Раздражимость и возбудимость клеток и тканей. Методы исследования электровозбудимых мембран. Потенциал покоя и механизм его форм
- Раздражимость и возбудимость клеток и тканей. Методы исследования электровозбудимых мембран. Потенциал покоя и механизм его форм

Содержание
- 2. РАЗДРАЖИМОСТЬ – ОБЩЕЕ СВОЙСТВО ВСЕХ ЖИВЫХ ОРГАНИЗМОВ
- 3. ВОЗБУДИМЫЕ ТКАНИ
- 4. Мера возбудимости – порог раздражения, т.е. минимальная сила раздражителя, вызывающая ответ. ВОЗБУЖДЕНИЕ – сложный биологический процесс,
- 5. РАЗДРАЖИТЕЛИ ПОДПОРОГОВЫЕ СВЕРХПОРОГОВЫЕ ПОРОГОВЫЕ
- 6. РАЗДРАЖИТЕЛИ ФИЗИЧЕСКИЕ (механические, звуковые, световые, температурные, электрические) ХИМИЧЕСКИЕ (щелочи, кислоты, гормоны, продукты обмена веществ) ФИЗИКО-ХИМИЧЕСКИЕ (изменения
- 7. РАЗДРАЖИТЕЛИ НЕАДЕКВАТНЫЕ АДЕКВАТНЫЕ
- 8. СООТНОШЕНИЕ МЕЖДУ ПОРОГОВОЙ СИЛОЙ РАЗДРАЖЕНИЯ И ЕГО ДЛИТЕЛЬНОСТЬЮ
- 9. КРИВАЯ «СИЛА – ДЛИТЕЛЬНОСТЬ» OA – реобаза ОС – полезное время OD – 2 реобазы OF
- 11. МЕТОДЫ ИССЛЕДОВАНИЯ ЭЛЕКТРОВОЗБУДИМЫХ ТКАНЕЙ
- 12. ПРОГРЕСС В ИССЛЕДОВАНИИ БИОПОТЕНЦИАЛОВ ОБЕСПЕЧЕН РАЗРАБОТКОЙ МИКРОЭЛЕКТРОДНОГО МЕТОДА ВНУТРИКЛЕТОЧНОГО ОТВЕДЕНИЯ ПОТЕНЦИАЛОВ СОЗДАНИЕ СПЕЦИАЛЬНЫХ УСИЛИТЕЛЕЙ БИОПОТЕНЦИАЛОВ ВЫБОР
- 13. ДИАМЕТР гигантского аксона – 0,5 – 1 мм
- 14. ОСНОВНЫЕ ЭЛЕМЕНТЫ УСТАНОВКИ ДЛЯ ИЗУЧЕНИЯ ВОЗБУДИМЫХ КЛЕТОК И ТКАНЕЙ 1) электроды для регистрации и стимуляции; 2)
- 15. a - внутриклеточное раздражение и отведение потенциалов гигантского аксона кальмара при коаксиальном введении электродов . б
- 16. НЕПОЛЯРИЗУЮЩИЕСЯ ЭЛЕКТРОДЫ: хлорсеребряный электрод Благодаря стабильности потенциала и простоте конструкции является одним из наиболее часто употребляемых
- 17. Микроэлектроды введены в 1946 американскими учёными Р. Джерардом и Дж. Лингом. Применяются для отведения электрических потенциалов
- 19. Внутриклеточная регистрация трансмембранных потенциалов и электростимуляция клеточной мембраны А — схема установки для изучения электрических характеристик
- 20. В лабораторных исследованиях используются металлические микроэлектроды с диаметром кончика порядка 1 мкм, стеклянные микропипетки с диаметром
- 21. РЕГИСТРАЦИЯ МП НЕРВНОЙ КЛЕТКИ А) путем введения микроэлектрода; Б) путем введения микроэлектрода внутрь аксона
- 23. Метод пэтч-кламп введен в лабораторную практику Э. Неером и Б. Сакманом в 1976 г.
- 24. Клеточная мембрана формирует очень плотный контакт с поверхностью кончика микроэлектрода. Между стеклом и мембранным фрагментом возникает
- 26. ИЗМЕРЕНИЯ НА ПРИКРЕПЛЕННОЙ, НО НЕПОВРЕЖДЕННОЙ КЛЕТКЕ ИЗМЕРЕНИЯ НА ЦЕЛОЙ КЛЕТКЕ ПРИ РАЗРУШЕНИИ МЕМБРАНЫ В КОНЧИКЕ МИКРОПИПЕТКИ
- 28. Мембранная теория возбуждения: при раздражении возбудимой клетки в её поверхностной мембране происходит молекулярная перестройка, которая приводит
- 29. Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю. Бернштейном (1902) ПОЛЯРИЗАЦИЯ КЛЕТОЧНОЙ МЕМБРАНЫ В ПОКОЕ
- 30. Развитие мембранной теории возбуждения получило в трудах английских учёных: П. Бойла и Э. Конуэя (1941), А.
- 31. Регистрация мембранного потенциала покоя А — микроэлектрод 1 еще не введен в нервное волокно; луч осциллографа
- 32. Потенциал покоя, механизм его формирования
- 33. Предположение Бернштейна: поверхностная мембрана возбудимой клетки в покое обладает избирательной проницаемостью: ионы K+ проходят через неё
- 34. ДИФФУЗИОННЫЙ ПОТЕНЦИАЛ Na+ Na+ Na+ Na+ Cl- Cl- Cl- + - ПОДВИЖНОСТЬ ИОНОВ ХЛОРА (v) ВЫШЕ,
- 35. МЕМБРАННЫЙ ПОТЕНЦИАЛ Cl- Cl- К+ К+ МЕМБРАНА НЕ ПРОНИЦАЕМА ДЛЯ ИОНОВ ХЛОРА - +
- 36. ЭЛЕКТРОХИМИЧЕСКИЙ ПОТЕНЦИАЛ УСЛОВИЕ РАВНОВЕСИЯ РАВНОВЕСНЫЕ ПОТЕНЦИАЛЫ: ENa=+35…+65 мВ EK = -70…-100 мВ
- 37. Возникновение электрохимического равновесия на полупроницаемой мембране. Диффузионное давление (1) в равновесии с противоположной электростатической силой (2),
- 39. Зависимость мембранного потенциала (МП) нервного волокна каракатицы от наружной концентрации К+(1) и величина МП, рассчитанная по
- 40. Зависимость величины ПП от [K+]е (расчетная и экспериментальная кривые). По оси абсцисс – содержание калия во
- 43. состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов (по сравнению с их общим
- 44. УРАВНЕНИЕ НЕРНСТА Здесь и далее индекс «е» относится к внешним концентрациям ионов, а индекс «i» -
- 46. ПП гигантского аксона кальмара ( - 70 мВ) близок к его Ек ( - 75 мВ),
- 47. Итоговая величина ПП, обусловленного переносом многих ионов может быть достаточно точно рассчитана по формуле Гольдмана.
- 48. УРАВНЕНИЕ ГОЛЬДМАНА PK : PNa: PCl = 1 : 0,04 : 0,45 Соотношение проницаемостей потенциалообразующих ионов
- 49. Вклад активного транспорта в формирование потенциала покоя
- 50. ПП складывается из Еконц и Енас
- 51. РОЛЬ Na/K НАСОСА В ГЕНЕРАЦИИ ПП Поддержание высокой концентрации К+ внутри клетки, что обеспечивает постоянство величины
- 52. Электротонический потенциал и локальный ответ, их сходство и различие
- 53. Схема регистрации мембранного потенциала клетки (А); мембранный потенциал клетки в состоянии покоя и его возможные изменения
- 54. Изменение мембранного потенциала клетки (А) при действии электрического тока различной силы (Б):
- 55. ЭЛЕКТРОТОНИЧЕСКИЕ ПОТЕНЦИАЛЫ И ЛОКАЛЬНЫЕ ОТВЕТЫ
- 56. ВОЛЬТ-АМПЕРНАЯ ХАРАКТЕРИСТИКА ЭЛЕКТРОТОНИЧЕСКОГО ПОТЕНЦИАЛА
- 58. Скачать презентацию

