучных клеток
FcRγI - CD64. FCGR1B. Высокоаффинный рецептор к IgG
FcεRI
Кластерные молекулы тучных клеток
FcRγI - CD64. FCGR1B. Высокоаффинный рецептор к IgG FcεRI
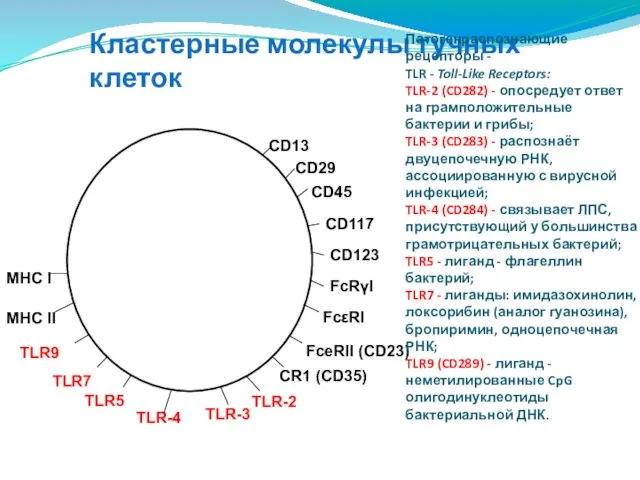
Кластерные молекулы тучных клеток
Патогенраспознающие рецепторы -
TLR - Toll-Like Receptors:
TLR-2 (CD282) -
Кластерные молекулы тучных клеток
Патогенраспознающие рецепторы - TLR - Toll-Like Receptors: TLR-2 (CD282) -
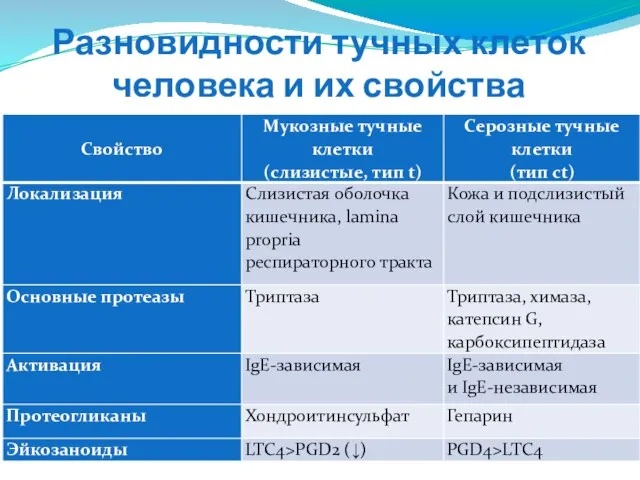
Разновидности тучных клеток человека и их свойства
Разновидности тучных клеток человека и их свойства
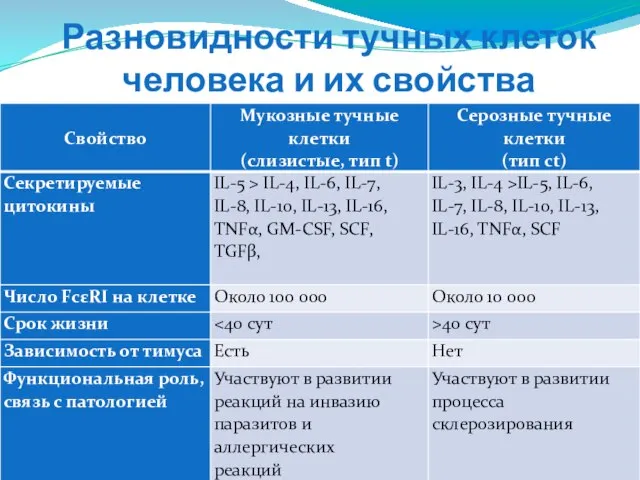
Разновидности тучных клеток человека и их свойства
Разновидности тучных клеток человека и их свойства
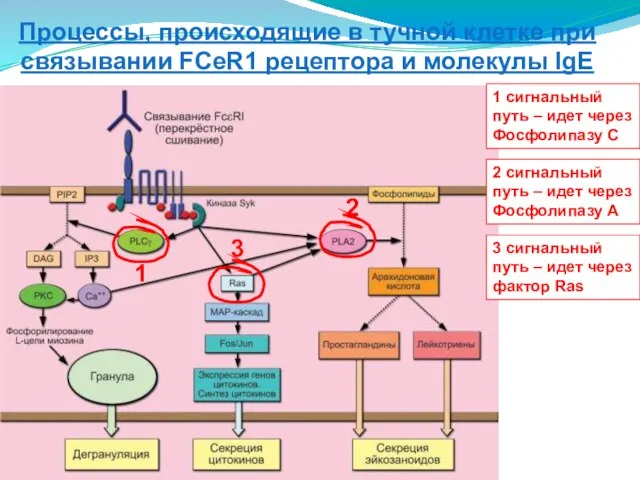
1
2
3
1 сигнальный путь – идет через Фосфолипазу С
2 сигнальный путь –
1
2
3
1 сигнальный путь – идет через Фосфолипазу С
2 сигнальный путь –
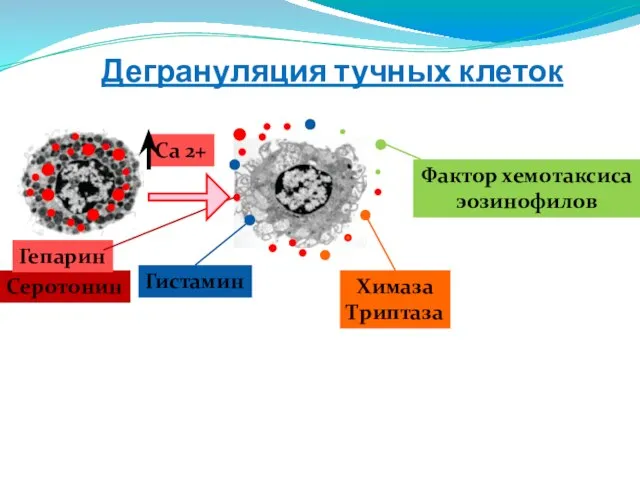
Дегрануляция тучных клеток
Гистамин
Химаза
Триптаза
Са 2+
Серотонин
Гепарин
Фактор хемотаксиса эозинофилов
Дегрануляция тучных клеток
Гистамин
Химаза
Триптаза
Са 2+
Серотонин
Гепарин
Фактор хемотаксиса эозинофилов
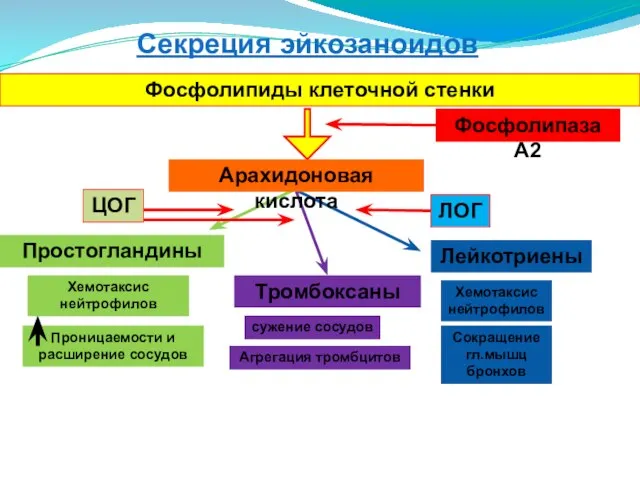
Секреция эйкозаноидов
Фосфолипиды клеточной стенки
Фосфолипаза А2
Простогландины
Лейкотриены
Арахидоновая кислота
ЦОГ
ЛОГ
Тромбоксаны
Проницаемости и расширение сосудов
Хемотаксис нейтрофилов
Хемотаксис нейтрофилов
Агрегация
Секреция эйкозаноидов
Фосфолипиды клеточной стенки
Фосфолипаза А2
Простогландины
Лейкотриены
Арахидоновая кислота
ЦОГ
ЛОГ
Тромбоксаны
Проницаемости и расширение сосудов
Хемотаксис нейтрофилов
Хемотаксис нейтрофилов
Агрегация

1) Участие в реакциях воспаления (гистамин, гепарин): повышение проницаемости сосудов, образование
1) Участие в реакциях воспаления (гистамин, гепарин): повышение проницаемости сосудов, образование
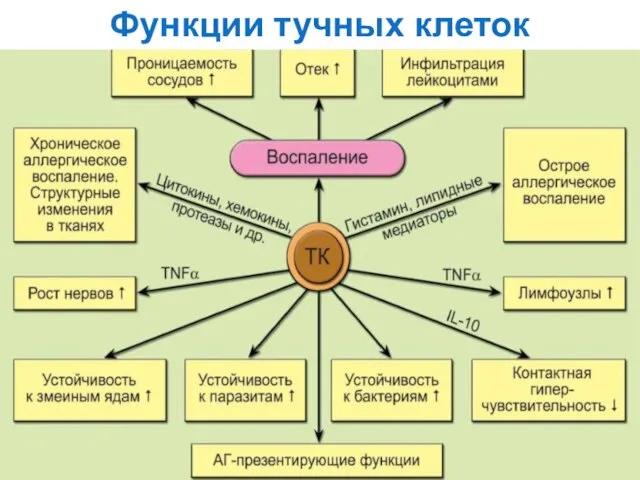
Функции тучных клеток
Функции тучных клеток

Дендритные клетки (ДК)
Это специализированные антигенпрезентирующие лейкоциты, способные активировать как
Дендритные клетки (ДК)
Это специализированные антигенпрезентирующие лейкоциты, способные активировать как
Места локализации
Основные места локализации: барьерные органы, слизистые оболочки и регионарные лимфатические
Места локализации
Основные места локализации: барьерные органы, слизистые оболочки и регионарные лимфатические
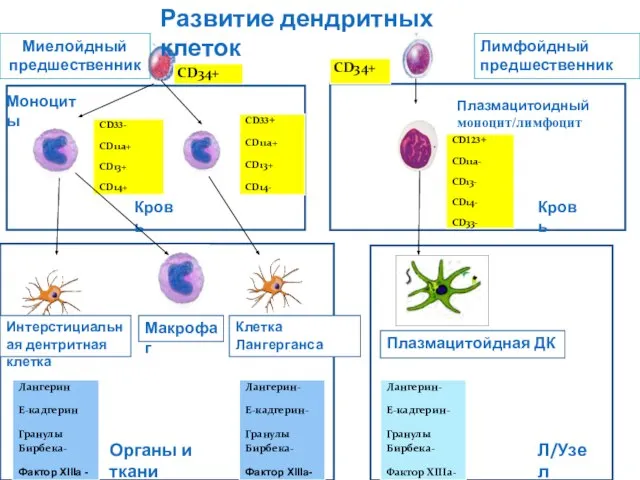
Макрофаг
Лимфойдный предшественник
Моноциты
Плазмацитоидный моноцит/лимфоцит
Кровь
Кровь
Интерстициальная дентритная клетка
Макрофаг
Клетка Лангерганса
Плазмацитойдная ДК
Органы и ткани
Л/Узел
Миелойдный предшественник
Развитие
Макрофаг
Лимфойдный предшественник
Моноциты
Плазмацитоидный моноцит/лимфоцит
Кровь
Кровь
Интерстициальная дентритная клетка
Макрофаг
Клетка Лангерганса
Плазмацитойдная ДК
Органы и ткани
Л/Узел
Миелойдный предшественник
Развитие

Миелоидные ДК и макрофаги
Миелоидные ДК и макрофаги имеют общего костномозгового предшественника,
Миелоидные ДК и макрофаги
Миелоидные ДК и макрофаги имеют общего костномозгового предшественника,

Плазмацитоидные ДК
Плазмацитоидные ДК также происходят из CD34 позитивного предшественника. Внешне походи
Плазмацитоидные ДК
Плазмацитоидные ДК также происходят из CD34 позитивного предшественника. Внешне походи
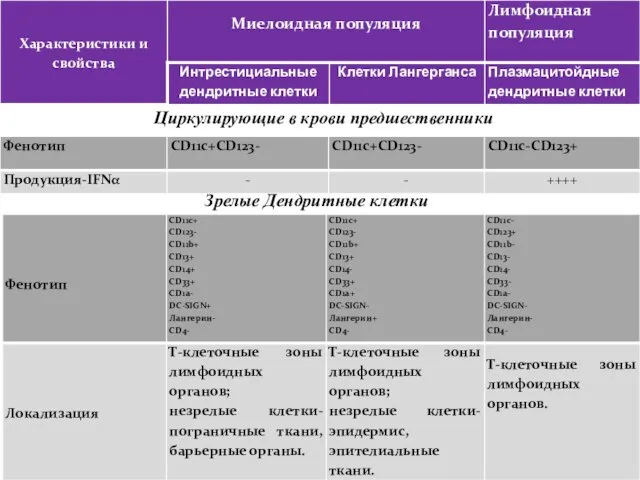
Циркулирующие в крови предшественники
Зрелые Дендритные клетки
Циркулирующие в крови предшественники
Зрелые Дендритные клетки
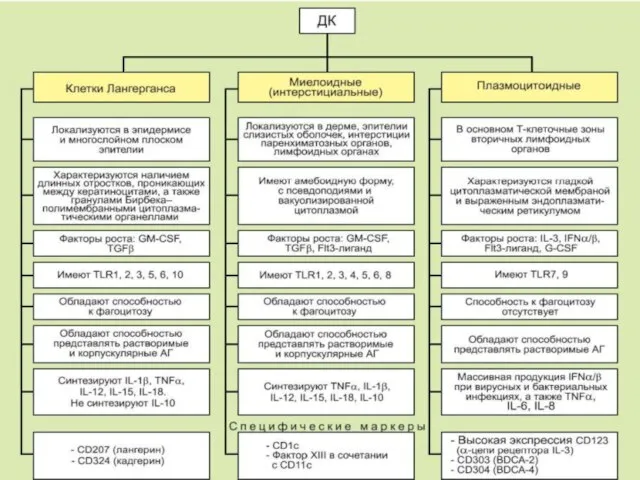
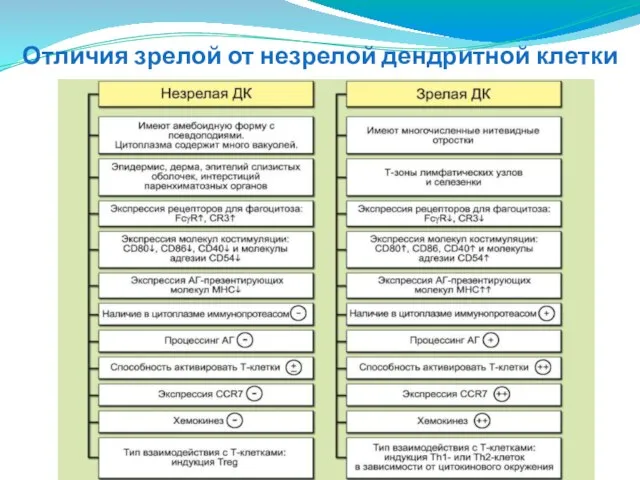
Отличия зрелой от незрелой дендритной клетки
Отличия зрелой от незрелой дендритной клетки
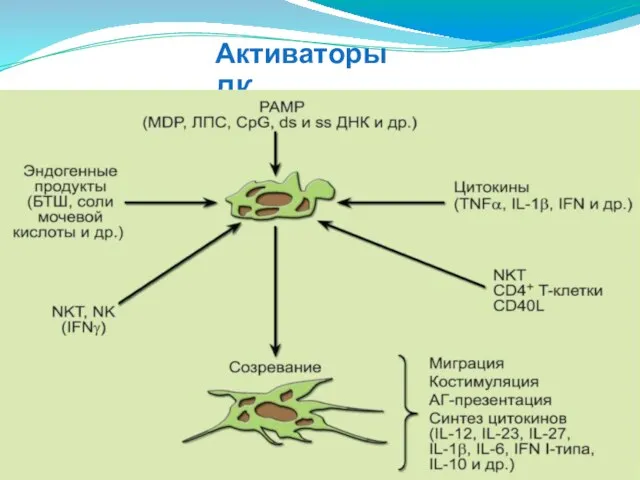
Активаторы ДК
Активаторы ДК

Миграция ДК
В зависимости от степени созревания, ДК клетки будут проявлять различную
Миграция ДК
В зависимости от степени созревания, ДК клетки будут проявлять различную
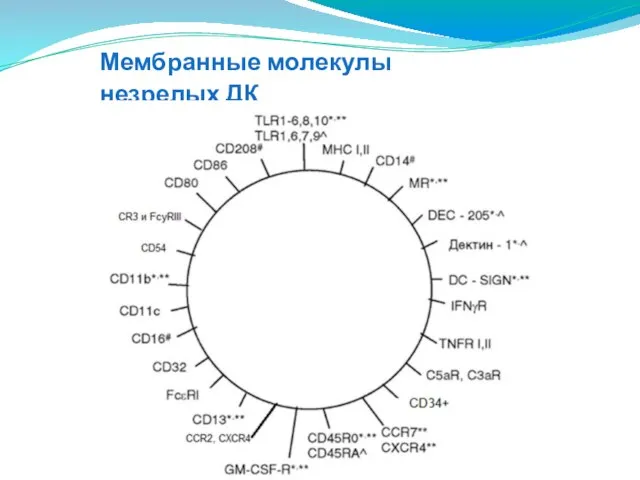
Мембранные молекулы незрелых ДК
Мембранные молекулы незрелых ДК
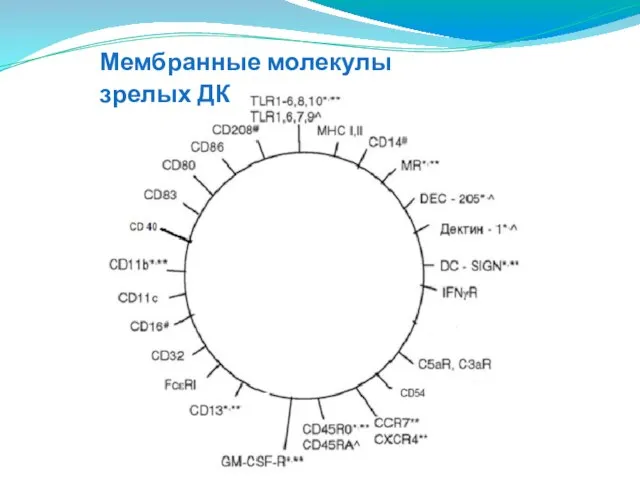
Мембранные молекулы зрелых ДК
Мембранные молекулы зрелых ДК
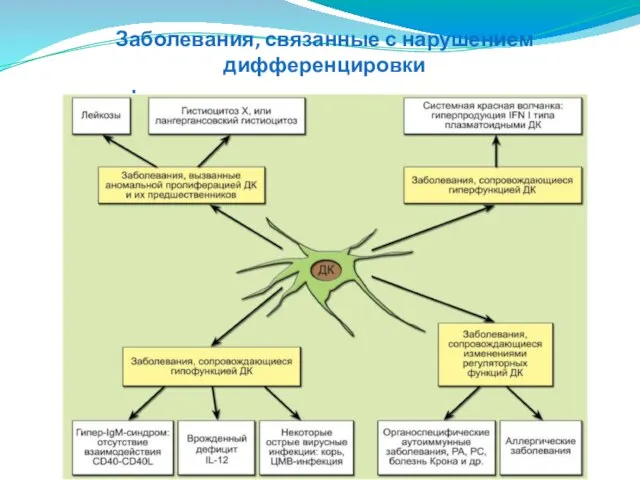
Заболевания, связанные с нарушением дифференцировки
и функционирования дендритных клеток
Заболевания, связанные с нарушением дифференцировки
и функционирования дендритных клеток

