- Биохимия мышц

Содержание
- 2. План: Морфология поперечно-полосатой мышцы. Мышечные белки. Небелковые вещества ткани мышц. Функциональная биохимия мышц. Механизм мышечного сокращения
- 3. Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора (реже гормона), проявляющаяся в уменьшении длины клетки.
- 4. Мышечная ткань составляет 40–42% от массы тела. При сокращении мышц осуществляется работа, связанная с превращением химической
- 5. МОРФОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ МЫШЦЫ Поперечно-полосатая мышца состоит из многочисленных удлиненных волокон, или мышечных клеток . Двигательные
- 6. Структура волокна скелетной мышцы I - А-диск; II - I-диск; III - Н-зона; 1 - Z-линия;
- 7. Повторяющийся элемент поперечно-полосатой миофибриллы - саркомер , ограниченный узкими Z-линиями. Миофибрилла состоит из нескольких сот саркомеров.
- 8. В мышечной ткани взрослых животных и человека содержится от 72 до 80% воды. Около 20–28% от
- 9. Мышечные белки. Белки мышечной ткани делят на три основные группы: миофибриллярные – 45% саркоплазматические -35% белки
- 10. Важнейшие белки миофибрилл: Сократительные – миозин, актин, актомиозин. Регуляторные- тропомиозин, тропонин.
- 11. Миозин – сократительный белок, составляет 50–55% от сухой массы миофибрилл, обладает АТФ-азной активностью, т.е. способностью катализировать
- 12. Молекула миозина имеет сильно вытянутую форму, длину 150 нм. Она может быть расщеплена на субъединицы: две
- 13. Актин – сократительный белок, составляет 20% от сухой массы миофибрилл. Известны две формы актина: глобулярный актин
- 14. Актомиозин -образуется при соединении миозина с F-актином., обладает АТФ-азной активностью. Актомиозин активируется ионами Mg2+ и ингибируется
- 15. Тропонин — регуляторный глобулярный белок. Составляет 2% от всех миофибриллярных белков. Содержится в скелетных мышцах и
- 16. В биохимии тропонин используется как диагностический маркер заболеваний миокарда. Два вида тропонинов - тропонин-I и тропонин-T
- 17. Тропомиозин — регуляторный фибриллярный белок (70 кДа). Состоит из двух перевитых α-спиралей. Тропомиозин связывается в единый
- 18. Белки саркоплазмы - протеины, растворимые в солевых средах с низкой ионной силой. К числу саркоплазматических белков
- 19. Миоглобин. Молекула миоглобина образована единичной полипептидной цепью (153 АК) и железосодержащим гемом, сходна по строению и
- 20. Белки стромы в поперечно-полосатой мускулатуре представлены в основном коллагеном и эластином.
- 21. Небелковые вещества мышечной ткани.