РАЗДРАЖИМОСТЬ – ОБЩЕЕ СВОЙСТВО ВСЕХ ЖИВЫХ ОРГАНИЗМОВ
РАЗДРАЖИМОСТЬ – ОБЩЕЕ СВОЙСТВО ВСЕХ ЖИВЫХ ОРГАНИЗМОВ
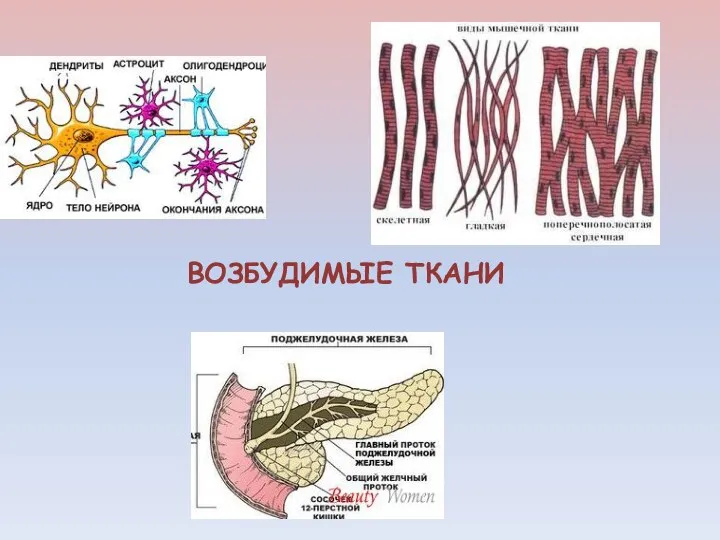
ВОЗБУДИМЫЕ ТКАНИ
ВОЗБУДИМЫЕ ТКАНИ

Мера возбудимости – порог раздражения, т.е. минимальная сила раздражителя, вызывающая ответ.
ВОЗБУЖДЕНИЕ
Мера возбудимости – порог раздражения, т.е. минимальная сила раздражителя, вызывающая ответ.
ВОЗБУЖДЕНИЕ
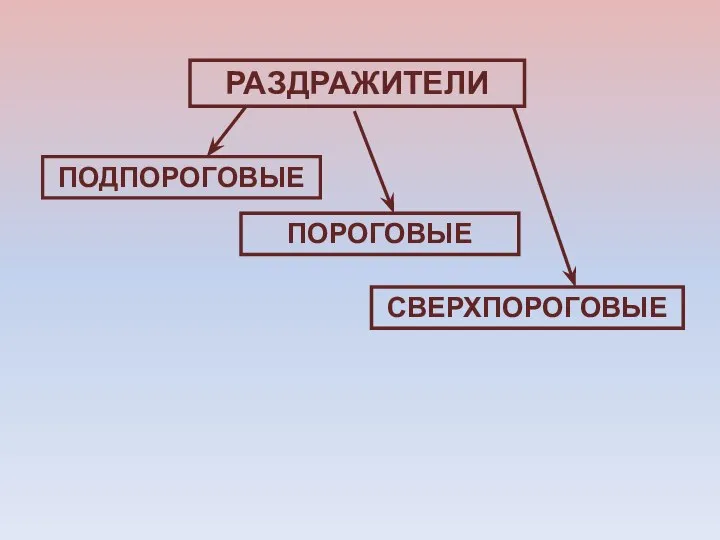
РАЗДРАЖИТЕЛИ
ПОДПОРОГОВЫЕ
СВЕРХПОРОГОВЫЕ
ПОРОГОВЫЕ
РАЗДРАЖИТЕЛИ
ПОДПОРОГОВЫЕ
СВЕРХПОРОГОВЫЕ
ПОРОГОВЫЕ

РАЗДРАЖИТЕЛИ
ФИЗИЧЕСКИЕ (механические, звуковые, световые, температурные, электрические)
ХИМИЧЕСКИЕ (щелочи, кислоты, гормоны, продукты обмена
РАЗДРАЖИТЕЛИ
ФИЗИЧЕСКИЕ (механические, звуковые, световые, температурные, электрические)
ХИМИЧЕСКИЕ (щелочи, кислоты, гормоны, продукты обмена
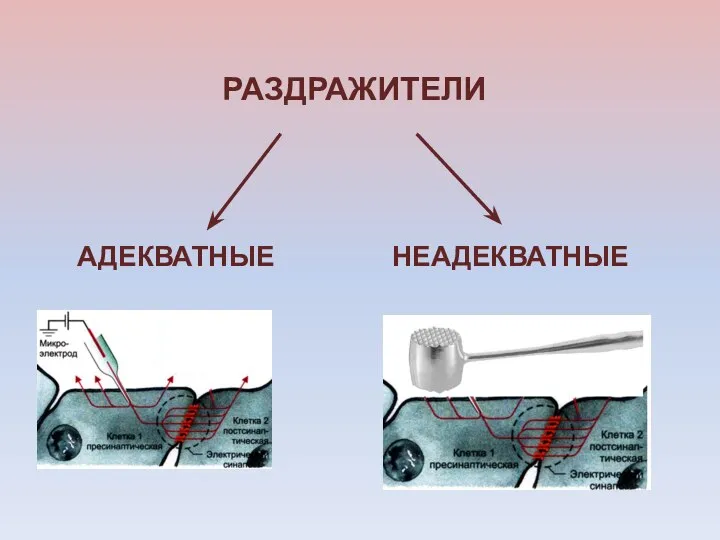
РАЗДРАЖИТЕЛИ
НЕАДЕКВАТНЫЕ
АДЕКВАТНЫЕ
РАЗДРАЖИТЕЛИ
НЕАДЕКВАТНЫЕ
АДЕКВАТНЫЕ