NK-клетки
1.Развитие NK-клеток
2.Морфология NK-клеток
3.Гранулы NK-клеток
4.Кластерные молекулы NK-клеток
5.Функции NK-клеток
6. Эффекторные функции NK-клеток.
а)Экзоцитоз гранул
б)Включение
NK-клетки
1.Развитие NK-клеток
2.Морфология NK-клеток
3.Гранулы NK-клеток
4.Кластерные молекулы NK-клеток
5.Функции NK-клеток
6. Эффекторные функции NK-клеток.
а)Экзоцитоз гранул
б)Включение
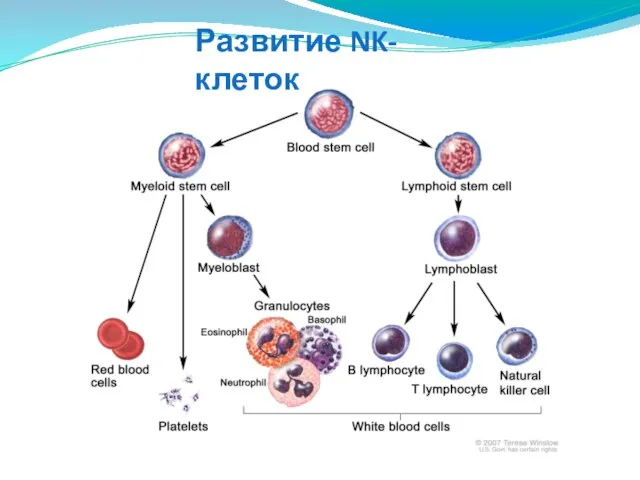
Развитие NK-клеток
Развитие NK-клеток
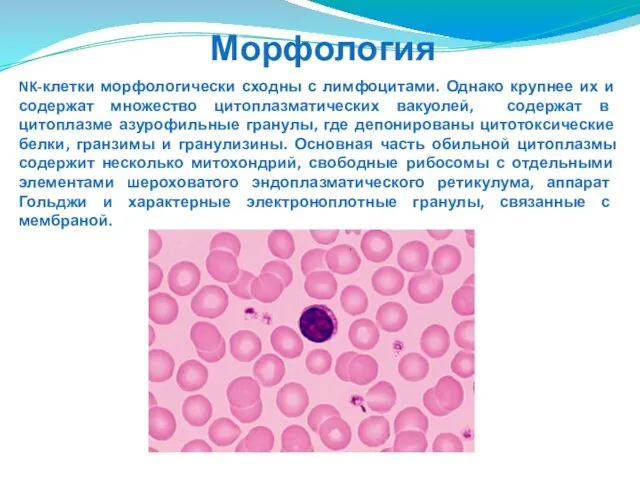
NK-клетки морфологически сходны с лимфоцитами. Однако крупнее их и содержат множество
NK-клетки морфологически сходны с лимфоцитами. Однако крупнее их и содержат множество

Гранулы
Перфорин представляет собой белок с высокой молекулярной массой. На поверхности клетки
Гранулы
Перфорин представляет собой белок с высокой молекулярной массой. На поверхности клетки

Гранулы
Гранулизин - апсониноподобный положительно заряженный белок с молекулярной массой 9 kDa.
Гранулы
Гранулизин - апсониноподобный положительно заряженный белок с молекулярной массой 9 kDa.

Гранулы
Гранзимы - протеазы химотрипсинового типа основной эффект гранзимов В активация протеазы
Гранулы
Гранзимы - протеазы химотрипсинового типа основной эффект гранзимов В активация протеазы
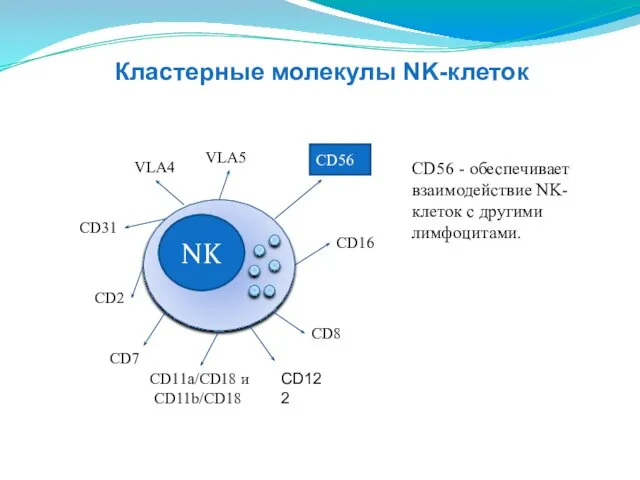
NK
CD16
CD8
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
СD56 - обеспечивает взаимодействие NK-клеток с другими лимфоцитами.
CD7
CD122
Кластерные молекулы
NK
CD16
CD8
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
СD56 - обеспечивает взаимодействие NK-клеток с другими лимфоцитами.
CD7
CD122
Кластерные молекулы
NK
CD8
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
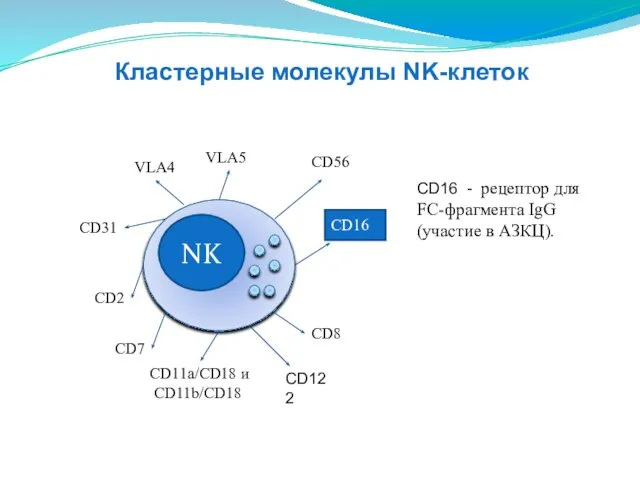
CD16
CD16 - рецептор для FC-фрагмента IgG (участие в АЗКЦ).
CD7
CD122
Кластерные
NK
CD8
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
CD16
CD16 - рецептор для FC-фрагмента IgG (участие в АЗКЦ).
CD7
CD122
Кластерные
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
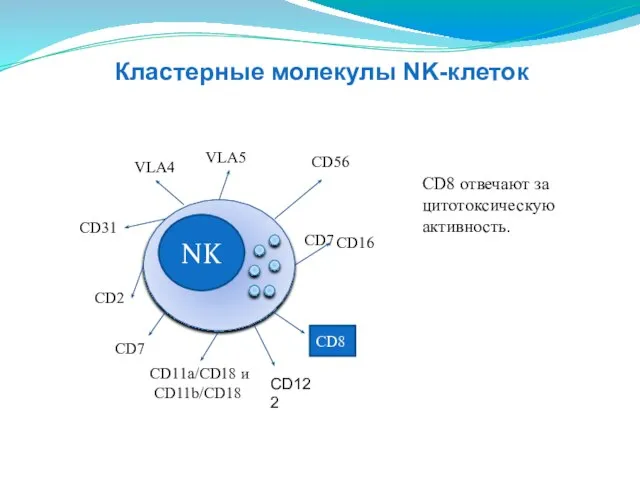
CD8
CD8 отвечают за цитотоксическую активность.
CD7
CD7
CD122
Кластерные молекулы NK-клеток
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
CD8
CD8 отвечают за цитотоксическую активность.
CD7
CD7
CD122
Кластерные молекулы NK-клеток
NK
CD16
CD2
СD31
VLA4
VLA5
СD56
CD8
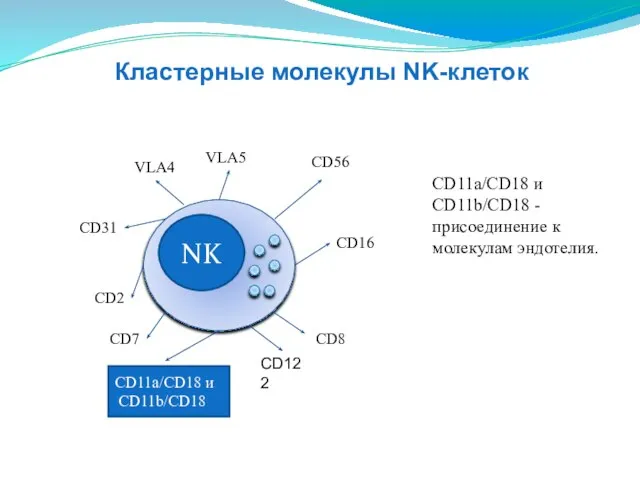
CD11a/CD18 и
CD11b/CD18
CD11a/CD18 и CD11b/CD18 - присоединение к молекулам эндотелия.
CD7
CD122
Кластерные молекулы
NK
CD16
CD2
СD31
VLA4
VLA5
СD56
CD8
CD11a/CD18 и
CD11b/CD18
CD11a/CD18 и CD11b/CD18 - присоединение к молекулам эндотелия.
CD7
CD122
Кластерные молекулы
NK
CD16
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
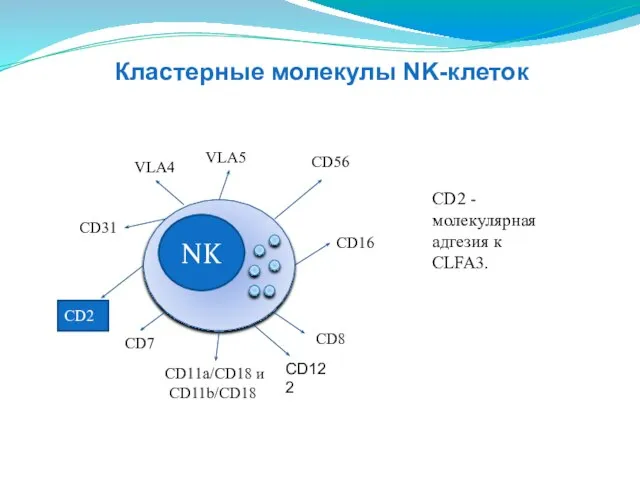
CD8
CD2
CD2 - молекулярная адгезия к CLFA3.
CD7
CD122
Кластерные молекулы NK-клеток
NK
CD16
CD11a/CD18 и
CD11b/CD18
СD31
VLA4
VLA5
СD56
CD8
CD2
CD2 - молекулярная адгезия к CLFA3.
CD7
CD122
Кластерные молекулы NK-клеток
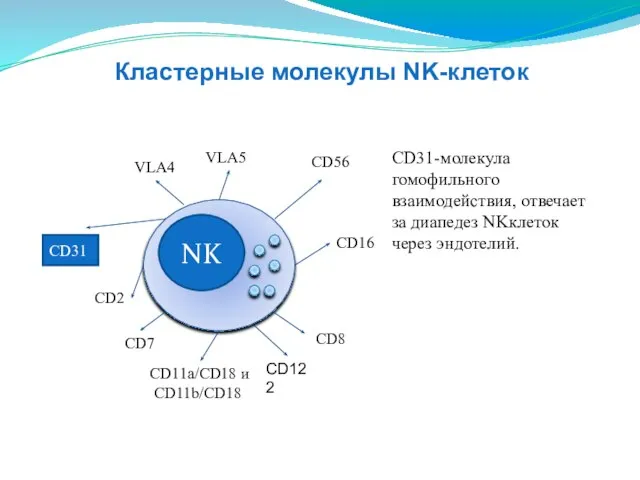
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
VLA4
VLA5
СD56
CD8
СD31
СD31-молекула гомофильного взаимодействия, отвечает за диапедез NKклеток через эндотелий.
CD7
CD122
Кластерные
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
VLA4
VLA5
СD56
CD8
СD31
СD31-молекула гомофильного взаимодействия, отвечает за диапедез NKклеток через эндотелий.
CD7
CD122
Кластерные
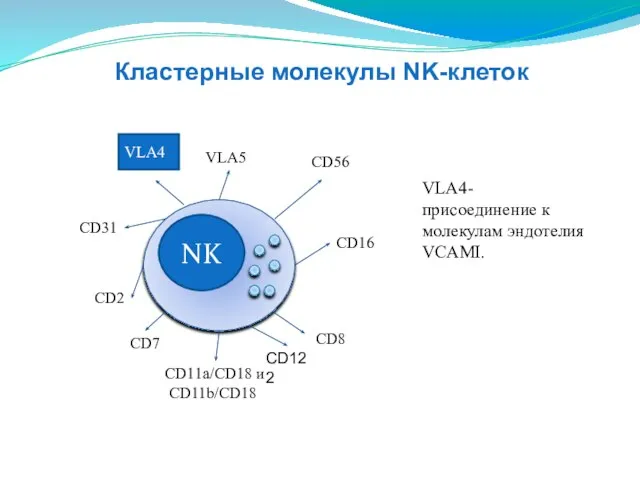
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA5
СD56
CD8
VLA4
VLA4-присоединение к молекулам эндотелия VCAMI.
CD7
CD122
Кластерные молекулы NK-клеток
NK
CD16
CD2
CD11a/CD18 и
CD11b/CD18
СD31
VLA5
СD56
CD8
VLA4
VLA4-присоединение к молекулам эндотелия VCAMI.
CD7
CD122
Кластерные молекулы NK-клеток
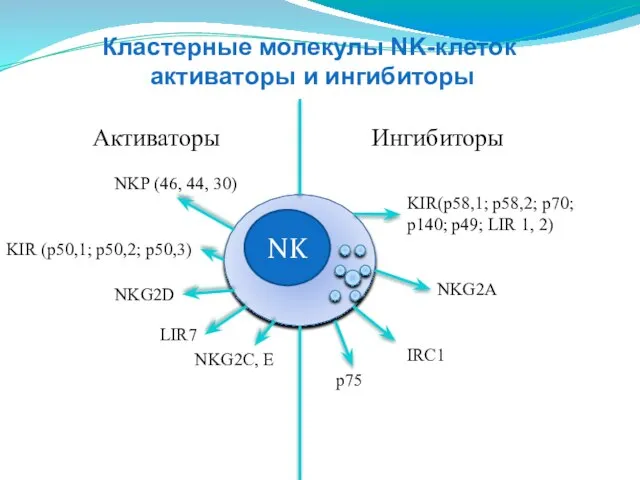
NK
Активаторы
Ингибиторы
NKP (46, 44, 30)
KIR (p50,1; p50,2; p50,3)
NKG2D
NKG2C, E
KIR(p58,1; p58,2; p70; p140;
NK
Активаторы
Ингибиторы
NKP (46, 44, 30)
KIR (p50,1; p50,2; p50,3)
NKG2D
NKG2C, E
KIR(p58,1; p58,2; p70; p140;

Регуляция процессов пролиферации, дифференцировки и элиминации стареющих соматических клеток организма
Модуляция клеток
Регуляция процессов пролиферации, дифференцировки и элиминации стареющих соматических клеток организма
Модуляция клеток