- 22. Кроме белков, в состав мышц входят азотистые небелковые вещества, безазотистые органические вещества и минеральные соли. Азотистые
- 23. Карнозин и ансерин увеличивают амплитуду мышечного сокращения, которая предварительно была снижена утомлением, путем повышения эффективности работы
- 24. Безазотистые органические вещества . представлены углеводами и липидами. Гликоген — до 2%, Свободная глюкоза —в следовых
- 25. Минеральные соли . 0,1 - 1,5% мышечной массы , представлены различными ионами. Из катионов преобладают К
- 26. Функциональная биохимия мышц. Основная функция мышц - движение( т.е. сокращение и расслабление). При сокращении мышц осуществляется
- 27. Источники энергии для мышечных сокращений. Креатинфосфат и АТФ –высокоэнергетические соединения мышц. Креатинфосфат содержится преимущественно в возбудимых
- 28. Концентрация креатинфосфата в покоящейся мышечной ткани в 3-8 раз превышает концентрацию АТФ, что позволяет компенсировать расход
- 29. Некоторое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции
- 30. Запасы креатинфосфата в мышце невелики, расход его постоянно возмещается синтезом АТФ. Для любой ткани есть два
- 31. Креатинфосфат в мышечной ткани способен выполнять не только роль депо фосфатных групп, но и роль транспортной
- 32. При умеренной нагрузке мышца может покрывать свои энергетические затраты за счет аэробных реакций. При больших нагрузках,
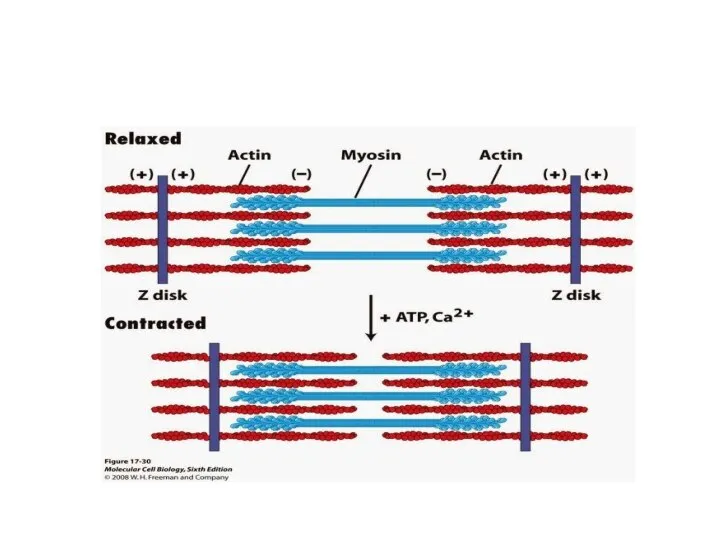
- 33. Механизм мышечного сокращения. Скольжение миозина относительно актина. Головки миозина расщепляют АТФ и за счет высвобождающейся энергии
- 36. - Источником энергии для мышечного сокращения являются молекулы АТФ. - Гидролиз АТФ катализируется при мышечном сокращении
- 37. В продвижении актиновых нитей вдоль миозиновых важную роль играют временно замыкающиеся между нитями поперечные мостики, которые
- 38. Биохимические изменения в мышцах при патологии Общими для большинства заболеваний мышц (мышечные дистрофии, атрофия, полимиозит, некоторые
- 39. Для многих форм патологии мышечной ткани характерны нарушение метаболизма креатина и его усиленное выделение с мочой
- 40. Миокард. Содержание АТФ и креатинфосфата в сердечной мышце ниже, чем в скелетной мускулатуре, а расход АТФ
- 41. Нарушение метаболизма сердечной мышцы при ишемической болезни При ишемии миокарда снижено окислительное фосфорилирование и повышен анаэробный
- 42. Размер повреждения миокарда при возникновении ишемии, можно оценить по активности ферментов сыворотки крови- Креатинкиназа -. Содержится
- 44. Скачать презентацию

