- Биохимия белков и пептидов

Содержание
- 2. Кафедра биохимии, 2006 (C) Пептиды и белки: общие сведения Белки При соединении аминокислот в цепочку образуется
- 3. Кафедра биохимии, 2006 (C) Примеры белков
- 4. Кафедра биохимии, 2006 (C) Структурообразующие функции Структурные белки отвечают за поддержание формы и стабильности клеток и
- 5. Кафедра биохимии, 2006 (C) Гистоны в хроматине К структурным белкам можно отнести также гистоны, функцией которых
- 6. Кафедра биохимии, 2006 (C) Транспортные функции (гемоглобин) Наиболее известным транспортным белком является гемоглобин эритроцитов, ответственный за
- 7. Кафедра биохимии, 2006 (C) Другие транспортные белки В плазме крови содержатся множество других белков, выполняющих транспортные
- 8. Кафедра биохимии, 2006 (C) Защитные функции Иммунная система защищает организм от возбудителей болезней и чужеродных веществ.
- 9. Кафедра биохимии, 2006 (C) Регуляторные функции В биохимических сигнальных цепях белки осуществляют функции сигнальных веществ (гормонов)
- 10. Кафедра биохимии, 2006 (C) В регуляции обмена веществ и процессов дифференцировки принимают решающее участие ДНК-ассоцированиые белки
- 11. Кафедра биохимии, 2006 (C) Ферменты Среди 2000 известных белков наиболее многочисленную группу составляют ферменты. Самые низкомолекулярные
- 12. Кафедра биохимии, 2006 (C) Двигательные функции Взаимодействие актина с миозином ответственно за мышечное сокращение и другие
- 13. Кафедра биохимии, 2006 (C) Запасные функции В растениях содержатся запасные белки, являющиеся ценными пищевыми веществами. В
- 14. Кафедра биохимии, 2006 (C) Пептидная связь Главными структурными единицами белкой и пептидов являются остатки аминокислот, связанные
- 15. Кафедра биохимии, 2006 (C) Пептидный синтез В клетках пептиды и белки синтезируются в процессе трансляции на
- 16. Кафедра биохимии, 2006 (C) Мезомерия пептидной связи Как всякая карбоксамидная связь, пептидная связь стабилизирована за счет
- 17. Кафедра биохимии, 2006 (C) Номенклатура пептидов Пептидная цепь имеет одно направление и два разных конца —
- 18. Кафедра биохимии, 2006 (C) Конформация полипептидной цепи Каждый аминокислотный остаток, за исключением концевых, принимает участие в
- 19. Кафедра биохимии, 2006 (C) Вторичные структуры белков Вторичные структуры стабилизированы водородными мостиками в пределах одной пептидной
- 20. Кафедра биохимии, 2006 (C) α-Спираль Наиболее распространенным элементом вторичной структуры является правая α-спираль (αR). Пептидная цепь
- 21. Кафедра биохимии, 2006 (C) Спираль коллагена Другая форма спирали присутствует в коллагене, важнейшем компоненте соединительных тканей.
- 22. Кафедра биохимии, 2006 (C) Складчатые структуры Две следующие почти вытянутые конформации пептидной цепи называются "β-складчатым листом",
- 23. Кафедра биохимии, 2006 (C) Складчатые структуры (α и β) В складчатых структурах α-С-атомы располагаются на перегибах,
- 24. Кафедра биохимии, 2006 (C) β-Петля В тех участках, где пептидная цепь изгибается достаточно круто, часто находится
- 25. Кафедра биохимии, 2006 (C) Структурные белки Структурным белком, построенным преимущественно в виде α-спирали, является α-кератин. Волосы
- 26. Кафедра биохимии, 2006 (C) Фиброин Ocновной белок шелка, фиброин, обладает структурой антипараллельного складчатого листа, причем сами
- 27. Кафедра биохимии, 2006 (C) Коллаген В организме млекопитающих коллаген — преобладающий в количественном отношении белок: он
- 28. Кафедра биохимии, 2006 (C) Глобулярные белки В отличие от нерастворимых фибриллярных белков растворимые белки имеют почти
- 29. Кафедра биохимии, 2006 (C) Инсулин: первичная структура Под первичной структурой понимают аминокислотную последовательность полипептидной цепи. Инсулин
- 30. Кафедра биохимии, 2006 (C) Инсулин: вторичная структура Вторичными структурами называются участки полипептидной цепи с упорядоченной конформацией,
- 31. Кафедра биохимии, 2006 (C) Инсулин. Третичная структура Трехмерные функционально активные конформации белков носят название третичной структуры.
- 32. Кафедра биохимии, 2006 (C) Инсулин: анализ третичной структуры Анализ третичной структуры инсулина показал, что в Α-цепи
- 33. Кафедра биохимии, 2006 (C) Инсулин. Четвертичная структура Белковые молекулы часто образуют симметрично построенные комплексы, стабилизированные за
- 34. Кафедра биохимии, 2006 (C) Свертывание белков При сравнении наиболее крупных глобулярных белков становится очевидным, что существует
- 35. Кафедра биохимии, 2006 (C) Свертывание белков: примеры Рассмотрим ряд примеров (α-спирали выделены красным цветом, плоскости складчатого
- 36. Кафедра биохимии, 2006 (C) Методы выделения и анализа белков Препараты высокоочищенных белков находят разнообразное применение в
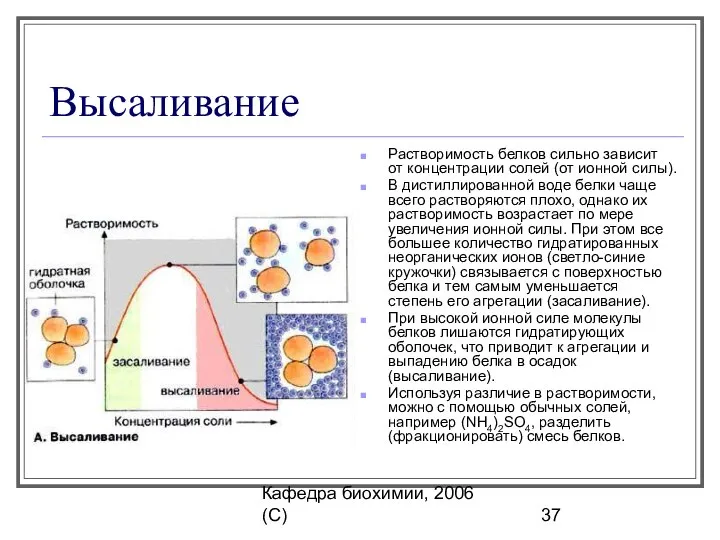
- 37. Кафедра биохимии, 2006 (C) Высаливание Растворимость белков сильно зависит от концентрации солей (от ионной силы). В
- 38. Кафедра биохимии, 2006 (C) Диализ Для отделения низкомолекулярных примесей или замены состава среды используют диализ. Метод
- 39. Кафедра биохимии, 2006 (C) Гель-фильтрация Гель-проникающая хроматография (гель-фильтрация) позволяет разделять белки по величине и форме молекул.
- 40. Кафедра биохимии, 2006 (C) Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия Метод основан на свойстве
- 41. Кафедра биохимии, 2006 (C) Под действием ДСН олигомерные белки диссоциируют на субъединицы и денатурируют. Развернутые полипептидные
- 42. Кафедра биохимии, 2006 (C) Электрофореграммы Электрофорез проводят в тонком слое полиакриламида (2). После завершения электрофореза, зоны
- 43. Кафедра биохимии, 2006 (C)
- 44. Кафедра биохимии, 2006 (C)
- 45. Кафедра биохимии, 2006 (C)
- 46. Кафедра биохимии, 2006 (C)
- 47. Кафедра биохимии, 2006 (C)
- 48. Кафедра биохимии, 2006 (C)
- 49. Кафедра биохимии, 2006 (C)
- 50. Кафедра биохимии, 2006 (C)
- 51. Кафедра биохимии, 2006 (C)
- 53. Скачать презентацию