Созревание предшественников CTL
Генерация вирус-специфических CTL
Созревание претимоцитов и тимоцитов
Функции NK-клеток
Созревание предшественников CTL
Генерация вирус-специфических CTL
Созревание претимоцитов и тимоцитов
Функции NK-клеток
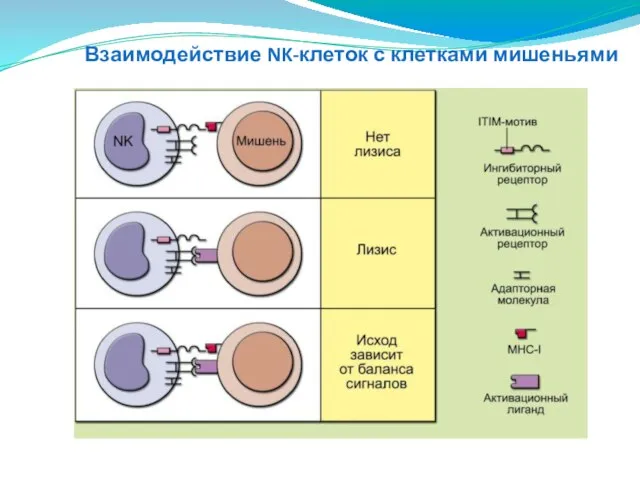
Взаимодействие NK-клеток с клетками мишеньями
Взаимодействие NK-клеток с клетками мишеньями
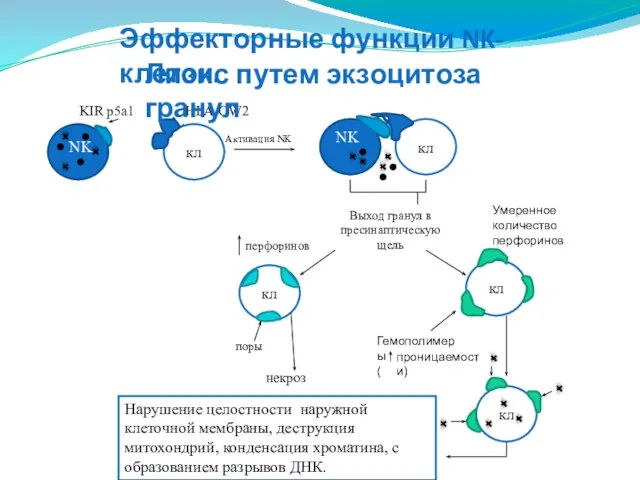
NK
KIR p5a1
кл
HLA-CW2
Активация NK
кл
NK
Выход гранул в пресинаптическую щель
кл
поры
некроз
перфоринов
кл
Умеренное количество перфоринов
Гемополимеры
(
проницаемости)
кл
Нарушение целостности
NK
KIR p5a1
кл
HLA-CW2
Активация NK
кл
NK
Выход гранул в пресинаптическую щель
кл
поры
некроз
перфоринов
кл
Умеренное количество перфоринов
Гемополимеры
(
проницаемости)
кл
Нарушение целостности
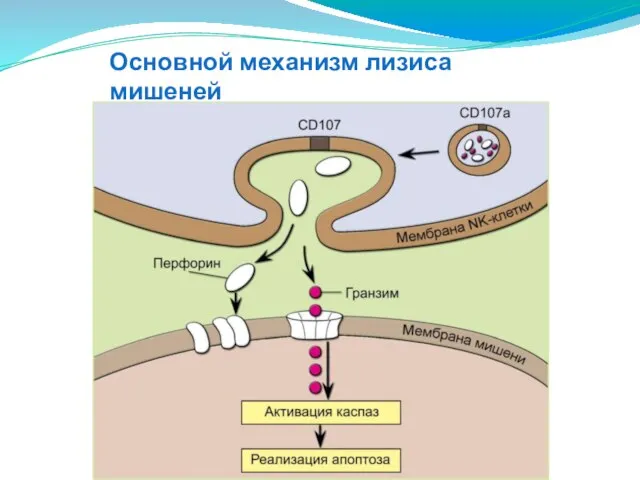
Основной механизм лизиса мишеней
Основной механизм лизиса мишеней
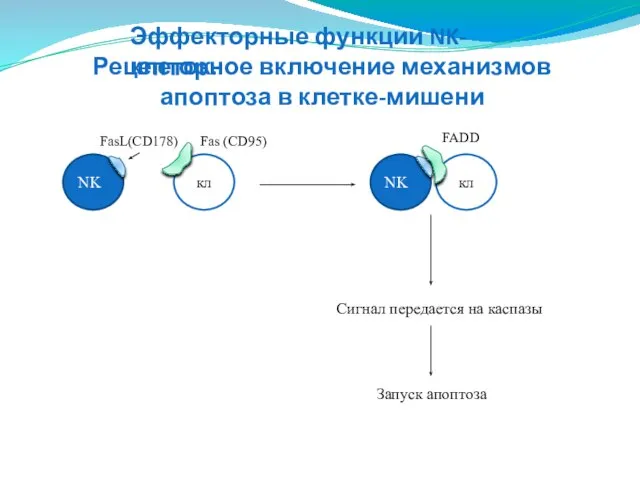
NK
кл
FasL(CD178)
Fas (CD95)
NK
кл
FADD
Сигнал передается на каспазы
Запуск апоптоза
Рецепторное включение механизмов апоптоза в
NK
кл
FasL(CD178)
Fas (CD95)
NK
кл
FADD
Сигнал передается на каспазы
Запуск апоптоза
Рецепторное включение механизмов апоптоза в
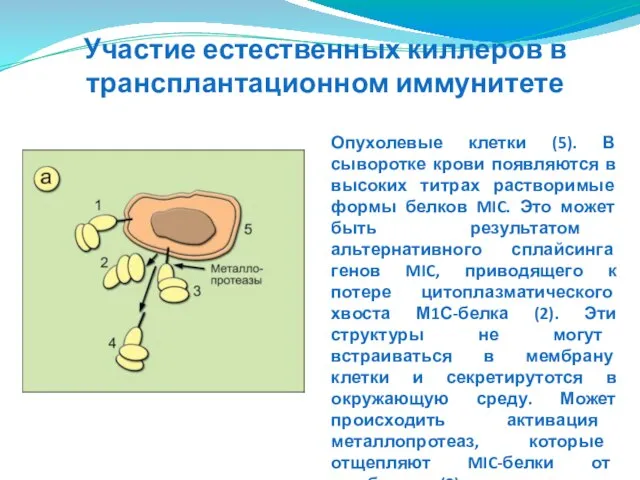
Опухолевые клетки (5). В сыворотке крови появляются в высоких титрах растворимые
Опухолевые клетки (5). В сыворотке крови появляются в высоких титрах растворимые
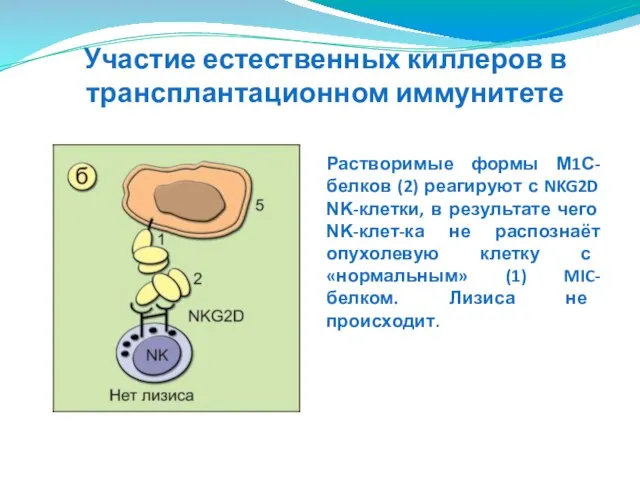
Растворимые формы М1С-белков (2) реагируют с NKG2D ΝΚ-клетки, в результате чего
Растворимые формы М1С-белков (2) реагируют с NKG2D ΝΚ-клетки, в результате чего
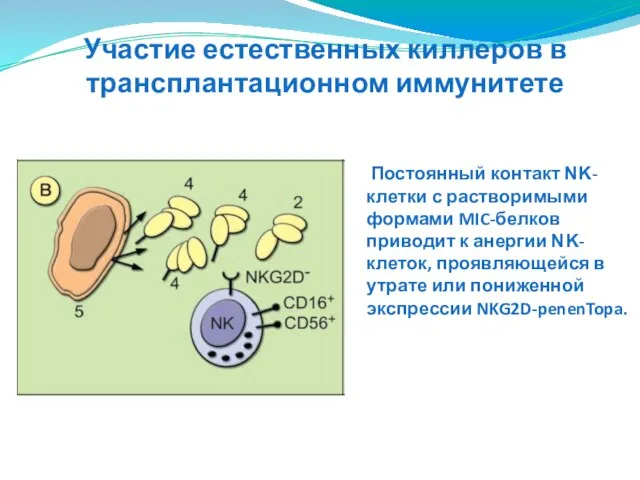
Постоянный контакт ΝΚ-клетки с растворимыми формами MIC-белков приводит к анергии ΝΚ-клеток,
Постоянный контакт ΝΚ-клетки с растворимыми формами MIC-белков приводит к анергии ΝΚ-клеток,
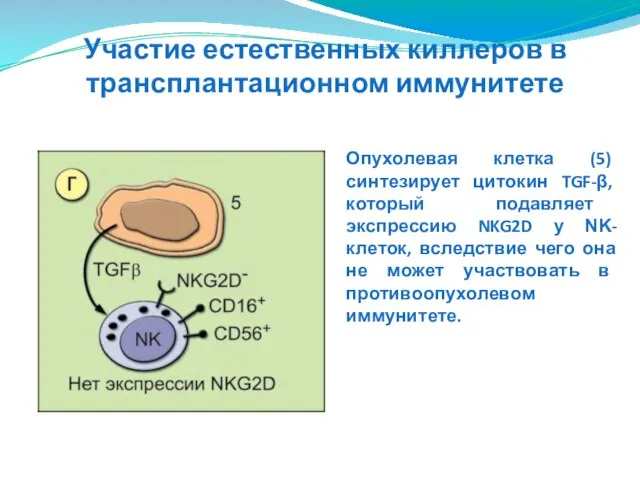
Опухолевая клетка (5) синтезирует цитокин TGF-β, который подавляет экспрессию NKG2D у
Опухолевая клетка (5) синтезирует цитокин TGF-β, который подавляет экспрессию NKG2D у
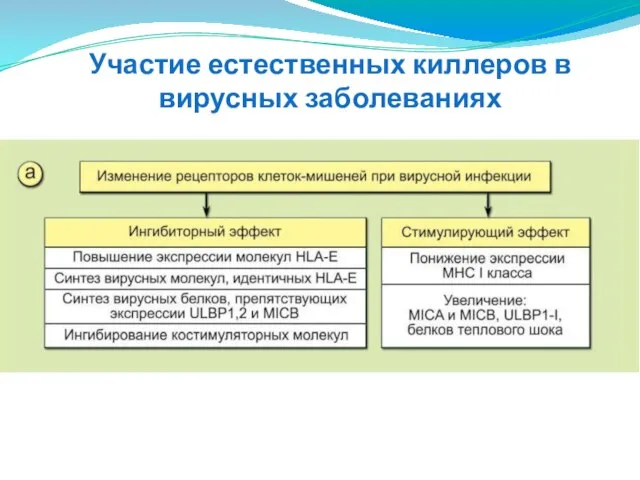
Участие естественных киллеров в вирусных заболеваниях
Участие естественных киллеров в вирусных заболеваниях
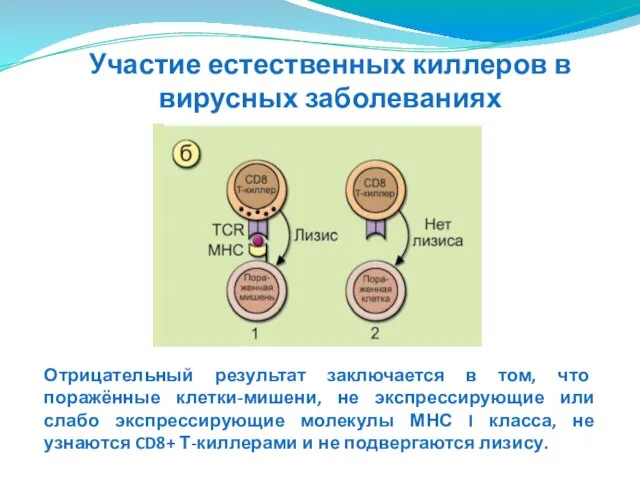
Отрицательный результат заключается в том, что поражённые клетки-мишени, не экспрессирующие или
Отрицательный результат заключается в том, что поражённые клетки-мишени, не экспрессирующие или
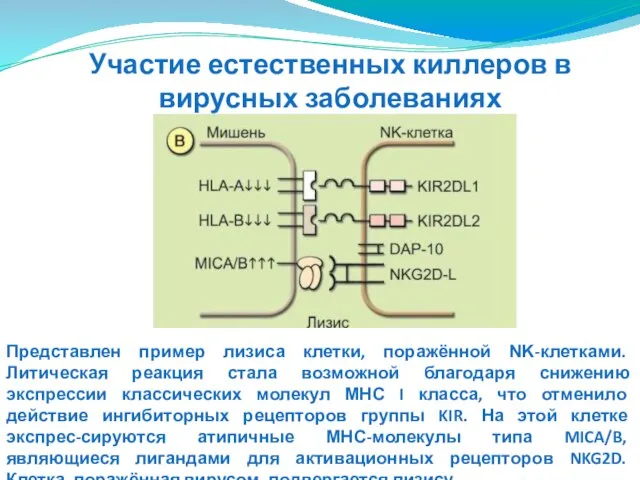
Представлен пример лизиса клетки, поражённой ΝΚ-клетками. Литическая реакция стала возможной благодаря
Представлен пример лизиса клетки, поражённой ΝΚ-клетками. Литическая реакция стала возможной благодаря

NK-клетки выступают в роли в роли LGL клеток. Их функции:
Регулируют
NK-клетки выступают в роли в роли LGL клеток. Их функции:
Регулируют

ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА

ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА

ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
Гуморальная составляющая врожденного иммунитета представлена несколькими взаимосвязанными системами –
ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО
ИММУНИТЕТА
Гуморальная составляющая врожденного иммунитета представлена несколькими взаимосвязанными системами –

Компоненты системы комплемента
Компоненты системы комплемента

Система комплемента является важным компонентом врождённого иммунитета, играющим большую роль в
Система комплемента является важным компонентом врождённого иммунитета, играющим большую роль в
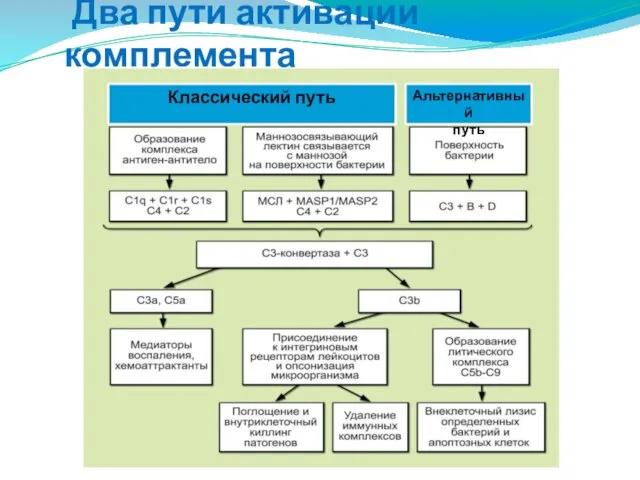
Два пути активации комплемента
Классический путь
Альтернативный
путь
Два пути активации комплемента
Классический путь
Альтернативный
путь
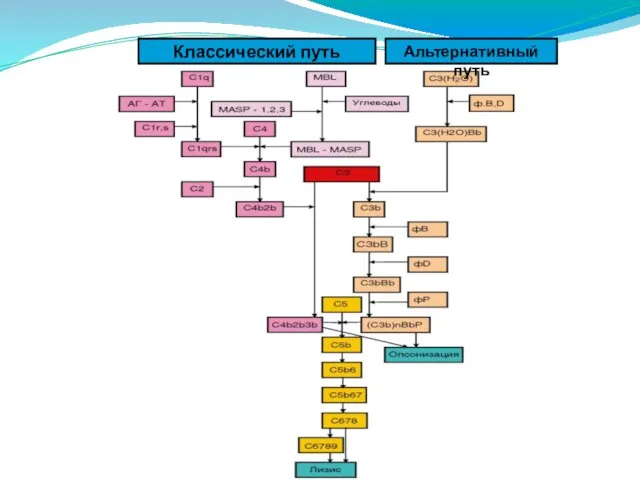
Классический путь
Альтернативный путь
Классический путь
Альтернативный путь
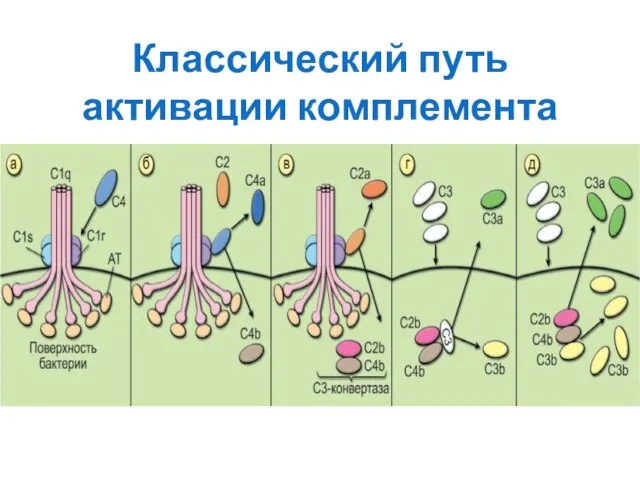
Классический путь активации комплемента
Классический путь активации комплемента

А) После присоединения AT класса IgM или некоторых изотипов IgG к поверхности бактериальной клетки у
А) После присоединения AT класса IgM или некоторых изотипов IgG к поверхности бактериальной клетки у
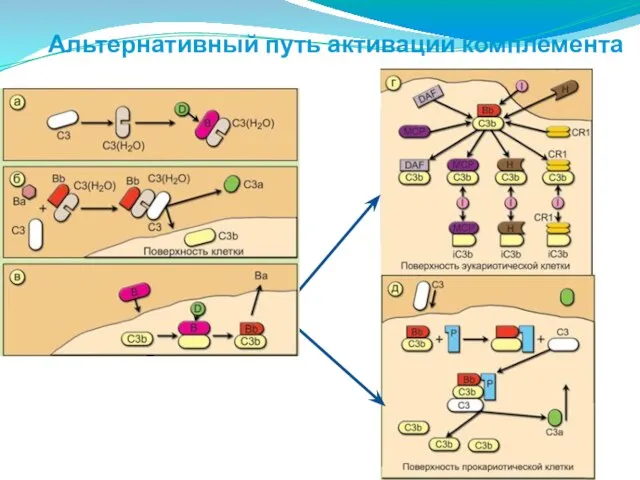
Альтернативный путь активации комплемента
Альтернативный путь активации комплемента

Альтернативный путь происходит в отсутствие AT на поверхности микробной клетки, приводя
Альтернативный путь происходит в отсутствие AT на поверхности микробной клетки, приводя

В сыворотке крови присутствует сложный белок, относящийся к семейству коллектинов, - маннозасвязывающий лектин
В сыворотке крови присутствует сложный белок, относящийся к семейству коллектинов, - маннозасвязывающий лектин

Белки острой фазы воспаления
Белки (реактанты) острой фазы представляют группу протеинов,
Белки острой фазы воспаления
Белки (реактанты) острой фазы представляют группу протеинов,
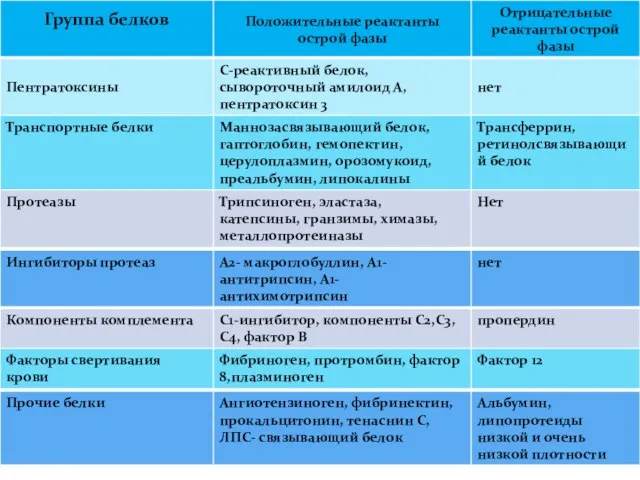

Пентраксины
Наиболее полно проявляют свойства реактантов острой фазы белки семейства пентраксинов:
Пентраксины
Наиболее полно проявляют свойства реактантов острой фазы белки семейства пентраксинов:

Пентраксины
Короткие петраксины
Длинные пентраксины
С-реактивный белок
Сывороточный амилоид Р
РТХ3 (пентраксин
Пентраксины
Короткие петраксины
Длинные пентраксины
С-реактивный белок
Сывороточный амилоид Р
РТХ3 (пентраксин
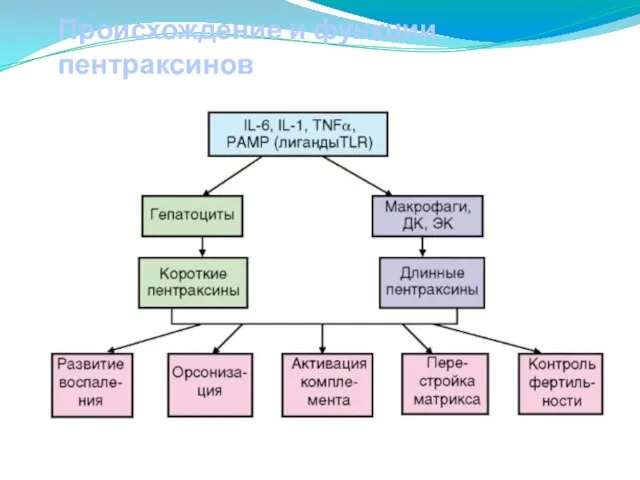
Происхождение и функции пентраксинов
Происхождение и функции пентраксинов

Распознавание и связывание РАМР (растворимые патогенраспознающие рецепторы )
Комплемент-активирующая(через Clq) и опсонизирующая
Распознавание и связывание РАМР (растворимые патогенраспознающие рецепторы )
Комплемент-активирующая(через Clq) и опсонизирующая
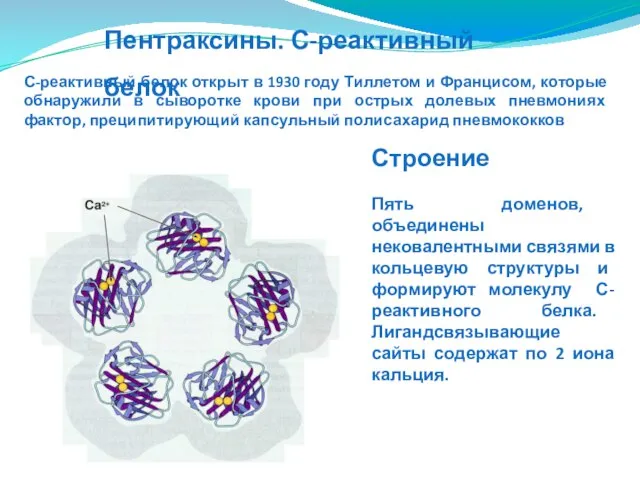
Пентраксины. С-реактивный белок
Строение
Пять доменов, объединены нековалентными связями в кольцевую структуры
Пентраксины. С-реактивный белок
Строение
Пять доменов, объединены нековалентными связями в кольцевую структуры

является предшественником Р-компонента амилоидных фибрилл - постоянной составной части амилоидных депозитов
является предшественником Р-компонента амилоидных фибрилл - постоянной составной части амилоидных депозитов