План:
Морфология поперечно-полосатой мышцы.
Мышечные белки.
Небелковые вещества ткани мышц.
Функциональная биохимия мышц.
Механизм мышечного сокращения
Биохимические
План:
Морфология поперечно-полосатой мышцы.
Мышечные белки.
Небелковые вещества ткани мышц.
Функциональная биохимия мышц.
Механизм мышечного сокращения
Биохимические

Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора (реже гормона), проявляющаяся в уменьшении длины клетки.
Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора (реже гормона), проявляющаяся в уменьшении длины клетки.

Мышечная ткань составляет 40–42% от массы тела. При сокращении мышц осуществляется работа, связанная
Мышечная ткань составляет 40–42% от массы тела. При сокращении мышц осуществляется работа, связанная

МОРФОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ МЫШЦЫ
Поперечно-полосатая мышца состоит из многочисленных удлиненных волокон, или
МОРФОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ МЫШЦЫ Поперечно-полосатая мышца состоит из многочисленных удлиненных волокон, или
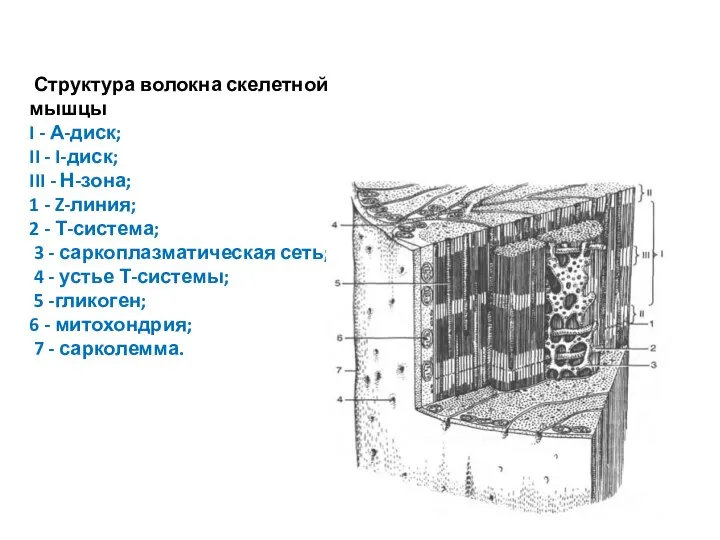
Структура волокна скелетной мышцы
I - А-диск;
II - I-диск;
III
Структура волокна скелетной мышцы I - А-диск; II - I-диск; III
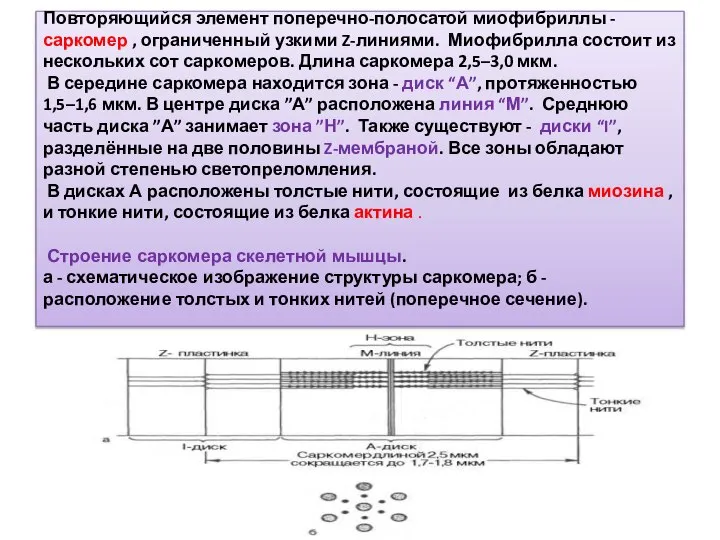
Повторяющийся элемент поперечно-полосатой миофибриллы - саркомер , ограниченный узкими Z-линиями. Миофибрилла
Повторяющийся элемент поперечно-полосатой миофибриллы - саркомер , ограниченный узкими Z-линиями. Миофибрилла
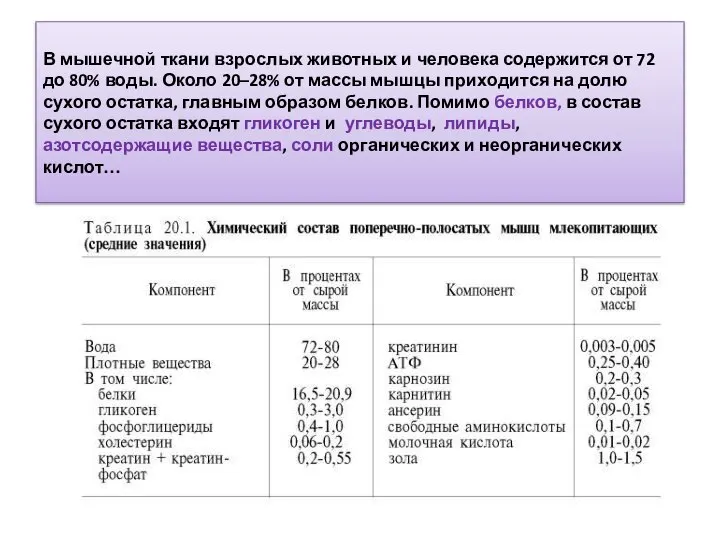
В мышечной ткани взрослых животных и человека содержится от 72 до 80% воды.
В мышечной ткани взрослых животных и человека содержится от 72 до 80% воды.

