- Аминокислоты, полипептиды, белки

Содержание
- 2. АМИНОКИСЛОТЫ Это производные углеводородов, содержащие амино- и карбоксильную группы H2N-CH2-COOH аминоуксусная кислота глицин (гликокол) * 2-аминопропановая
- 3. пара-амино- бензойная кислота (ПАБК) анестезин (этиловый эфир ПАБК) новокаин (β-диэтиламино- этиловый эфир ПАБК)
- 4. В белки входят 20 α-аминокислот * алифатические ароматические гетероцикли- ческие изолейцин * * * фенилаланин пролин
- 5. С Т Е Р Е О И З О М Е Р И Я Относительная конфигурация
- 6. D-аланин L-аланин
- 7. Некоторые аминокислоты, например изолейцин, треонин, цистин и гидроксипролин содержат 2 асимметрических атома углерода, поэтому они существуют
- 9. Получение аминокислот 1. Кислотный гидролиз белков 2. Гидроксинитрильный способ (циангидринный, реакция Зелинского-Стадникова) R-CH=O HCN гидроксинитрил NН3
- 11. 3. Взаимодействие аммиака с галогенопроизвод- ными карбоновых кислот (реакция Гофмана) Сl2 - HCl 2NН3 - NH4Cl
- 12. Кислотно-основные свойства аминокислот Образование внутренних солей (биполярных или амфотерных ионов) : R± 2. Амфотерность HCl NaOH
- 13. 3. Аминокислоты - комплексоны Хелатный эффект [Ni(NH3)6]2+ Кнест. ≈ 10-8 Кнест. ≈ 10-18 Хелатный эффект заклю-
- 14. Причины устойчивости хелатных соединений: увеличение энтропии системы; 2) образование пяти- и щестичленных циклов Хелатный эффект иначе
- 15. пентацин Хелатотерапия антидотная терапия при отравлении солями тяжелых металлов (Pb2+, Hg2+, Cd2+, Cu2+ и др.) 2)
- 16. Изоэлектрическая точка белка Это то значение рН = рI, при котором число разно- именных зарядов в
- 17. ИЭТ аминокислоты - это то значение рН, при котором концентрация катионов (R+) равна кон- центрации анионов
- 18. [Н+] = Ка1 . Ка2 ; рН = рI = pКа1 + pКа2 2 Для глицина:
- 19. Аминокислоты кислые нейтральные основные моноамино- дикарбоновые моноамино- монокарбоновые диамино- монокарбоновые 1 NH2-группа, 2 COOН-группы 1 NH2-группа,
- 20. Методы определения ИЭТ белков I. Прямой метод - метод электрофореза (это наблю- дение за поведением чaстиц
- 21. u электрофоретическая подвижность рН ИЭТ
- 22. Электрофоретическое разделение белков буфер (рН=8,6) буфер (рН=8,6) + - Хроматографическая бумага Смесь белков (*) рI α-глобулин