Они играют роль переносчиков метаболитов, ионов металлов, физиологически активных факторов.
К
Они играют роль переносчиков метаболитов, ионов металлов, физиологически активных факторов.
К

Протеазы и антипротеазы
Другую группу образуют протеазы, активация которых необходима для формирования
Протеазы и антипротеазы
Другую группу образуют протеазы, активация которых необходима для формирования

Цитокины
Цитокины –это белковые или полипептидные факторы, лишенные специфичности в отношении
Цитокины
Цитокины –это белковые или полипептидные факторы, лишенные специфичности в отношении

Интерфероны
Общее свойство интерферонов - наличие у них противовирусной активности. В то
Интерфероны
Общее свойство интерферонов - наличие у них противовирусной активности. В то

Функции основных интерферонов
Функции основных интерферонов

Приобретенный иммунитет
Приобретенный иммунитет
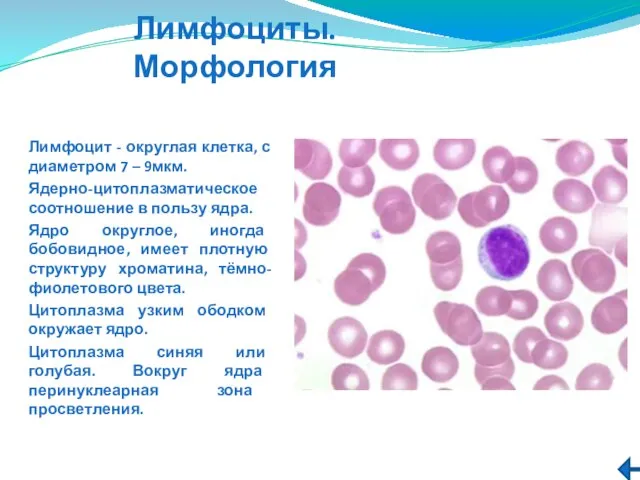
Лимфоциты.Морфология
Лимфоцит - округлая клетка, с диаметром 7 – 9мкм.
Ядерно-цитоплазматическое соотношение
Лимфоциты.Морфология
Лимфоцит - округлая клетка, с диаметром 7 – 9мкм.
Ядерно-цитоплазматическое соотношение
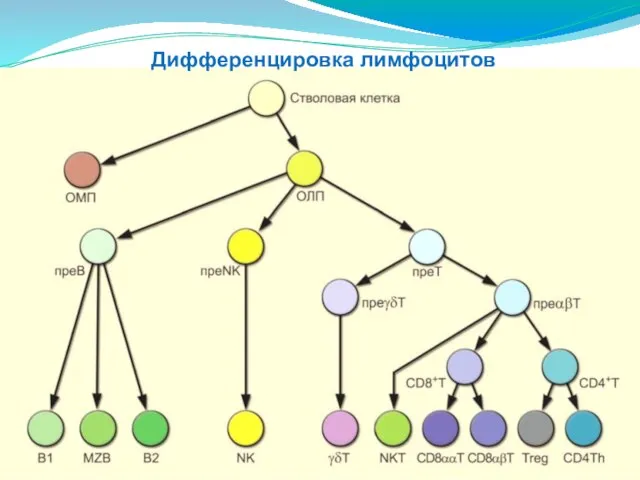
Дифференцировка лимфоцитов
Дифференцировка лимфоцитов
Органы иммунной системы
Центральные
Периферические
Костный мозг
Тимус
Селезенка
Аппендикс
Пееровые бляшки
Печень
Лимфоузлы
Органы иммунной системы
Центральные
Периферические
Костный мозг
Тимус
Селезенка
Аппендикс
Пееровые бляшки
Печень
Лимфоузлы
Органы иммунной системы
Неинкапсулированные
MALT (Лимфоидная ткань слизистых оболочек )
GALT (с пищеварительным трактом)
Селезенка
Тимус
Лимфоузлы
Инкапсулированные
BALT
Органы иммунной системы
Неинкапсулированные
MALT (Лимфоидная ткань слизистых оболочек )
GALT (с пищеварительным трактом)
Селезенка
Тимус
Лимфоузлы
Инкапсулированные
BALT
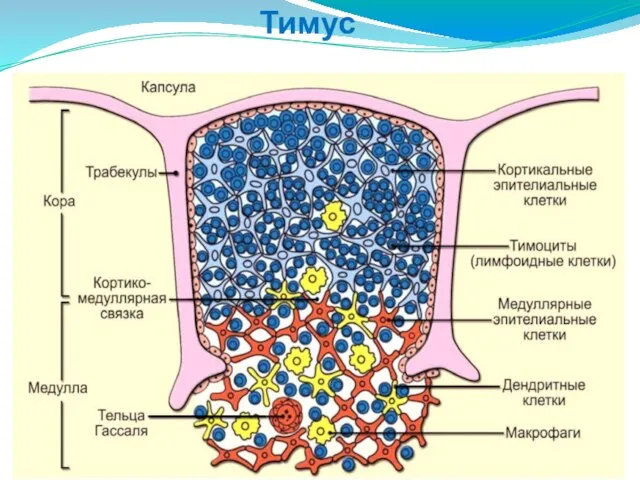
Тимус (вилочковая железа)
Тимус расположен за рукояткой грудины. У человека и большинства
Тимус (вилочковая железа)
Тимус расположен за рукояткой грудины. У человека и большинства
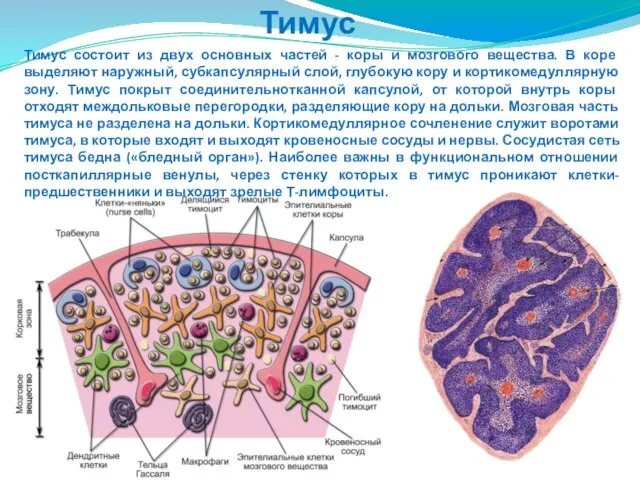
Тимус
Тимус состоит из двух основных частей - коры и мозгового вещества.
Тимус
Тимус состоит из двух основных частей - коры и мозгового вещества.
Тимус
Тимус

1 - корковое вещество;
2 - мозговое вещество;
3 - капсула;
1 - корковое вещество;
2 - мозговое вещество;
3 - капсула;
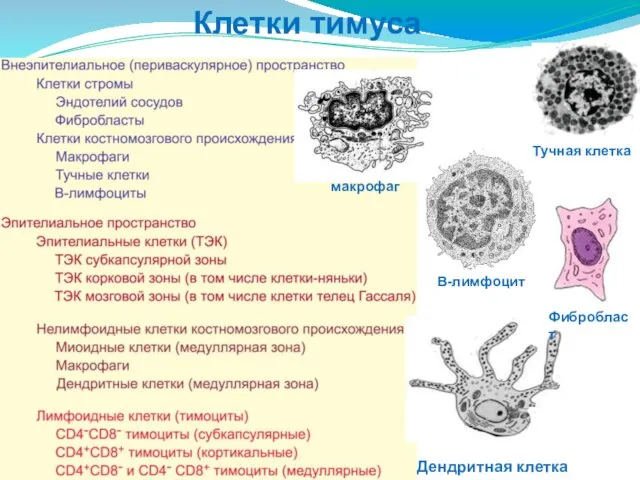
Клетки тимуса
макрофаг
В-лимфоцит
Тучная клетка
Дендритная клетка
Фибробласт
Клетки тимуса
макрофаг
В-лимфоцит
Тучная клетка
Дендритная клетка
Фибробласт
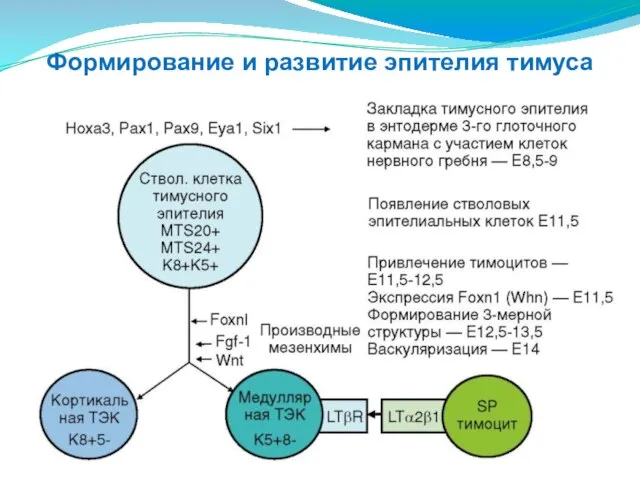
Формирование и развитие эпителия тимуса
Формирование и развитие эпителия тимуса
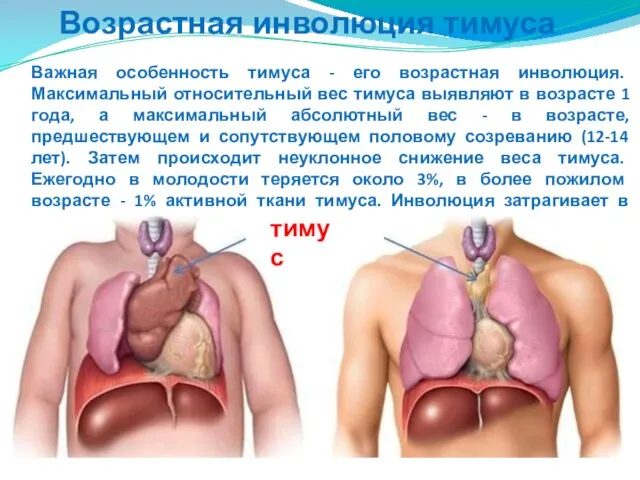
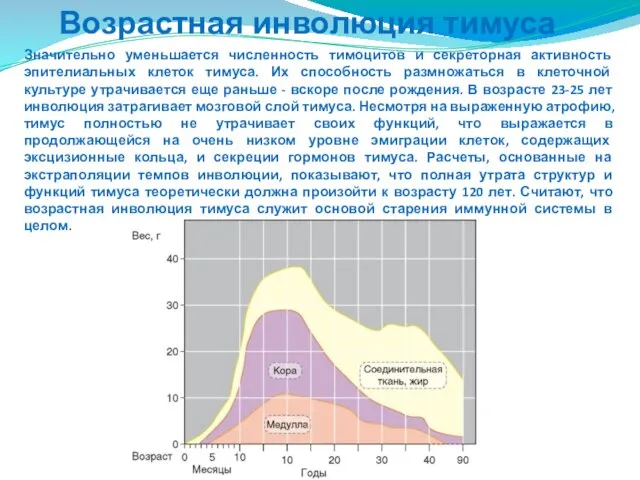
Возрастная инволюция тимуса
Важная особенность тимуса - его возрастная инволюция. Максимальный относительный
Возрастная инволюция тимуса
Важная особенность тимуса - его возрастная инволюция. Максимальный относительный
Возрастная инволюция тимуса
Значительно уменьшается численность тимоцитов и секреторная активность эпителиальных клеток
Возрастная инволюция тимуса
Значительно уменьшается численность тимоцитов и секреторная активность эпителиальных клеток
Лимфатические узлы
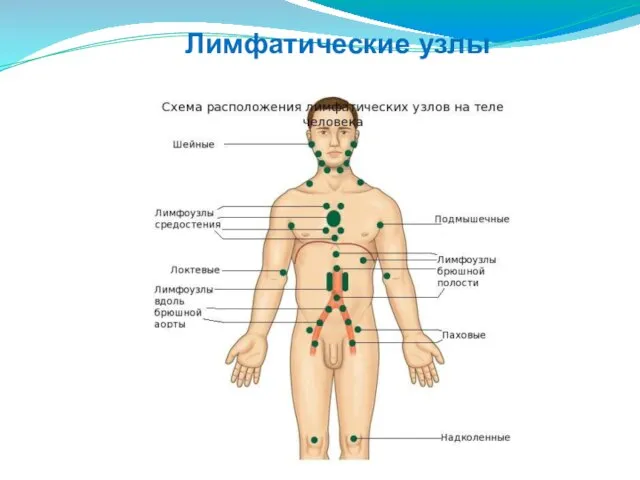
Лимфатические узлы
Лимфатические узлы - бобовидные образования величиной 0,2-1,0 см, образующие группы или цепочки.
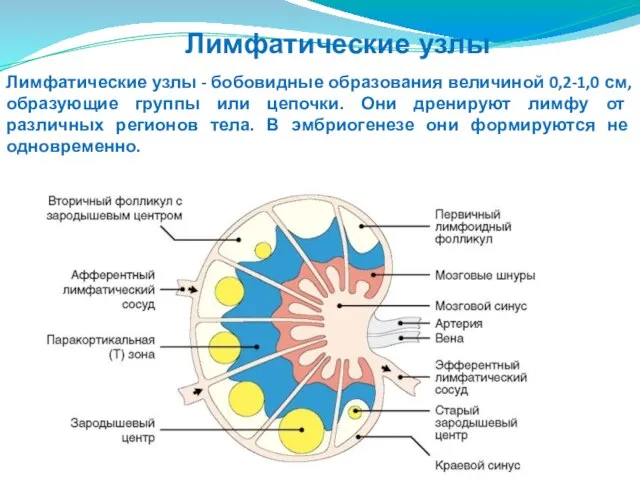
Лимфатические узлы - бобовидные образования величиной 0,2-1,0 см, образующие группы или цепочки.
Лимфатические узлы
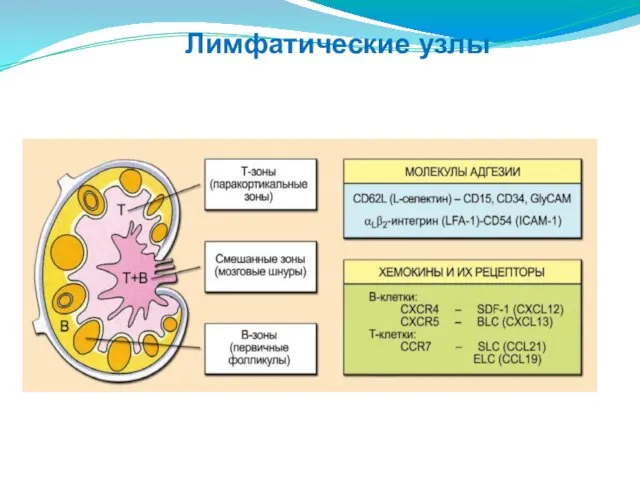
Лимфатические узлы

Селезенка служит иммунным барьером на путях гематогенного распространения патогенов и других
Селезенка служит иммунным барьером на путях гематогенного распространения патогенов и других
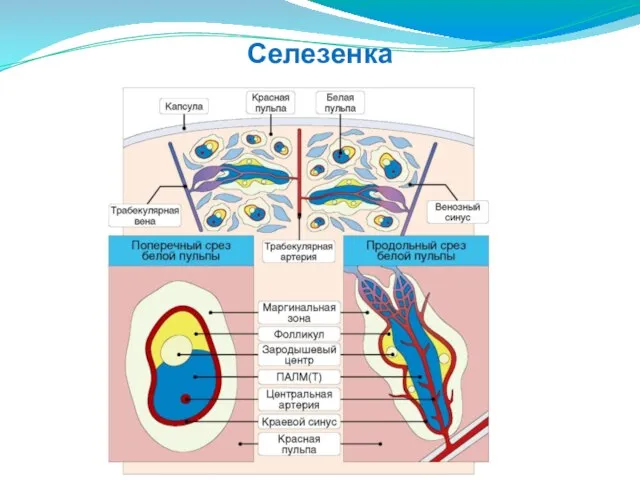
Селезенка
Селезенка
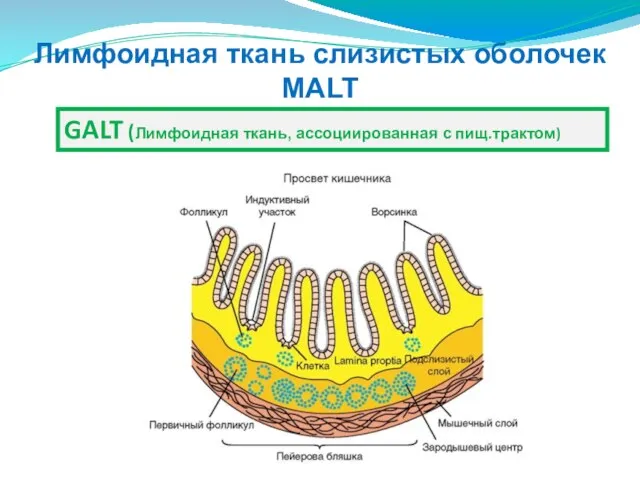
Лимфоидная ткань слизистых оболочек
MALT
GALT (Лимфоидная ткань, ассоциированная с пищ.трактом)
Лимфоидная ткань слизистых оболочек
MALT
GALT (Лимфоидная ткань, ассоциированная с пищ.трактом)