Кафедра биохимии, 2006 (C)
Пептиды и белки: общие сведения
Белки
При соединении аминокислот в
Кафедра биохимии, 2006 (C)
Пептиды и белки: общие сведения
Белки
При соединении аминокислот в
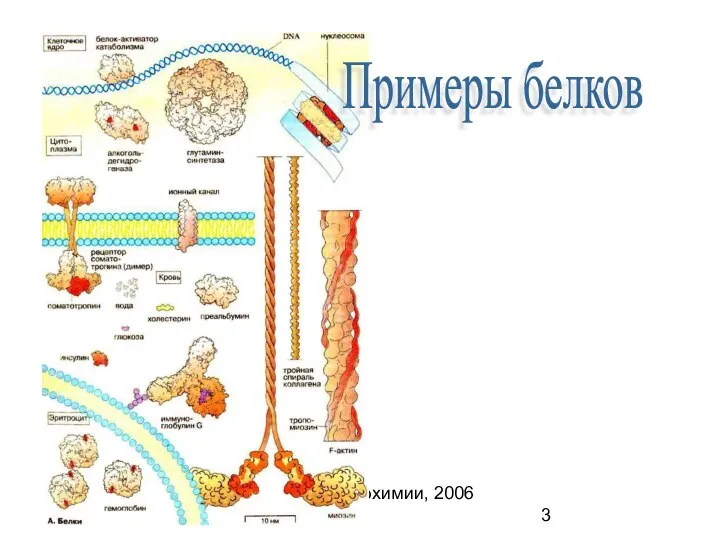
Кафедра биохимии, 2006 (C)
Примеры белков
Кафедра биохимии, 2006 (C)
Примеры белков

