- Загальна характеристика родини Iridoviridae

Содержание
- 2. “Назва походить від імен богів веселки у Давній Греції, тому що тканини хворих комах та осад,
- 3. Iridoviridae Вперше виявлені у 1954 р.у личинці багатоніжки Вірус розповсюджений по всьому світі
- 4. Кристалоподібні скупчення в клітинах уражених комах. Така “упаковка” вірусу надає комахам блакитного кольору, що опалесціює.
- 5. Iridoviridae Виявлені у безхребетних та деяких хребетних (не ссавців) 1. Рід Chloriridovirus – великі іридісцентні віруси
- 6. Iridoviridae
- 7. Нуклеокапсид діаметром 125-300 нм, складається з 72 капсомерів. Основний капсидний білок - МСР. Морфологія вірусних часток
- 8. Будова віріону Віріони складаються із трьох концентричних структур: зовнішнього білкового капсиду, середнього ліпідного шару, асоційованого з
- 9. Геном Геном – лінійна 2л ДНК, 150 дo 280 тис.п.о. Наявні термінальні повторів та циклічні перестановки
- 10. Геном Геном містить близько 100 відкритих рамок зчитування на обох ланцюгах ДНК. Високий рівень метилювання ДНК
- 11. Білки У структурі віріону виявляють від 25 до 75 структурних білків, які мають М від 12
- 12. Ліпіди входять до складу внутрішнього шару. Їх вміст становить від 3 до 14% маси віріону. Структура
- 14. Фізико-хімічні властивості Віруси дуже стабільні до умов зовнішнього середовища, поза організмом хазяїна Стабільні в широкому діапазоні
- 15. Реплікація вірусів
- 16. Вірусні частки проникають у середину клітини шляхом ендоцитозу, після цього наступає стадія роздягання. Вірусна ДНК транспортується
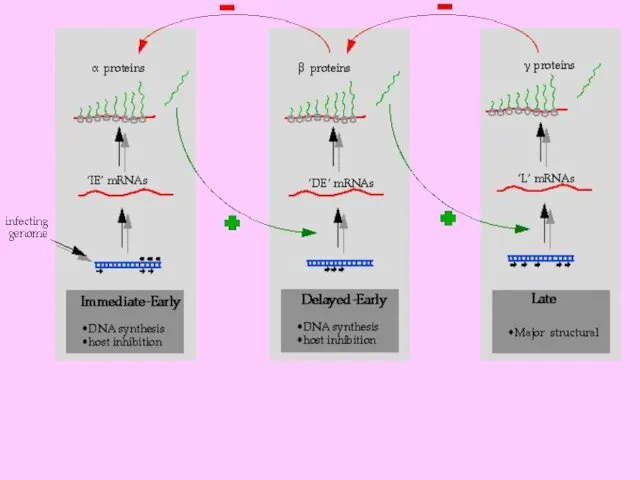
- 17. 1 етап реплікації здійснюється в ядрі інфікованої клітини: синтез копій геному, неповних з 5’-кінця кожного ланцюга
- 19. Можливі типи рекомбінації при реплікації іридовірусів
- 20. Wiseana iridescent virus
- 21. Симптоми хвороб Хронічні або ураження легкого ступеня (лізис, виразки), Збільшення клітин деяких органів. Виявляються проблематично. У
- 22. Патогенез Патогенез – температурозалежний процес. Іридовіруси у чорної мухи Simulium spp. викликають дві форми інфекції: приховану
- 23. Симптоми захворювання 1. Контроль (ін’єкція водою) 2. Інапарантна інфекція 3. Ранні симптоми 4. Пізня стадія інфекції,
- 24. Патогенез Електронномікроскопічне зображення епітелію трахей при ураженні довгоносика Diaprepes abbreviatus IIV-6.
- 25. Патогенез При летальній інфекції комах початковими місцями реплікації вірусів є жирові тіла та гемоцити (що в
- 26. Епідеміологія Передача: Горизонтальна Вертикальна Віруси передаються за допомогою води, на пір’ї та дзьобах птахів, за рахунок
- 27. Передача IIV-6 у D. abbreviatus
- 28. Можливе застосування комахи: біоконтроль У комах вірус був виділений з мошок (Similiidae), хрущика японського (Popillia japonica),
- 29. Хребетні: аквакультура, тваринництво, зоопарки; Передача – контактним шлях (при контакті з хворими тваринами та інфікованими предметами).
- 30. Cимптоми Проявляються на всіх стадіях розвитку амфібій: У пуголовків- зниження активності, асцит, вогнищеві крововиливи У молодих
- 31. Іридовіруси Ranavirus Уражуються жаби, саламандри та інші амфібії. Північно Американський вид тігрової саламандри Ambystoma tigrinum під
- 32. Ranavirus виділений з морського окуня Epinephelus tauvina (Сингапур)
- 33. 1994р.- “Сонні”захворювання морського окуня (SGD), летальність 90%
- 34. Лімфоцистит LDCV-1 викликає хронічну контагіозну хворобу багатьох видів морських та прісноводних риб.
- 36. Іридовірусні захворювання риб
- 37. Географічне розповсюдження
- 38. Діагностика Вибір методу діагностики інфекції, спричиненої іридовірусами визначається епізоотичною ситуацією з врахуваннями клінічних та патолого-морфологічних даних.
- 39. Діагностика Іридовірусні інфекції деяких комах, зокрема комарів та мошок, легко виявляти за характерним райдужним забарвленням тіла
- 41. Скачать презентацию