- 23. + - * смесь белков α Чем больше (pH - pI), тем больше ско- рость дви-
- 24. Электрофорез белков плазмы α β γ альбумин
- 25. II. Косвенные методы Ряд свойств белков экстремален (max или min) в изоэлектрической точке 1. Гидратация -
- 26. Термические превращения аминокислот 1) α-аминокислоты α α + t0 - 2H2O 2,5-диалкил-3,6-дикето- пиперазин
- 27. 2) β-аминокислоты α β t0 - NH3 непредельная кислота 3) γ-аминокислоты γ t0 - H2O γ-лактам
- 28. Другие общие химические свойства аминокислот по NH2-группе по СООН-группе - СО2 R-CH2-NH2 HO-N=O - N2 C2H5OH
- 29. по NH2-группе по СООН-группе PCl5 CH3COCl - HCl - HCl NH3 - H2O R-C=O R -
- 30. Превращения аминокислот в организме Аминокислота метио- нин (Мет) – поставщик СН3-группы в реакциях алкилирования (например, коламин
- 31. 3) Дезаминирование (генетическая связь α-аланина, молочной кислоты и ПВК) Н2O [O] [H] NН3 4) Декарбоксилирование H2N-(CH2)5-NH2
- 32. Полипептиды Это продукты поликонденсации аминокислот (с отщеплением воды) и образованием амидной (пептидной) связи В общем виде
- 33. Пример. γ-Глутатион (трипептид) – активатор ферментов, содержится во всех клетках организма, участвует в ox-red процессах, т.к.
- 34. Специфичность белков определяется: аминокислотным составом – природой и количеством аминокислот 2) аминокислотной последовательностью – первичной структурой
- 35. Пространственное строение полипептидов Вторичная структура белка 1. Пептидная связь а) частичная двоесвязанность (из-за р-π сопряжения электронной
- 36. б) трансоидная конформация Сравните! lС=O 0,121 нм lN-С 0,147 нм длиннее! короче!
- 37. 2. α-Спираль и β-складчатая структура Упорядоченное свертывание «остова» полипептидной цепи в результате образования водородных связей между
- 39. α-Спираль
- 40. β-складчатая структура N-конец С-конец (преобладает в фибрилляр- ных белках)
- 41. β-складка
- 42. β-Складка
- 43. Полипептид
- 44. Тройная спираль коллагена
- 45. Третичная структура белка Трехмерное свертывание полипептидной цепи, приводя- щее к наложению одних участ- ков α-спирали или
- 47. Ион-ионные взаимодействия Ковалентное связывание N-конец С-конец
- 48. Следует иметь в виду, что изгибы полипептидной цепи обусловлены наличием аминокислот со вторичной аминогруппой – пролина