Кафедра биохимии, 2006 (C)
Структурообразующие функции
Структурные белки отвечают за поддержание формы и
Кафедра биохимии, 2006 (C)
Структурообразующие функции
Структурные белки отвечают за поддержание формы и
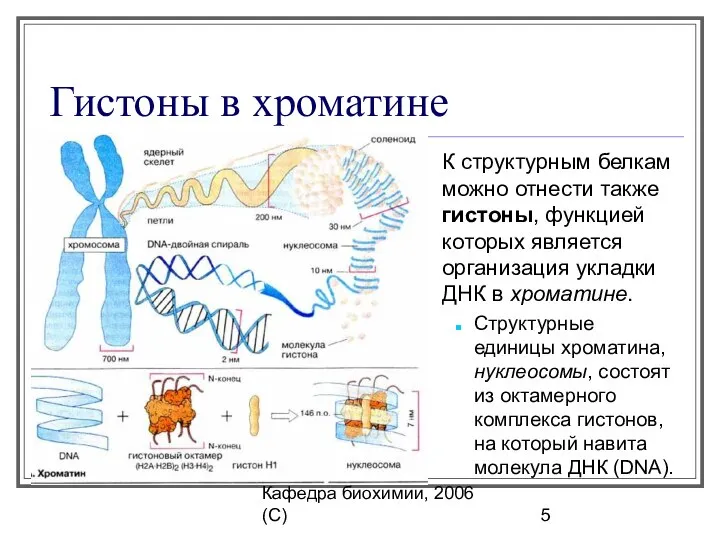
Кафедра биохимии, 2006 (C)
Гистоны в хроматине
К структурным белкам можно отнести также
Кафедра биохимии, 2006 (C)
Гистоны в хроматине
К структурным белкам можно отнести также
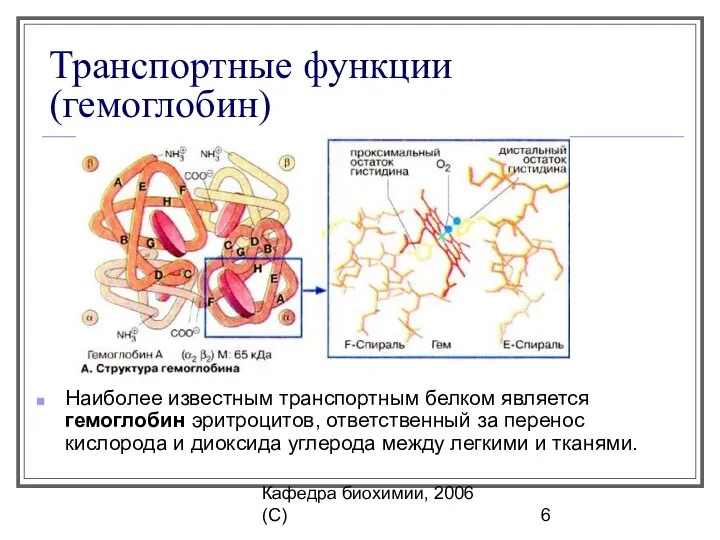
Кафедра биохимии, 2006 (C)
Транспортные функции (гемоглобин)
Наиболее известным транспортным белком является гемоглобин
Кафедра биохимии, 2006 (C)
Транспортные функции (гемоглобин)
Наиболее известным транспортным белком является гемоглобин
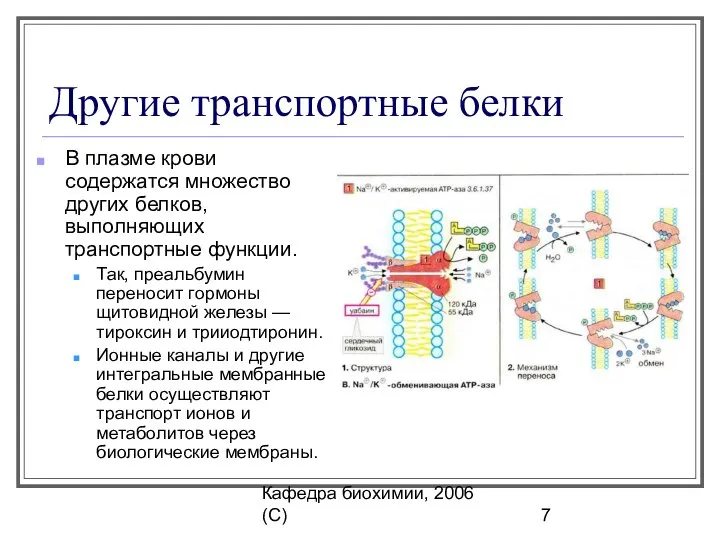
Кафедра биохимии, 2006 (C)
Другие транспортные белки
В плазме крови содержатся множество других
Кафедра биохимии, 2006 (C)
Другие транспортные белки
В плазме крови содержатся множество других
Кафедра биохимии, 2006 (C)
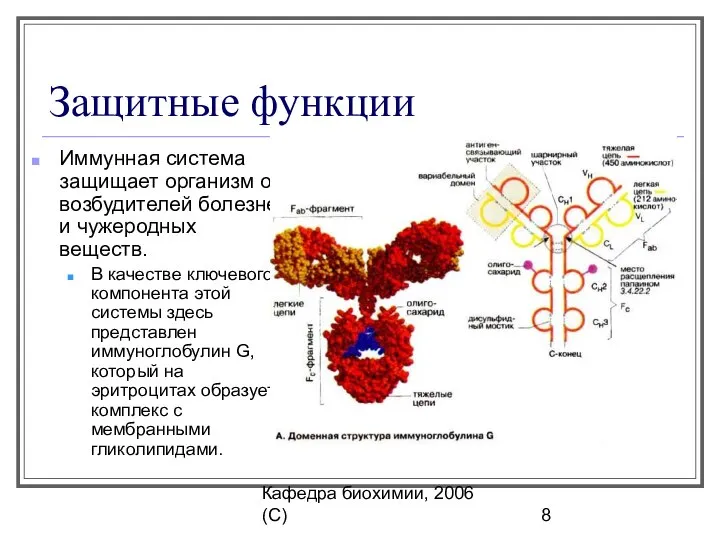
Защитные функции
Иммунная система защищает организм от возбудителей болезней
Кафедра биохимии, 2006 (C)
Защитные функции
Иммунная система защищает организм от возбудителей болезней
Кафедра биохимии, 2006 (C)
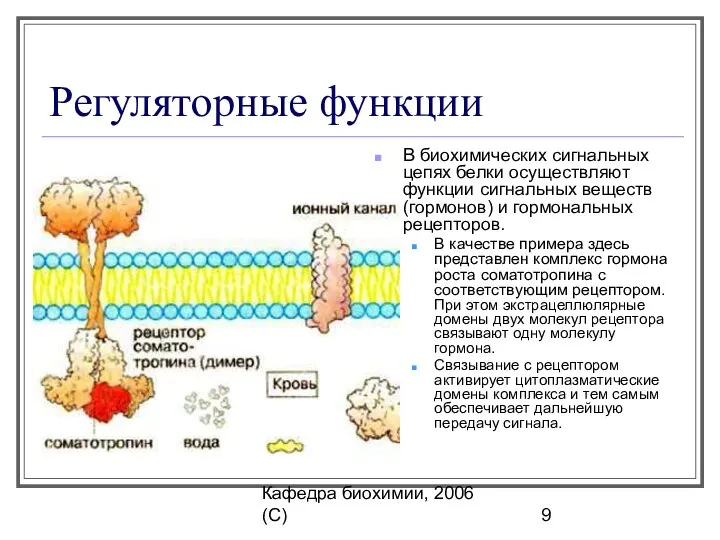
Регуляторные функции
В биохимических сигнальных цепях белки осуществляют функции
Кафедра биохимии, 2006 (C)
Регуляторные функции
В биохимических сигнальных цепях белки осуществляют функции
Кафедра биохимии, 2006 (C)
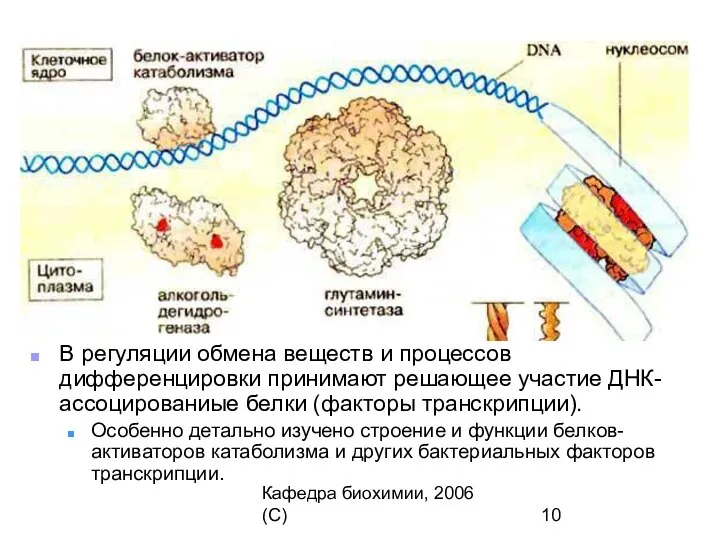
В регуляции обмена веществ и процессов дифференцировки принимают
Кафедра биохимии, 2006 (C)
В регуляции обмена веществ и процессов дифференцировки принимают
Кафедра биохимии, 2006 (C)
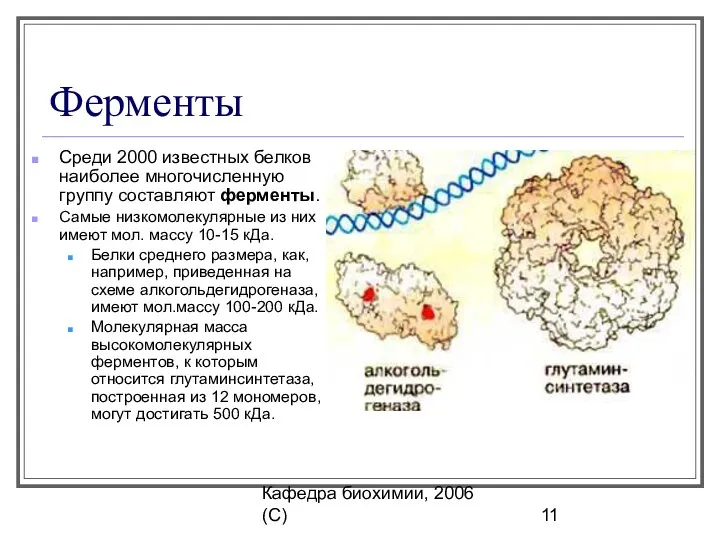
Ферменты
Среди 2000 известных белков наиболее многочисленную группу составляют
Кафедра биохимии, 2006 (C)
Ферменты
Среди 2000 известных белков наиболее многочисленную группу составляют