“Назва походить від імен богів веселки у Давній Греції, тому що
“Назва походить від імен богів веселки у Давній Греції, тому що
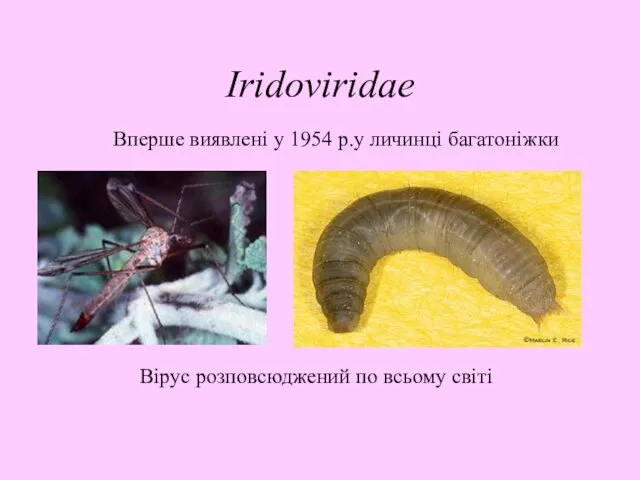
Iridoviridae
Вперше виявлені у 1954 р.у личинці багатоніжки
Вірус розповсюджений по всьому світі
Iridoviridae
Вперше виявлені у 1954 р.у личинці багатоніжки
Вірус розповсюджений по всьому світі
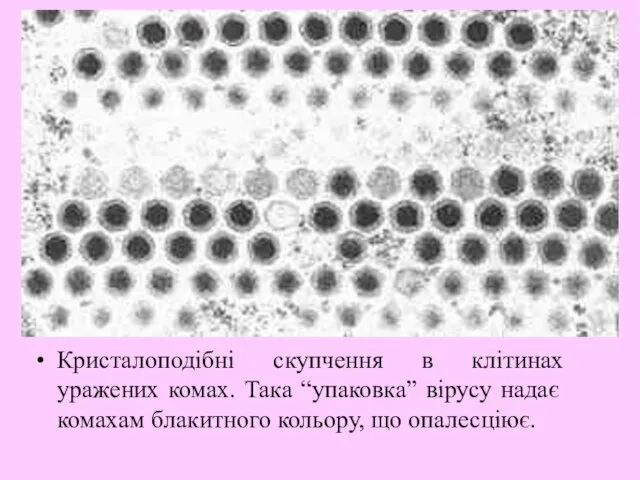
Кристалоподібні скупчення в клітинах уражених комах. Така “упаковка” вірусу надає комахам
Кристалоподібні скупчення в клітинах уражених комах. Така “упаковка” вірусу надає комахам