- 49. Некоторые белки, например гемоглобин, обладают чет- вертичной структурой; при этом одинаковые или сход- ные субъединицы соединя-
- 50. Полипептид
- 52. Скачать презентацию
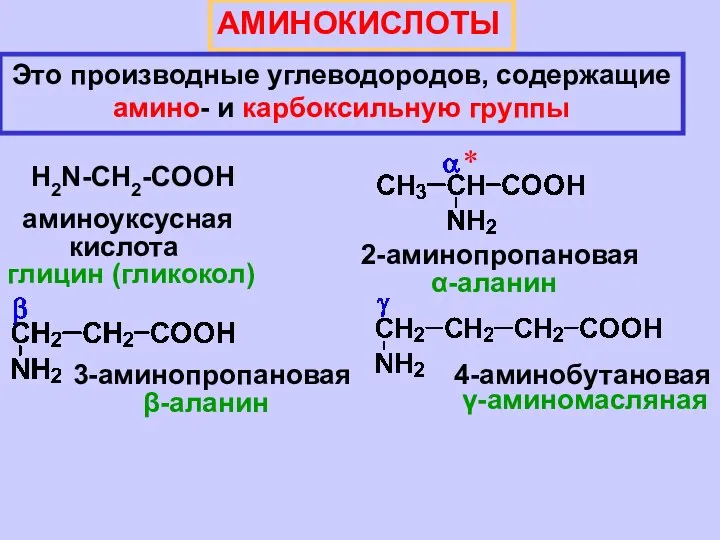
АМИНОКИСЛОТЫ
Это производные углеводородов, содержащие
амино- и карбоксильную группы
H2N-CH2-COOH
аминоуксусная
кислота
глицин (гликокол)
*
2-аминопропановая
α-аланин
3-аминопропановая
АМИНОКИСЛОТЫ
Это производные углеводородов, содержащие
амино- и карбоксильную группы
H2N-CH2-COOH
аминоуксусная
кислота
глицин (гликокол)
*
2-аминопропановая
α-аланин
3-аминопропановая
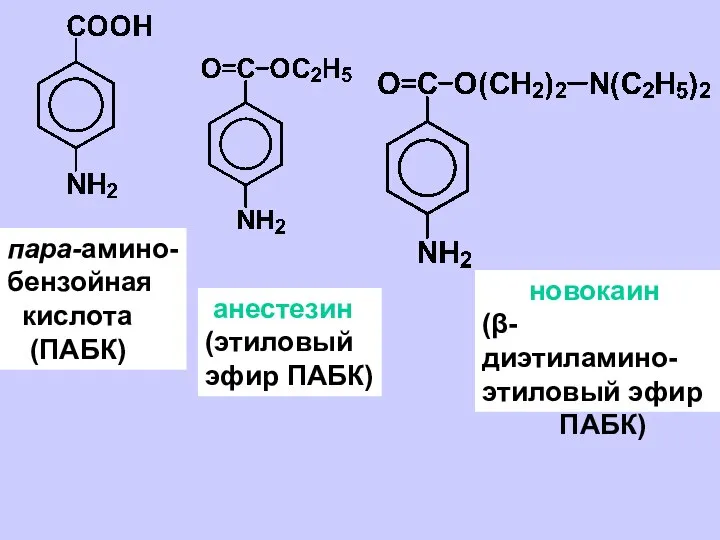
пара-амино-
бензойная
кислота
(ПАБК)
анестезин
(этиловый
эфир ПАБК)
новокаин
(β-диэтиламино-
этиловый эфир
ПАБК)
пара-амино-
бензойная
кислота
(ПАБК)
анестезин
(этиловый
эфир ПАБК)
новокаин
(β-диэтиламино-
этиловый эфир
ПАБК)
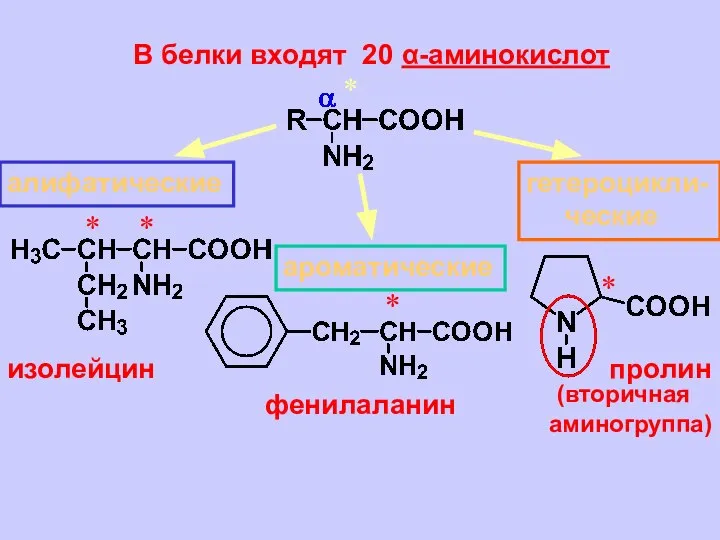
В белки входят 20 α-аминокислот
*
алифатические
ароматические
гетероцикли-
ческие
изолейцин
*
*
*
фенилаланин
пролин
*
(вторичная
аминогруппа)
*
алифатические
ароматические
гетероцикли-
ческие
изолейцин
*
*
*
фенилаланин
пролин
*
(вторичная
аминогруппа)
С Т Е Р Е О И З О М Е
С Т Е Р Е О И З О М Е
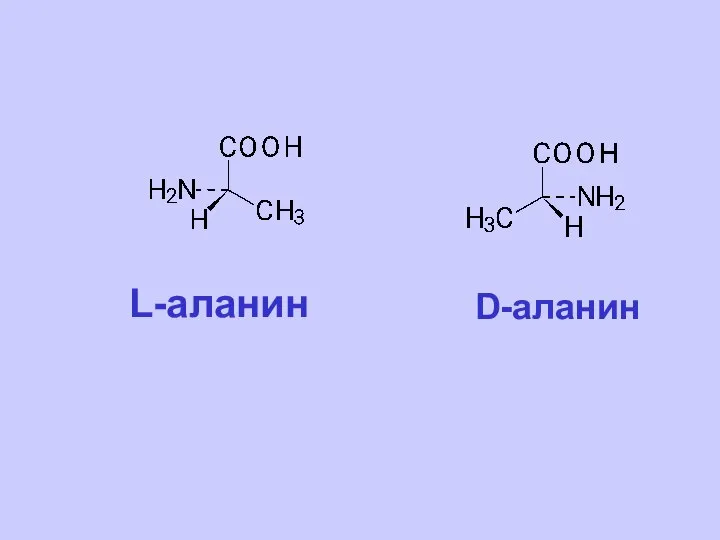
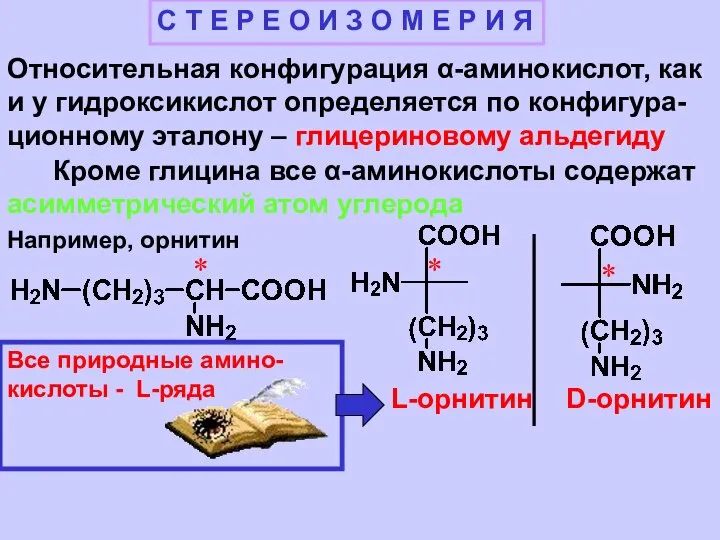
D-аланин
L-аланин
D-аланин
L-аланин
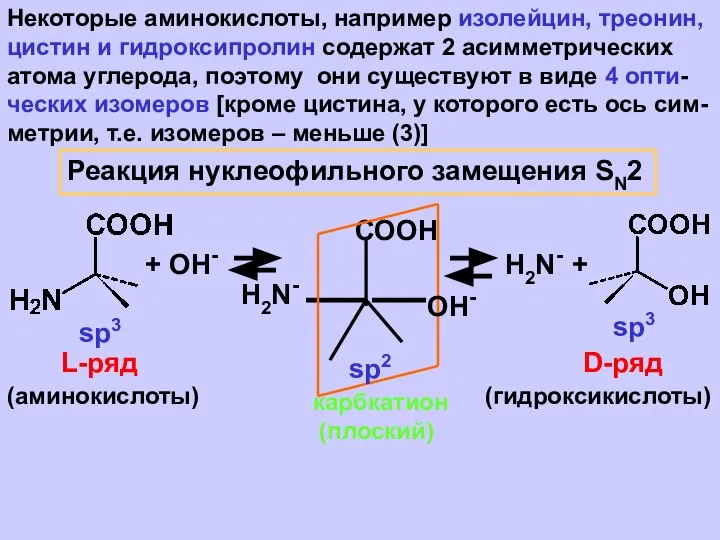
Некоторые аминокислоты, например изолейцин, треонин,
цистин и гидроксипролин содержат 2 асимметрических
атома углерода,
Некоторые аминокислоты, например изолейцин, треонин,
цистин и гидроксипролин содержат 2 асимметрических
атома углерода,
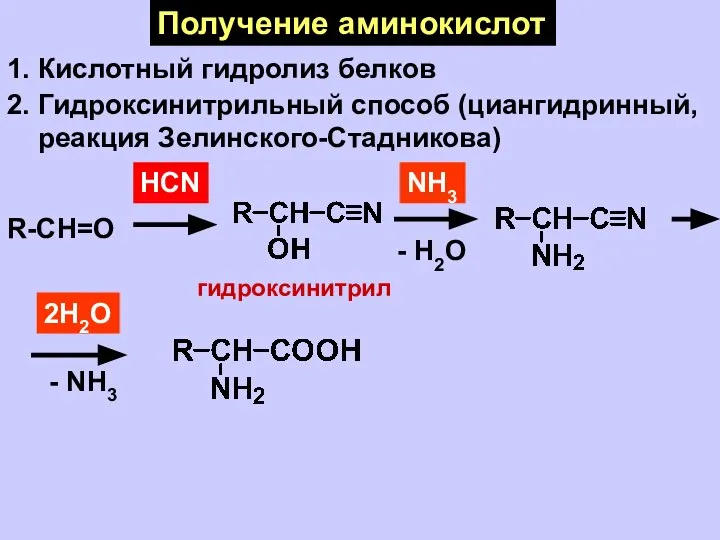
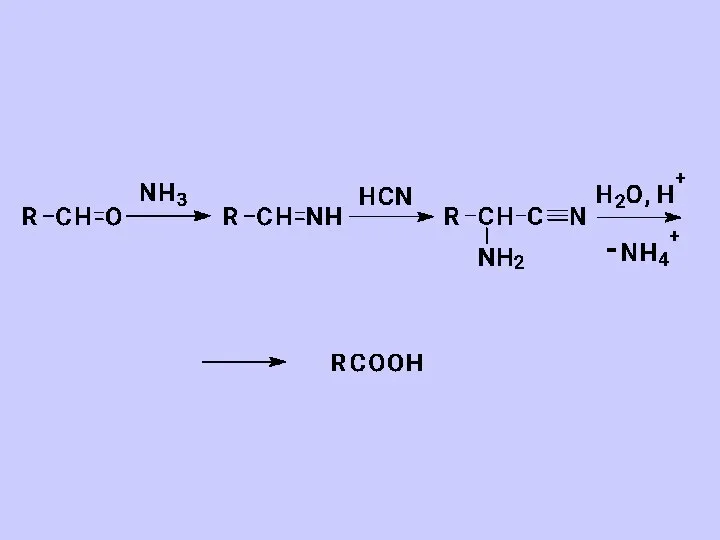
Получение аминокислот
1. Кислотный гидролиз белков
2. Гидроксинитрильный способ (циангидринный,
реакция Зелинского-Стадникова)
R-CH=O
HCN
гидроксинитрил
NН3
-
Получение аминокислот
1. Кислотный гидролиз белков
2. Гидроксинитрильный способ (циангидринный,
реакция Зелинского-Стадникова)
R-CH=O
HCN
гидроксинитрил
NН3
-
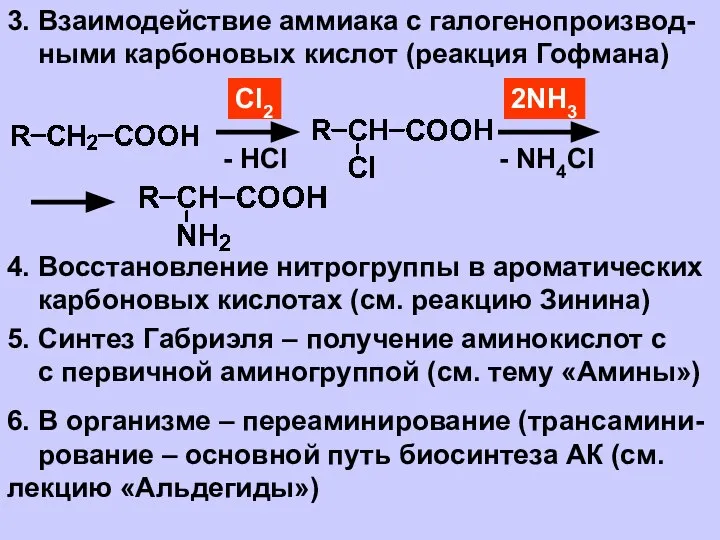
3. Взаимодействие аммиака с галогенопроизвод-
ными карбоновых кислот (реакция Гофмана)
Сl2
- HCl
2NН3
-
3. Взаимодействие аммиака с галогенопроизвод-