Кафедра биохимии, 2006 (C)
Двигательные функции
Взаимодействие актина с миозином ответственно за мышечное
Кафедра биохимии, 2006 (C)
Двигательные функции
Взаимодействие актина с миозином ответственно за мышечное

Кафедра биохимии, 2006 (C)
Запасные функции
В растениях содержатся запасные белки, являющиеся ценными
Кафедра биохимии, 2006 (C)
Запасные функции
В растениях содержатся запасные белки, являющиеся ценными

Кафедра биохимии, 2006 (C)
Пептидная связь
Главными структурными единицами белкой и пептидов являются
Кафедра биохимии, 2006 (C)
Пептидная связь
Главными структурными единицами белкой и пептидов являются
Кафедра биохимии, 2006 (C)
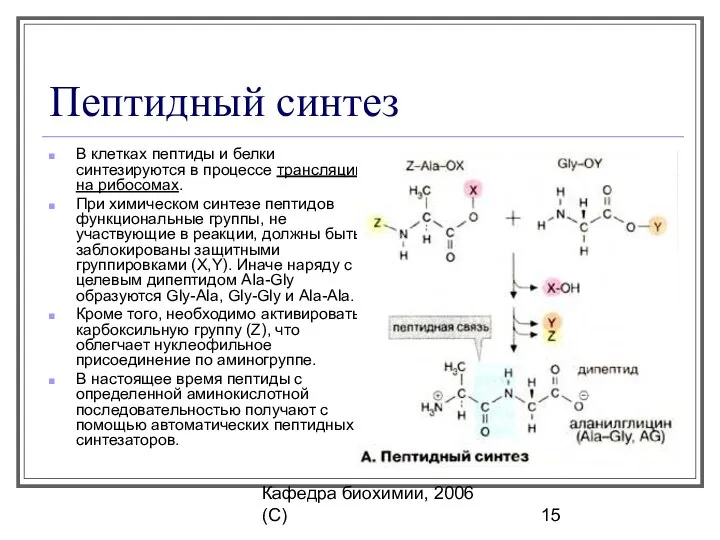
Пептидный синтез
В клетках пептиды и белки синтезируются в
Кафедра биохимии, 2006 (C)
Пептидный синтез
В клетках пептиды и белки синтезируются в
Кафедра биохимии, 2006 (C)
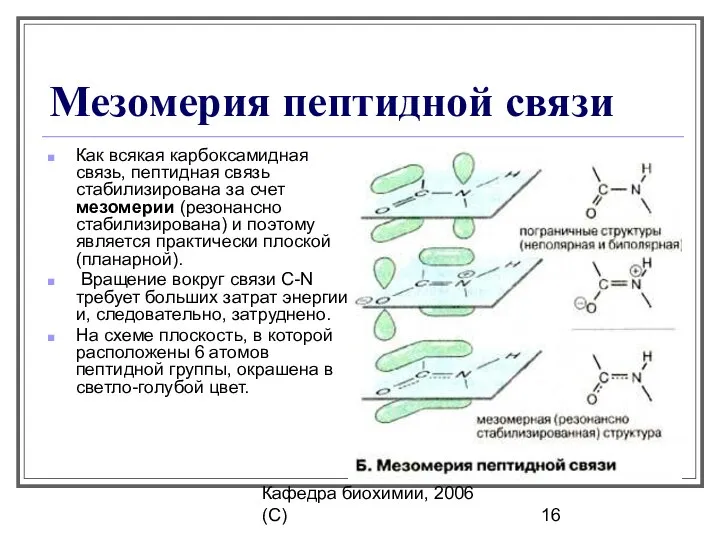
Мезомерия пептидной связи
Как всякая карбоксамидная связь, пептидная связь
Кафедра биохимии, 2006 (C)
Мезомерия пептидной связи
Как всякая карбоксамидная связь, пептидная связь
Кафедра биохимии, 2006 (C)
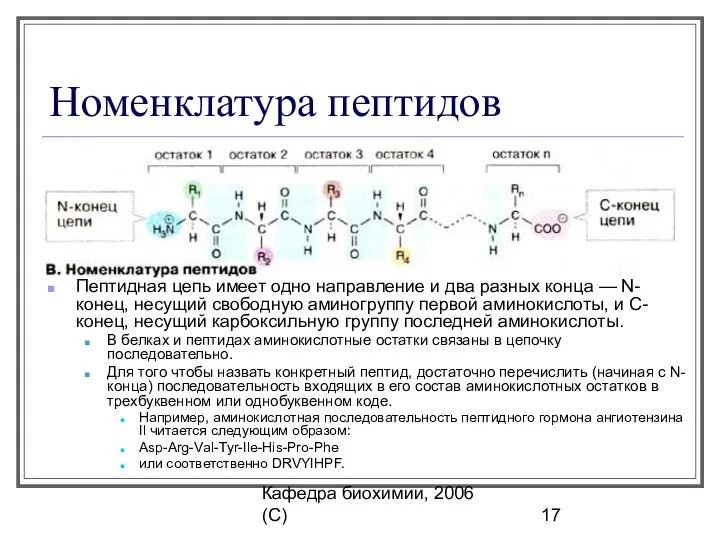
Номенклатура пептидов
Пептидная цепь имеет одно направление и два
Кафедра биохимии, 2006 (C)
Номенклатура пептидов
Пептидная цепь имеет одно направление и два