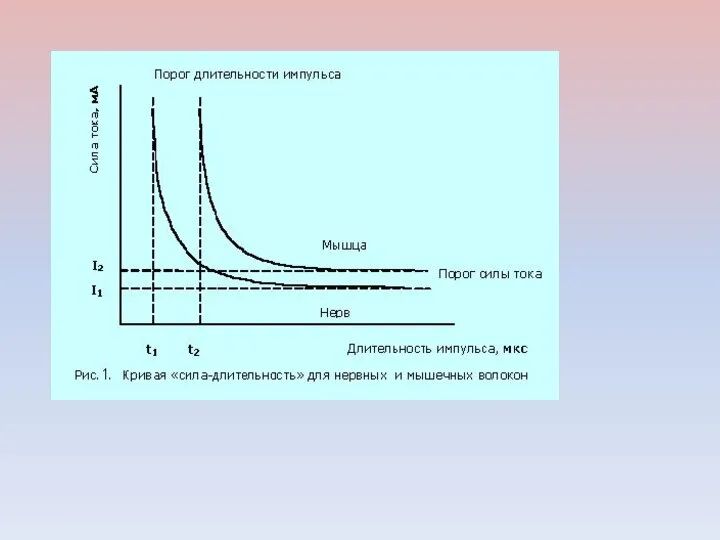
СООТНОШЕНИЕ МЕЖДУ ПОРОГОВОЙ СИЛОЙ РАЗДРАЖЕНИЯ И ЕГО ДЛИТЕЛЬНОСТЬЮ
СООТНОШЕНИЕ МЕЖДУ ПОРОГОВОЙ СИЛОЙ РАЗДРАЖЕНИЯ И ЕГО ДЛИТЕЛЬНОСТЬЮ
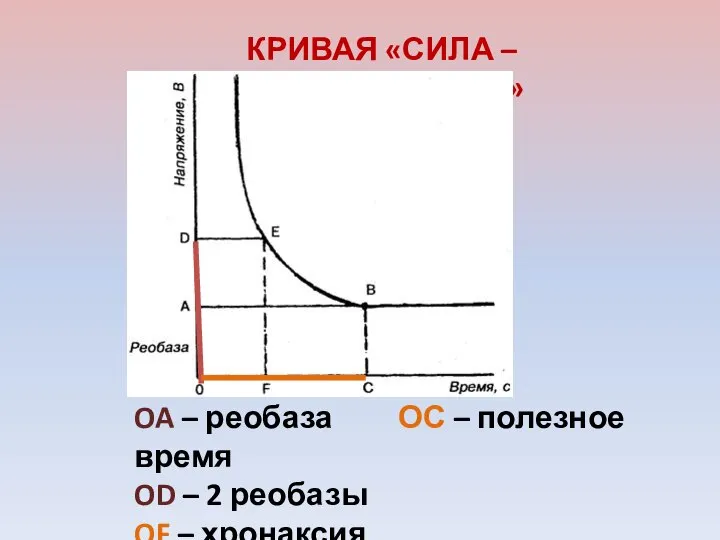
КРИВАЯ «СИЛА – ДЛИТЕЛЬНОСТЬ»
OA – реобаза ОС – полезное время
OD
КРИВАЯ «СИЛА – ДЛИТЕЛЬНОСТЬ»
OA – реобаза ОС – полезное время
OD

МЕТОДЫ ИССЛЕДОВАНИЯ ЭЛЕКТРОВОЗБУДИМЫХ ТКАНЕЙ
МЕТОДЫ ИССЛЕДОВАНИЯ ЭЛЕКТРОВОЗБУДИМЫХ ТКАНЕЙ

ПРОГРЕСС В ИССЛЕДОВАНИИ БИОПОТЕНЦИАЛОВ ОБЕСПЕЧЕН
РАЗРАБОТКОЙ МИКРОЭЛЕКТРОДНОГО МЕТОДА ВНУТРИКЛЕТОЧНОГО ОТВЕДЕНИЯ ПОТЕНЦИАЛОВ
СОЗДАНИЕ СПЕЦИАЛЬНЫХ
ПРОГРЕСС В ИССЛЕДОВАНИИ БИОПОТЕНЦИАЛОВ ОБЕСПЕЧЕН
РАЗРАБОТКОЙ МИКРОЭЛЕКТРОДНОГО МЕТОДА ВНУТРИКЛЕТОЧНОГО ОТВЕДЕНИЯ ПОТЕНЦИАЛОВ
СОЗДАНИЕ СПЕЦИАЛЬНЫХ

ДИАМЕТР гигантского аксона – 0,5 – 1 мм
ДИАМЕТР гигантского аксона – 0,5 – 1 мм

ОСНОВНЫЕ ЭЛЕМЕНТЫ УСТАНОВКИ ДЛЯ ИЗУЧЕНИЯ ВОЗБУДИМЫХ КЛЕТОК И ТКАНЕЙ
1) электроды для
ОСНОВНЫЕ ЭЛЕМЕНТЫ УСТАНОВКИ ДЛЯ ИЗУЧЕНИЯ ВОЗБУДИМЫХ КЛЕТОК И ТКАНЕЙ
1) электроды для
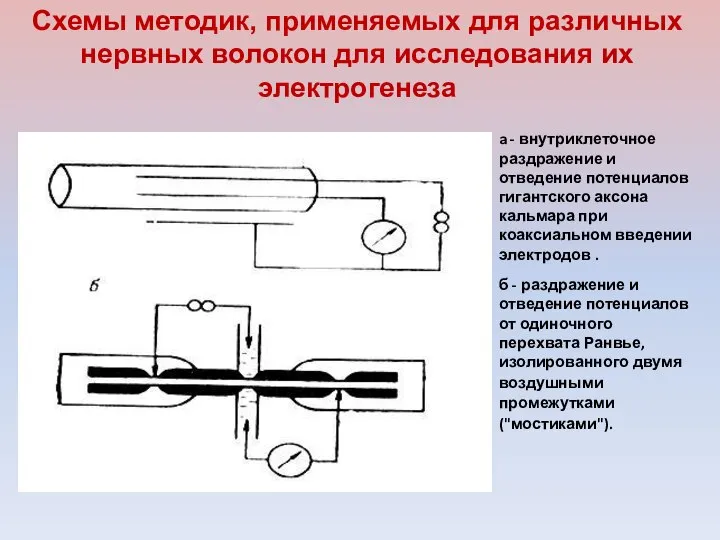
a - внутриклеточное раздражение и отведение потенциалов гигантского аксона кальмара при
a - внутриклеточное раздражение и отведение потенциалов гигантского аксона кальмара при

