- Ферменты. ИСТОРИЯ

Содержание
- 2. Ферменты. История. Манассеина в споре с Бюхнером c доказывает, что ферменты могут работать как в клетке,
- 3. Ферменты Практически все реакции в клетке идут при участии ферментов. Ферменты –катализаторы белковой природы. Ферменты –простые
- 4. Сходство и различие с неорганическими катализаторами Ферменты –как и неорганические катализаторы : выходят из реакции в
- 5. Субстратная специфичность ферментов Абсолютная (фермент катализирует реакцию только с одним субстратом):аргиназа, уреаза, ДНК-полимераза, аденилатциклаза Относительная или
- 7. Ферменты Конститутивные (синтезируются постоянно): ферменты гликолиза, окисления жирных кислот ит.д. Адаптивные, индуцибельные (синтезируются в определенных ситуациях,
- 8. Организация и структурированность ферментов Ферменты экскретируемые (работают во внешней среде, вне клеток): гидролазы пищеварительного тракта Ферменты
- 9. Ферменты клеточного метаболизма Растворимые (в цитозоле, матриксе лизосом, митохондрий): фосфофруктокиназа, кислая фосфатаза, малатдегидрогеназа) «Структурированные», ассоциированные с
- 10. СТРОЕНИЕ ФЕРМЕНТОВ Активный центр (субстратный + каталитический участки) Аллостерические ферменты имеют еще регуляторный центр Активный центр
- 11. СТРОЕНИЕ ФЕРМЕНТОВ Каталитический центр «отвечает» за механизм катализа, большие группы ферментов могут иметь одинаковое строение каталитического
- 13. КАТАЛИТИЧЕСКИЙ УЧАСТОК ФЕРМЕНТОВ Каталитический центр простых ферментов состоит из реакционоспособных аминокислот: серин, треонин (ОН), аргинин (гуанидин),
- 14. Как работают ферменты? Снижают энергию активации молекул субстрата. Скорость реакции пропорциональна количеству молекул, свободная энергия которых
- 17. Механизмы катализа (гипотезы) Э.Фишер, 1890г. – гипотеза «шаблона», (ключ-замок). Объясняет только абсолютную специфичность ферментов. Кошланд –
- 20. Химизм катализа Кислотно-основной: молекулы ферментов в активном центре содержат функциональные группы, служащие донорами или акцепторами протонов
- 21. КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА L.Michaelis, M.Menten,1913г. k1 k3 S + E ES E + P k2 k4
- 22. КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА Л. Михаэлис и М. Ментен решают уравнение относительно [ES], когда реакция находится в
- 24. КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА [E] [S] [ES] = [S] + (k3 + k2) / k1 k3 +
- 26. Классификация ферментов JUBMB ввел международную классификацию ферментов . Шифр каждого фермента состоит из 4 цифр: 1.
- 27. Классификация ферментов. 1.ОКСИДОРЕДУКТАЗЫ 1.1 Дегидрогеназы SH2 + S* = S + S*H2 (ЛДГ,МДГ,АДГ) 1.2 Оксидазы SH2
- 28. Классификация ферментов 2.ТРАНСФЕРАЗЫ 2.1. Аминотрансферазы 2.2. Фосфотрансферазы 2.3. Метилтрансферазы 2.4. Глюкуронилтрансферазы 2.5. Сульфотрансферазы 2.7. РНК- и
- 29. Классификация ферментов 3. ГИДРОЛАЗЫ 3.1.Эстеразы 3.2 Гликозидазы 3.3. Пептидазы 3.4. Тиолазы 3.5. Рибонуклеазы 3.6. Амидазы 3.7.
- 30. Классификация ферментов 4. ЛИАЗЫ Отщепление каких-либо групп негидролитическим и неокислительным путем или присоединение групп (чаще всего
- 31. Классификация ферментов 5. ИЗОМЕРАЗЫ Внутримолекулярные превращения: внутримолекулярный перенос групп; перенос связей. Изомеразы, Мутазы, Рацемазы.