Кафедра биохимии, 2006 (C)
Конформация полипептидной цепи
Каждый аминокислотный остаток, за исключением концевых,
Кафедра биохимии, 2006 (C)
Конформация полипептидной цепи
Каждый аминокислотный остаток, за исключением концевых,

Кафедра биохимии, 2006 (C)
Вторичные структуры белков
Вторичные структуры стабилизированы водородными мостиками в
Кафедра биохимии, 2006 (C)
Вторичные структуры белков
Вторичные структуры стабилизированы водородными мостиками в
Кафедра биохимии, 2006 (C)
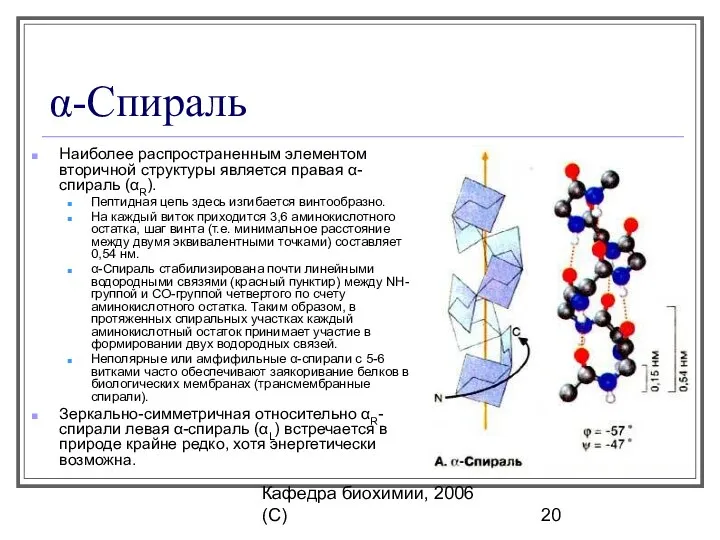
α-Спираль
Наиболее распространенным элементом вторичной структуры является правая α-спираль
Кафедра биохимии, 2006 (C)
α-Спираль
Наиболее распространенным элементом вторичной структуры является правая α-спираль
Кафедра биохимии, 2006 (C)
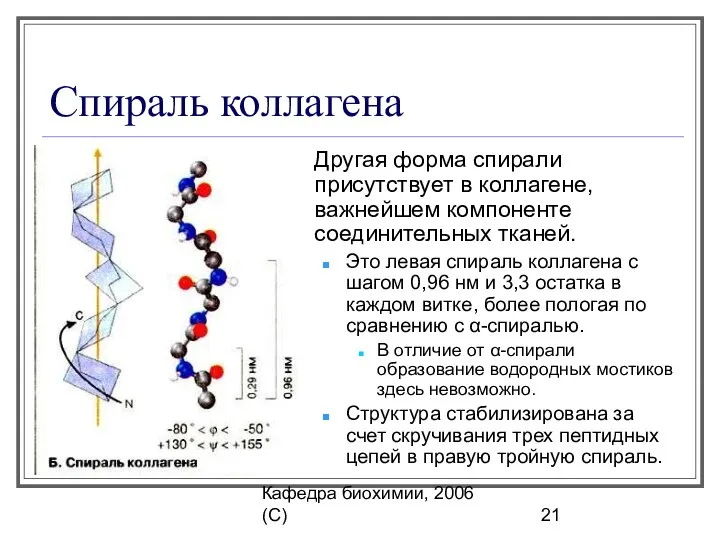
Спираль коллагена
Другая форма спирали присутствует в коллагене, важнейшем
Кафедра биохимии, 2006 (C)
Спираль коллагена
Другая форма спирали присутствует в коллагене, важнейшем