НЕПОЛЯРИЗУЮЩИЕСЯ ЭЛЕКТРОДЫ: хлорсеребряный электрод
Благодаря стабильности потенциала и простоте конструкции является одним
НЕПОЛЯРИЗУЮЩИЕСЯ ЭЛЕКТРОДЫ: хлорсеребряный электрод
Благодаря стабильности потенциала и простоте конструкции является одним
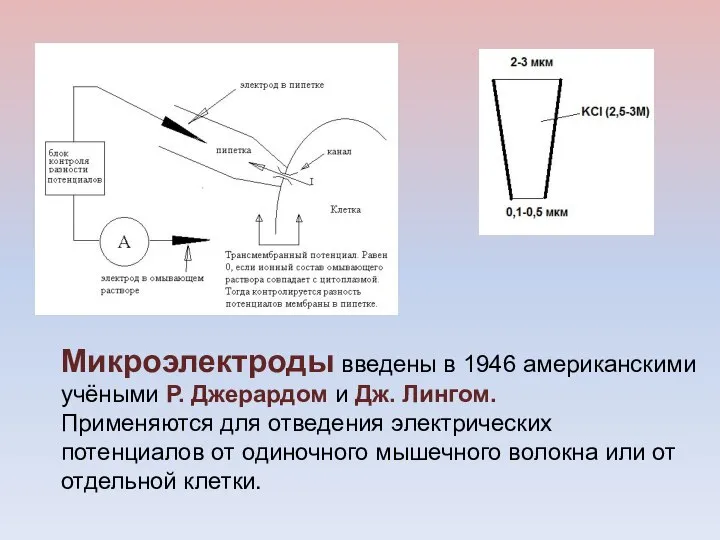
Микроэлектроды введены в 1946 американскими учёными Р. Джерардом и Дж. Лингом.
Микроэлектроды введены в 1946 американскими учёными Р. Джерардом и Дж. Лингом.
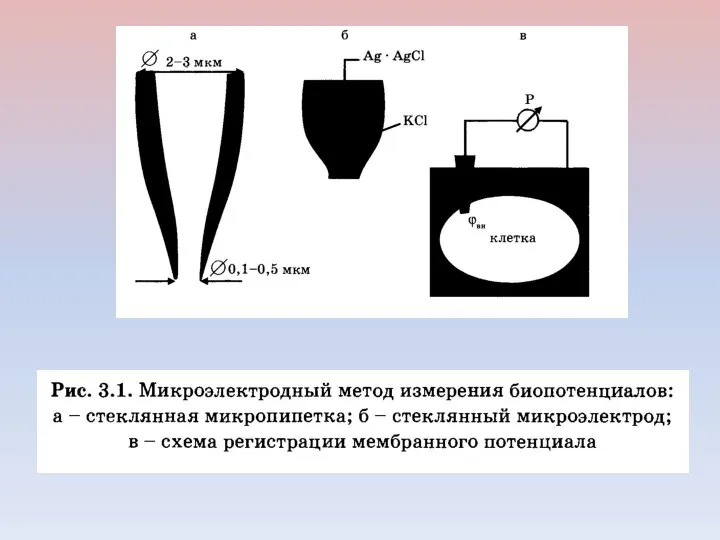
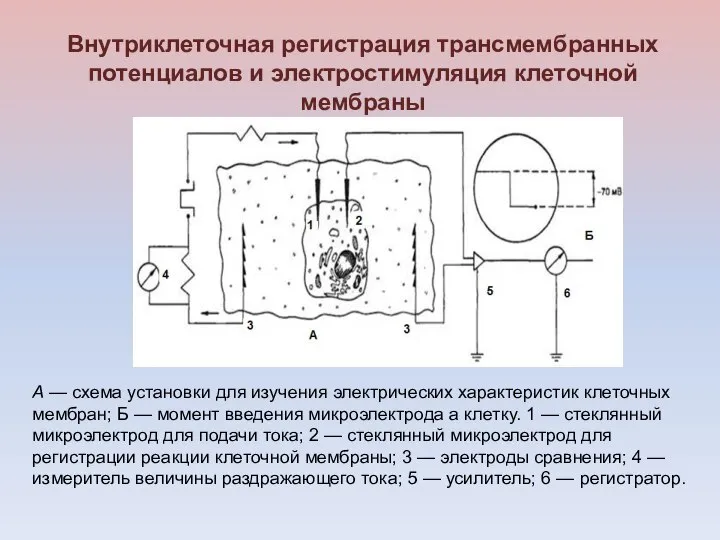
Внутриклеточная регистрация трансмембранных потенциалов и электростимуляция клеточной мембраны
А — схема установки
Внутриклеточная регистрация трансмембранных потенциалов и электростимуляция клеточной мембраны
А — схема установки

В лабораторных исследованиях используются
металлические микроэлектроды с диаметром кончика порядка 1
металлические микроэлектроды с диаметром кончика порядка 1
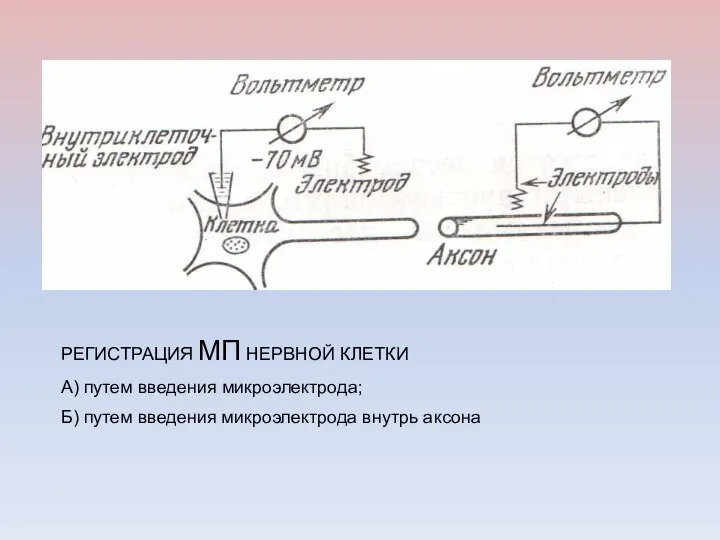
РЕГИСТРАЦИЯ МП НЕРВНОЙ КЛЕТКИ
А) путем введения микроэлектрода;
Б) путем введения микроэлектрода
РЕГИСТРАЦИЯ МП НЕРВНОЙ КЛЕТКИ
А) путем введения микроэлектрода;
Б) путем введения микроэлектрода
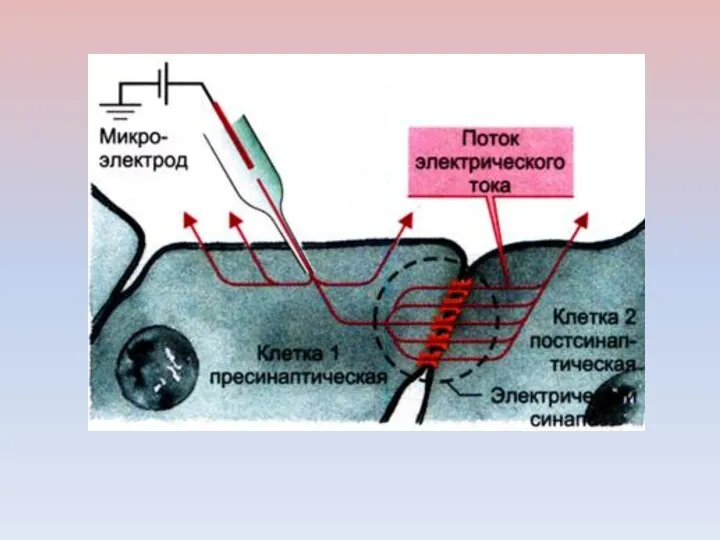

Метод пэтч-кламп введен в лабораторную практику Э. Неером и Б. Сакманом
Метод пэтч-кламп введен в лабораторную практику Э. Неером и Б. Сакманом

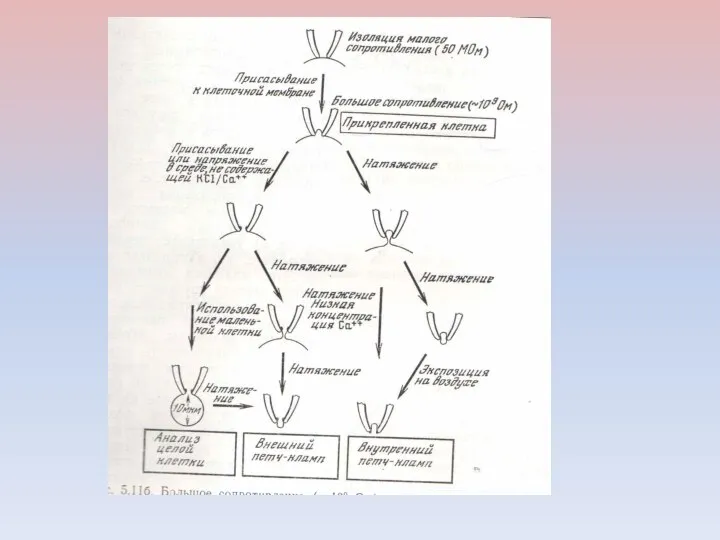
Клеточная мембрана формирует очень плотный контакт с поверхностью кончика микроэлектрода. Между
Клеточная мембрана формирует очень плотный контакт с поверхностью кончика микроэлектрода. Между