- 32. Классификация ферментов 6. ЛИГАЗЫ (СИНТЕТАЗЫ) Соединение 2-х молекул с использованием знергии гидролиза макроэргической связи АТФ (синтетаза,
- 33. Определение активности ферментов Содержание (количество) фермента определить невозможно. Определяют активность фермента = скорость реакции. Для этого
- 34. Единицы ферментативной активности Общая активность : количество молей субстрата (или продукта) за единицу времени.(моль/час, ммоль/мин, мкмоль/сек).
- 35. Единицы ферментативной активности 1МЕ соответствует количеству фермента, катализирующего превращение 1 мкмоля субстрата за 1 мин 1
- 36. Регуляция активности ферментов Скорость ферментативных реакций (активность ферментов) зависит от: [ E ], [S ], [
- 37. Регуляция активности ферментов Количество фермента находится под генетическим контролем, определяется скоростью синтеза и распада ферментных молекул.
- 38. Регуляция активности ферментов Зависимость от рН среды –специфическое свойство биокатализаторов. Каждый фермент имеет opt значение рН
- 39. Регуляция активности ферментов Зависимость активности ферментов от t0 среды подчиняется законам термодинамики. Для ферментов коэффициент Q10
- 40. Регуляция активности ферментов Зависимость скорости реакции от [S] различна для «Михаэлисовских» ферментов и аллостерических. Для всех
- 41. Регуляция активности ферментов Зависимость скорости реакции от [P]: На начальных этапах реакции, когда [P] мала, равновесие
- 42. Регуляция активности ферментов Профермент – активный фермент: Лимитированный протеолиз Ковалентная модификация (фосфорилирование, ацетилирование, аденилирование и т.д.)
- 43. Регуляция активности ферментов Лимитированный (ограниченный) протеолиз: Профермент синтезируется в виде крупной молекулы –предшественника Подвергается гидролизу специфическими
- 45. Регуляция активности ферментов Ковалентная модификация: Е – ОН ? Е – О-РО3 (протеинфосфотранфераза фосфорилирует ферментный белок
- 47. Регуляция активности ферментов Белок –белковые взаимодействия: Ассоциация – диссоциация белковых субъединиц: G – белковый комплекс, фосфопротеинкиназа
- 48. Регуляция активности ферментов Действие активаторов и ингибиторов: Ингибиторы по прочности связывания с ферментом м.б. необратимыми (комплекс
- 50. Регуляция активности ферментов Ингибиторы по механизму действия: Конкурентные (ингибитор- структурный аналог субстрата, связывается в активном центре
- 52. Конкурентное и неконкурентное ингибирование активности ферментов Кинетика конкурентного и неконкурентного ингибирования различна: Конкурентный ингибитор снижает сродство
- 54. Конкурентное и неконкурентное ингибирование активности ферментов Неконкурентный ингибитор снижает скорость реакции до величины Vmax / (1
- 57. Изоферменты –молекулярные формы ферментов Разные молекулярные формы одного фермента (катализируют одну и ту же реакцию) в
- 58. Компартментализация Важный момент регуляции скорости метаболических путей в эукариотической клетке. Участие мембран заключается и в интеграции
- 59. Аллостерические ферменты «ИНЫЕ»! Не подчиняются кинетике Михаэлиса – Ментен. Кроме каталитического центра имеют еще один, другой
- 60. Аллостерические ферменты Эффекторами чаще всего служат метаболиты данного ферментативного процесса. Неспецифическими регуляторами часто служат нуклеотиды: NAD,
- 62. Аллостерические ферменты Как правило, аллостерические ферменты катализируют ключевые (лимитирующие скорость) реакции метаболических путей. Аллостерические ферменты катализируют
- 64. Кофакторы и коферменты Холофермент = кофермент (кофактор) + + апофермент. Коферменты (кофакторы) – низкомолекулярные, термостабильные простетические
- 65. Кофакторы) (неорганической природы) Ионы металлов: Стабилизируют субстрат (Е – S – Me), Mg2+- АТФ. Стабилизируют активный
- 72. Коферменты Производные витаминов (фосфорилированные: пиридоксальфосфат, тиаминпирофосфат или аденилированные: НАД, ФМН, ФАД) Гем (ы) Нуклеотиды (АТР, ГТР)
- 73. ВИТАМИНЫ Низкомолекулярные органические вещества различного строения, которые животные должны получать извне полностью или частично (микронутриенты). Не
- 74. Из истории витаминологии Научные основы витаминологии – начало 19 века: Н.И. Лунин, Ф. Мажанди, К. Танаки
- 75. История витаминологии А.Виндаус, 1928 г. – открытие витамина Д и холестерола. У.Хеворс и П. Каррер,1937 г.