ными карбоновых кислот (реакция Гофмана)
Сl2
- HCl
2NН3
-
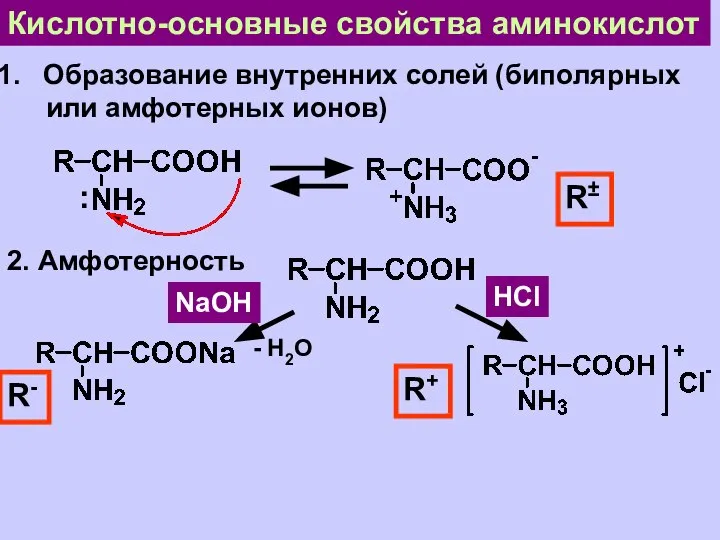
Кислотно-основные свойства аминокислот
Образование внутренних солей (биполярных
или амфотерных ионов)
:
R±
2. Амфотерность
HCl
NaOH
-
Кислотно-основные свойства аминокислот
Образование внутренних солей (биполярных
или амфотерных ионов)
:
R±
2. Амфотерность
HCl
NaOH
-
![3. Аминокислоты - комплексоны Хелатный эффект [Ni(NH3)6]2+ Кнест. ≈ 10-8 Кнест.](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1300353/slide-12.jpg)
3. Аминокислоты - комплексоны
Хелатный эффект
[Ni(NH3)6]2+
Кнест. ≈ 10-8
Кнест. ≈ 10-18
Хелатный эффект
3. Аминокислоты - комплексоны
Хелатный эффект
[Ni(NH3)6]2+
Кнест. ≈ 10-8
Кнест. ≈ 10-18
Хелатный эффект
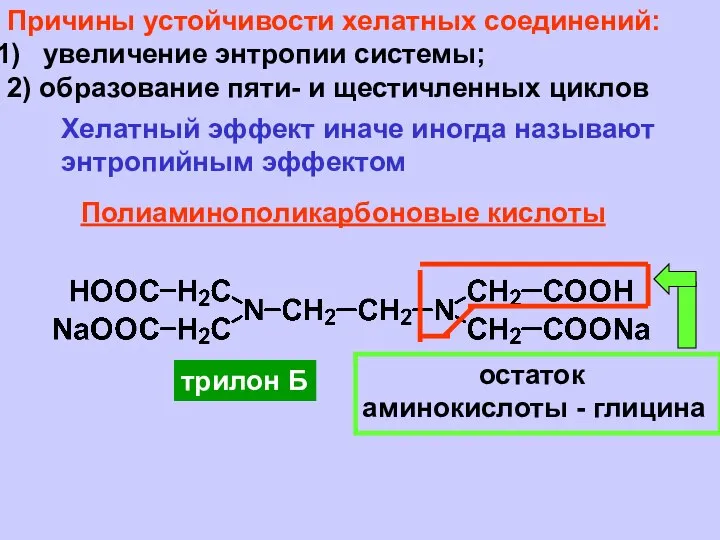
Причины устойчивости хелатных соединений:
увеличение энтропии системы;
2) образование пяти- и щестичленных циклов
Хелатный
Причины устойчивости хелатных соединений:
увеличение энтропии системы;
2) образование пяти- и щестичленных циклов
Хелатный
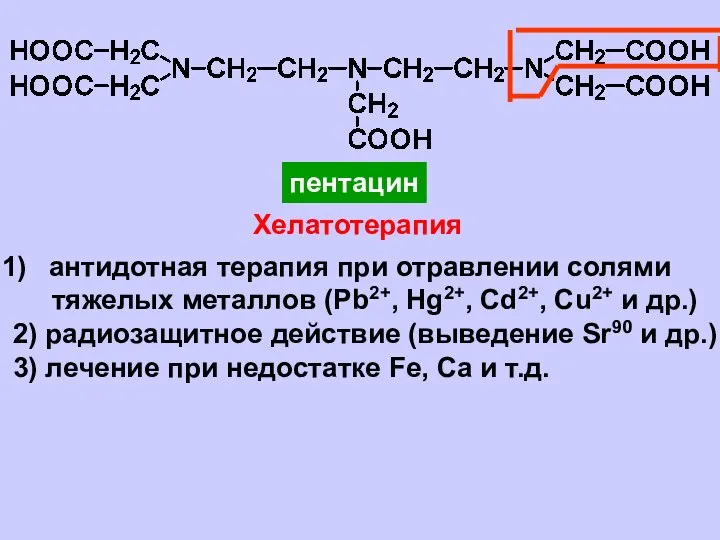
пентацин
Хелатотерапия
антидотная терапия при отравлении солями
тяжелых металлов (Pb2+, Hg2+, Cd2+, Cu2+
пентацин
Хелатотерапия
антидотная терапия при отравлении солями
тяжелых металлов (Pb2+, Hg2+, Cd2+, Cu2+
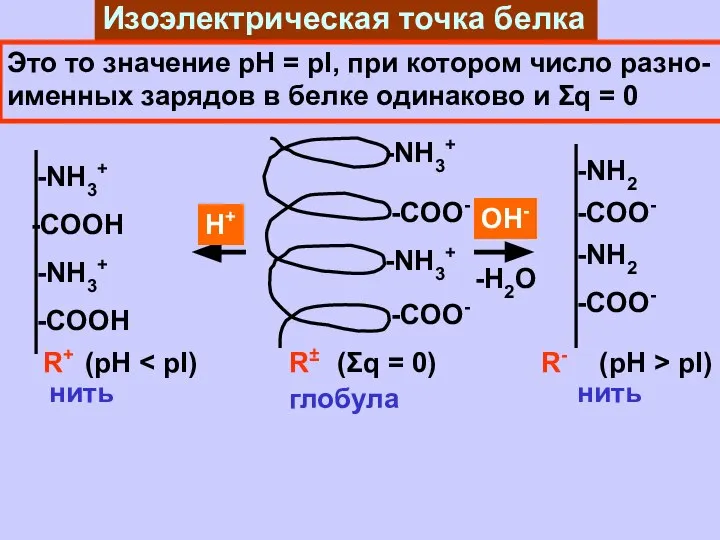
Изоэлектрическая точка белка
Это то значение рН = рI, при котором число
Изоэлектрическая точка белка
Это то значение рН = рI, при котором число
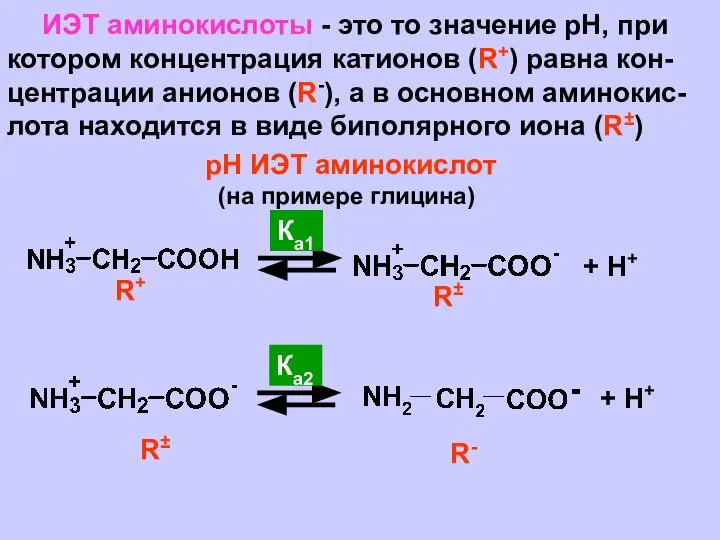
ИЭТ аминокислоты - это то значение рН, при
котором концентрация катионов
ИЭТ аминокислоты - это то значение рН, при
котором концентрация катионов
![[Н+] = Ка1 . Ка2 ; рН = рI = pКа1](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1300353/slide-17.jpg)
[Н+] =
Ка1 . Ка2 ;
рН = рI =
pКа1 + pКа2
2
Для глицина:
[Н+] =
Ка1 . Ка2 ;
рН = рI =
pКа1 + pКа2
2
Для глицина:
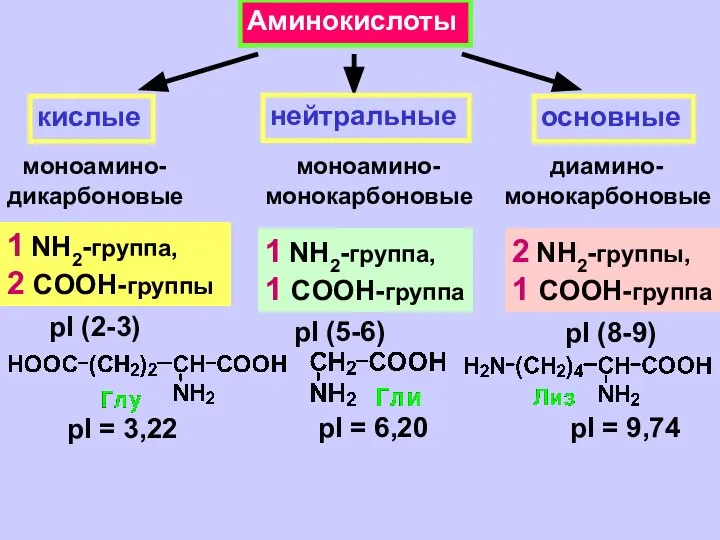
Аминокислоты
кислые
нейтральные
основные
моноамино-
дикарбоновые
моноамино-
монокарбоновые
диамино-
монокарбоновые
1 NH2-группа,
2 COOН-группы
1 NH2-группа,
1 COOН-группа
2 NH2-группы,
1 COOН-группа
рI
Аминокислоты
кислые
нейтральные
основные
моноамино-
дикарбоновые
моноамино-
монокарбоновые
диамино-
монокарбоновые
1 NH2-группа,