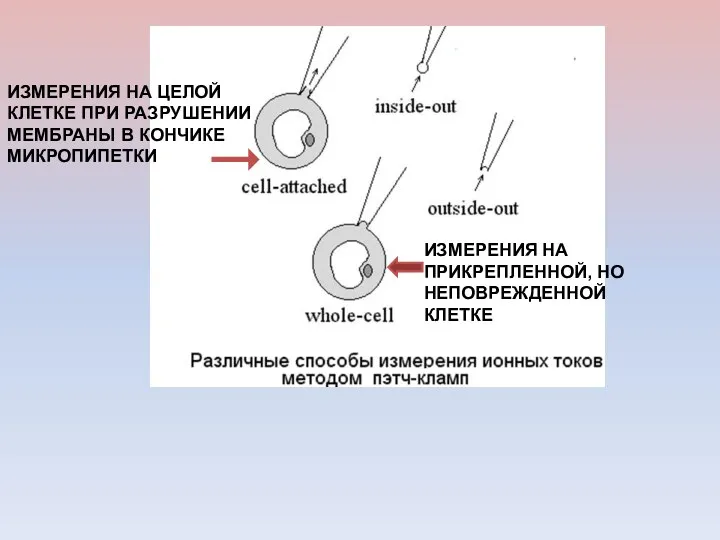
ИЗМЕРЕНИЯ НА ПРИКРЕПЛЕННОЙ, НО НЕПОВРЕЖДЕННОЙ КЛЕТКЕ
ИЗМЕРЕНИЯ НА ЦЕЛОЙ КЛЕТКЕ ПРИ РАЗРУШЕНИИ
ИЗМЕРЕНИЯ НА ПРИКРЕПЛЕННОЙ, НО НЕПОВРЕЖДЕННОЙ КЛЕТКЕ
ИЗМЕРЕНИЯ НА ЦЕЛОЙ КЛЕТКЕ ПРИ РАЗРУШЕНИИ

Мембранная теория возбуждения:
при раздражении возбудимой клетки в её поверхностной мембране
Мембранная теория возбуждения:
при раздражении возбудимой клетки в её поверхностной мембране
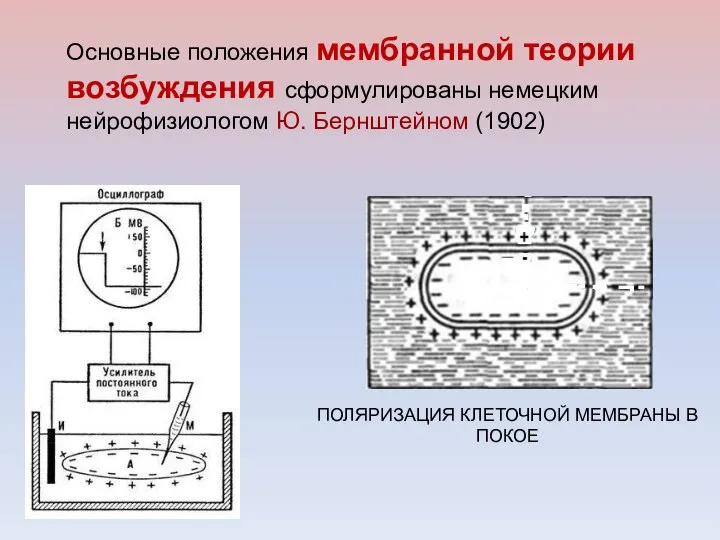
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю. Бернштейном (1902)
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю. Бернштейном (1902)

Развитие мембранной теории возбуждения получило в трудах английских учёных: П. Бойла
Развитие мембранной теории возбуждения получило в трудах английских учёных: П. Бойла
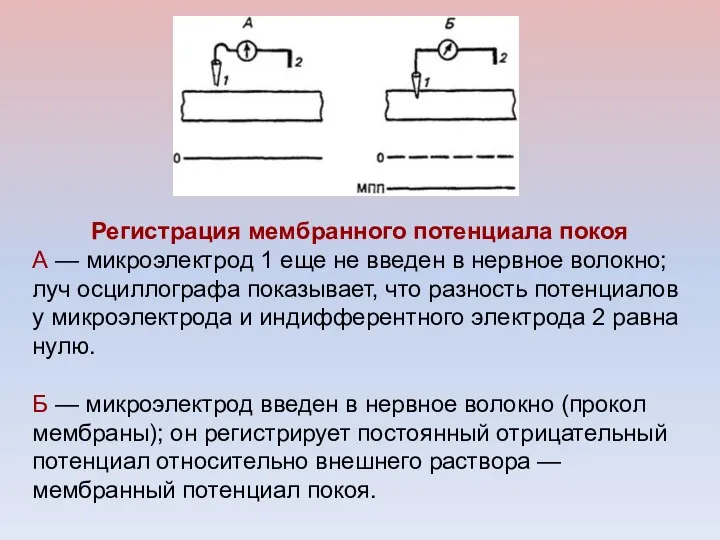
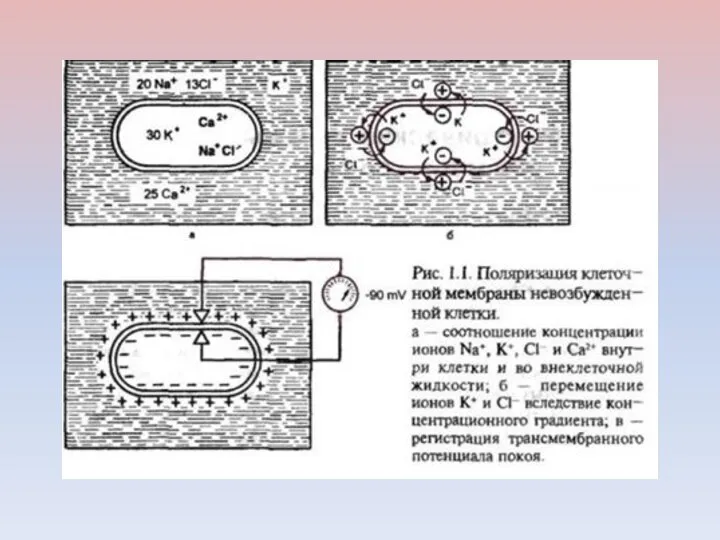
Регистрация мембранного потенциала покоя
А — микроэлектрод 1 еще не введен в
Регистрация мембранного потенциала покоя
А — микроэлектрод 1 еще не введен в

Потенциал покоя, механизм его формирования
Потенциал покоя, механизм его формирования