Кафедра биохимии, 2006 (C)
Складчатые структуры
Две следующие почти вытянутые конформации пептидной цепи
Кафедра биохимии, 2006 (C)
Складчатые структуры
Две следующие почти вытянутые конформации пептидной цепи
Кафедра биохимии, 2006 (C)
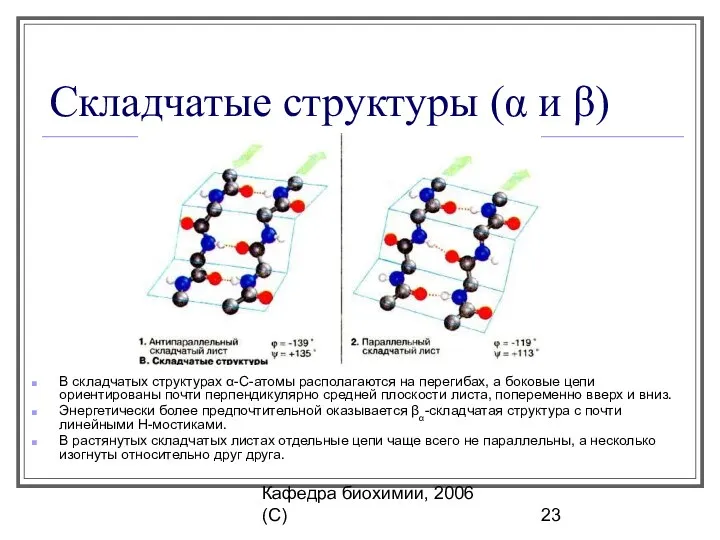
Складчатые структуры (α и β)
В складчатых структурах α-С-атомы
Кафедра биохимии, 2006 (C)
Складчатые структуры (α и β)
В складчатых структурах α-С-атомы
Кафедра биохимии, 2006 (C)
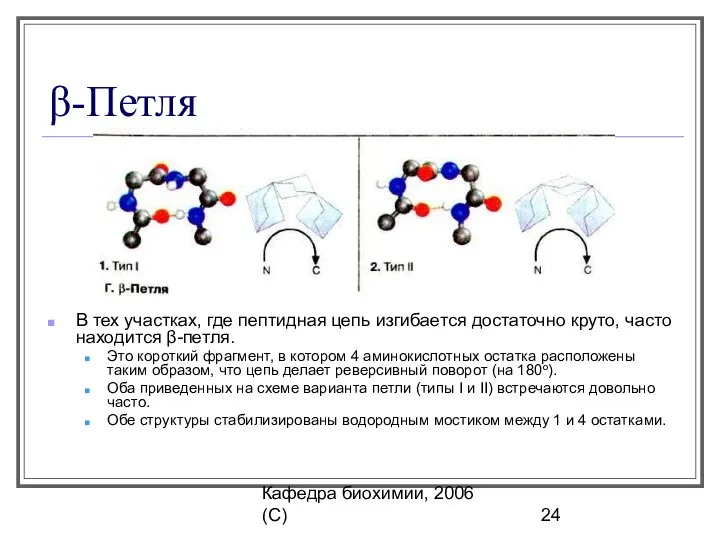
β-Петля
В тех участках, где пептидная цепь изгибается достаточно
Кафедра биохимии, 2006 (C)
β-Петля
В тех участках, где пептидная цепь изгибается достаточно
Кафедра биохимии, 2006 (C)
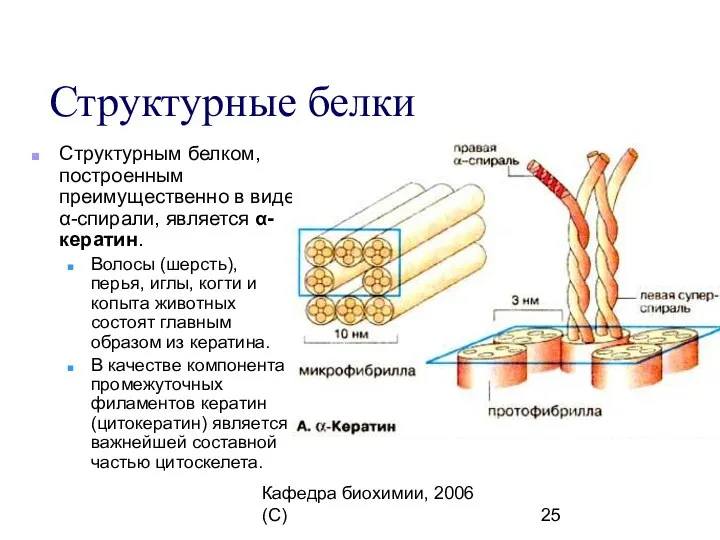
Структурные белки
Структурным белком, построенным преимущественно в виде α-спирали,
Кафедра биохимии, 2006 (C)
Структурные белки
Структурным белком, построенным преимущественно в виде α-спирали,

Кафедра биохимии, 2006 (C)
Фиброин
Ocновной белок шелка, фиброин, обладает структурой антипараллельного складчатого
Кафедра биохимии, 2006 (C)
Фиброин
Ocновной белок шелка, фиброин, обладает структурой антипараллельного складчатого

Кафедра биохимии, 2006 (C)
Коллаген
В организме млекопитающих коллаген — преобладающий в количественном
Кафедра биохимии, 2006 (C)
Коллаген
В организме млекопитающих коллаген — преобладающий в количественном