2 COOН-группы
1 NH2-группа,
1 COOН-группа
2 NH2-группы,
1 COOН-группа
рI
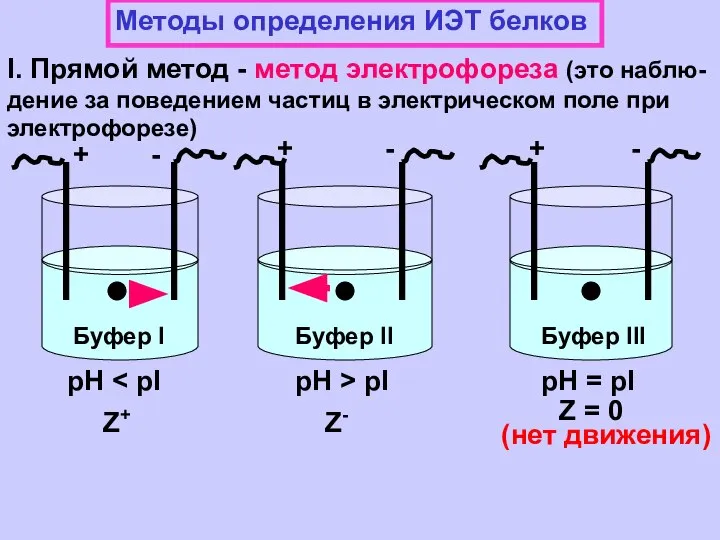
Методы определения ИЭТ белков
I. Прямой метод - метод электрофореза (это наблю-
дение
Методы определения ИЭТ белков
I. Прямой метод - метод электрофореза (это наблю-
дение
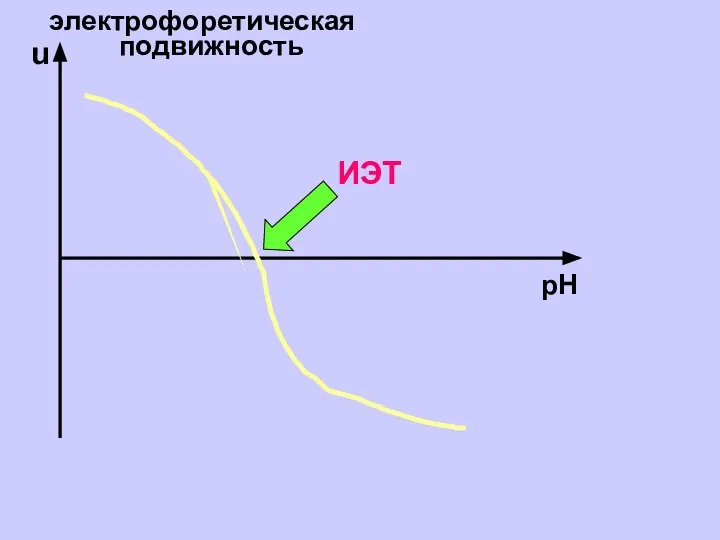
u
электрофоретическая
подвижность
рН
ИЭТ
u
электрофоретическая
подвижность
рН
ИЭТ
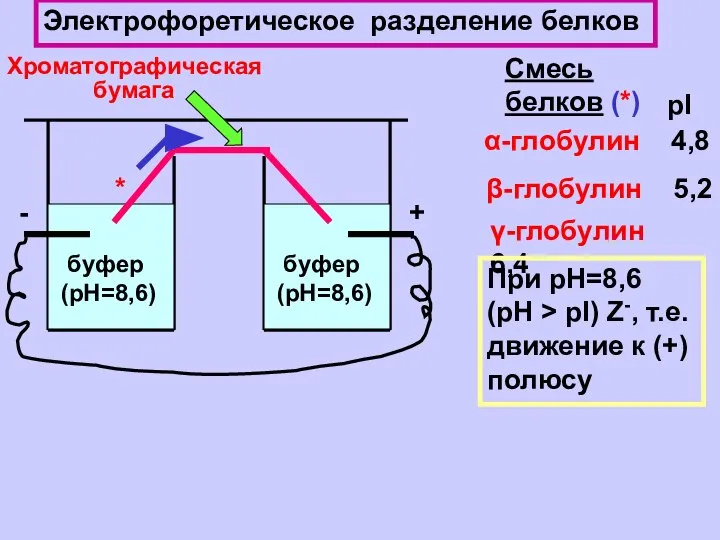
Электрофоретическое разделение белков
буфер
(рН=8,6)
буфер
(рН=8,6)
+
-
Хроматографическая
бумага
Смесь
белков (*)
рI
α-глобулин 4,8
γ-глобулин 6,4
β-глобулин 5,2
*
При рН=8,6
(pH
Электрофоретическое разделение белков
буфер
(рН=8,6)
буфер
(рН=8,6)
+
-
Хроматографическая
бумага
Смесь
белков (*)
рI
α-глобулин 4,8
γ-глобулин 6,4
β-глобулин 5,2
*
При рН=8,6
(pH
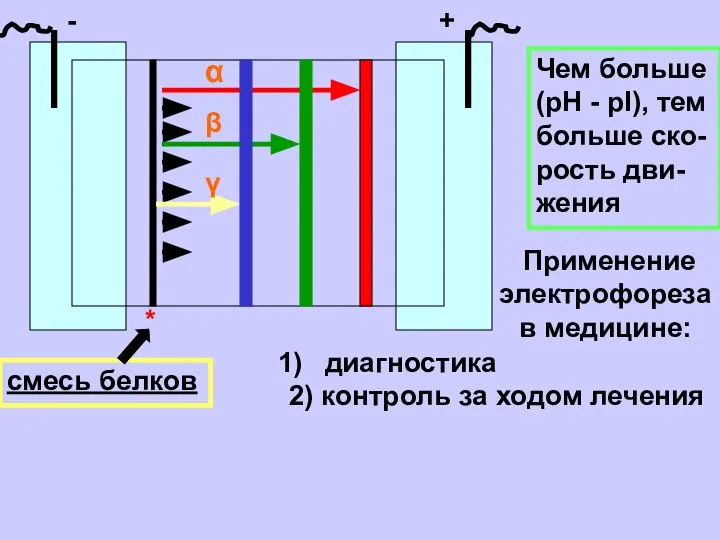
+
-
*
смесь белков
α
Чем больше
(pH - pI), тем
больше ско-
рость дви-
жения
Применение
электрофореза
в медицине:
диагностика
2) контроль
+
-
*
смесь белков
α
Чем больше
(pH - pI), тем
больше ско-
рость дви-
жения
Применение
электрофореза
в медицине:
диагностика
2) контроль
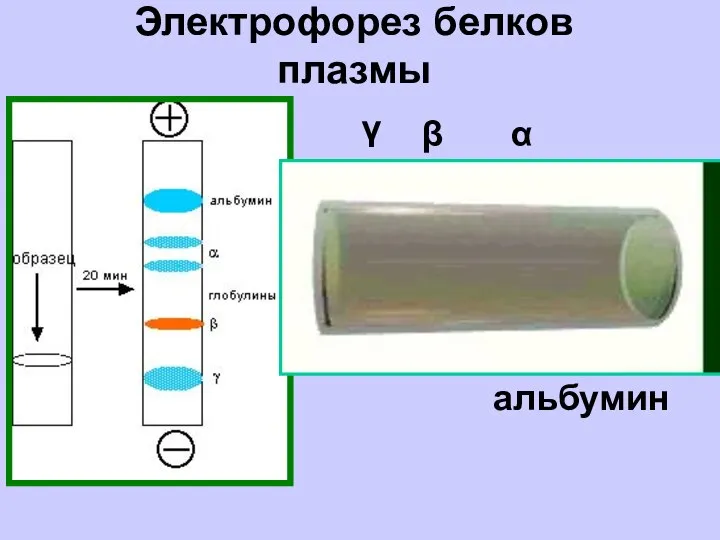
Электрофорез белков плазмы
α
β
γ
альбумин
Электрофорез белков плазмы
α
β
γ
альбумин

II. Косвенные методы
Ряд свойств белков экстремален (max или min)
в изоэлектрической
II. Косвенные методы
Ряд свойств белков экстремален (max или min)
в изоэлектрической
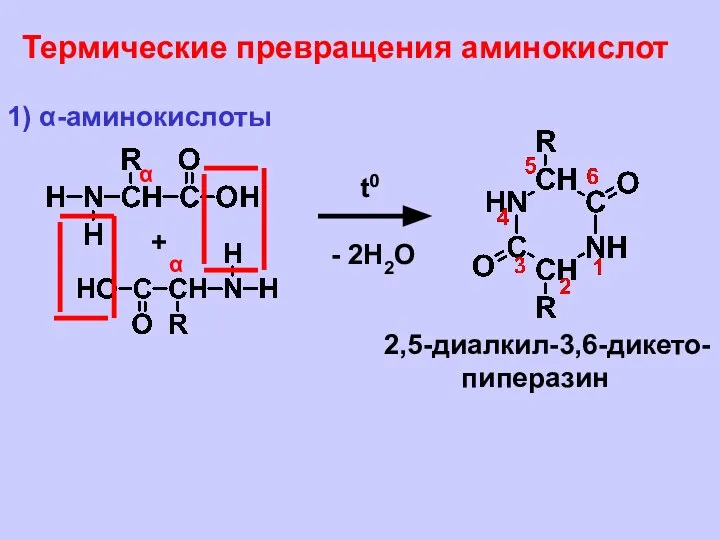
Термические превращения аминокислот
1) α-аминокислоты
α
α
+
t0
- 2H2O
2,5-диалкил-3,6-дикето-
пиперазин
Термические превращения аминокислот
1) α-аминокислоты
α
α
+
t0
- 2H2O
2,5-диалкил-3,6-дикето-
пиперазин
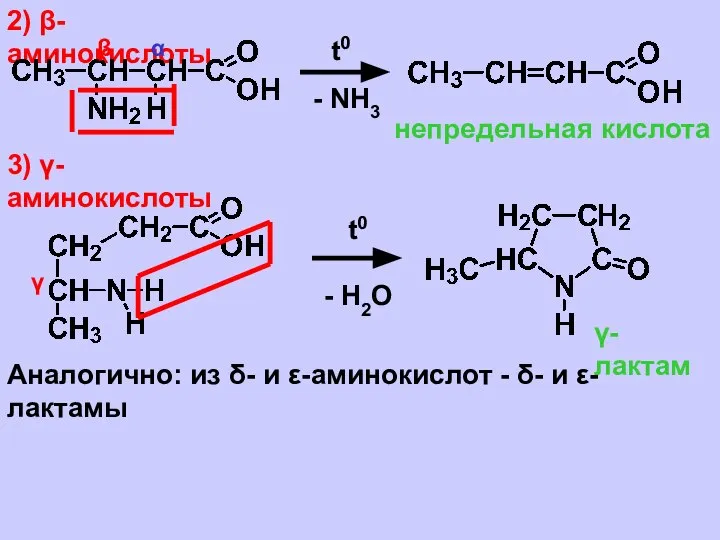
2) β-аминокислоты
α
β
t0
- NH3
непредельная кислота
3) γ-аминокислоты
γ
t0
- H2O
γ-лактам
Аналогично: из δ- и ε-аминокислот -
2) β-аминокислоты
α
β
t0
- NH3
непредельная кислота
3) γ-аминокислоты
γ
t0
- H2O
γ-лактам
Аналогично: из δ- и ε-аминокислот -
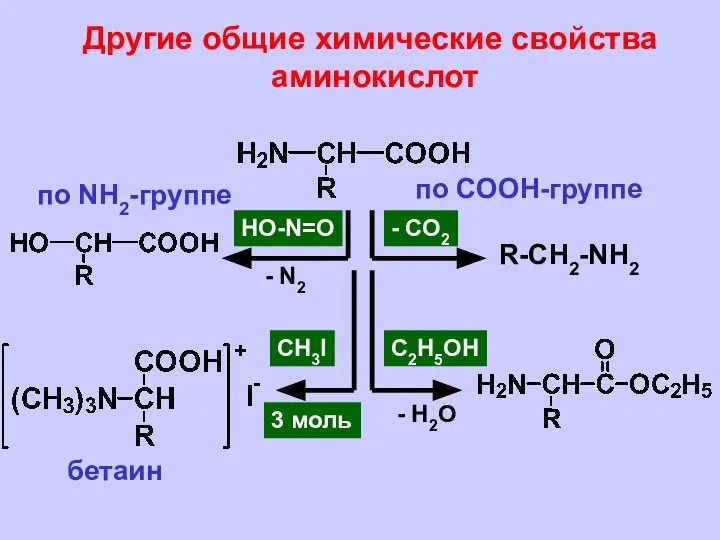
Другие общие химические свойства
аминокислот
по NH2-группе
по СООН-группе
- СО2
R-CH2-NH2
HO-N=O
- N2
C2H5OH
- H2O
CH3I
3 моль
бетаин
Другие общие химические свойства
аминокислот
по NH2-группе
по СООН-группе