Мышечные белки.
Белки мышечной ткани делят на три основные группы:
миофибриллярные – 45%
саркоплазматические
Мышечные белки. Белки мышечной ткани делят на три основные группы: миофибриллярные – 45% саркоплазматические

Важнейшие белки миофибрилл:
Сократительные – миозин, актин, актомиозин.
Регуляторные- тропомиозин, тропонин.
Важнейшие белки миофибрилл:
Сократительные – миозин, актин, актомиозин.
Регуляторные- тропомиозин, тропонин.
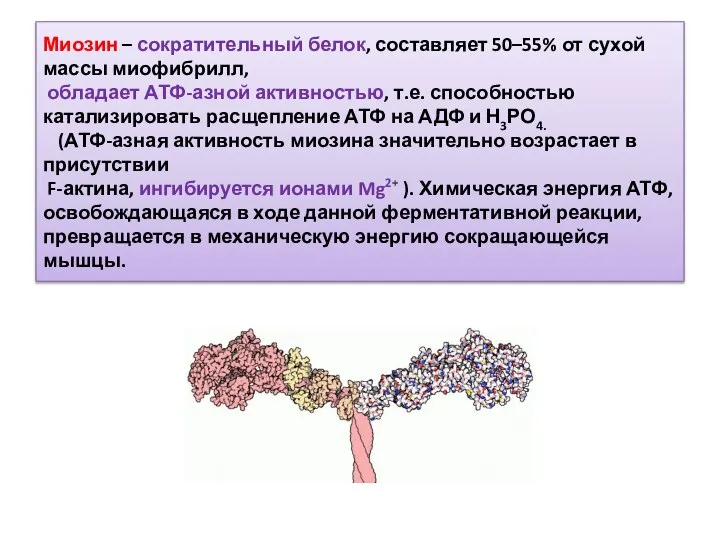
Миозин – сократительный белок, составляет 50–55% от сухой массы миофибрилл,
обладает
Миозин – сократительный белок, составляет 50–55% от сухой массы миофибрилл, обладает

Молекула миозина имеет сильно вытянутую форму, длину 150 нм. Она может быть
Молекула миозина имеет сильно вытянутую форму, длину 150 нм. Она может быть
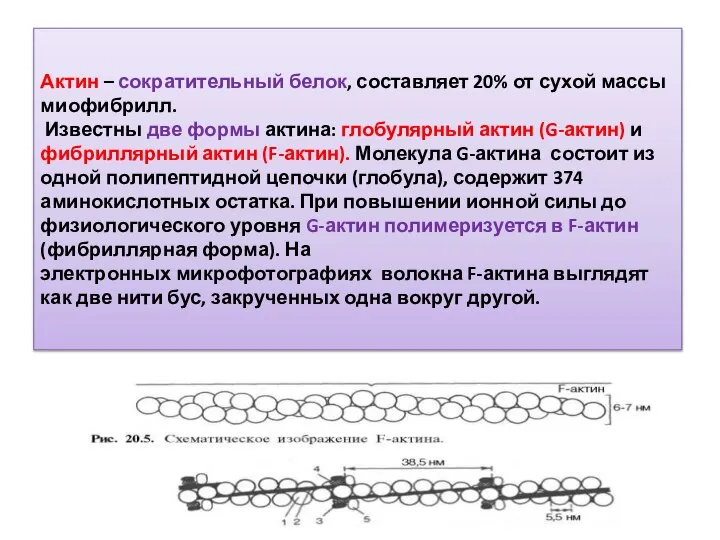
Актин – сократительный белок, составляет 20% от сухой массы миофибрилл. Известны
Актин – сократительный белок, составляет 20% от сухой массы миофибрилл. Известны

Актомиозин -образуется при соединении миозина с F-актином., обладает АТФ-азной активностью.
Актомиозин активируется ионами Mg2+ и ингибируется высокой концентрацией АТФ.
Актомиозин -образуется при соединении миозина с F-актином., обладает АТФ-азной активностью.
Актомиозин активируется ионами Mg2+ и ингибируется высокой концентрацией АТФ.