Т-лимфоциты
Т-лимфоциты
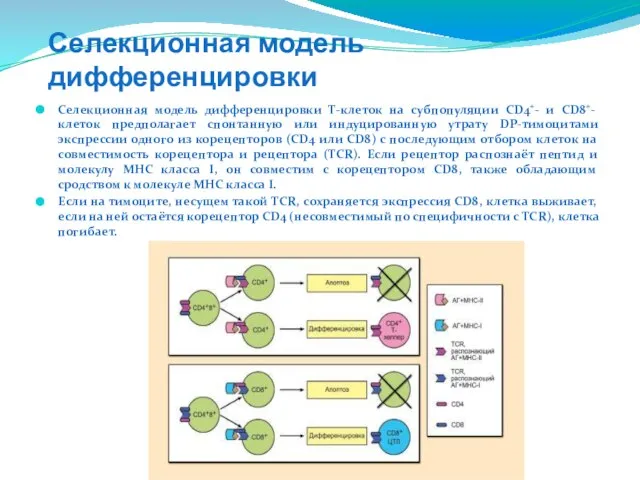
Селекционная модель дифференцировки
Селекционная модель дифференцировки Τ-клеток на субпопуляции CD4+- и CD8+-
Селекционная модель дифференцировки
Селекционная модель дифференцировки Τ-клеток на субпопуляции CD4+- и CD8+-
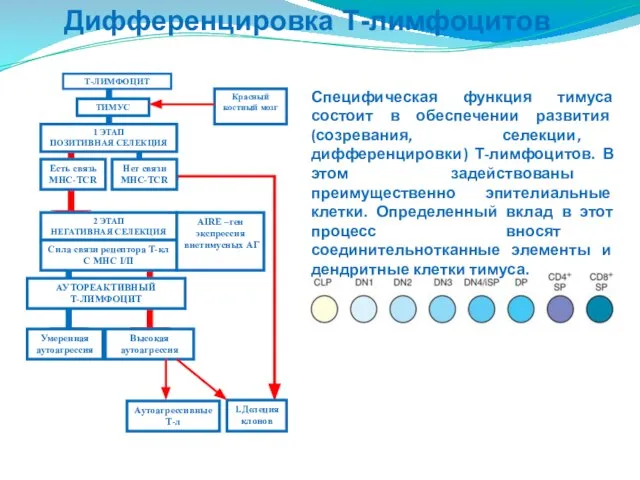
Дифференцировка Т-лимфоцитов
Красный костный мозг
ТИМУС
1 ЭТАП
ПОЗИТИВНАЯ СЕЛЕКЦИЯ
2 ЭТАП
НЕГАТИВНАЯ СЕЛЕКЦИЯ
Есть связь
МНС-ТСR
Сила связи
Дифференцировка Т-лимфоцитов
Красный костный мозг
ТИМУС
1 ЭТАП
ПОЗИТИВНАЯ СЕЛЕКЦИЯ
2 ЭТАП
НЕГАТИВНАЯ СЕЛЕКЦИЯ
Есть связь
МНС-ТСR
Сила связи
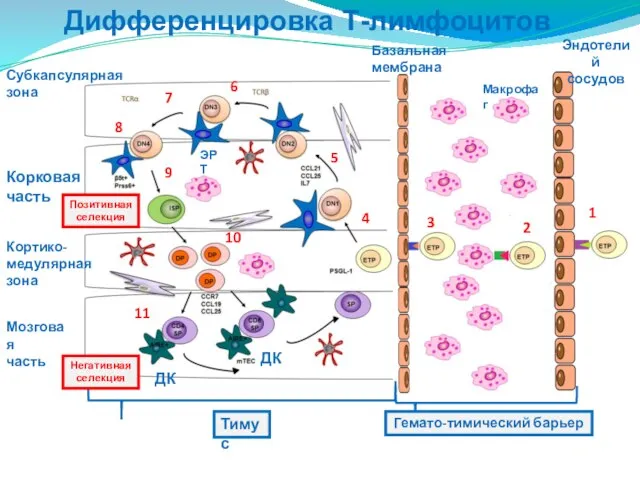
Дифференцировка Т-лимфоцитов
Субкапсулярная
зона
Корковая
часть
Кортико-
медулярная
зона
Мозговая
часть
Позитивная
селекция
Базальная
мембрана
Эндотелий
сосудов
Тимус
Гемато-тимический барьер
1
2
3
4
5
6
7
8
9
Негативная
селекция
10
11
Макрофаг
ЭРТ
ДК
ДК
Дифференцировка Т-лимфоцитов
Субкапсулярная
зона
Корковая
часть
Кортико-
медулярная
зона
Мозговая
часть
Позитивная
селекция
Базальная
мембрана
Эндотелий
сосудов
Тимус
Гемато-тимический барьер
1
2
3
4
5
6
7
8
9
Негативная
селекция
10
11
Макрофаг
ЭРТ
ДК
ДК
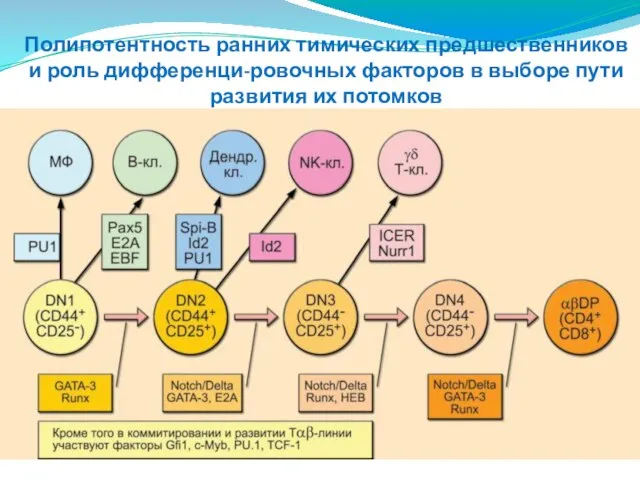
Полипотентность ранних тимических предшественников и роль дифференци-ровочных факторов в выборе пути
Полипотентность ранних тимических предшественников и роль дифференци-ровочных факторов в выборе пути

Кластерные молекулы Т-лмфоцитов
Популяции Т- и В-клеток имеют клональную структуру: в процессе
Кластерные молекулы Т-лмфоцитов
Популяции Т- и В-клеток имеют клональную структуру: в процессе
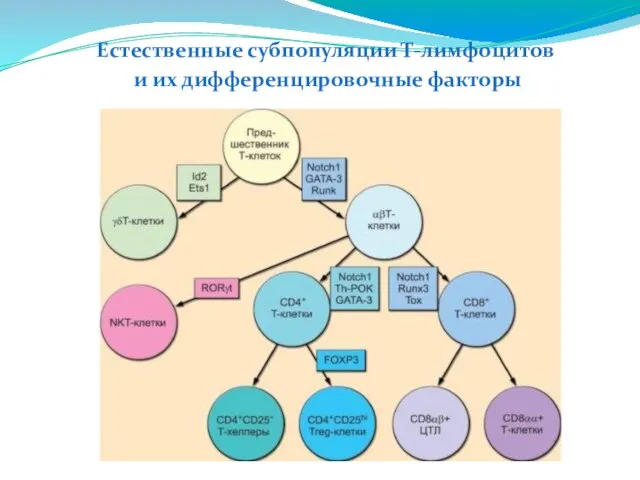
Естественные субпопуляции Т-лимфоцитов
и их дифференцировочные факторы
Естественные субпопуляции Т-лимфоцитов
и их дифференцировочные факторы
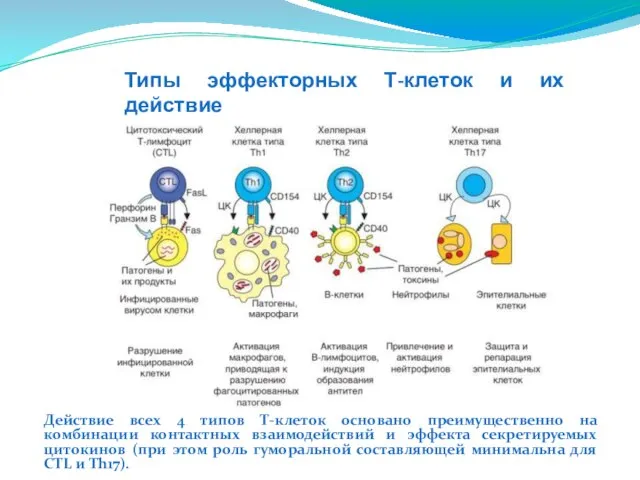
Типы эффекторных Т-клеток и их действие
Действие всех 4 типов Т-клеток основано
Типы эффекторных Т-клеток и их действие
Действие всех 4 типов Т-клеток основано

СD 4+ клетки
СD 4+ клетки
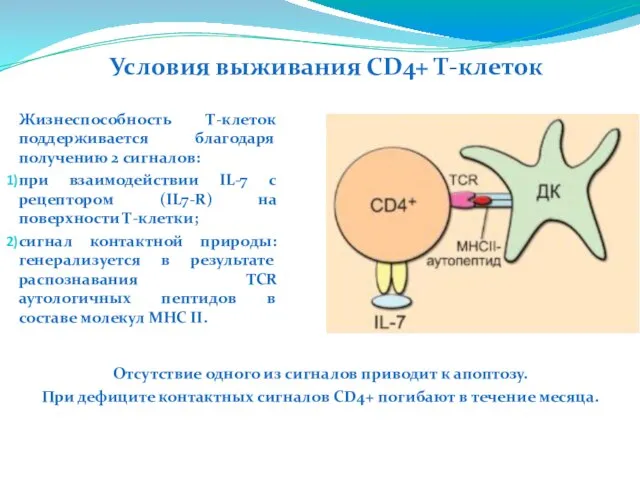
Жизнеспособность Т-клеток поддерживается благодаря получению 2 сигналов:
при взаимодействии IL-7 с рецептором
при взаимодействии IL-7 с рецептором
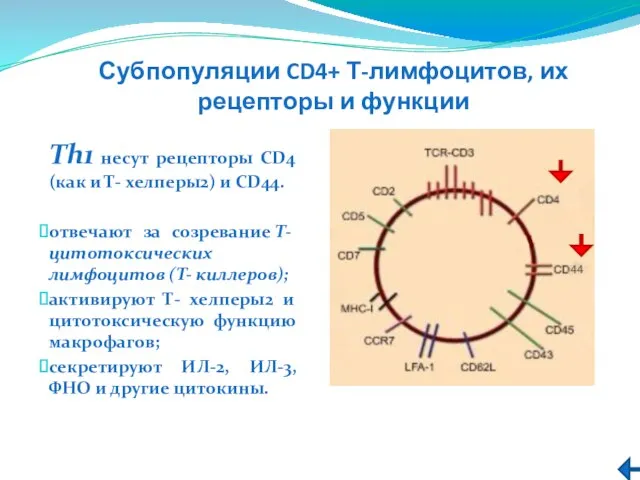
Субпопуляции CD4+ Т-лимфоцитов, их рецепторы и функции
Тh1 несут рецепторы CD4 (как и
Субпопуляции CD4+ Т-лимфоцитов, их рецепторы и функции
Тh1 несут рецепторы CD4 (как и
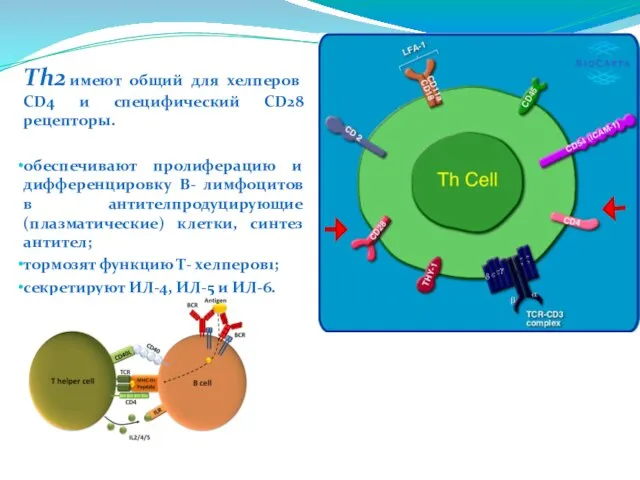
Тh2 имеют общий для хелперов CD4 и специфический CD28 рецепторы.
обеспечивают пролиферацию и
Тh2 имеют общий для хелперов CD4 и специфический CD28 рецепторы.
обеспечивают пролиферацию и

Маркерные цитокины и рецепторы
Th1- и Th2-клеток
Маркерные цитокины и рецепторы
Th1- и Th2-клеток
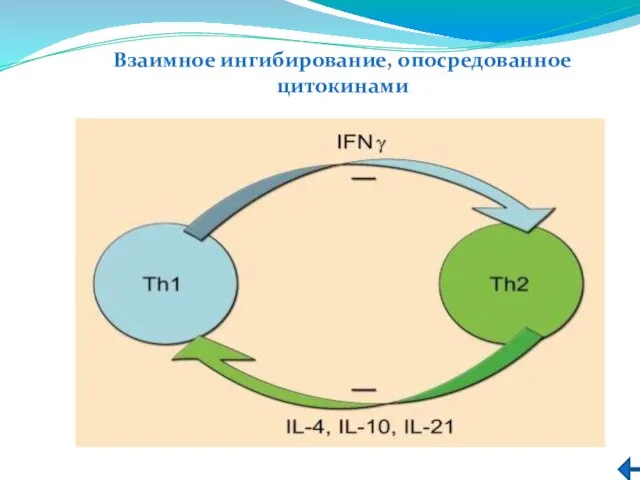
Взаимное ингибирование, опосредованное цитокинами
Взаимное ингибирование, опосредованное цитокинами
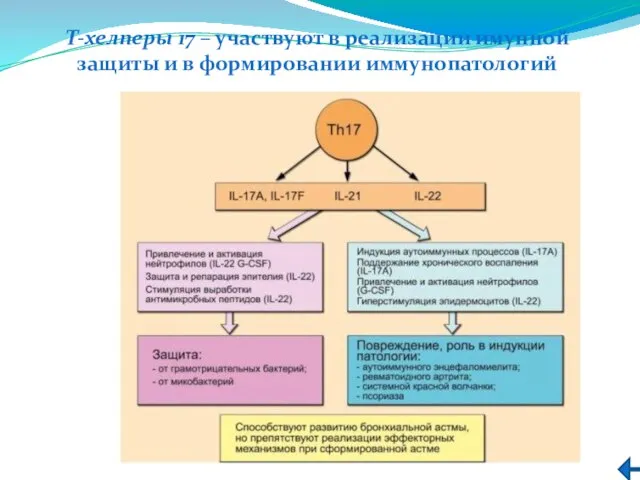
T-хелперы 17 – участвуют в реализации имунной защиты и в формировании
T-хелперы 17 – участвуют в реализации имунной защиты и в формировании
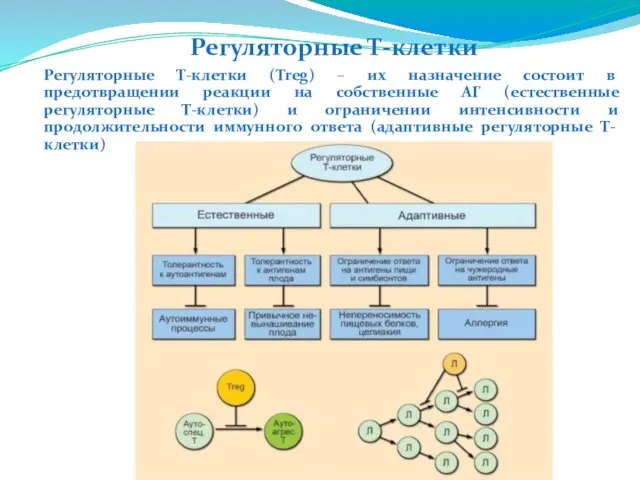
Регуляторные Т-клетки (Treg) – их назначение состоит в предотвращении реакции на
Регуляторные Т-клетки (Treg) – их назначение состоит в предотвращении реакции на
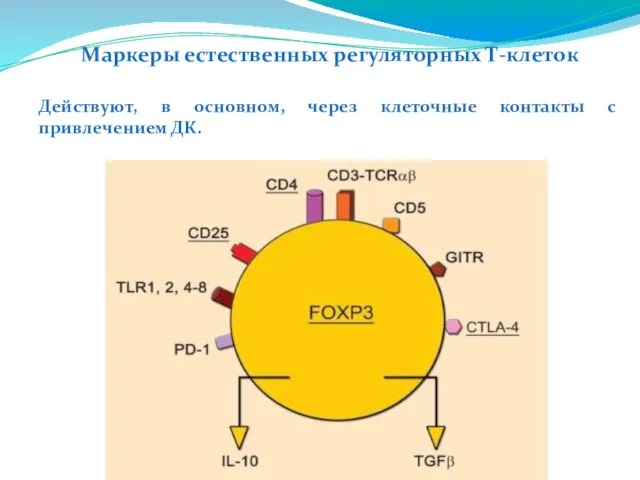
Маркеры естественных регуляторных Т-клеток
Действуют, в основном, через клеточные контакты с привлечением
Маркеры естественных регуляторных Т-клеток
Действуют, в основном, через клеточные контакты с привлечением
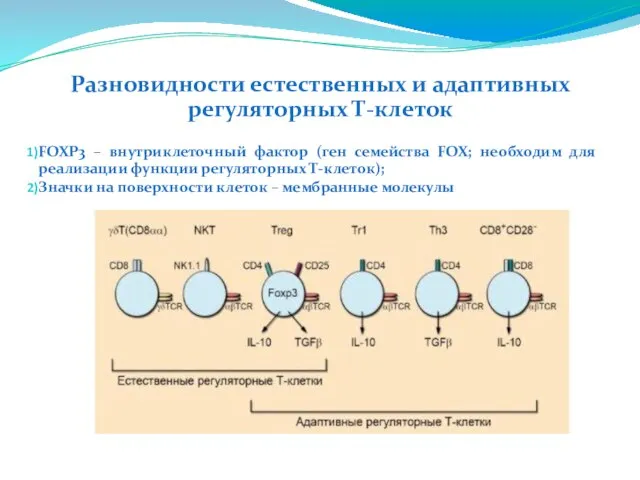
Разновидности естественных и адаптивных регуляторных Т-клеток
FOXP3 – внутриклеточный фактор (ген семейства
Разновидности естественных и адаптивных регуляторных Т-клеток
FOXP3 – внутриклеточный фактор (ген семейства
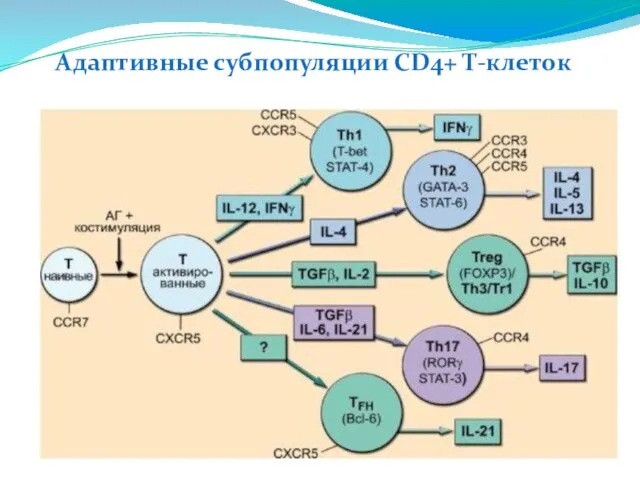
Адаптивные субпопуляции CD4+ Т-клеток
Адаптивные субпопуляции CD4+ Т-клеток
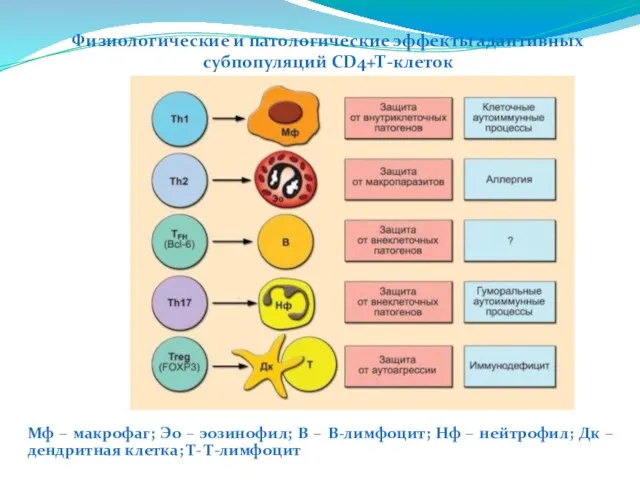
Физиологические и патологические эффекты адаптивных субпопуляций CD4+Т-клеток
Мф – макрофаг; Эо –
Физиологические и патологические эффекты адаптивных субпопуляций CD4+Т-клеток
Мф – макрофаг; Эо –
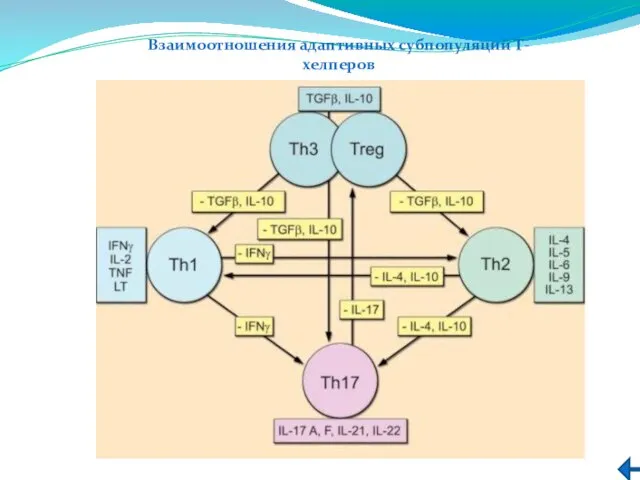
Взаимоотношения адаптивных субпопуляций Т-хелперов
Взаимоотношения адаптивных субпопуляций Т-хелперов
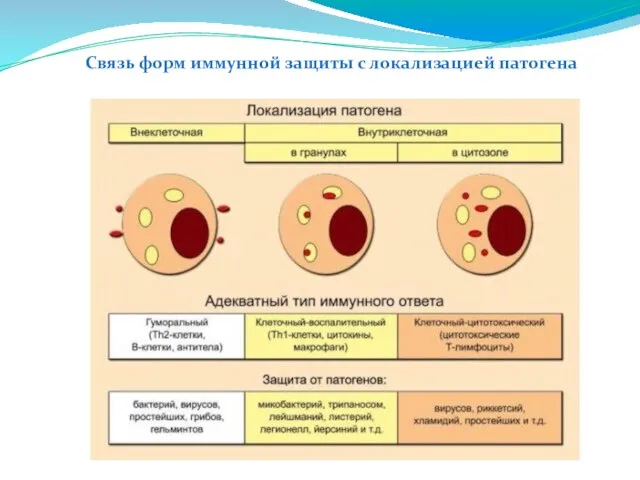
Связь форм иммунной защиты с локализацией патогена
Связь форм иммунной защиты с локализацией патогена
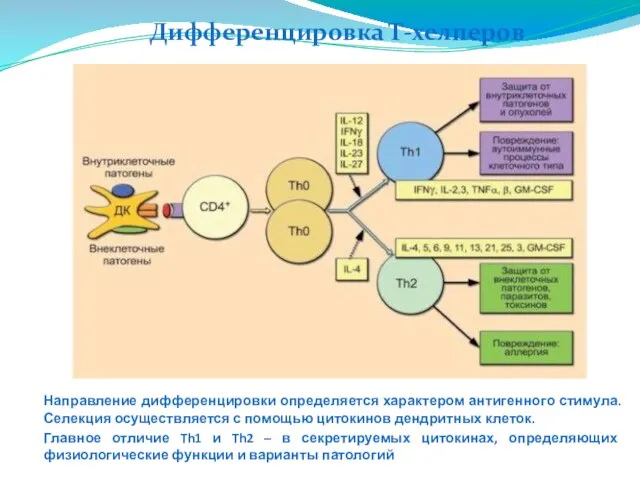
Дифференцировка Т-хелперов
Направление дифференцировки определяется характером антигенного стимула. Селекция осуществляется с помощью
Дифференцировка Т-хелперов
Направление дифференцировки определяется характером антигенного стимула. Селекция осуществляется с помощью