Предположение Бернштейна:
поверхностная мембрана возбудимой клетки в покое обладает избирательной проницаемостью: ионы
Предположение Бернштейна:
поверхностная мембрана возбудимой клетки в покое обладает избирательной проницаемостью: ионы
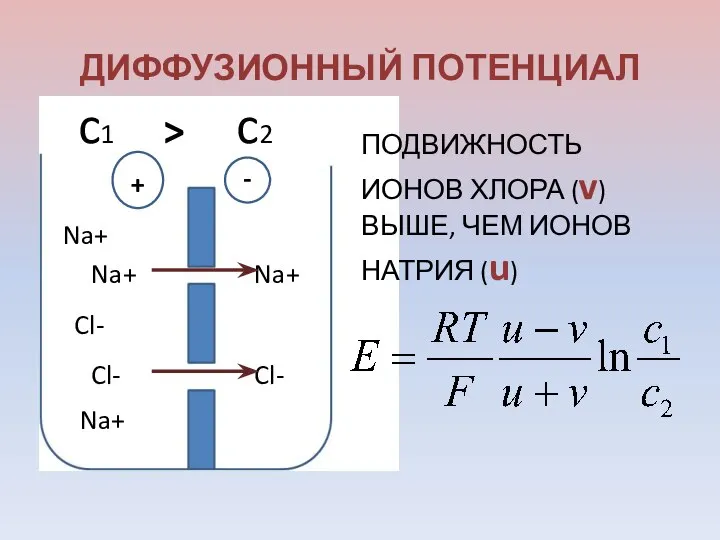
ДИФФУЗИОННЫЙ ПОТЕНЦИАЛ
Na+
Na+
Na+
Na+
Cl-
Cl-
Cl-
+
-
ПОДВИЖНОСТЬ ИОНОВ ХЛОРА (v) ВЫШЕ, ЧЕМ ИОНОВ НАТРИЯ (u)
ДИФФУЗИОННЫЙ ПОТЕНЦИАЛ
Na+
Na+
Na+
Na+
Cl-
Cl-
Cl-
+
-
ПОДВИЖНОСТЬ ИОНОВ ХЛОРА (v) ВЫШЕ, ЧЕМ ИОНОВ НАТРИЯ (u)
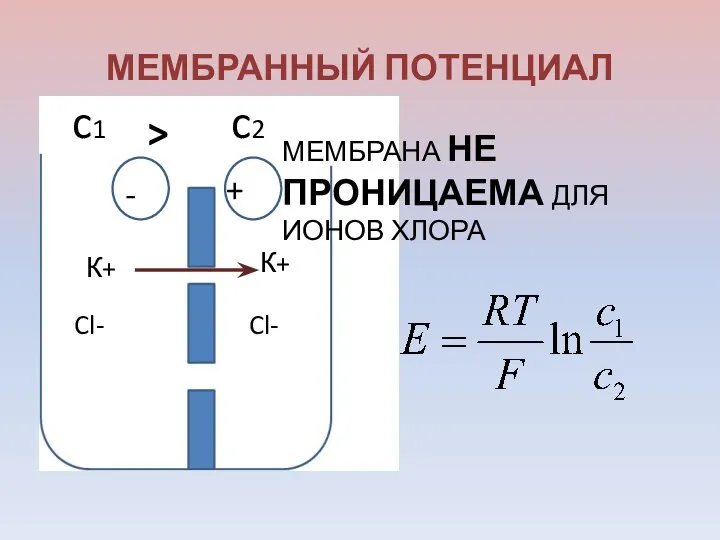
МЕМБРАННЫЙ ПОТЕНЦИАЛ
Cl-
Cl-
К+
К+
МЕМБРАНА НЕ ПРОНИЦАЕМА ДЛЯ ИОНОВ ХЛОРА
-
+
МЕМБРАННЫЙ ПОТЕНЦИАЛ
Cl-
Cl-
К+
К+
МЕМБРАНА НЕ ПРОНИЦАЕМА ДЛЯ ИОНОВ ХЛОРА
-
+

ЭЛЕКТРОХИМИЧЕСКИЙ ПОТЕНЦИАЛ
УСЛОВИЕ РАВНОВЕСИЯ
РАВНОВЕСНЫЕ ПОТЕНЦИАЛЫ:
ENa=+35…+65 мВ
EK = -70…-100 мВ
ЭЛЕКТРОХИМИЧЕСКИЙ ПОТЕНЦИАЛ
УСЛОВИЕ РАВНОВЕСИЯ
РАВНОВЕСНЫЕ ПОТЕНЦИАЛЫ:
ENa=+35…+65 мВ
EK = -70…-100 мВ
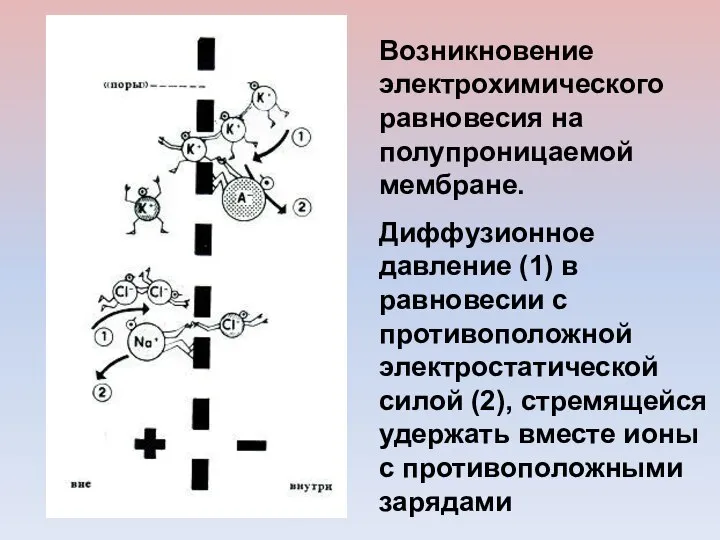
Возникновение электрохимического равновесия на полупроницаемой мембране.
Диффузионное давление (1) в равновесии
Возникновение электрохимического равновесия на полупроницаемой мембране.
Диффузионное давление (1) в равновесии

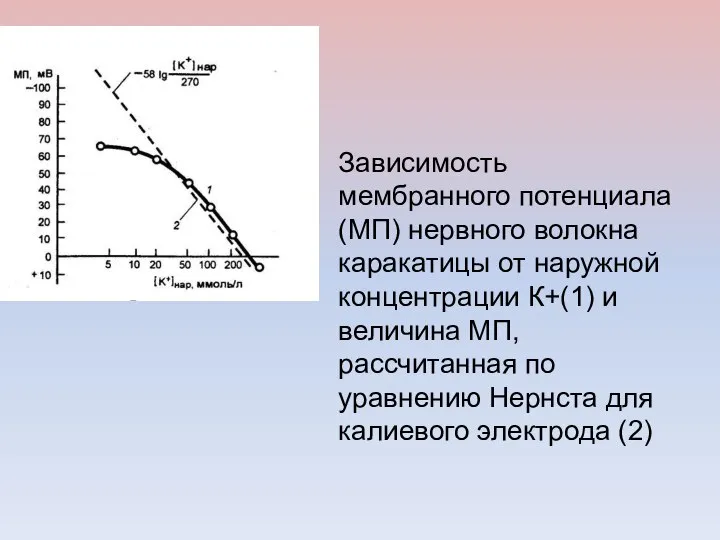
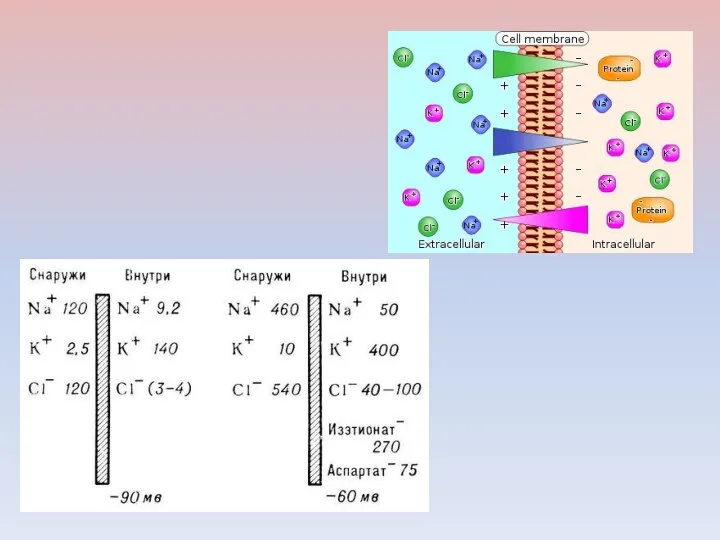
Зависимость мембранного потенциала (МП) нервного волокна каракатицы от наружной концентрации К+(1)
Зависимость мембранного потенциала (МП) нервного волокна каракатицы от наружной концентрации К+(1)
![Зависимость величины ПП от [K+]е (расчетная и экспериментальная кривые). По оси](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1302927/slide-39.jpg)
Зависимость величины ПП от [K+]е (расчетная и экспериментальная кривые).
По оси
Зависимость величины ПП от [K+]е (расчетная и экспериментальная кривые).
По оси

состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов
состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов

УРАВНЕНИЕ НЕРНСТА
Здесь и далее индекс «е» относится к внешним концентрациям ионов,
УРАВНЕНИЕ НЕРНСТА
Здесь и далее индекс «е» относится к внешним концентрациям ионов,

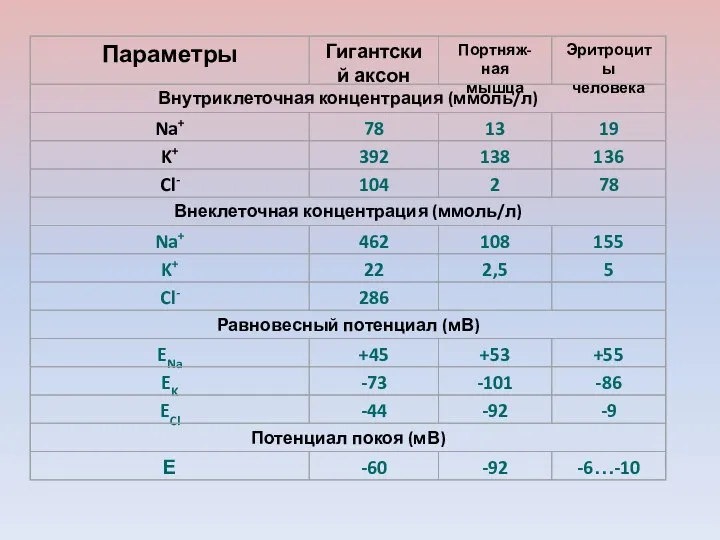
ПП гигантского аксона кальмара ( - 70 мВ) близок к его
ПП гигантского аксона кальмара ( - 70 мВ) близок к его

Итоговая величина ПП, обусловленного переносом многих ионов может быть достаточно точно
Итоговая величина ПП, обусловленного переносом многих ионов может быть достаточно точно

УРАВНЕНИЕ ГОЛЬДМАНА
PK : PNa: PCl = 1 : 0,04 : 0,45
Соотношение
УРАВНЕНИЕ ГОЛЬДМАНА
PK : PNa: PCl = 1 : 0,04 : 0,45
Соотношение

Вклад активного транспорта в формирование потенциала покоя
Вклад активного транспорта в формирование потенциала покоя
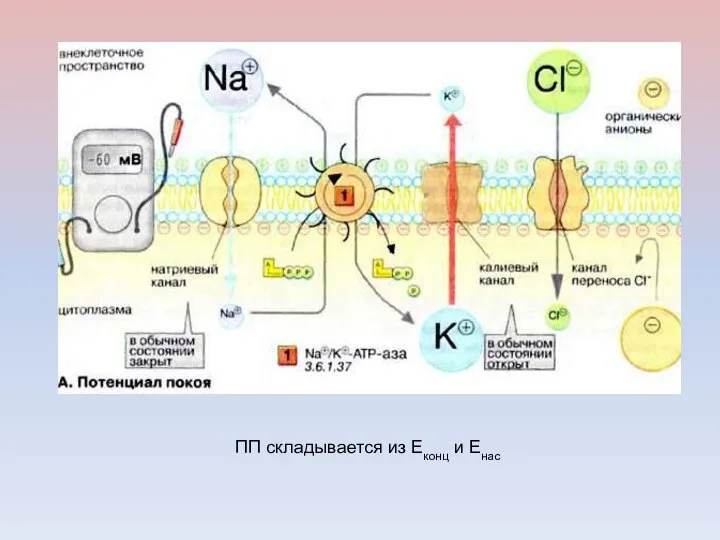
ПП складывается из Еконц и Енас
ПП складывается из Еконц и Енас

РОЛЬ Na/K НАСОСА В ГЕНЕРАЦИИ ПП
Поддержание высокой концентрации К+ внутри клетки,
РОЛЬ Na/K НАСОСА В ГЕНЕРАЦИИ ПП
Поддержание высокой концентрации К+ внутри клетки,