Тропонин — регуляторный глобулярный белок. Составляет 2% от всех миофибриллярных белков. Содержится в скелетных
Тропонин — регуляторный глобулярный белок. Составляет 2% от всех миофибриллярных белков. Содержится в скелетных

В биохимии тропонин используется как диагностический маркер заболеваний миокарда. Два вида тропонинов
В биохимии тропонин используется как диагностический маркер заболеваний миокарда. Два вида тропонинов

Тропомиозин — регуляторный фибриллярный белок (70 кДа). Состоит из двух перевитых α-спиралей. Тропомиозин связывается в единый
Тропомиозин — регуляторный фибриллярный белок (70 кДа). Состоит из двух перевитых α-спиралей. Тропомиозин связывается в единый

Белки саркоплазмы - протеины, растворимые в солевых средах с низкой ионной силой.
Белки саркоплазмы - протеины, растворимые в солевых средах с низкой ионной силой.

Миоглобин.
Молекула миоглобина образована единичной полипептидной цепью (153 АК) и железосодержащим гемом,
Миоглобин. Молекула миоглобина образована единичной полипептидной цепью (153 АК) и железосодержащим гемом,

Белки стромы в поперечно-полосатой мускулатуре представлены в основном коллагеном и эластином.
Белки стромы в поперечно-полосатой мускулатуре представлены в основном коллагеном и эластином.

Небелковые вещества мышечной ткани.
Небелковые вещества мышечной ткани.
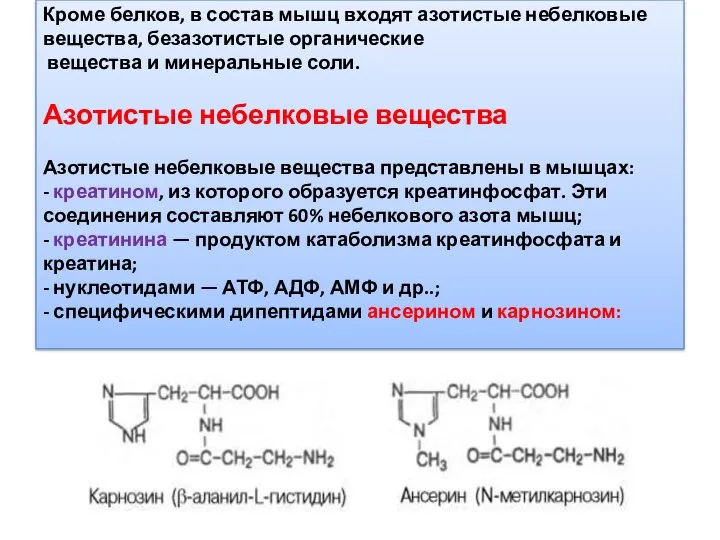
Кроме белков, в состав мышц входят азотистые небелковые вещества, безазотистые органические
Кроме белков, в состав мышц входят азотистые небелковые вещества, безазотистые органические

Карнозин и ансерин увеличивают амплитуду мышечного сокращения, которая предварительно была снижена
Карнозин и ансерин увеличивают амплитуду мышечного сокращения, которая предварительно была снижена

Безазотистые органические вещества .
представлены углеводами и липидами.
Гликоген — до 2%,
Свободная глюкоза
Безазотистые органические вещества . представлены углеводами и липидами. Гликоген — до 2%, Свободная глюкоза