Iridoviridae
Виявлені у безхребетних та деяких хребетних (не ссавців)
1. Рід Chloriridovirus
Iridoviridae
Виявлені у безхребетних та деяких хребетних (не ссавців)
1. Рід Chloriridovirus
Iridoviridae
Iridoviridae
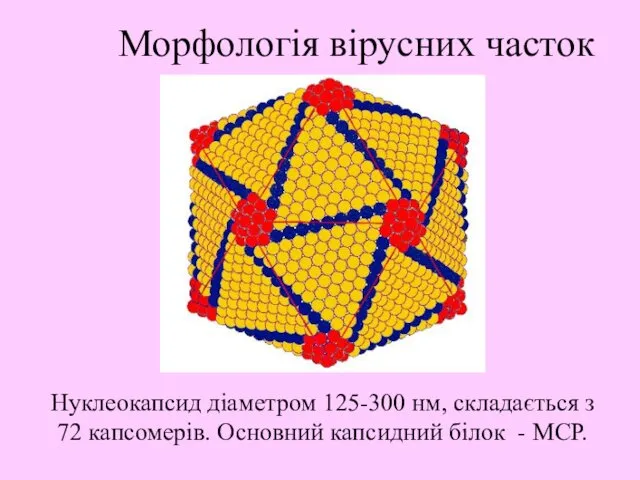
Нуклеокапсид діаметром 125-300 нм, складається з 72 капсомерів. Основний капсидний білок
Нуклеокапсид діаметром 125-300 нм, складається з 72 капсомерів. Основний капсидний білок
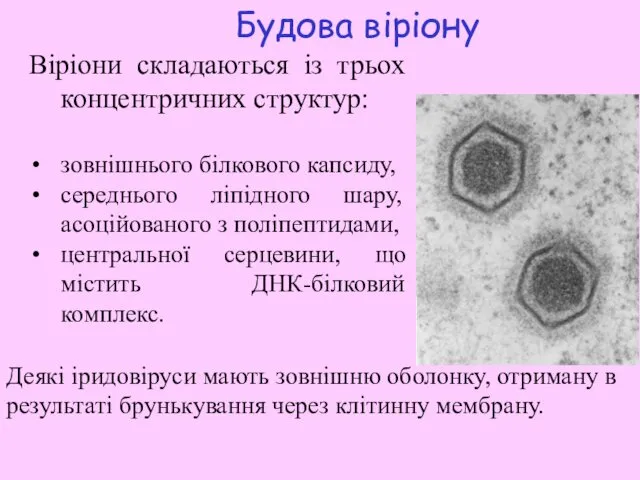
Будова віріону
Віріони складаються із трьох концентричних структур:
зовнішнього білкового капсиду,
середнього ліпідного
Будова віріону
Віріони складаються із трьох концентричних структур:
зовнішнього білкового капсиду,
середнього ліпідного
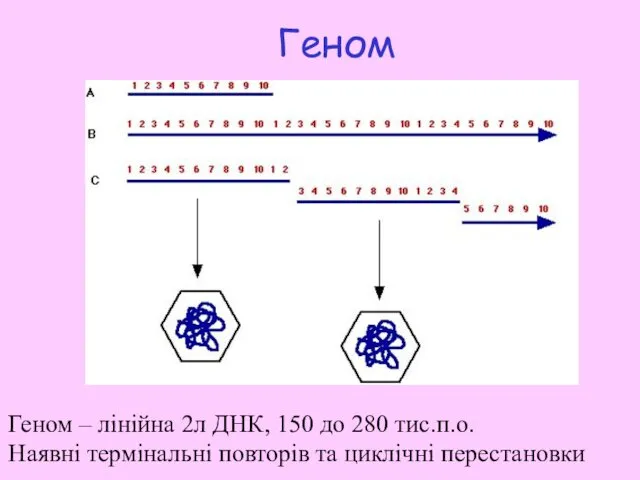
Геном
Геном – лінійна 2л ДНК, 150 дo 280 тис.п.о.
Наявні термінальні
Геном
Геном – лінійна 2л ДНК, 150 дo 280 тис.п.о.
Наявні термінальні

Геном
Геном містить близько 100 відкритих рамок зчитування на обох ланцюгах
Геном
Геном містить близько 100 відкритих рамок зчитування на обох ланцюгах

Білки
У структурі віріону виявляють від 25 до 75 структурних білків,
Білки
У структурі віріону виявляють від 25 до 75 структурних білків,

Ліпіди входять до складу внутрішнього шару. Їх вміст становить від 3
Ліпіди входять до складу внутрішнього шару. Їх вміст становить від 3