Кафедра биохимии, 2006 (C)
Глобулярные белки
В отличие от нерастворимых фибриллярных белков растворимые
Кафедра биохимии, 2006 (C)
Глобулярные белки
В отличие от нерастворимых фибриллярных белков растворимые
Кафедра биохимии, 2006 (C)
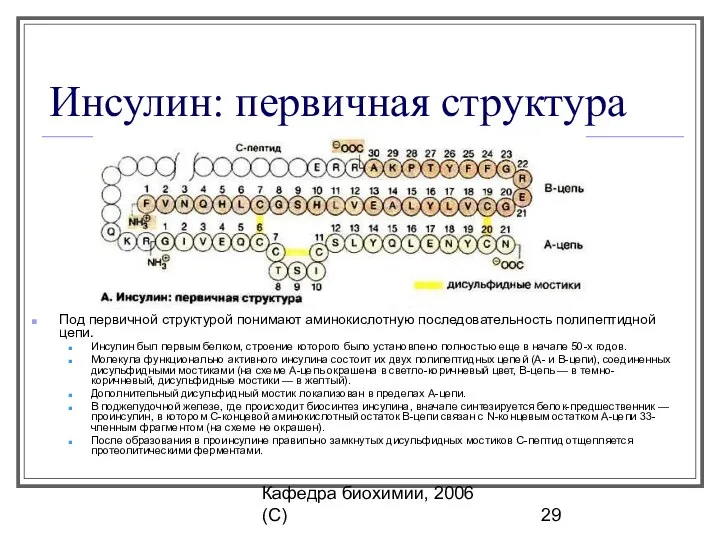
Инсулин: первичная структура
Под первичной структурой понимают аминокислотную последовательность
Кафедра биохимии, 2006 (C)
Инсулин: первичная структура
Под первичной структурой понимают аминокислотную последовательность
Кафедра биохимии, 2006 (C)
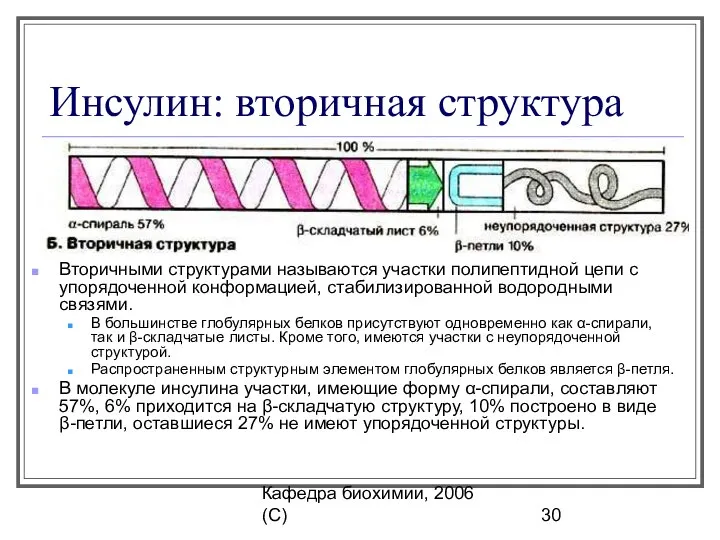
Инсулин: вторичная структура
Вторичными структурами называются участки полипептидной цепи
Кафедра биохимии, 2006 (C)
Инсулин: вторичная структура
Вторичными структурами называются участки полипептидной цепи
Кафедра биохимии, 2006 (C)
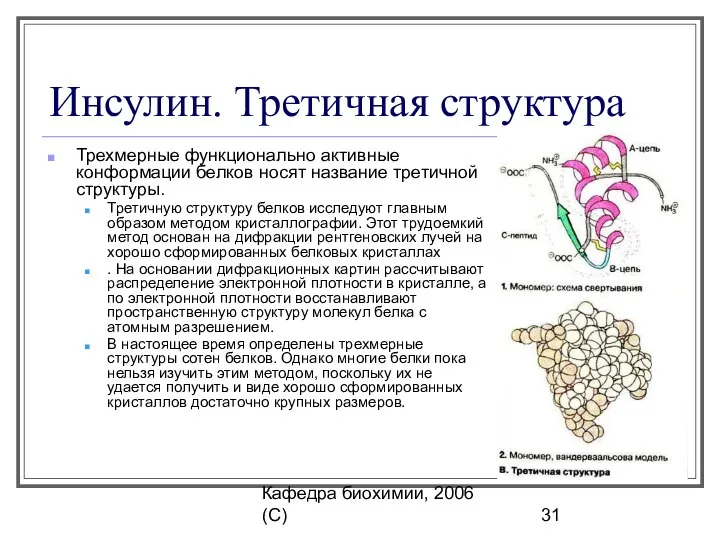
Инсулин. Третичная структура
Трехмерные функционально активные конформации белков носят
Кафедра биохимии, 2006 (C)
Инсулин. Третичная структура
Трехмерные функционально активные конформации белков носят

Кафедра биохимии, 2006 (C)
Инсулин: анализ третичной структуры
Анализ третичной структуры инсулина показал,
Кафедра биохимии, 2006 (C)
Инсулин: анализ третичной структуры
Анализ третичной структуры инсулина показал,
Кафедра биохимии, 2006 (C)
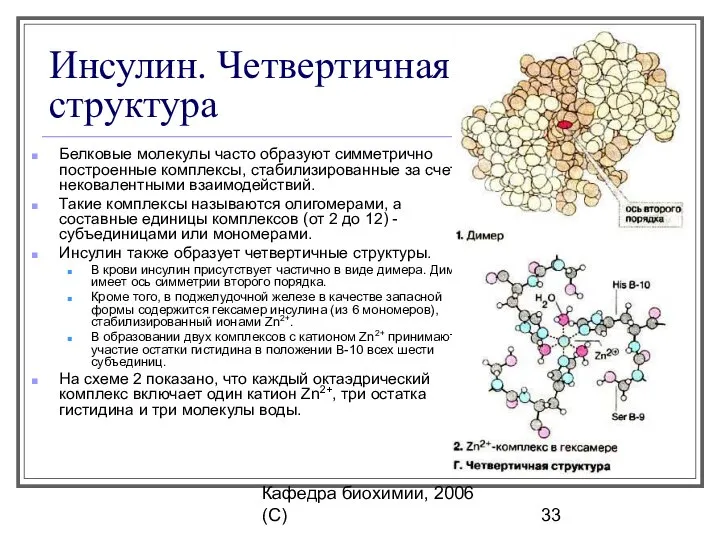
Инсулин. Четвертичная структура
Белковые молекулы часто образуют симметрично построенные
Кафедра биохимии, 2006 (C)
Инсулин. Четвертичная структура
Белковые молекулы часто образуют симметрично построенные

Кафедра биохимии, 2006 (C)
Свертывание белков
При сравнении наиболее крупных глобулярных белков становится
Кафедра биохимии, 2006 (C)
Свертывание белков
При сравнении наиболее крупных глобулярных белков становится