- СО2
R-CH2-NH2
HO-N=O
- N2
C2H5OH
- H2O
CH3I
3 моль
бетаин
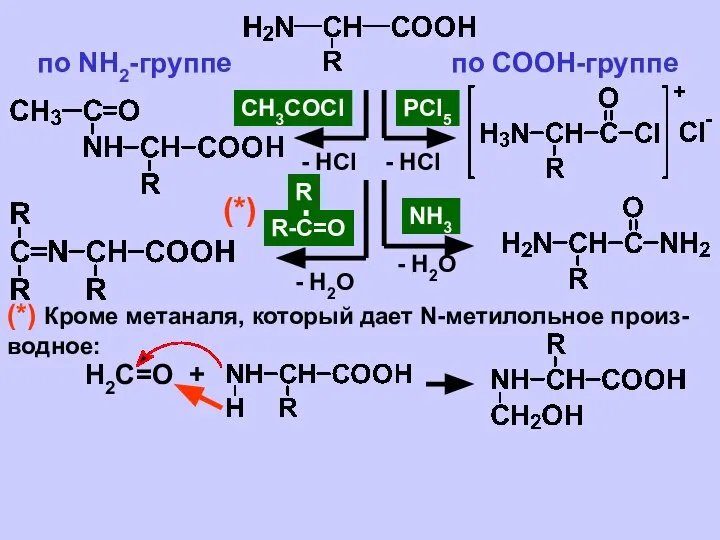
по NH2-группе
по СООН-группе
PCl5
CH3COCl
- HCl
- HCl
NH3
- H2O
R-C=O
R
- H2O
(*)
(*) Кроме метаналя, который дает
по NH2-группе
по СООН-группе
PCl5
CH3COCl
- HCl
- HCl
NH3
- H2O
R-C=O
R
- H2O
(*)
(*) Кроме метаналя, который дает
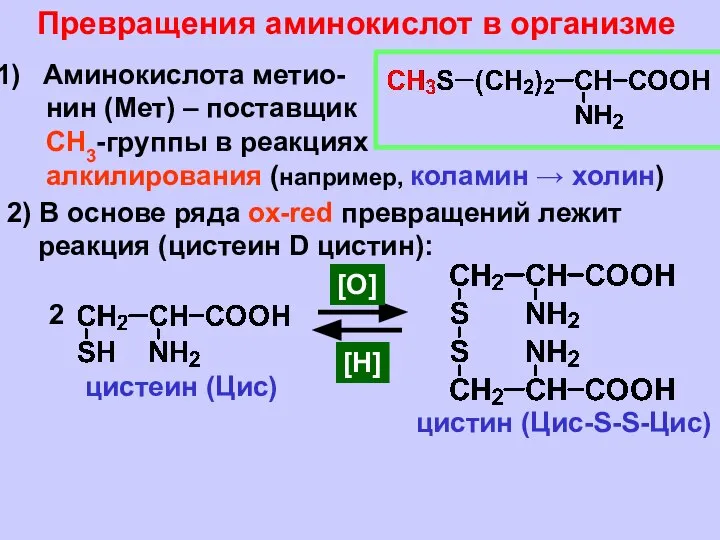
Превращения аминокислот в организме
Аминокислота метио-
нин (Мет) – поставщик
СН3-группы в
Превращения аминокислот в организме
Аминокислота метио-
нин (Мет) – поставщик
СН3-группы в
![3) Дезаминирование (генетическая связь α-аланина, молочной кислоты и ПВК) Н2O [O]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1300353/slide-30.jpg)
3) Дезаминирование (генетическая связь
α-аланина, молочной кислоты и ПВК)
Н2O
[O]
[H]
NН3
4) Декарбоксилирование
H2N-(CH2)5-NH2
-
3) Дезаминирование (генетическая связь
α-аланина, молочной кислоты и ПВК)
Н2O
[O]
[H]
NН3
4) Декарбоксилирование
H2N-(CH2)5-NH2
-
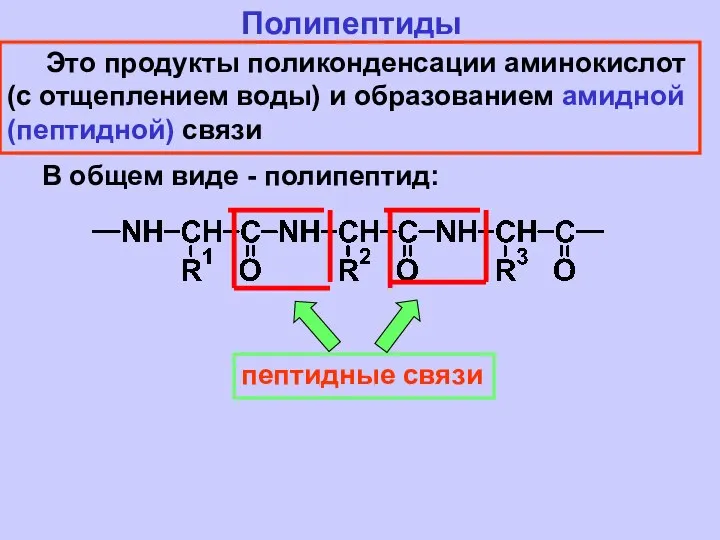
Полипептиды
Это продукты поликонденсации аминокислот
(с отщеплением воды) и образованием амидной
(пептидной) связи
В
Полипептиды
Это продукты поликонденсации аминокислот
(с отщеплением воды) и образованием амидной
(пептидной) связи
В
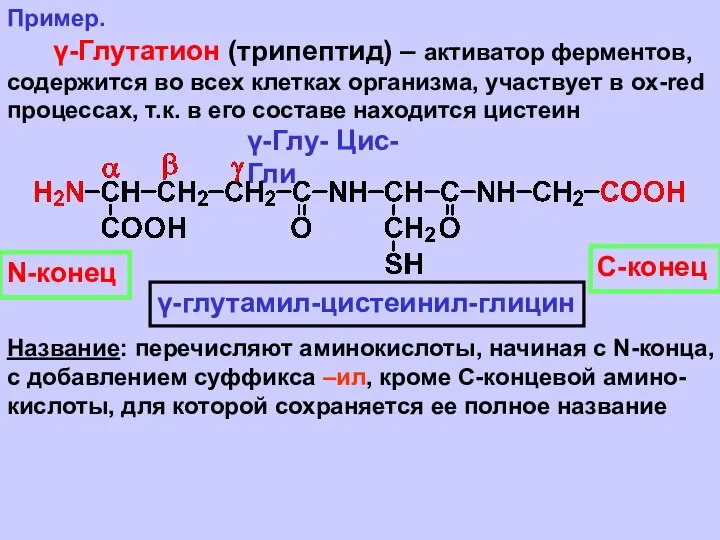
Пример.
γ-Глутатион (трипептид) – активатор ферментов,
содержится во всех клетках организма, участвует
Пример.
γ-Глутатион (трипептид) – активатор ферментов,
содержится во всех клетках организма, участвует

Специфичность белков определяется:
аминокислотным составом – природой и
количеством аминокислот
2) аминокислотной последовательностью
Специфичность белков определяется:
аминокислотным составом – природой и
количеством аминокислот
2) аминокислотной последовательностью

Пространственное строение полипептидов
Вторичная структура белка
1. Пептидная связь
а) частичная двоесвязанность (из-за р-π
Пространственное строение полипептидов
Вторичная структура белка
1. Пептидная связь
а) частичная двоесвязанность (из-за р-π
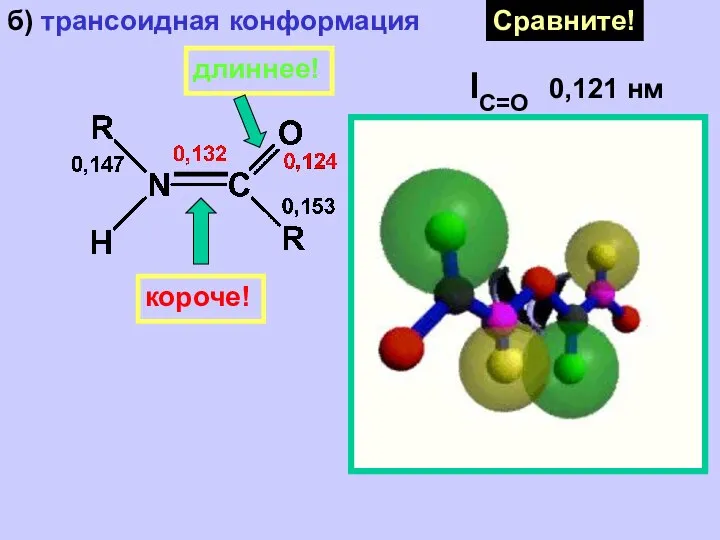
б) трансоидная конформация
Сравните!
lС=O 0,121 нм
lN-С 0,147 нм
длиннее!
короче!
б) трансоидная конформация
Сравните!
lС=O 0,121 нм
lN-С 0,147 нм
длиннее!
короче!