Факторы, определяющие преобладание иммунного ответа Th1- или Th2-типа
Факторы, определяющие преобладание иммунного ответа Th1- или Th2-типа

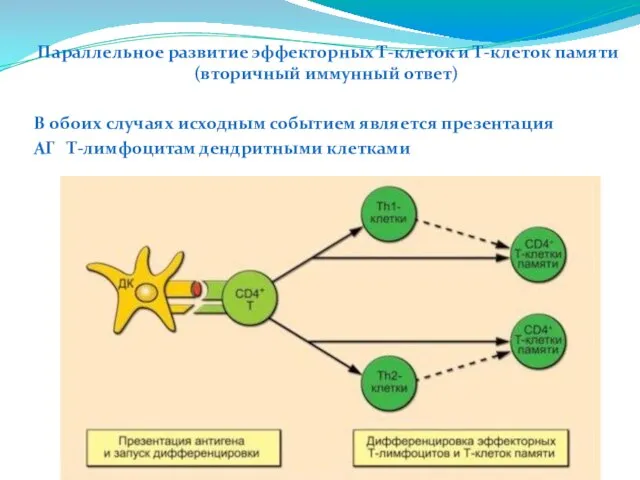
Параллельное развитие эффекторных Т-клеток и Т-клеток памяти (вторичный иммунный ответ)
В обоих
Параллельное развитие эффекторных Т-клеток и Т-клеток памяти (вторичный иммунный ответ)
В обоих

СD 8+ клетки
СD 8+ клетки
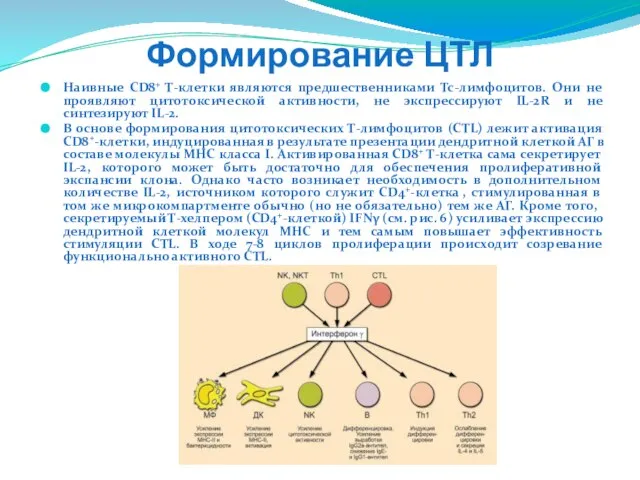
Формирование ЦТЛ
Наивные CD8+ Т-клетки являются предшественниками Tc-лимфоцитов. Они не проявляют цитотоксической
Формирование ЦТЛ
Наивные CD8+ Т-клетки являются предшественниками Tc-лимфоцитов. Они не проявляют цитотоксической
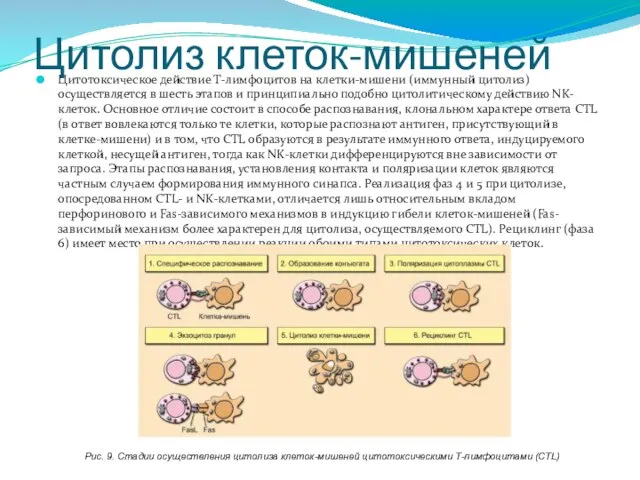
Цитолиз клеток-мишеней
Цитотоксическое действие Т-лимфоцитов на клетки-мишени (иммунный цитолиз) осуществляется в шесть
Цитолиз клеток-мишеней
Цитотоксическое действие Т-лимфоцитов на клетки-мишени (иммунный цитолиз) осуществляется в шесть

Гранулы ЦТЛ и апоптоз клеток-мишеней
В гранулах содержатся белки, участвующие в реализации
Гранулы ЦТЛ и апоптоз клеток-мишеней
В гранулах содержатся белки, участвующие в реализации
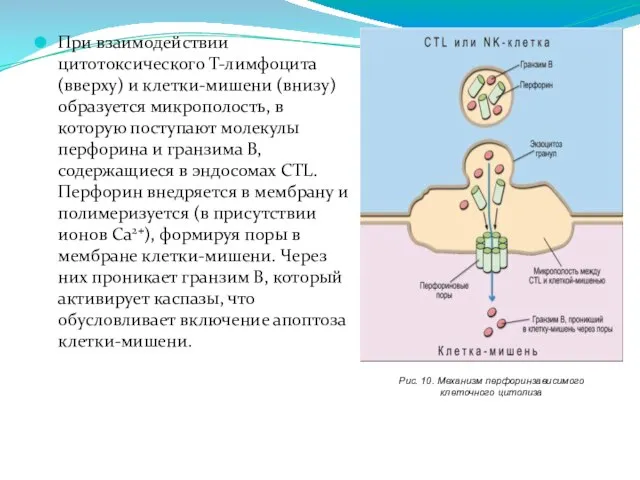
При взаимодействии цитотоксического Т-лимфоцита (вверху) и клетки-мишени (внизу) образуется микрополость, в
При взаимодействии цитотоксического Т-лимфоцита (вверху) и клетки-мишени (внизу) образуется микрополость, в
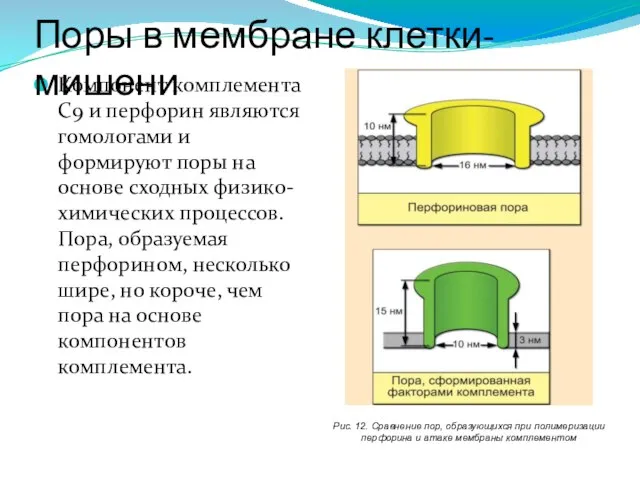
Компонент комплемента С9 и перфорин являются гомологами и формируют поры на
Компонент комплемента С9 и перфорин являются гомологами и формируют поры на
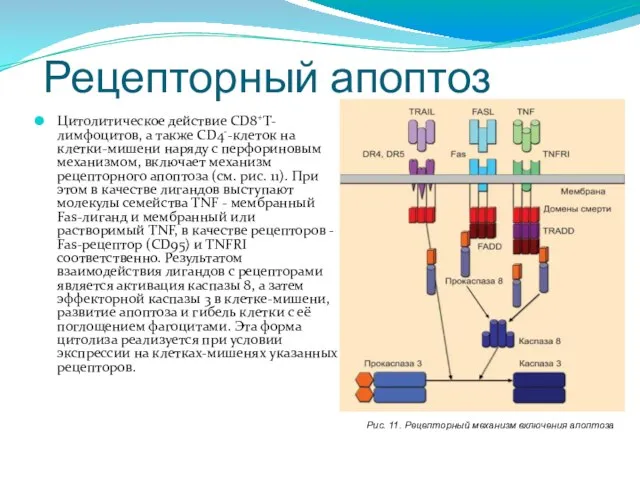
Рецепторный апоптоз
Цитолитическое действие СD8+Т-лимфоцитов, а также CD4--клеток на клетки-мишени наряду с
Рецепторный апоптоз
Цитолитическое действие СD8+Т-лимфоцитов, а также CD4--клеток на клетки-мишени наряду с

Направленность действия
Действие Tc-лимфоцитов направлено, главным образом, против вирус-инфицированных и опухолевых клеток.
Направленность действия
Действие Tc-лимфоцитов направлено, главным образом, против вирус-инфицированных и опухолевых клеток.
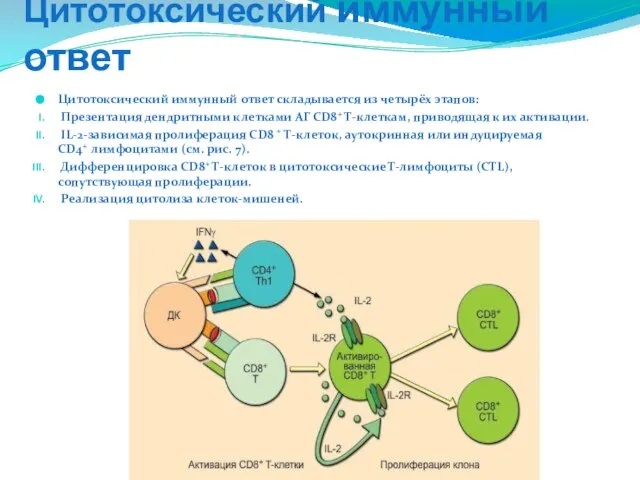
Цитотоксический иммунный ответ
Цитотоксический иммунный ответ складывается из четырёх этапов:
Презентация дендритными клетками
Цитотоксический иммунный ответ
Цитотоксический иммунный ответ складывается из четырёх этапов:
Презентация дендритными клетками
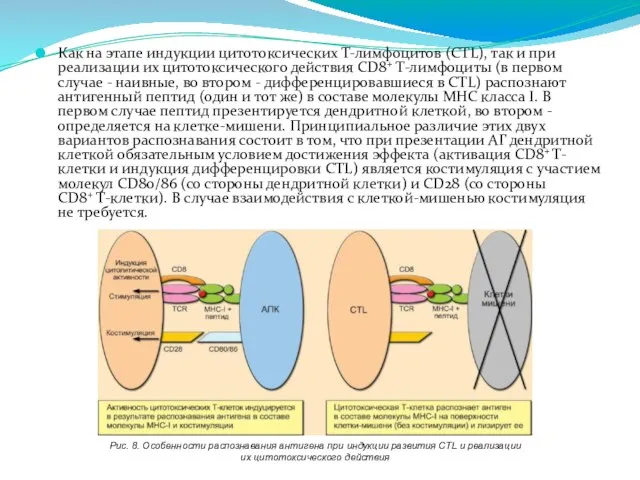
Как на этапе индукции цитотоксических Т-лимфоцитов (CTL), так и при реализации
Как на этапе индукции цитотоксических Т-лимфоцитов (CTL), так и при реализации
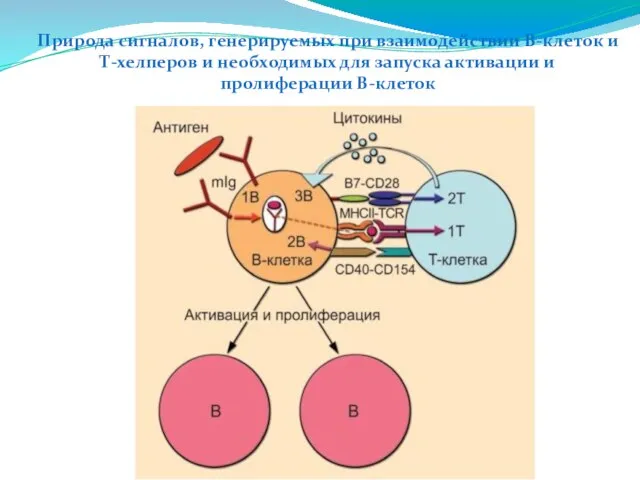
Природа сигналов, генерируемых при взаимодействии В-клеток и Т-хелперов и необходимых для
Природа сигналов, генерируемых при взаимодействии В-клеток и Т-хелперов и необходимых для
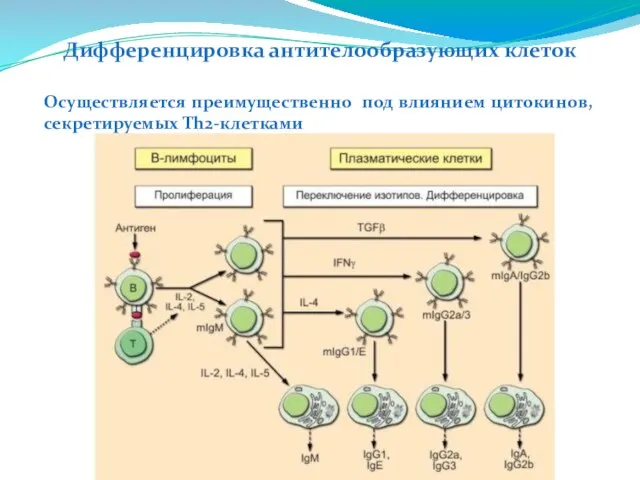
Дифференцировка антителообразующих клеток
Осуществляется преимущественно под влиянием цитокинов, секретируемых Th2-клетками
Дифференцировка антителообразующих клеток
Осуществляется преимущественно под влиянием цитокинов, секретируемых Th2-клетками
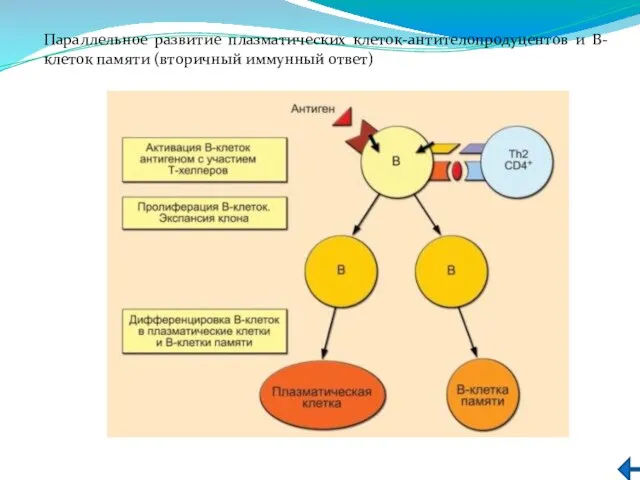
Параллельное развитие плазматических клеток-антителопродуцентов и В-клеток памяти (вторичный иммунный ответ)
Параллельное развитие плазматических клеток-антителопродуцентов и В-клеток памяти (вторичный иммунный ответ)
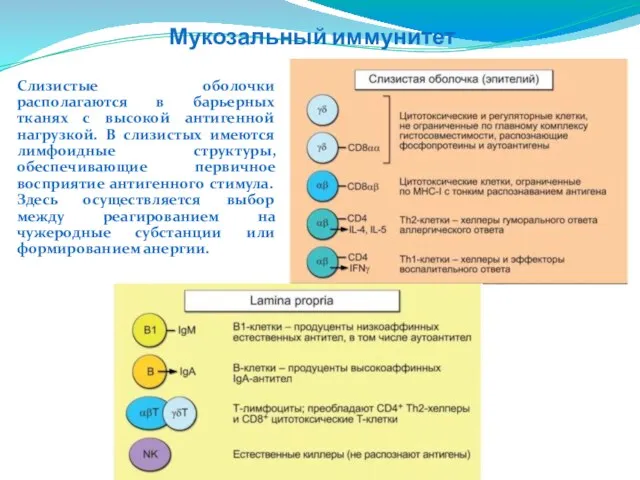
Мукозальный иммунитет
Слизистые оболочки располагаются в барьерных тканях с высокой антигенной нагрузкой.
Мукозальный иммунитет
Слизистые оболочки располагаются в барьерных тканях с высокой антигенной нагрузкой.
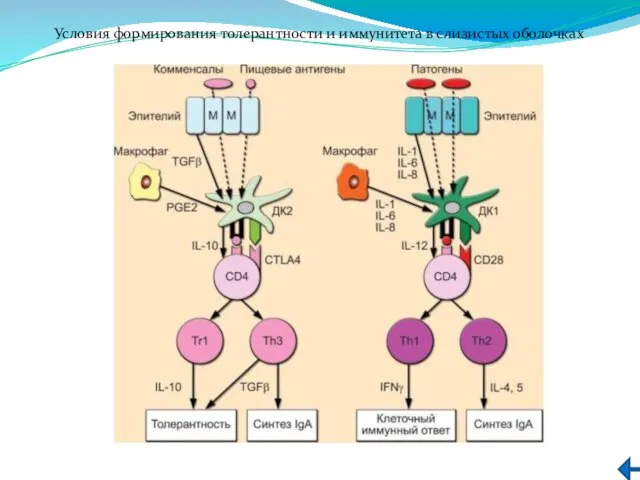
Условия формирования толерантности и иммунитета в слизистых оболочках
Условия формирования толерантности и иммунитета в слизистых оболочках