Минеральные соли .
0,1 - 1,5% мышечной массы , представлены различными
Минеральные соли . 0,1 - 1,5% мышечной массы , представлены различными
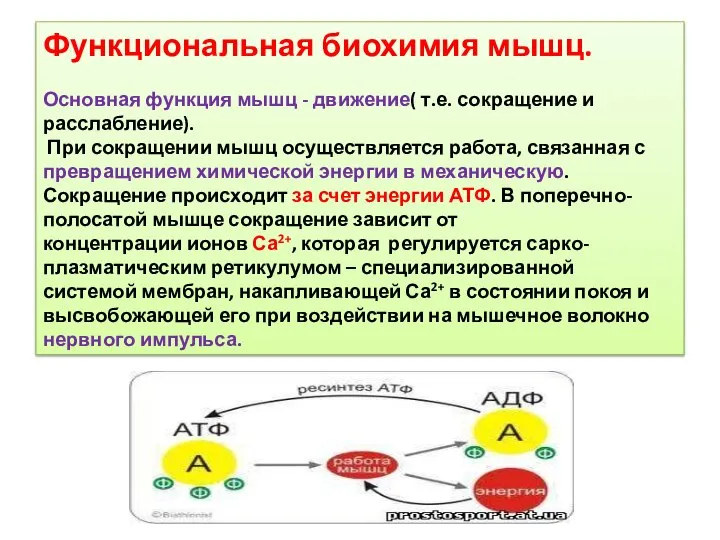
Функциональная биохимия мышц.
Основная функция мышц - движение( т.е. сокращение и расслабление).
Функциональная биохимия мышц. Основная функция мышц - движение( т.е. сокращение и расслабление).

Источники энергии для мышечных сокращений.
Креатинфосфат и АТФ –высокоэнергетические соединения мышц.
Креатинфосфат
Источники энергии для мышечных сокращений. Креатинфосфат и АТФ –высокоэнергетические соединения мышц. Креатинфосфат

Концентрация креатинфосфата в покоящейся мышечной ткани в 3-8 раз превышает концентрацию
Концентрация креатинфосфата в покоящейся мышечной ткани в 3-8 раз превышает концентрацию

Некоторое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции
Некоторое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции

Запасы креатинфосфата в мышце невелики, расход его постоянно возмещается синтезом АТФ. Для любой ткани есть
Запасы креатинфосфата в мышце невелики, расход его постоянно возмещается синтезом АТФ. Для любой ткани есть
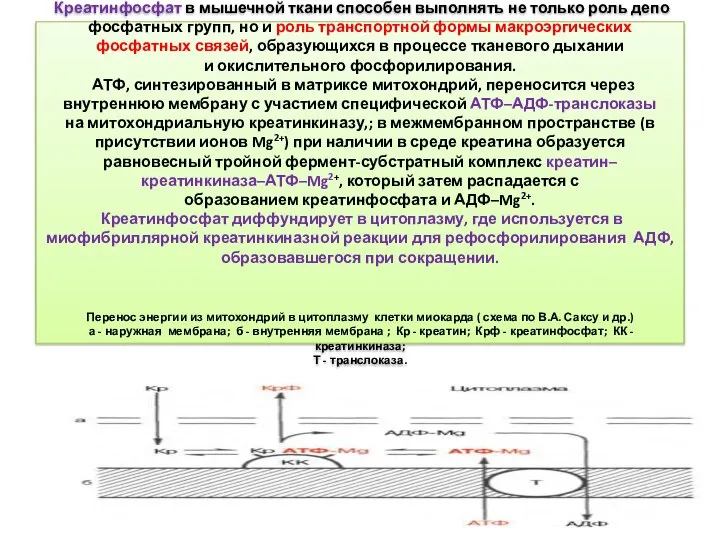
Креатинфосфат в мышечной ткани способен выполнять не только роль депо фосфатных
Креатинфосфат в мышечной ткани способен выполнять не только роль депо фосфатных

При умеренной нагрузке мышца может покрывать свои энергетические затраты за счет
При умеренной нагрузке мышца может покрывать свои энергетические затраты за счет