Электротонический потенциал и локальный ответ, их сходство и различие
Электротонический потенциал и локальный ответ, их сходство и различие
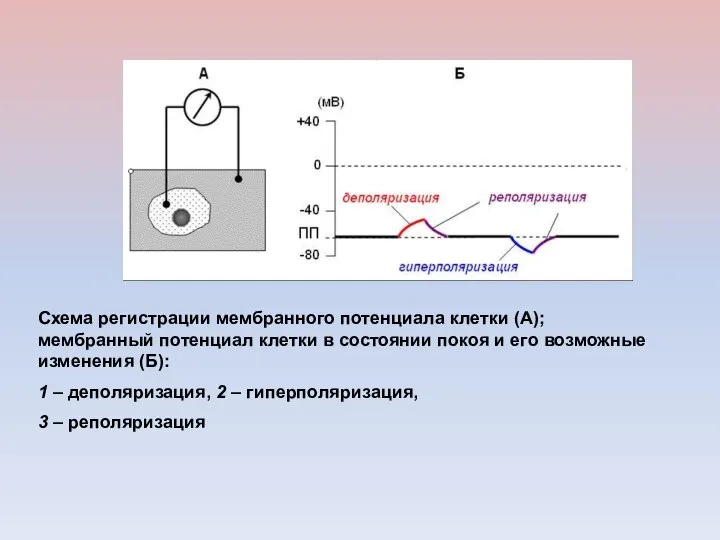
Схема регистрации мембранного потенциала клетки (А); мембранный потенциал клетки в состоянии
Схема регистрации мембранного потенциала клетки (А); мембранный потенциал клетки в состоянии
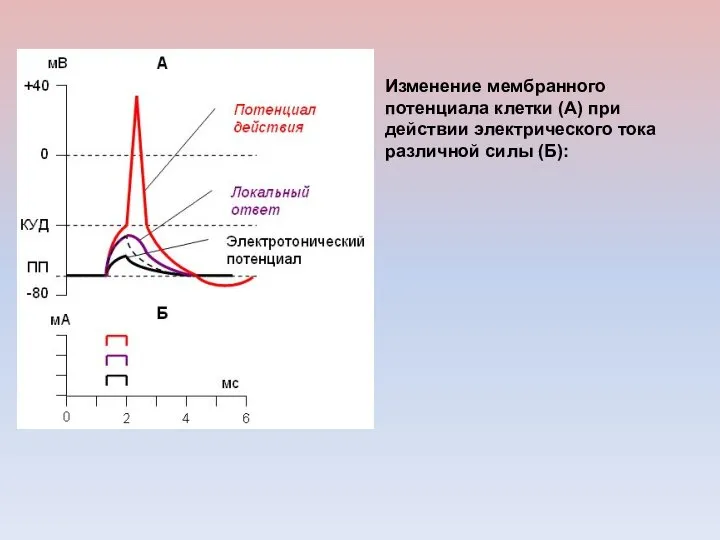
Изменение мембранного потенциала клетки (А) при действии электрического тока различной силы
Изменение мембранного потенциала клетки (А) при действии электрического тока различной силы
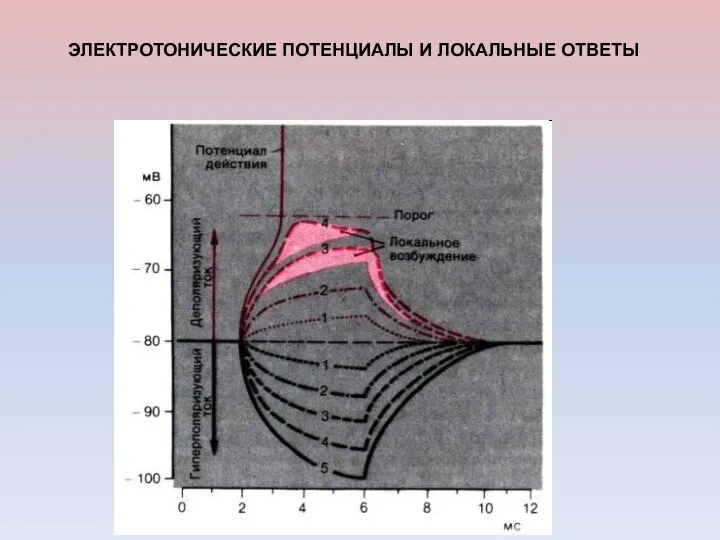
ЭЛЕКТРОТОНИЧЕСКИЕ ПОТЕНЦИАЛЫ И ЛОКАЛЬНЫЕ ОТВЕТЫ
ЭЛЕКТРОТОНИЧЕСКИЕ ПОТЕНЦИАЛЫ И ЛОКАЛЬНЫЕ ОТВЕТЫ
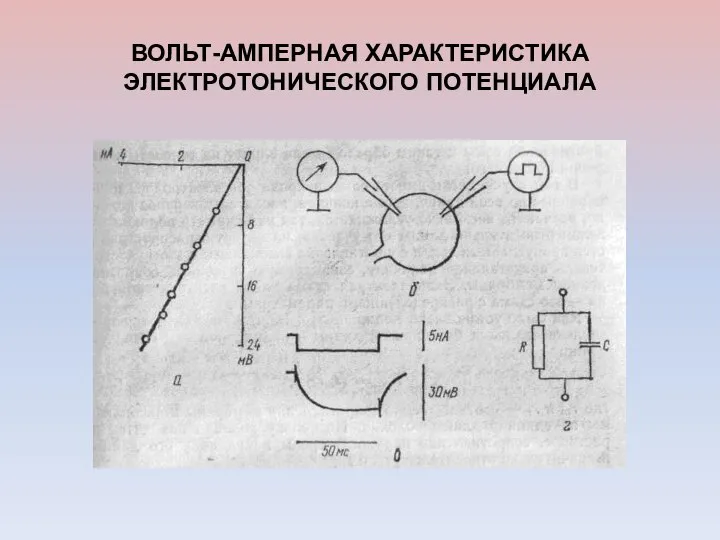
ВОЛЬТ-АМПЕРНАЯ ХАРАКТЕРИСТИКА ЭЛЕКТРОТОНИЧЕСКОГО ПОТЕНЦИАЛА
ВОЛЬТ-АМПЕРНАЯ ХАРАКТЕРИСТИКА ЭЛЕКТРОТОНИЧЕСКОГО ПОТЕНЦИАЛА
 Порядок приема на работу. Порядок заключения и расторжения трудового договора
Порядок приема на работу. Порядок заключения и расторжения трудового договора ПРОГРАММА РАЗВИТИЯ ШКОЛЫ В ЛОГИКЕ НАЦИОНАЛЬНОЙ ОБРАЗОВАТЕЛЬНОЙ ИНИЦИАТИВЫ «НАША НОВАЯ ШКОЛА» РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГ
ПРОГРАММА РАЗВИТИЯ ШКОЛЫ В ЛОГИКЕ НАЦИОНАЛЬНОЙ ОБРАЗОВАТЕЛЬНОЙ ИНИЦИАТИВЫ «НАША НОВАЯ ШКОЛА» РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГ Изгибаемые элементы. Расчет прочности наклонных сечений железобетонных конструкций
Изгибаемые элементы. Расчет прочности наклонных сечений железобетонных конструкций О внесении изменений в Земельный кодекс Российской Федерации и отдельные законодательные акты Российской Федерации
О внесении изменений в Земельный кодекс Российской Федерации и отдельные законодательные акты Российской Федерации Мебель. Ассортимент
Мебель. Ассортимент Театр ХХ века Экспрессионизм (Германия) Футуризм (Италия) Конструктивизм (Россия) Сюрреализм (Франция)
Театр ХХ века Экспрессионизм (Германия) Футуризм (Италия) Конструктивизм (Россия) Сюрреализм (Франция) Параметры зубошевинговальных станков
Параметры зубошевинговальных станков Основные понятия. Механические характеристики материалов
Основные понятия. Механические характеристики материалов Ресвератрол – научный прорыв!
Ресвератрол – научный прорыв! Графическая информация и средства её обработки
Графическая информация и средства её обработки Внедрение информатики в УМК «Гармония», «Начальная школа 21 века» и «Школа 2000-2100» в начальной школе ,
Внедрение информатики в УМК «Гармония», «Начальная школа 21 века» и «Школа 2000-2100» в начальной школе , Политическая система России
Политическая система России Обучение охране труда
Обучение охране труда Минимализм
Минимализм Наладчики аппаратно-программного обеспечения. Области применения профессии
Наладчики аппаратно-программного обеспечения. Области применения профессии Текущий ремонт разъединителя РНДЗ-1-35/1000
Текущий ремонт разъединителя РНДЗ-1-35/1000 Viera Plasma Display. PC Board Recycling. Component Level Repair
Viera Plasma Display. PC Board Recycling. Component Level Repair Урок развития речи во 2 классе МОУ гимназии №9 Изложение по готовому плану учитель Сизонова Ирина Александровна
Урок развития речи во 2 классе МОУ гимназии №9 Изложение по готовому плану учитель Сизонова Ирина Александровна Политические партии и движения
Политические партии и движения Імпульсні модулятори (заняття № 3.1)
Імпульсні модулятори (заняття № 3.1) Иерархическая система многоуровневой организации ЭВМ
Иерархическая система многоуровневой организации ЭВМ Проектирование Последовательных схем
Проектирование Последовательных схем Презентация на тему "22" - скачать презентации по Медицине
Презентация на тему "22" - скачать презентации по Медицине Кубок Love Radio по мини-футболу. Самый яркий праздник спорта и весны!
Кубок Love Radio по мини-футболу. Самый яркий праздник спорта и весны! Языки программирования
Языки программирования Платон и Аристотель об обществе и государстве.
Платон и Аристотель об обществе и государстве.  Средообразующая и рекреационная роль леса
Средообразующая и рекреационная роль леса Архітектура .NET. Основи C#
Архітектура .NET. Основи C#