В-лимфоциты представляют собой клетки адаптивного иммунитета – предшественники плазматических клеток, продуцирующих
В-лимфоциты представляют собой клетки адаптивного иммунитета – предшественники плазматических клеток, продуцирующих
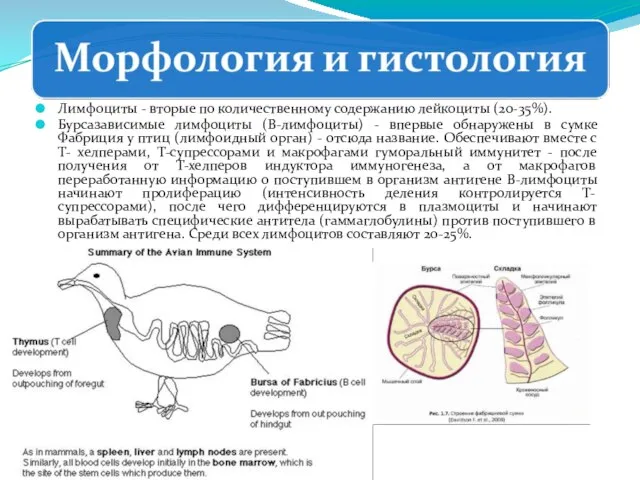
Лимфоциты - вторые по количественному содержанию лейкоциты (20-35%).
Бурсазависимые лимфоциты (В-лимфоциты) -
Лимфоциты - вторые по количественному содержанию лейкоциты (20-35%).
Бурсазависимые лимфоциты (В-лимфоциты) -
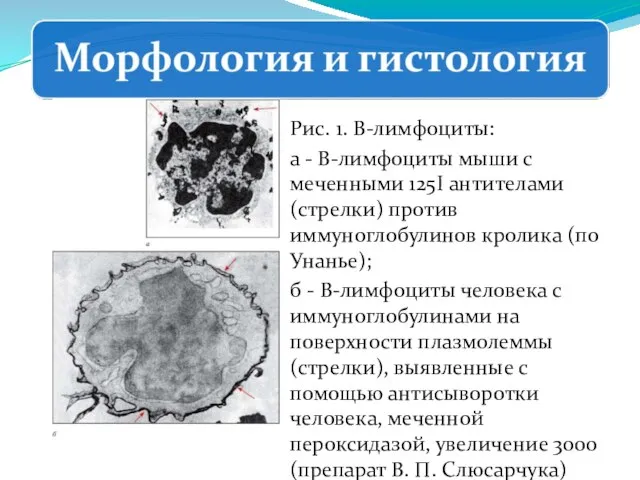
Рис. 1. В-лимфоциты:
а - В-лимфоциты мыши с меченными 125I антителами (стрелки)
Рис. 1. В-лимфоциты:
а - В-лимфоциты мыши с меченными 125I антителами (стрелки)

По морфологическим признакам В- и Т-лимфоциты и их субпопуляции различать затруднительно
По морфологическим признакам В- и Т-лимфоциты и их субпопуляции различать затруднительно
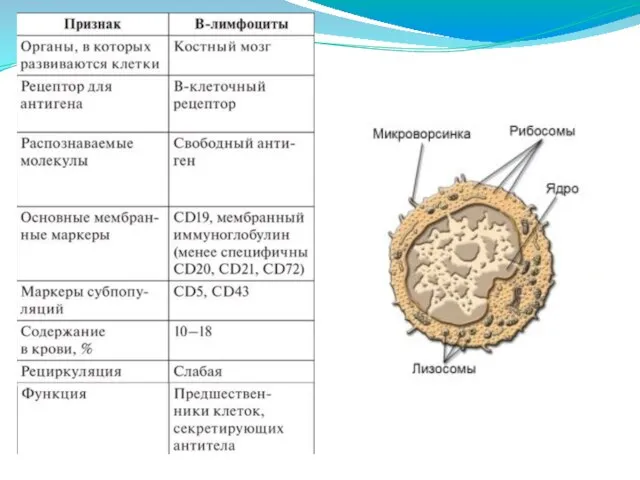

Образование В-лимфоцитов начинается в эмбриональном периоде и продолжается в течение всей
Образование В-лимфоцитов начинается в эмбриональном периоде и продолжается в течение всей
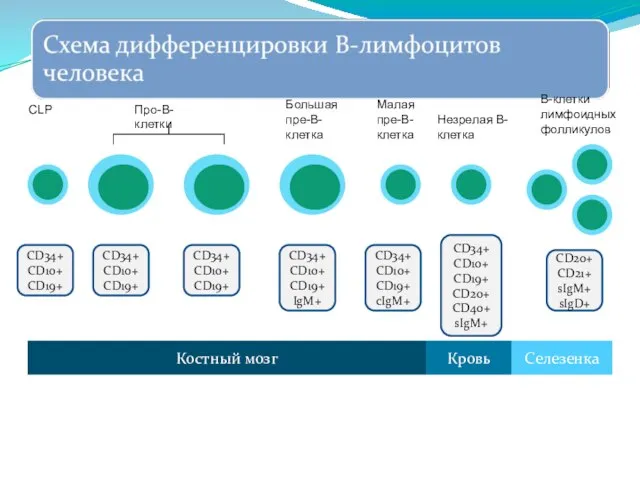
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
IgM+
CD34+
CD10+
CD19+
cIgM+
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
CD34+
CD10+
CD19+
CD20+
CD21+
sIgM+
sIgD+
Костный мозг
Селезенка
Кровь
CLP
Про-В-клетки
Большая пре-В-клетка
Малая пре-В-клетка
Незрелая В-клетка
В-клетки лимфоидных фолликулов
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
IgM+
CD34+
CD10+
CD19+
cIgM+
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
CD34+
CD10+
CD19+
CD20+
CD21+
sIgM+
sIgD+
Костный мозг
Селезенка
Кровь
CLP
Про-В-клетки
Большая пре-В-клетка
Малая пре-В-клетка
Незрелая В-клетка
В-клетки лимфоидных фолликулов
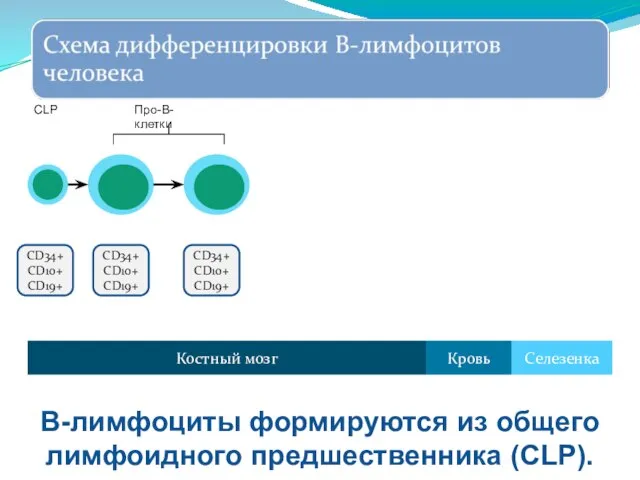
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
Костный мозг
Селезенка
Кровь
CLP
Про-В-клетки
В-лимфоциты формируются из общего лимфоидного предшественника (CLP).
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
Костный мозг
Селезенка
Кровь
CLP
Про-В-клетки
В-лимфоциты формируются из общего лимфоидного предшественника (CLP).
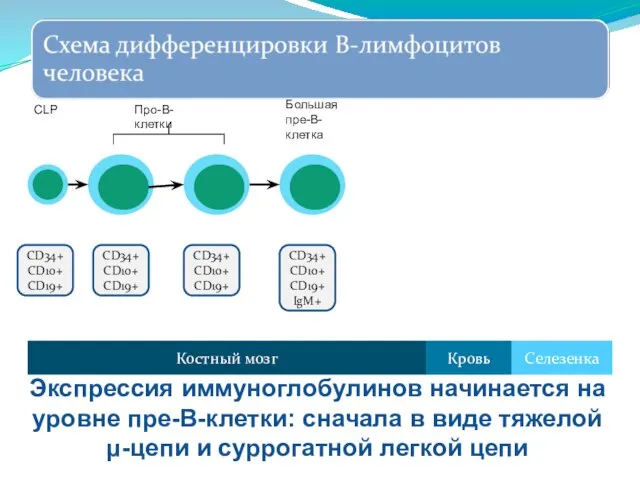
CD34+
CD10+
CD19+
IgM+
Костный мозг
Селезенка
Кровь
Большая пре-В-клетка
Экспрессия иммуноглобулинов начинается на уровне пре-В-клетки: сначала в виде
CD34+
CD10+
CD19+
IgM+
Костный мозг
Селезенка
Кровь
Большая пре-В-клетка
Экспрессия иммуноглобулинов начинается на уровне пре-В-клетки: сначала в виде
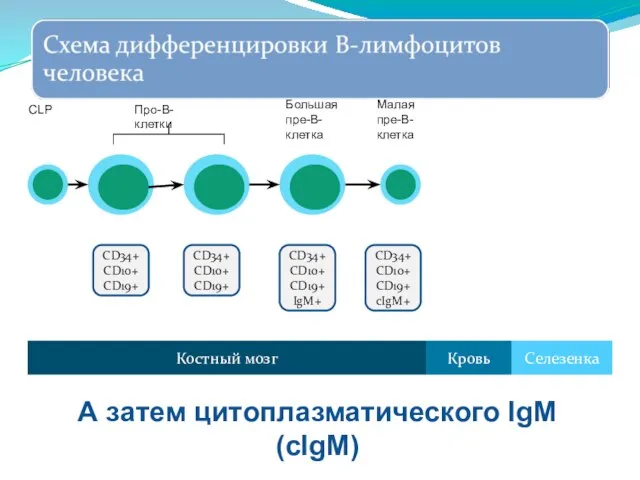
CD34+
CD10+
CD19+
cIgM+
Костный мозг
Селезенка
Кровь
Малая пре-В-клетка
А затем цитоплазматического IgM (cIgM)
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CLP
Про-В-клетки
CD34+
CD10+
CD19+
IgM+
Большая пре-В-клетка
CD34+
CD10+
CD19+
cIgM+
Костный мозг
Селезенка
Кровь
Малая пре-В-клетка
А затем цитоплазматического IgM (cIgM)
CD34+
CD10+
CD19+
CD34+
CD10+
CD19+
CLP
Про-В-клетки
CD34+
CD10+
CD19+
IgM+
Большая пре-В-клетка
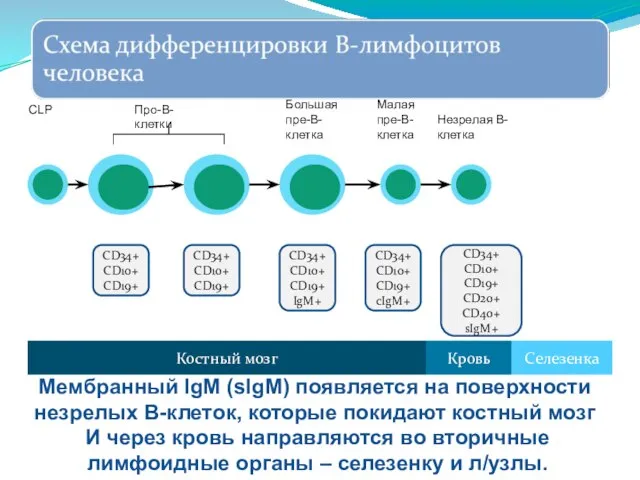
CD34+
CD10+
CD19+
cIgM+
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
Костный мозг
Селезенка
Кровь
Малая пре-В-клетка
Незрелая В-клетка
Мембранный IgM (sIgM) появляется на поверхности незрелых В-клеток,
CD34+
CD10+
CD19+
cIgM+
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
Костный мозг
Селезенка
Кровь
Малая пре-В-клетка
Незрелая В-клетка
Мембранный IgM (sIgM) появляется на поверхности незрелых В-клеток,
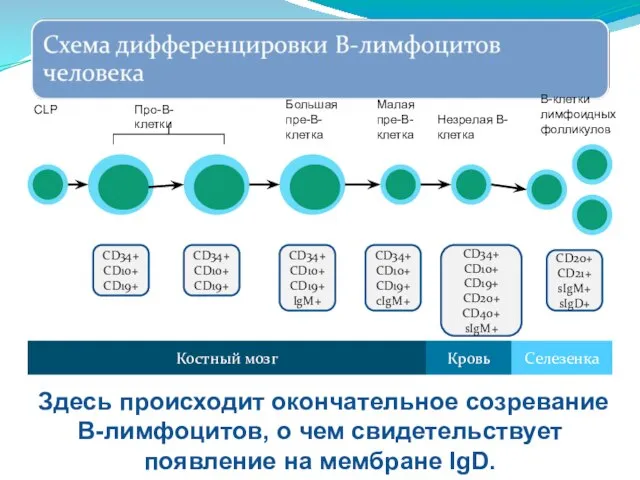
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
CD20+
CD21+
sIgM+
sIgD+
Костный мозг
Селезенка
Кровь
Незрелая В-клетка
В-клетки лимфоидных фолликулов
Здесь происходит окончательное созревание В-лимфоцитов, о чем
CD34+
CD10+
CD19+
CD20+
CD40+
sIgM+
CD20+
CD21+
sIgM+
sIgD+
Костный мозг
Селезенка
Кровь
Незрелая В-клетка
В-клетки лимфоидных фолликулов
Здесь происходит окончательное созревание В-лимфоцитов, о чем
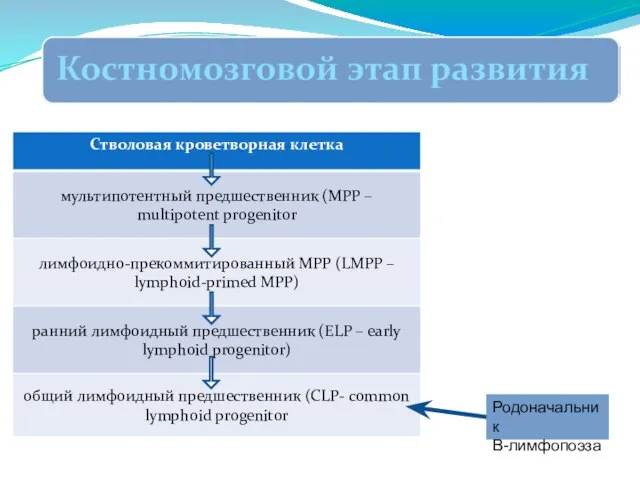
Родоначальник
В-лимфопоэза
Родоначальник
В-лимфопоэза

Негативная селекция и делеция клонов. Связывание мембранного антигена незрелой B-клеткой (экспрессирует
Негативная селекция и делеция клонов. Связывание мембранного антигена незрелой B-клеткой (экспрессирует
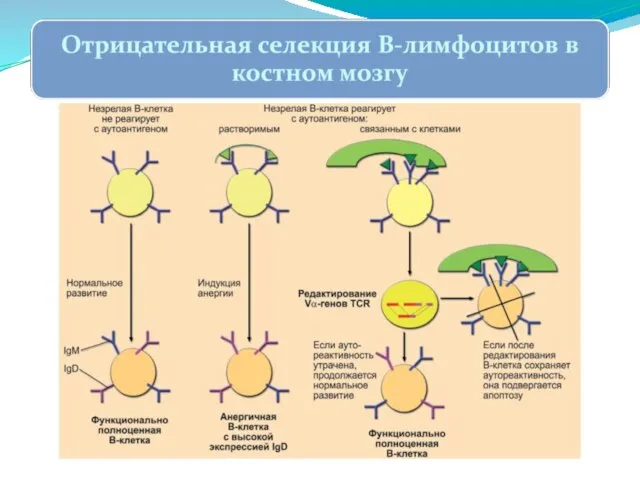
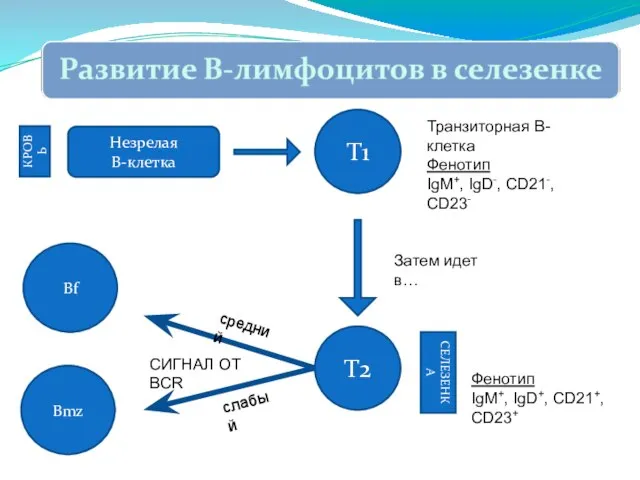
Незрелая
В-клетка
КРОВЬ
Т1
Транзиторная В-клетка
Фенотип
IgM+, IgD-, CD21-, CD23-
Т2
СЕЛЕЗЕНКА
Фенотип
IgM+, IgD+, CD21+, CD23+
Bf
Bmz
СИГНАЛ ОТ BCR
средний
слабый
Затем
Незрелая
В-клетка
КРОВЬ
Т1
Транзиторная В-клетка
Фенотип
IgM+, IgD-, CD21-, CD23-
Т2
СЕЛЕЗЕНКА
Фенотип
IgM+, IgD+, CD21+, CD23+
Bf
Bmz
СИГНАЛ ОТ BCR
средний
слабый
Затем

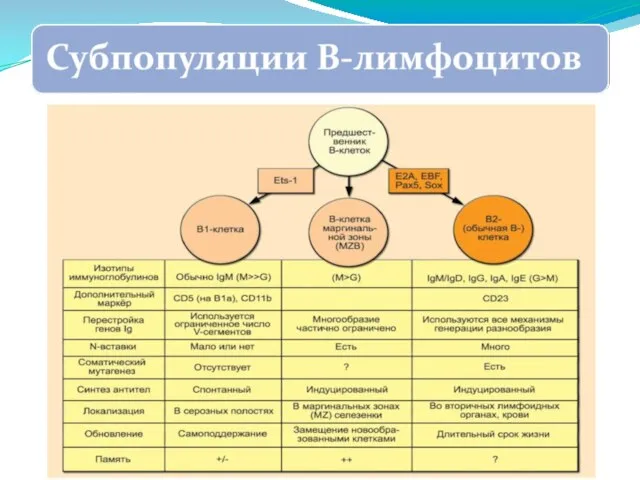
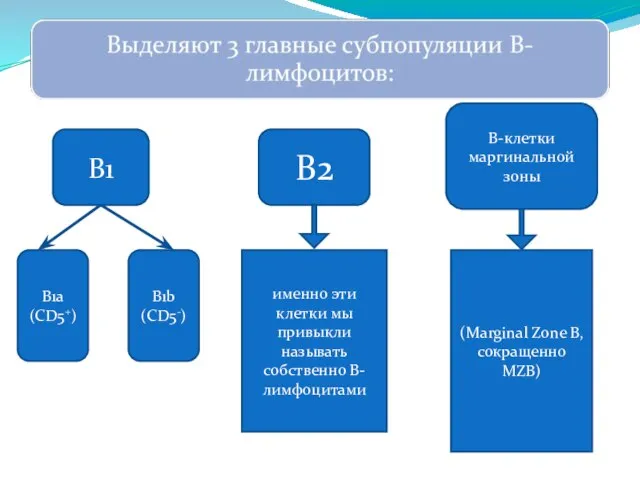
В1
В2
В-клетки маргинальной зоны
именно эти клетки мы привыкли называть собственно В-лимфоцитами
(Marginal Zone
В1
В2
В-клетки маргинальной зоны
именно эти клетки мы привыкли называть собственно В-лимфоцитами
(Marginal Zone