Механизм мышечного сокращения.
Скольжение миозина относительно актина.
Головки миозина расщепляют АТФ и за
Механизм мышечного сокращения. Скольжение миозина относительно актина. Головки миозина расщепляют АТФ и за
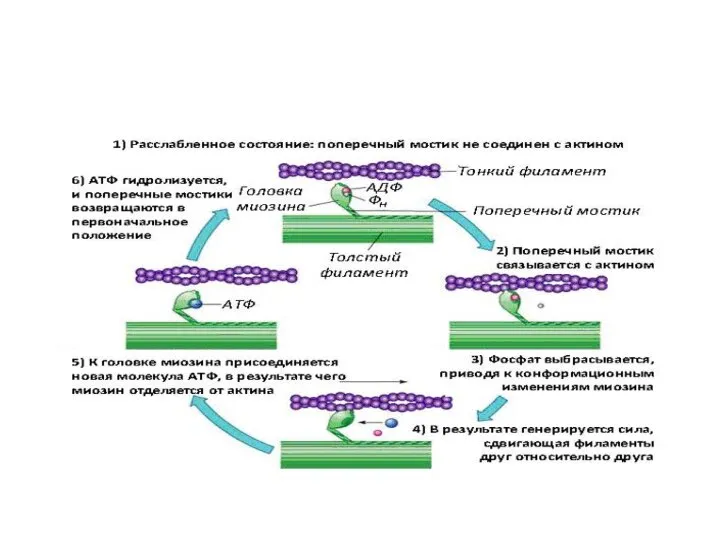

- Источником энергии для мышечного сокращения являются молекулы АТФ.
- Гидролиз
- Источником энергии для мышечного сокращения являются молекулы АТФ. - Гидролиз

В продвижении актиновых нитей вдоль миозиновых важную роль играют временно замыкающиеся
В продвижении актиновых нитей вдоль миозиновых важную роль играют временно замыкающиеся

Биохимические изменения в мышцах при патологии
Общими для большинства заболеваний мышц (мышечные
Биохимические изменения в мышцах при патологии Общими для большинства заболеваний мышц (мышечные
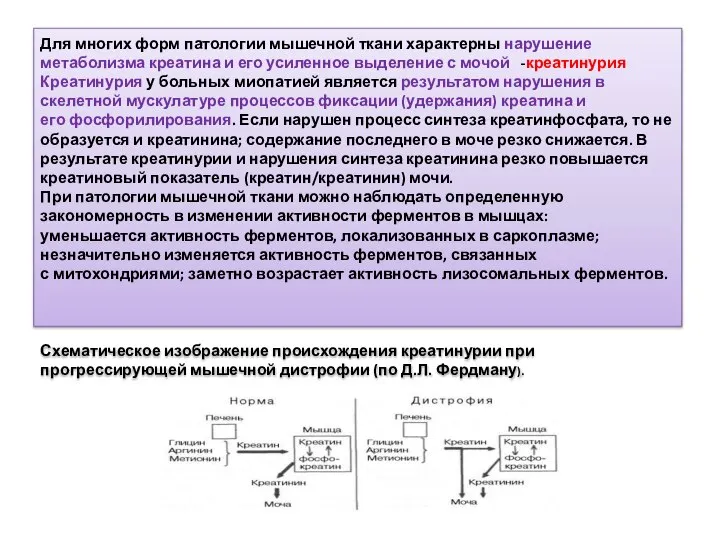
Для многих форм патологии мышечной ткани характерны нарушение метаболизма креатина и его усиленное выделение
Для многих форм патологии мышечной ткани характерны нарушение метаболизма креатина и его усиленное выделение

Миокард.
Содержание АТФ и креатинфосфата в сердечной мышце ниже, чем в скелетной мускулатуре, а расход АТФ велик. В
Миокард. Содержание АТФ и креатинфосфата в сердечной мышце ниже, чем в скелетной мускулатуре, а расход АТФ велик. В