Фізико-хімічні властивості
Віруси дуже стабільні до умов зовнішнього середовища, поза організмом хазяїна
Фізико-хімічні властивості
Віруси дуже стабільні до умов зовнішнього середовища, поза організмом хазяїна
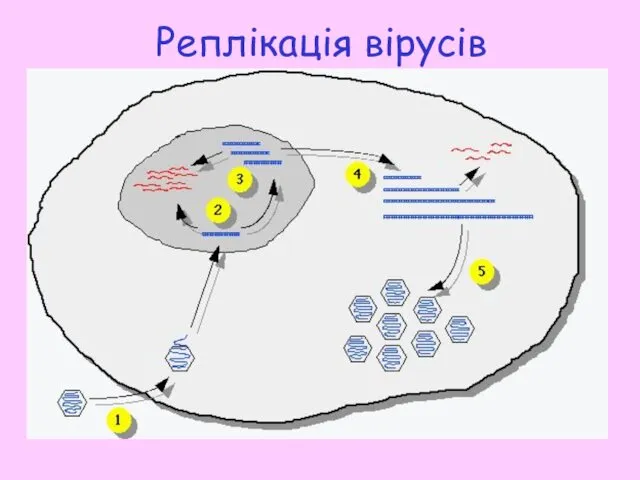
Реплікація вірусів
Реплікація вірусів

Вірусні частки проникають у середину клітини шляхом ендоцитозу, після цього наступає
Вірусні частки проникають у середину клітини шляхом ендоцитозу, після цього наступає

1 етап реплікації здійснюється в ядрі інфікованої клітини: синтез копій геному,
1 етап реплікації здійснюється в ядрі інфікованої клітини: синтез копій геному,
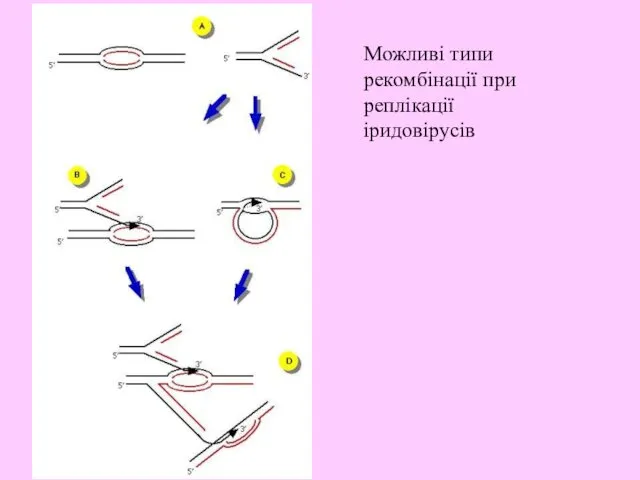
Можливі типи рекомбінації при реплікації іридовірусів
Можливі типи рекомбінації при реплікації іридовірусів
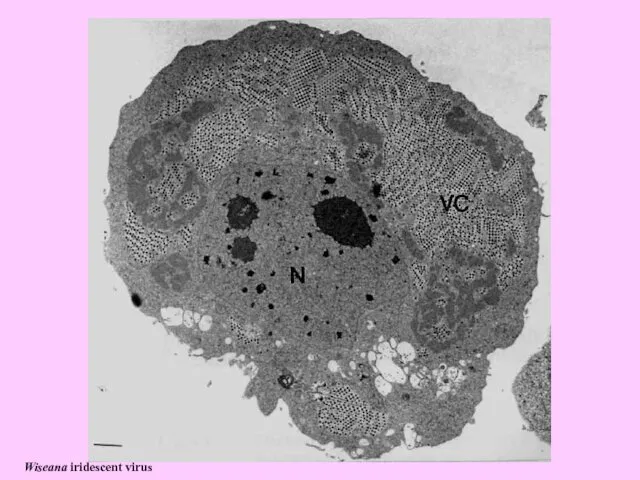
Wiseana iridescent virus
Wiseana iridescent virus

Симптоми хвороб
Хронічні або ураження легкого ступеня (лізис, виразки),
Збільшення клітин деяких
Симптоми хвороб
Хронічні або ураження легкого ступеня (лізис, виразки),
Збільшення клітин деяких

Патогенез
Патогенез – температурозалежний процес. Іридовіруси у чорної мухи Simulium spp. викликають
Патогенез
Патогенез – температурозалежний процес. Іридовіруси у чорної мухи Simulium spp. викликають