Предназначение B1-лимфоцитов - быстрый ответ на проникающие в организм широко распространённые
Предназначение B1-лимфоцитов - быстрый ответ на проникающие в организм широко распространённые

Предполагают, что основная функция B1a-клеток - секреция естественных антител.
Считается, что
Предполагают, что основная функция B1a-клеток - секреция естественных антител.
Считается, что
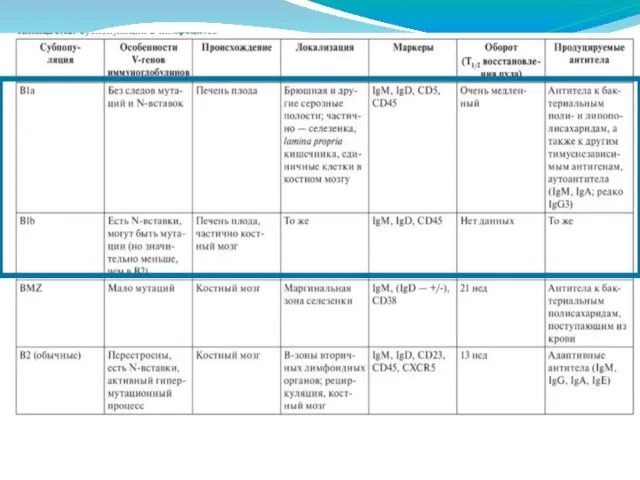
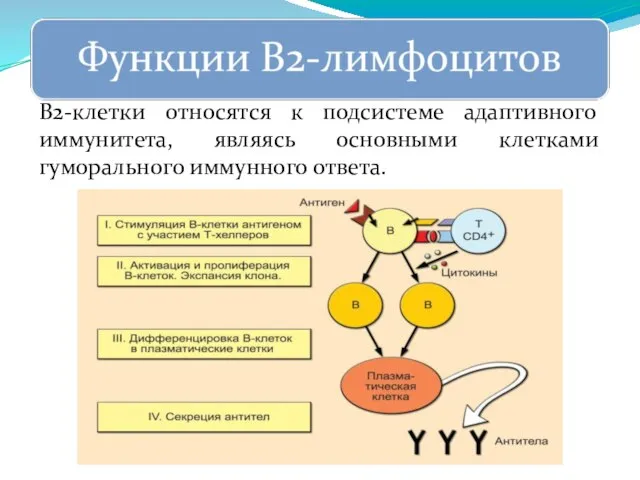
В2-клетки относятся к подсистеме адаптивного иммунитета, являясь основными клетками гуморального иммунного
В2-клетки относятся к подсистеме адаптивного иммунитета, являясь основными клетками гуморального иммунного
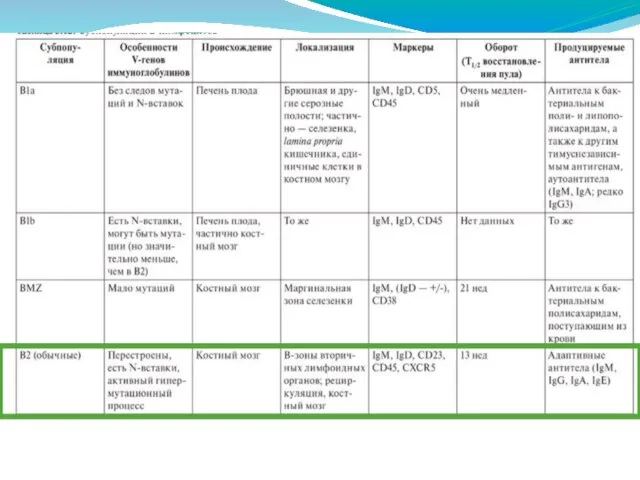

Их основной задачей является ответ на Т-независимые антигены, находящиеся в крови.
Их основной задачей является ответ на Т-независимые антигены, находящиеся в крови.

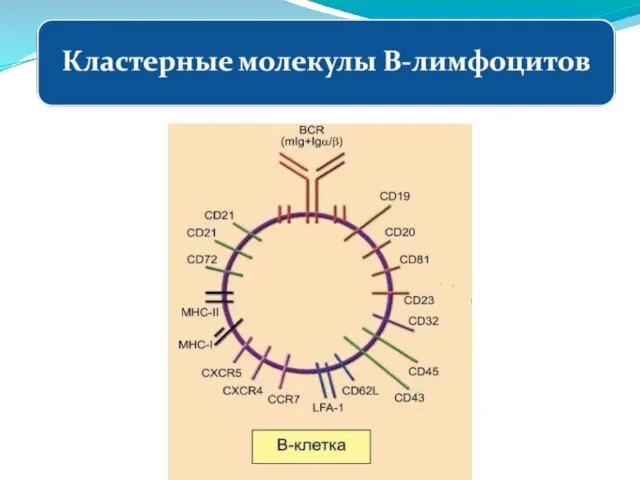

Антитела
Антигенраспознающие молекулы (АРМ) являются основными молекулами системы адаптивного иммунитета.
Секретируемые иммуноглобулины
Антитела
Антигенраспознающие молекулы (АРМ) являются основными молекулами системы адаптивного иммунитета.
Секретируемые иммуноглобулины

История открытия
Антитела были открыты в 1890 г., когда Э. Беринг и С.
История открытия
Антитела были открыты в 1890 г., когда Э. Беринг и С.

Строение иммуноглобулинов
Молекула Ig - мономер, состоит из двух лёгких (от light) и двух
Строение иммуноглобулинов
Молекула Ig - мономер, состоит из двух лёгких (от light) и двух
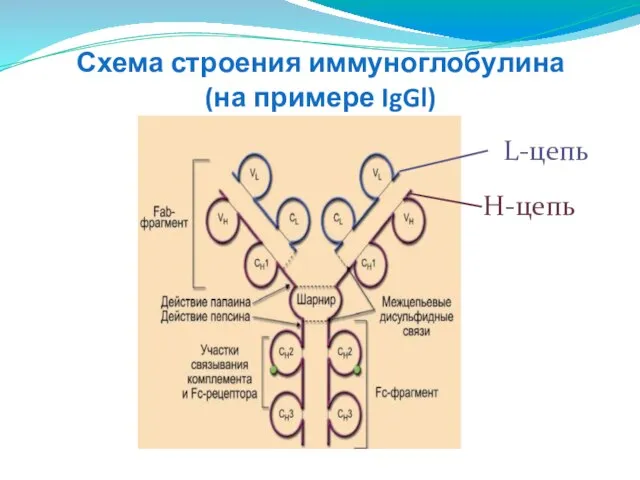
Схема строения иммуноглобулина
(на примере IgGl)
L-цепь
H-цепь
Схема строения иммуноглобулина
(на примере IgGl)
L-цепь
H-цепь
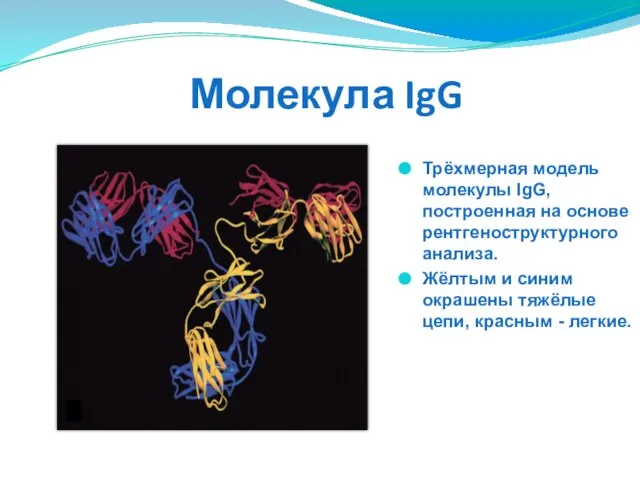
Молекула IgG
Трёхмерная модель молекулы IgG, построенная на основе рентгеноструктурного анализа.
Жёлтым и
Молекула IgG
Трёхмерная модель молекулы IgG, построенная на основе рентгеноструктурного анализа.
Жёлтым и

Изотипия иммуноглобулинов
Изотипические детерминанты специфичны для разновидностей Н- и L-цепей. Они локализуются
Изотипия иммуноглобулинов
Изотипические детерминанты специфичны для разновидностей Н- и L-цепей. Они локализуются
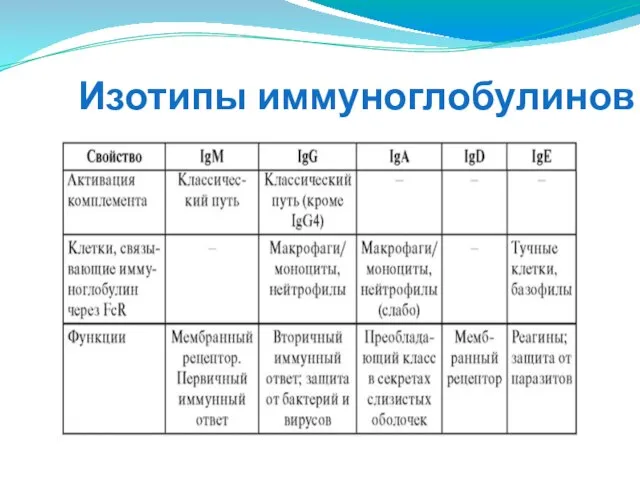
Изотипы иммуноглобулинов
Изотипы иммуноглобулинов

Аллотипия и идиотипия иммуноглобулинов
Аллотипические детерминанты отражают генетический полиморфизм полипептидных цепей иммуноглобулинов,
Аллотипия и идиотипия иммуноглобулинов
Аллотипические детерминанты отражают генетический полиморфизм полипептидных цепей иммуноглобулинов,
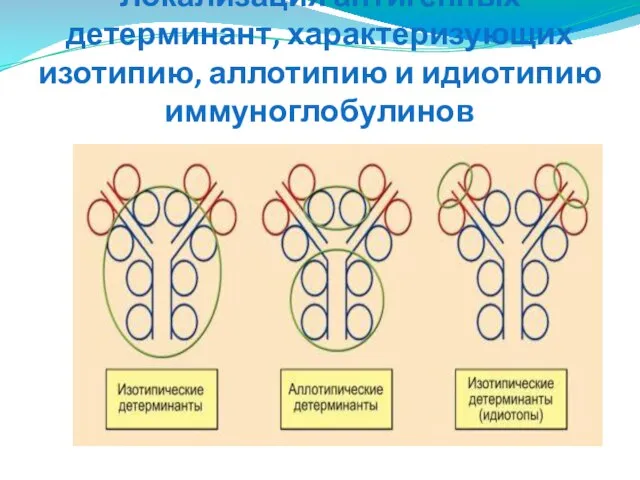
Локализация антигенных детерминант, характеризующих изотипию, аллотипию и идиотипию иммуноглобулинов
Локализация антигенных детерминант, характеризующих изотипию, аллотипию и идиотипию иммуноглобулинов

Функции антител
Проявлением эффекторной функции собственно антител является блокада АГ, приводящая к
Функции антител
Проявлением эффекторной функции собственно антител является блокада АГ, приводящая к
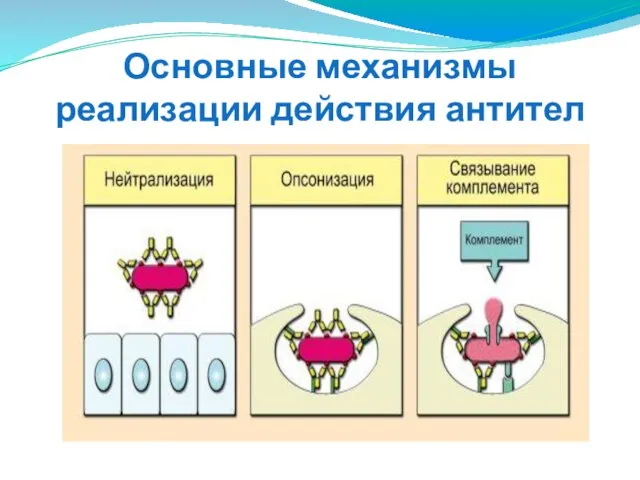
Основные механизмы реализации действия антител
Основные механизмы реализации действия антител
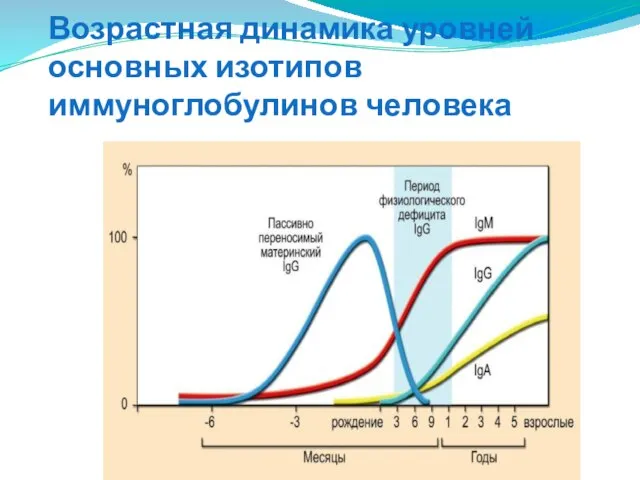
Возрастная динамика уровней основных изотипов иммуноглобулинов человека
Возрастная динамика уровней основных изотипов иммуноглобулинов человека

К моменту рождения успевает в значительной степени сформироваться только механизм выработки
К моменту рождения успевает в значительной степени сформироваться только механизм выработки
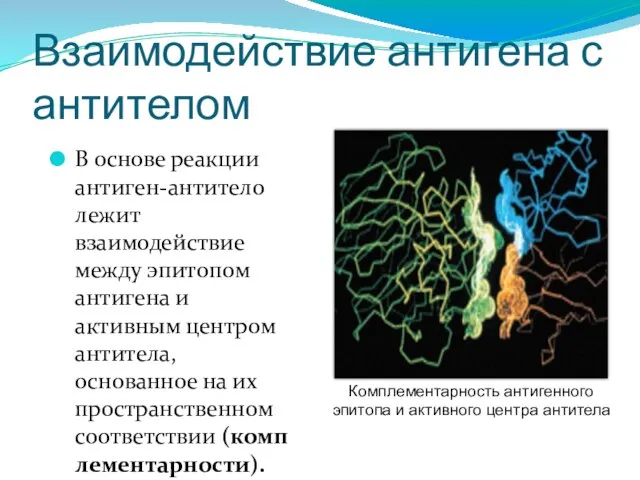
Взаимодействие антигена с антителом
В основе реакции антиген-антитело лежит взаимодействие между эпитопом
Взаимодействие антигена с антителом
В основе реакции антиген-антитело лежит взаимодействие между эпитопом
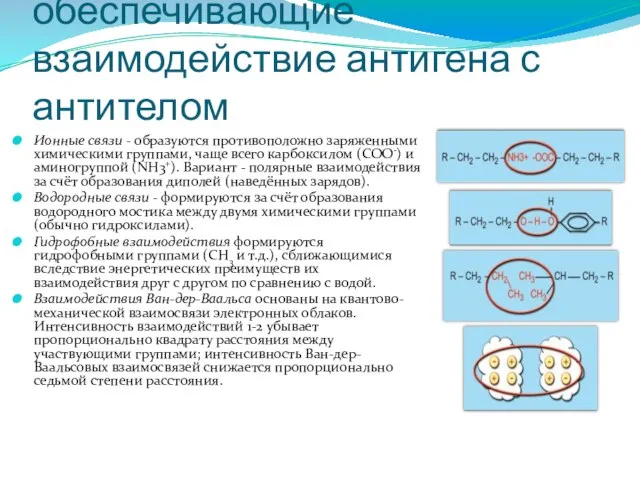
Нековалентные связи, обеспечивающие взаимодействие антигена с антителом
Ионные связи - образуются противоположно
Нековалентные связи, обеспечивающие взаимодействие антигена с антителом
Ионные связи - образуются противоположно

Афинность антител
Аффинность антител рассматривается как показатель специфичности антител к данному эпитопу.
Афинность антител
Аффинность антител рассматривается как показатель специфичности антител к данному эпитопу.

Авидность и валентность АТ
Авидность (функциональная аффинность) - суммарное сродство. Это сила,
Авидность и валентность АТ
Авидность (функциональная аффинность) - суммарное сродство. Это сила,
 Современные методы исследования в офтальмологии
Современные методы исследования в офтальмологии Эскапел новое средство для экстренной контрацепции
Эскапел новое средство для экстренной контрацепции Об этике и долге врача, врачебной деонтологии и врачевании
Об этике и долге врача, врачебной деонтологии и врачевании Пре- и постнатальный онтогенез
Пре- и постнатальный онтогенез Гастрит в детском возрасте
Гастрит в детском возрасте Визначення туберкульозу, як наукової та практичної проблеми. Історія розвитку фтизіатрії. (Лекція 1)
Визначення туберкульозу, як наукової та практичної проблеми. Історія розвитку фтизіатрії. (Лекція 1) Имплантация для чайников
Имплантация для чайников Для снятия зрительной нагрузки, рекомендую просмотр стереограмм (бумажных и электронных)
Для снятия зрительной нагрузки, рекомендую просмотр стереограмм (бумажных и электронных) Арт терапия
Арт терапия Лихорадка
Лихорадка Произведение ребенка 448 группа ОМФ 2011-2012 травматология
Произведение ребенка 448 группа ОМФ 2011-2012 травматология Антенатальная охрана плода
Антенатальная охрана плода Современные подходы к организации медицинской профилактической работы
Современные подходы к организации медицинской профилактической работы 5 игр на развитие памяти
5 игр на развитие памяти Болезни слизистой полости рта
Болезни слизистой полости рта Проходження практики у ветеринарній клініці
Проходження практики у ветеринарній клініці Микробиологическая диагностика инфекционных заболеваний. Микроскопический метод
Микробиологическая диагностика инфекционных заболеваний. Микроскопический метод Самопроизвольное прерывание беременности
Самопроизвольное прерывание беременности Тіс шірігі,тіс шірігіне қатысатын микроорганизмдер. Патогенезі. Тіс тасы , оның құрамы және тіс шірігі дамуына қатысуы
Тіс шірігі,тіс шірігіне қатысатын микроорганизмдер. Патогенезі. Тіс тасы , оның құрамы және тіс шірігі дамуына қатысуы Геморрагические диатезы
Геморрагические диатезы Применение классификации BI-RADS при ультразвуковом скрининге рака молочной железы
Применение классификации BI-RADS при ультразвуковом скрининге рака молочной железы Дрига_322_леч_электронное здравоохранение_аккредитация
Дрига_322_леч_электронное здравоохранение_аккредитация Типология конфликтов
Типология конфликтов Опухоли. Этиология, патогенез, морфогенез опухоли
Опухоли. Этиология, патогенез, морфогенез опухоли Гонорея
Гонорея Кома. Коматозное состояние
Кома. Коматозное состояние Судебная медицина. Организация судебномедицинской службы
Судебная медицина. Организация судебномедицинской службы Организация борьбы с материнской смертностью, борьба с гемморрагическим шоком, инфузионно-трансфузионная терапия
Организация борьбы с материнской смертностью, борьба с гемморрагическим шоком, инфузионно-трансфузионная терапия