Нарушение метаболизма сердечной мышцы при ишемической болезни
При ишемии миокарда снижено окислительное фосфорилирование и повышен
Нарушение метаболизма сердечной мышцы при ишемической болезни При ишемии миокарда снижено окислительное фосфорилирование и повышен

Размер повреждения миокарда при возникновении ишемии, можно оценить по активности ферментов
Размер повреждения миокарда при возникновении ишемии, можно оценить по активности ферментов
 Роль показателей в реализации статистических методов изучения правонарушений
Роль показателей в реализации статистических методов изучения правонарушений Разработка гастрономического тура выходного дня в Таджикистане
Разработка гастрономического тура выходного дня в Таджикистане Vorbereitung für die kontrollarbeit (2)
Vorbereitung für die kontrollarbeit (2) Экономические показатели предприятия
Экономические показатели предприятия  Артикуляционный отдел
Артикуляционный отдел  Современная деревянная архитектура и традиции
Современная деревянная архитектура и традиции Микрофлора организма человека
Микрофлора организма человека «Практика применения вывозных таможенных пошлин (на примере нефти сырой и нефтепродуктов» Выполнили студенты 3 курса Гр.
«Практика применения вывозных таможенных пошлин (на примере нефти сырой и нефтепродуктов» Выполнили студенты 3 курса Гр.  Культура Месопотамии
Культура Месопотамии Место и роль динамической метеорологии
Место и роль динамической метеорологии  Трансформатор для дуговой электросварки
Трансформатор для дуговой электросварки  Спеціалізований туризм. Екстремальний туризм
Спеціалізований туризм. Екстремальний туризм ПОДГОТОВКА К ПЕДАГОГИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ПРЕПОДАВАТЕЛЕЙ В ММА им. И.М. Сеченова
ПОДГОТОВКА К ПЕДАГОГИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ПРЕПОДАВАТЕЛЕЙ В ММА им. И.М. Сеченова  Школьный кошелек. Оплата питания в школе без проблем
Школьный кошелек. Оплата питания в школе без проблем История футбола
История футбола Презентация Соление рыбы. Мороженная рыба.
Презентация Соление рыбы. Мороженная рыба. выполнил: студентка 13 гр. филологич.фак-та Савинова Татьяна проверил: Шереметьева О. А.
выполнил: студентка 13 гр. филологич.фак-та Савинова Татьяна проверил: Шереметьева О. А. Всемирное наследие в России
Всемирное наследие в России Файлы
Файлы Наноэлектроника. Физические основы. Методы формирования наноразмерных структур. Перенос носителей заряда
Наноэлектроника. Физические основы. Методы формирования наноразмерных структур. Перенос носителей заряда Пословицы и поговорки как выражение народной мудрости
Пословицы и поговорки как выражение народной мудрости НАДЕЛЕНИЕ ОРГАНОВ МЕСТНОГО САМОУПРАВЛЕНИЯ ОТДЕЛЬНЫМИ ГОСУДАРСТВЕННЫМИ ПОЛНОМОЧИЯМИ
НАДЕЛЕНИЕ ОРГАНОВ МЕСТНОГО САМОУПРАВЛЕНИЯ ОТДЕЛЬНЫМИ ГОСУДАРСТВЕННЫМИ ПОЛНОМОЧИЯМИ Мышечная ткань
Мышечная ткань Тренер в Зеленограде. Самбо. Дзюдо. Джиу-джитсу. ММА. Бокс. Художественная гимнастика. Самооборона. Йога
Тренер в Зеленограде. Самбо. Дзюдо. Джиу-джитсу. ММА. Бокс. Художественная гимнастика. Самооборона. Йога Любовь
Любовь Об особенностях финансовой поддержки экспорта услуг Подготовил Ивантей М. группа Т-093
Об особенностях финансовой поддержки экспорта услуг Подготовил Ивантей М. группа Т-093  Презентация "Оборудование предприятий" - скачать презентации по Экономике
Презентация "Оборудование предприятий" - скачать презентации по Экономике Жизнь средневекового человека
Жизнь средневекового человека