Симптоми захворювання
1. Контроль (ін’єкція водою)
2. Інапарантна інфекція
3. Ранні симптоми
4. Пізня стадія
Симптоми захворювання
1. Контроль (ін’єкція водою)
2. Інапарантна інфекція
3. Ранні симптоми
4. Пізня стадія
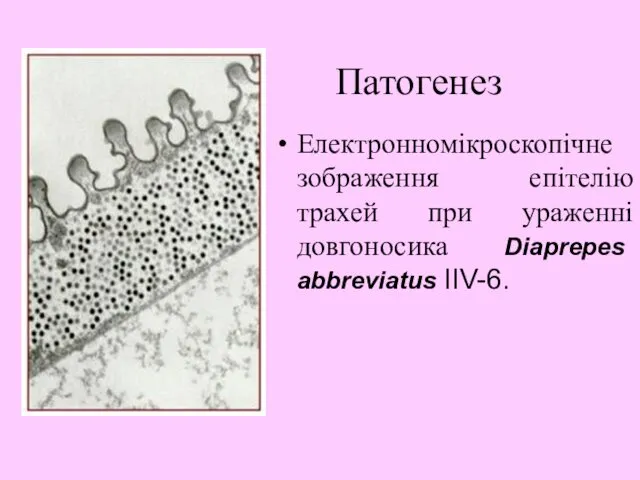
Патогенез
Електронномікроскопічне зображення епітелію трахей при ураженні довгоносика Diaprepes abbreviatus IIV-6.
Патогенез
Електронномікроскопічне зображення епітелію трахей при ураженні довгоносика Diaprepes abbreviatus IIV-6.

Патогенез
При летальній інфекції комах початковими місцями реплікації вірусів є жирові
Патогенез
При летальній інфекції комах початковими місцями реплікації вірусів є жирові

Епідеміологія
Передача:
Горизонтальна
Вертикальна
Віруси передаються за допомогою води,
на пір’ї та дзьобах птахів,
Епідеміологія
Передача:
Горизонтальна
Вертикальна
Віруси передаються за допомогою води,
на пір’ї та дзьобах птахів,
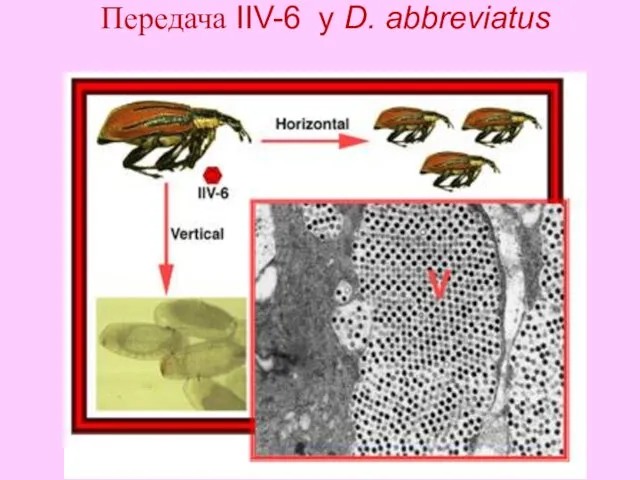
Передача IIV-6 у D. abbreviatus
Передача IIV-6 у D. abbreviatus

Можливе застосування
комахи: біоконтроль
У комах вірус був виділений з мошок (Similiidae),
Можливе застосування
комахи: біоконтроль
У комах вірус був виділений з мошок (Similiidae),

Хребетні: аквакультура, тваринництво, зоопарки;
Передача – контактним шлях (при контакті з хворими
Хребетні: аквакультура, тваринництво, зоопарки;
Передача – контактним шлях (при контакті з хворими

Cимптоми
Проявляються на всіх стадіях розвитку амфібій:
У пуголовків- зниження активності, асцит, вогнищеві
Cимптоми
Проявляються на всіх стадіях розвитку амфібій:
У пуголовків- зниження активності, асцит, вогнищеві

Іридовіруси
Ranavirus
Уражуються жаби, саламандри та інші амфібії.
Північно Американський вид тігрової
Іридовіруси
Ranavirus
Уражуються жаби, саламандри та інші амфібії.
Північно Американський вид тігрової