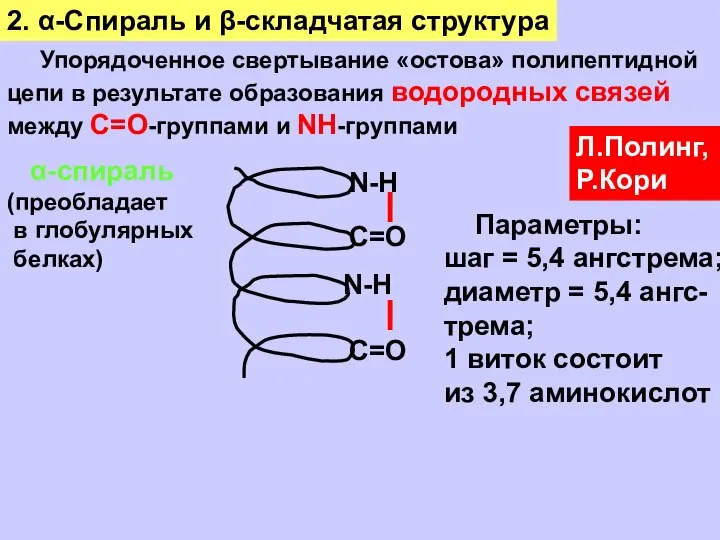
2. α-Спираль и β-складчатая структура
Упорядоченное свертывание «остова» полипептидной
цепи в результате
2. α-Спираль и β-складчатая структура
Упорядоченное свертывание «остова» полипептидной
цепи в результате
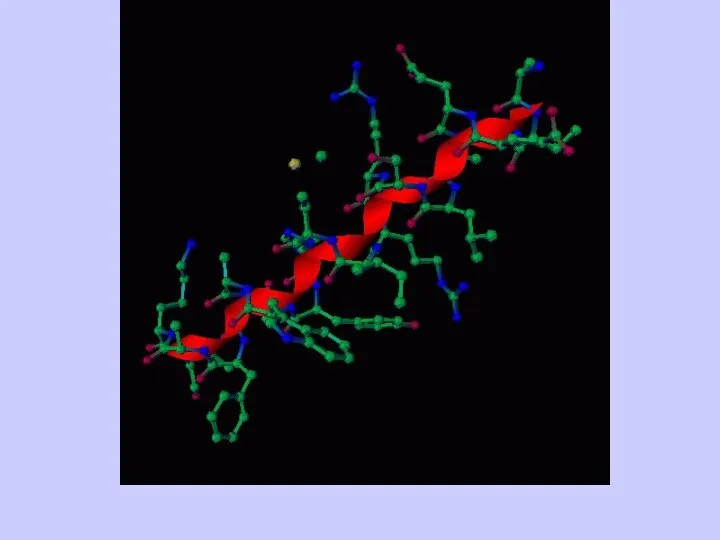
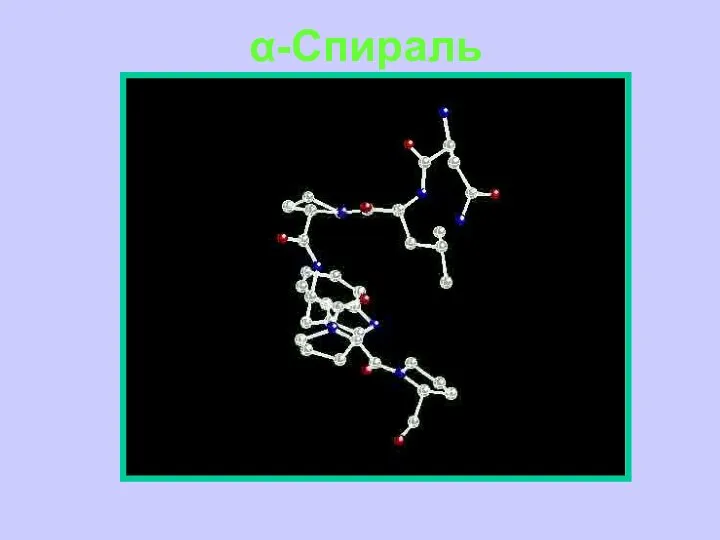
α-Спираль
α-Спираль
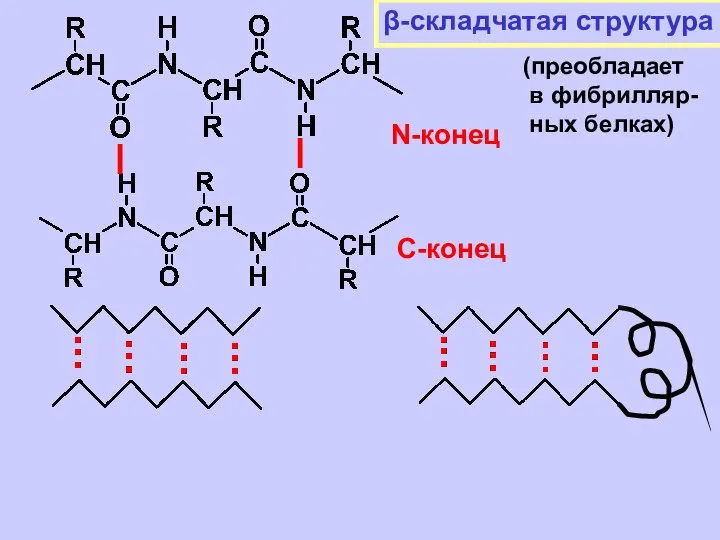
β-складчатая структура
N-конец
С-конец
(преобладает
в фибрилляр-
ных белках)
β-складчатая структура
N-конец
С-конец
(преобладает
в фибрилляр-
ных белках)
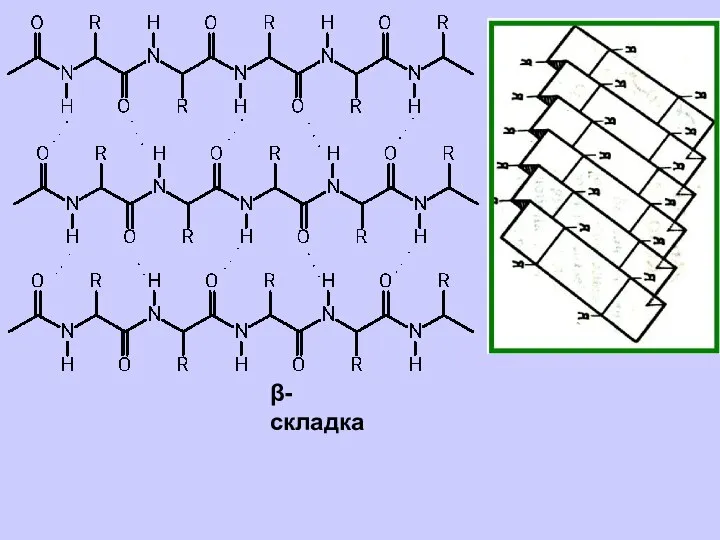
β-складка
β-складка
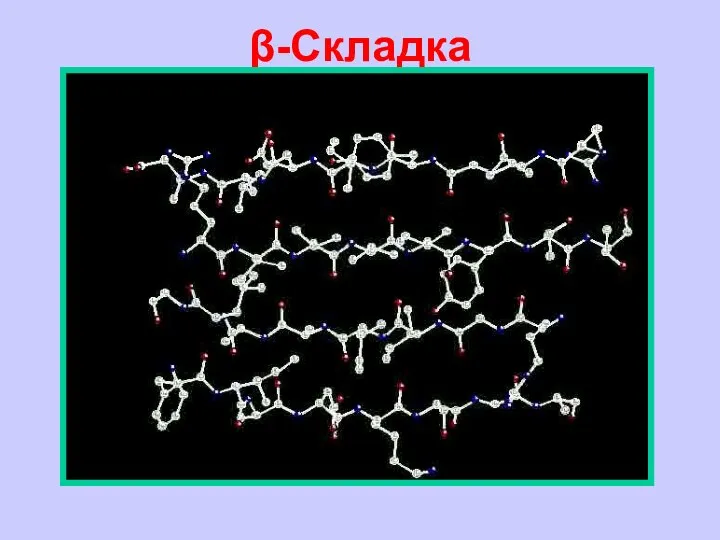
β-Складка
β-Складка

Полипептид
Полипептид
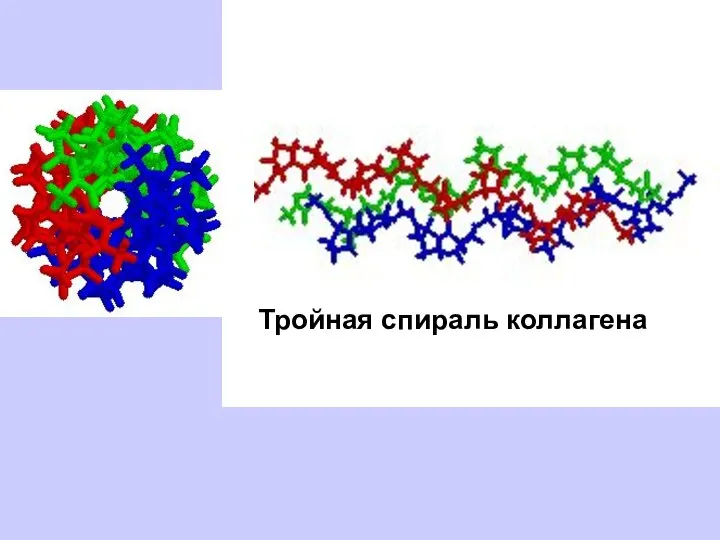
Тройная спираль коллагена
Тройная спираль коллагена

Третичная структура белка
Трехмерное свертывание
полипептидной цепи, приводя-
щее к наложению одних
Третичная структура белка
Трехмерное свертывание
полипептидной цепи, приводя-
щее к наложению одних
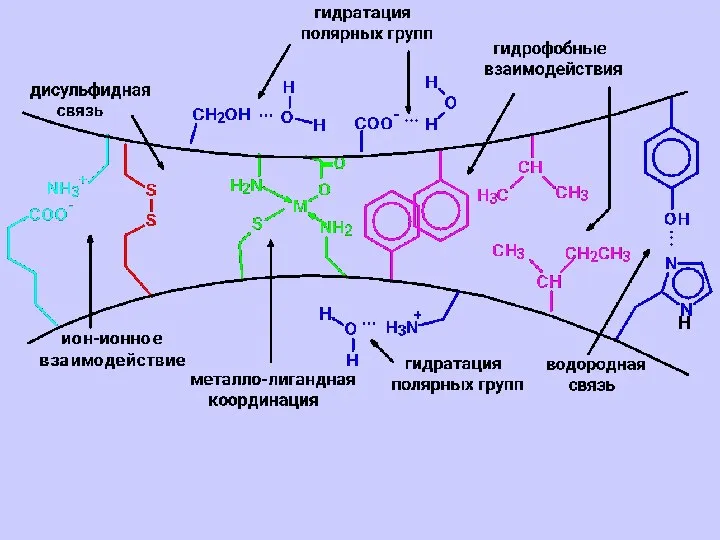
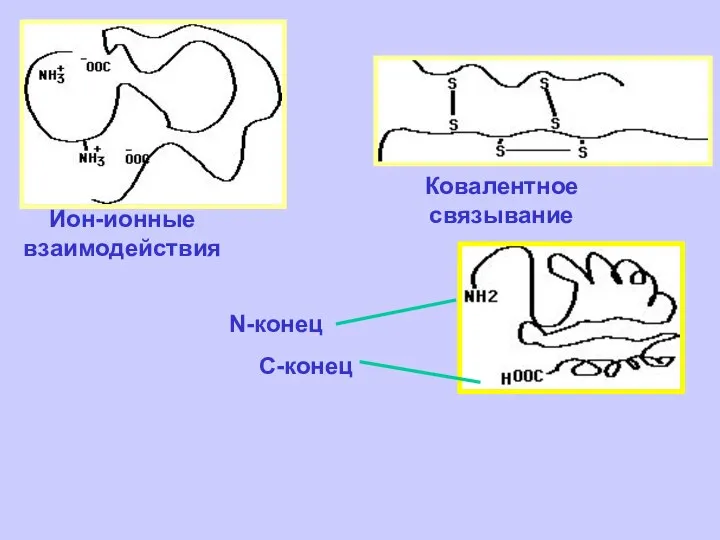
Ион-ионные
взаимодействия
Ковалентное
связывание
N-конец
С-конец
Ион-ионные
взаимодействия
Ковалентное
связывание
N-конец
С-конец
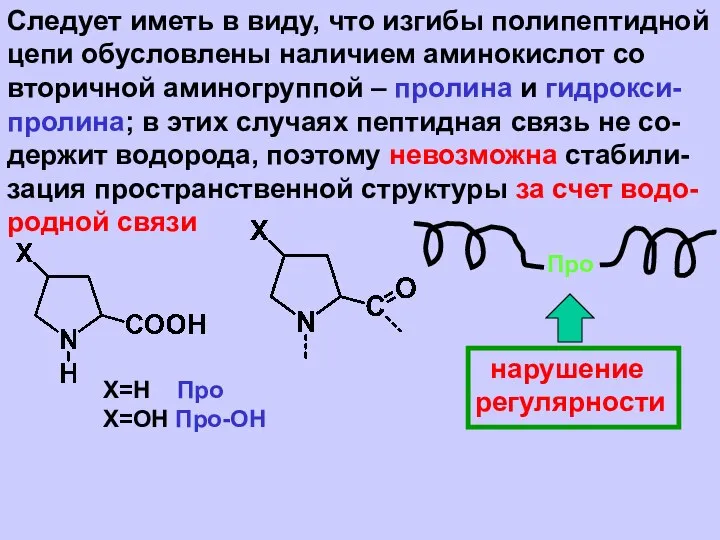
Следует иметь в виду, что изгибы полипептидной
цепи обусловлены наличием аминокислот со
вторичной
Следует иметь в виду, что изгибы полипептидной
цепи обусловлены наличием аминокислот со
вторичной

Некоторые белки, например
гемоглобин, обладают чет-
вертичной структурой; при
этом одинаковые или сход-
ные субъединицы
Некоторые белки, например
гемоглобин, обладают чет-
вертичной структурой; при
этом одинаковые или сход-
ные субъединицы
Полипептид
Полипептид
 Операторы Делегаты и события. Атрибуты. Свойства и индексаторы. Пространство имен. (Лекция 5)
Операторы Делегаты и события. Атрибуты. Свойства и индексаторы. Пространство имен. (Лекция 5) А.C. Пушкин в живописи Презентацию подготовила учитель русского языка, литературы и МХК МБОУСОШ № 119 г.Новосибирска Рябизо
А.C. Пушкин в живописи Презентацию подготовила учитель русского языка, литературы и МХК МБОУСОШ № 119 г.Новосибирска Рябизо Ауызекі сөйлеу стилі
Ауызекі сөйлеу стилі Христианство и палеохристиане
Христианство и палеохристиане Введение в объектно-ориентированное программирование
Введение в объектно-ориентированное программирование Технология изготовления и организация производства двутавровой балки
Технология изготовления и организация производства двутавровой балки Қазақ халқының қолөнері