Кафедра биохимии, 2006 (C)
Свертывание белков: примеры
Рассмотрим ряд примеров (α-спирали выделены красным
Кафедра биохимии, 2006 (C)
Свертывание белков: примеры
Рассмотрим ряд примеров (α-спирали выделены красным

Кафедра биохимии, 2006 (C)
Методы выделения и анализа белков
Препараты высокоочищенных белков находят
Кафедра биохимии, 2006 (C)
Методы выделения и анализа белков
Препараты высокоочищенных белков находят
Кафедра биохимии, 2006 (C)
Высаливание
Растворимость белков сильно зависит от концентрации солей (от
Кафедра биохимии, 2006 (C)
Высаливание
Растворимость белков сильно зависит от концентрации солей (от

Кафедра биохимии, 2006 (C)
Диализ
Для отделения низкомолекулярных примесей или замены состава среды
Кафедра биохимии, 2006 (C)
Диализ
Для отделения низкомолекулярных примесей или замены состава среды

Кафедра биохимии, 2006 (C)
Гель-фильтрация
Гель-проникающая хроматография (гель-фильтрация) позволяет разделять белки по величине
Кафедра биохимии, 2006 (C)
Гель-фильтрация
Гель-проникающая хроматография (гель-фильтрация) позволяет разделять белки по величине

Кафедра биохимии, 2006 (C)
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия
Метод
Кафедра биохимии, 2006 (C)
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия
Метод

Кафедра биохимии, 2006 (C)
Под действием ДСН олигомерные белки диссоциируют на субъединицы
Кафедра биохимии, 2006 (C)
Под действием ДСН олигомерные белки диссоциируют на субъединицы
Кафедра биохимии, 2006 (C)
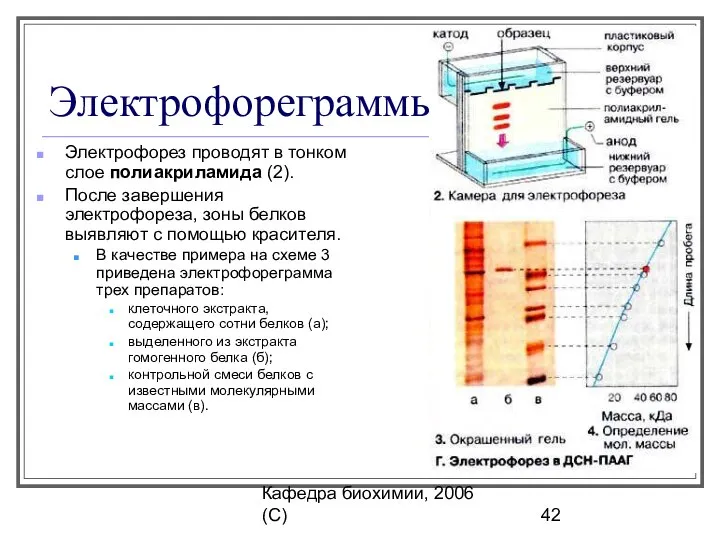
Электрофореграммы
Электрофорез проводят в тонком слое полиакриламида (2).
После
Кафедра биохимии, 2006 (C)
Электрофореграммы
Электрофорез проводят в тонком слое полиакриламида (2).
После
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
Кафедра биохимии, 2006 (C)
 БАЗЫ ДАННЫХ. ACCESS 2007 Запросы
БАЗЫ ДАННЫХ. ACCESS 2007 Запросы Ch1.ppt
Ch1.ppt Добро пожаловать. ETHEREUMPRO
Добро пожаловать. ETHEREUMPRO Требования к строительным машинам
Требования к строительным машинам Классификация персональных компьютеров
Классификация персональных компьютеров Аттестация социальных работников
Аттестация социальных работников МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ АВТОНОМНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕ
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ АВТОНОМНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕ Фруктово-овощной ребрейдинг остановок
Фруктово-овощной ребрейдинг остановок Финансы хозяйствующих субъектов
Финансы хозяйствующих субъектов Лекция 03 - Рак желудка (слайды).ppt
Лекция 03 - Рак желудка (слайды).ppt Подвеска колес автомобиля
Подвеска колес автомобиля Эвристические комбинаторные алгоритмы
Эвристические комбинаторные алгоритмы Презентация "Сравнительный подход к оценке недвижимости" - скачать презентации по Экономике
Презентация "Сравнительный подход к оценке недвижимости" - скачать презентации по Экономике Понятие и специфические признаки гражданского правоотношения Выполнил: Трушко Павел олегович
Понятие и специфические признаки гражданского правоотношения Выполнил: Трушко Павел олегович  Гостиничная сеть Choice Hotels International
Гостиничная сеть Choice Hotels International  Порядок образования туристской фирмы
Порядок образования туристской фирмы Технология отделочных работ и систем КНАУФ
Технология отделочных работ и систем КНАУФ Classification of computer
Classification of computer Презентация "Бюджет государства" - скачать презентации по Экономике
Презентация "Бюджет государства" - скачать презентации по Экономике ФИЗИЧЕСКАЯ КУЛЬТУРА КАК ВИД КУЛЬТУРЫ ОБЩЕСТВА
ФИЗИЧЕСКАЯ КУЛЬТУРА КАК ВИД КУЛЬТУРЫ ОБЩЕСТВА Программирование циклов
Программирование циклов Александр Порфирьевич Бородин
Александр Порфирьевич Бородин Достоинства человека
Достоинства человека  2. Java Spring Core 2. Maven
2. Java Spring Core 2. Maven Возникновение ислама
Возникновение ислама ОРГАНИЗАЦИЯ СПЕЦИАЛЬНЫХ СОБЫТИЙ: Церемонии открытия Приемы
ОРГАНИЗАЦИЯ СПЕЦИАЛЬНЫХ СОБЫТИЙ: Церемонии открытия Приемы  Без названия
Без названия «Социальные нормы и отклоняющееся поведение» Учитель: Чистякова Н.Ф.
«Социальные нормы и отклоняющееся поведение» Учитель: Чистякова Н.Ф.