Ranavirus виділений з морського окуня Epinephelus tauvina (Сингапур)
Ranavirus виділений з морського окуня Epinephelus tauvina (Сингапур)
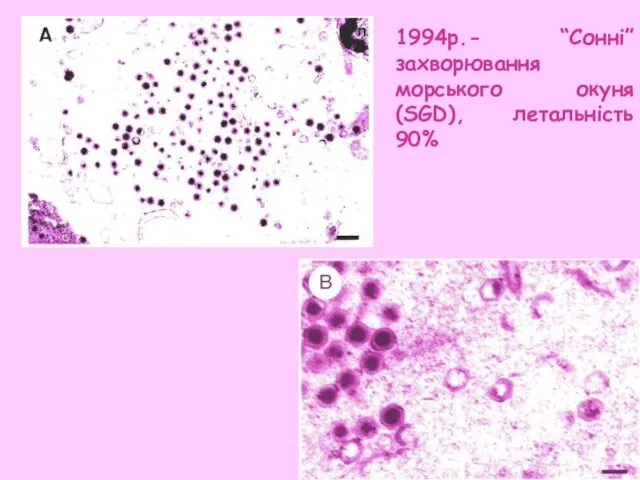
1994р.- “Сонні”захворювання морського окуня (SGD), летальність 90%
1994р.- “Сонні”захворювання морського окуня (SGD), летальність 90%
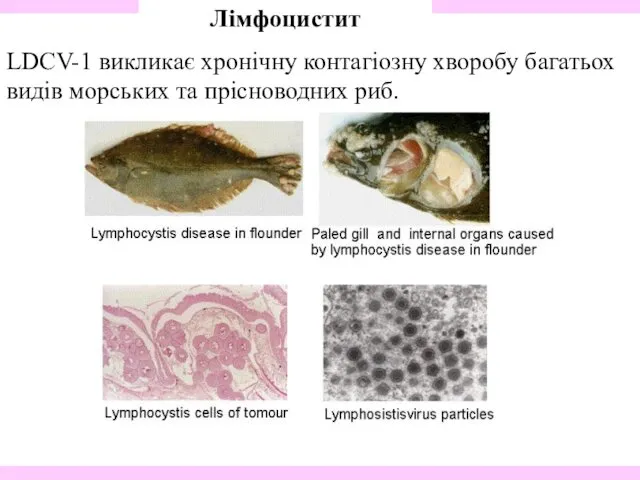
Лімфоцистит
LDCV-1 викликає хронічну контагіозну хворобу багатьох видів морських та прісноводних
Лімфоцистит
LDCV-1 викликає хронічну контагіозну хворобу багатьох видів морських та прісноводних

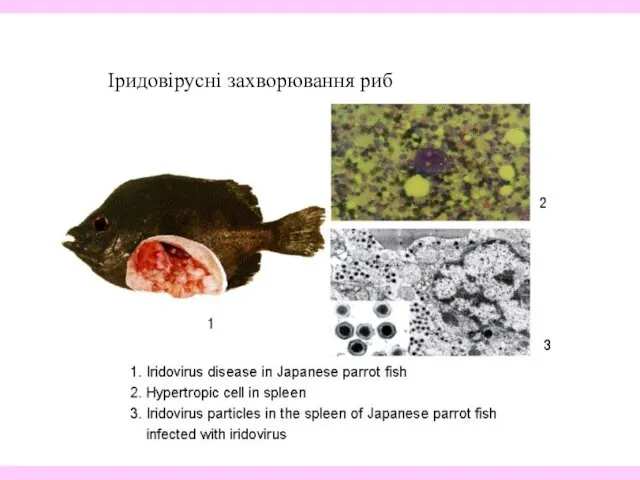
Іридовірусні захворювання риб
Іридовірусні захворювання риб
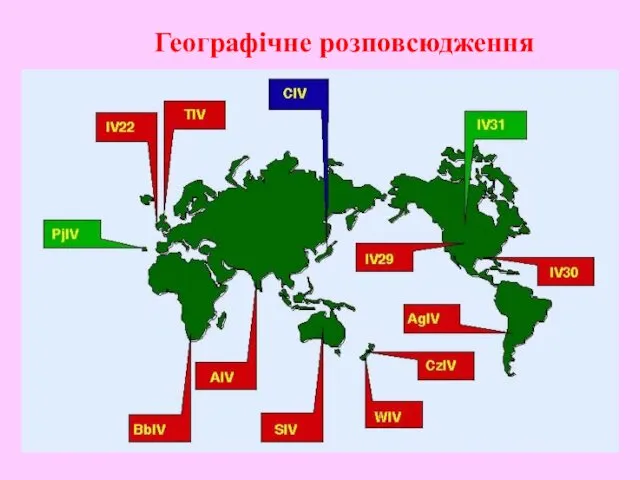
Географічне розповсюдження
Географічне розповсюдження