Қазақ халқының қолөнері Презентация "Мастера земли Калевальской" - скачать презентации по МХК
Презентация "Мастера земли Калевальской" - скачать презентации по МХК грибы - презентация для начальной школы
грибы - презентация для начальной школы Бизнес-курс. Начало. Этапы прохождения курса
Бизнес-курс. Начало. Этапы прохождения курса Законы и принципы психологии
Законы и принципы психологии урок 4
урок 4 Księdzem Bosco
Księdzem Bosco Презентация на тему "Учебно-методические комплекты" - скачать презентации по Педагогике
Презентация на тему "Учебно-методические комплекты" - скачать презентации по Педагогике Выпускная квалификационная работа: Сравнительный анализ систем связи 4G и 5G
Выпускная квалификационная работа: Сравнительный анализ систем связи 4G и 5G Презентация на тему "Эпилепсия. Органические, включая симптоматические расстройства" - скачать презентации по Медицине
Презентация на тему "Эпилепсия. Органические, включая симптоматические расстройства" - скачать презентации по Медицине Презентация: Практические аспекты применения критериев достаточной переработки товара при определении его страны происхождения
Презентация: Практические аспекты применения критериев достаточной переработки товара при определении его страны происхождения  Презентация на тему "Родительское собрание будущих первоклассников" - скачать презентации по Педагогике
Презентация на тему "Родительское собрание будущих первоклассников" - скачать презентации по Педагогике Подготовка мясных грузов к доставке
Подготовка мясных грузов к доставке Игра по темам: «Основные направления и перспективы развития ТД», «Научно-технический прогресс в ТД» Выполнила: студентка 3-го
Игра по темам: «Основные направления и перспективы развития ТД», «Научно-технический прогресс в ТД» Выполнила: студентка 3-го  Сетевые протоколы
Сетевые протоколы Аэродинамика. Самолёты и планеры
Аэродинамика. Самолёты и планеры Подход компании «Сименс» к реализации цифровых сетей
Подход компании «Сименс» к реализации цифровых сетей Розробка програмної системи «Автобазар»
Розробка програмної системи «Автобазар» Правописание гласных в корне орфография
Правописание гласных в корне орфография Презентация "«Модель денег»" - скачать презентации по Экономике
Презентация "«Модель денег»" - скачать презентации по Экономике Международный кредит
Международный кредит Проект на тему: «Технологии, повышающие качество жизни школьников».
Проект на тему: «Технологии, повышающие качество жизни школьников».