Діагностика
Вибір методу діагностики інфекції, спричиненої іридовірусами визначається епізоотичною ситуацією з врахуваннями
Діагностика
Вибір методу діагностики інфекції, спричиненої іридовірусами визначається епізоотичною ситуацією з врахуваннями

Діагностика
Іридовірусні інфекції деяких комах, зокрема комарів та мошок, легко виявляти
Діагностика
Іридовірусні інфекції деяких комах, зокрема комарів та мошок, легко виявляти
 Эпидемиологическая ситуация по ВИЧ-инфекции в Курганской области
Эпидемиологическая ситуация по ВИЧ-инфекции в Курганской области Клиническое обследование. Clinical examination. Лекция 2
Клиническое обследование. Clinical examination. Лекция 2 Современные методы диагностики и хирургического лечения язвенной болезни желудка и 12-перстной кишки
Современные методы диагностики и хирургического лечения язвенной болезни желудка и 12-перстной кишки Этилметилгидроксипиридина сукцинат
Этилметилгидроксипиридина сукцинат Планирование беременности
Планирование беременности Деятельность телефонов доверия
Деятельность телефонов доверия Тіс жегі қуысын егеп-тазалау үшін жоғарғы энергетикалық лазерлерді қолдану
Тіс жегі қуысын егеп-тазалау үшін жоғарғы энергетикалық лазерлерді қолдану Презентация по медицине Фенилкетонурия ФКУ финилпировиноградная олигофрения болезнь Фёллинга
Презентация по медицине Фенилкетонурия ФКУ финилпировиноградная олигофрения болезнь Фёллинга  Бетаблокаторы в детской кардиологической практике
Бетаблокаторы в детской кардиологической практике Хирургия желудочно-кишечного тракта. Кишечные швы
Хирургия желудочно-кишечного тракта. Кишечные швы Оценка новорожденного ребенка сразу после рождения и принципы ухода за ним
Оценка новорожденного ребенка сразу после рождения и принципы ухода за ним Клинико-педагогическая классификация речевых нарушений. Виды речевых нарушений у детей
Клинико-педагогическая классификация речевых нарушений. Виды речевых нарушений у детей Областное родительское собрание «Профилактика потребления несовершеннолетними наркотических средств и психотропных веществ»
Областное родительское собрание «Профилактика потребления несовершеннолетними наркотических средств и психотропных веществ» Электр қозғалтқыштар, түрлері, жұмыс жасау принципі
Электр қозғалтқыштар, түрлері, жұмыс жасау принципі Особенности течения эпидемического процесса. Количественная и качественная характеристики эпидемического процесса
Особенности течения эпидемического процесса. Количественная и качественная характеристики эпидемического процесса Тотальное и субтотальное затемнение. Рентгенография грудной клетки
Тотальное и субтотальное затемнение. Рентгенография грудной клетки Дисциркуляторная энцефалопатия
Дисциркуляторная энцефалопатия Босануды жүргізу, интранатальды кезең
Босануды жүргізу, интранатальды кезең Неотложная помощь при остром отеке Квинке
Неотложная помощь при остром отеке Квинке СМП при выполнении инъекций и введении внутрисосудистых катетеров. Профессиональные риски и безопасность медицинских работников
СМП при выполнении инъекций и введении внутрисосудистых катетеров. Профессиональные риски и безопасность медицинских работников Воспаление. Воспалительная реакция
Воспаление. Воспалительная реакция Мектеп бітірушілердің психологиялық-физиологиялық жағдайын ескере отырып қобалжу деңгейін төмендету
Мектеп бітірушілердің психологиялық-физиологиялық жағдайын ескере отырып қобалжу деңгейін төмендету Л - 8 ( 2.4.) Питание
Л - 8 ( 2.4.) Питание Катетеризация мочевого пузыря
Катетеризация мочевого пузыря Диагностика ревматизма
Диагностика ревматизма Гипертоническая болезнь и Трансфер Фактор. Обзор научных исследований по применению
Гипертоническая болезнь и Трансфер Фактор. Обзор научных исследований по применению Методическая разработка раздела «Соотношения между сторонами и углами треугольника»
Методическая разработка раздела «Соотношения между сторонами и углами треугольника» Бүйрек алмастырушы ем қағидалары
Бүйрек алмастырушы ем қағидалары