- 76. Номенклатура витаминов Буквенные символы (А, В,С…) Химические названия (тиамин,рибофлавин, никотинамид….) Биологическая классификация (антирахитический, антигеморрагический, антиневритный…) Жиро
- 77. Биохимические функции витаминов 1. Обеспечивают активность ферментов (кофакторная ф –ция): В1,В2.,В6 ,РР, Н, К, фолиевая кислота,
- 83. Метаболизм витаминов Обмен очень интенсивный. Не накапливаются! (Исключение – жирорастворимые витамины, частично депонируются в печени и
- 84. Источники витаминов: Пищевые: желток яиц, печень, икра, проростки злаков, дрожжи. Микрофлора кишечника. КАК СОХРАНИТЬ ВИТАМИНЫ В
- 85. Причины гиповитаминозов: Первичный (экзогенный) гипо- или авитаминоз: алиментарная недостаточность. Голодание, «однобокая» диета, потребление «рафинированных» продуктов. Вторичные
- 86. Причины гиповитаминозов: Нарушение всасывания в ЖКТ жирорастворимых витаминов (снижение желчеобразования); В12 (недостаток фактора Касла). Нарушение микрофлоры
- 87. Взаимодействие витаминов Витамин влияет на метаболизм другого витамина (Токоферол препятствует переокислению витамина А; полиеновые кислоты (витамин
- 88. ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. А (ретинол, ретиналь), антиксерофтальмический. Источник: продукты животного происхождения (печень рыб);растительные продукты содержат каротин –предшественник
- 89. ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. Д (кальциферол), антирахитический. Источники: эндогенный синтез (УФ), продукты животного происхождения, дрожжи содержат эргостерин. Увеличивает
- 90. ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. Е (токоферол), антистерильный Источники: растительные масла. Обеспечивает стабильность клеточных мембран, являясь антиоксидантом и снижая
- 91. ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. К (филлохиноны), антигеморрагический Источники: зелень, печень, дрожжи, синтез микрофлорой. Является кофактором карбоксилазы глутамата ,
- 92. Водорастворимые витамины. В1 (тиамин), антиневритный Кофакторная форма – ТДФ (тиаминдифосфат) Работает в составе дегидрогеназных комплексов, обеспечивая
- 94. Водорастворимые витамины. В2 (рибофлавин), витамин роста Образует кофакторы ФМН, ФАД. Работает в составе анаэробных дегидрогеназ различных
- 97. Водорастворимые витамины. В3 (пантотеновая кислота), универсальный витамин. Входит в состав КоА, активатора и переносчика ацетильных и
- 98. Водорастворимые витамины. В5, РР (никотинамид), антипеллагрический. Образует кофакторы НАД, НАДФ. Входит в состав анаэробных дегидрогеназ, обратимо
- 101. Водорастворимые витамины. В6(пиридоксаль), антидерматитный Образует кофактор пиридоксальфосфат. Входит в состав трансфераз и декарбоксилаз аминокислот, моно- и
- 102. Водорастворимые витамины. Н (биотин), антисеборейный.
- 104. Скачать презентацию

Ферменты. История.
Манассеина в споре с Бюхнером c доказывает, что ферменты могут
Ферменты. История.
Манассеина в споре с Бюхнером c доказывает, что ферменты могут

Ферменты
Практически все реакции в клетке идут при участии ферментов.
Ферменты –катализаторы белковой
Ферменты
Практически все реакции в клетке идут при участии ферментов.
Ферменты –катализаторы белковой

Сходство и различие с неорганическими катализаторами
Ферменты –как и неорганические катализаторы :
Сходство и различие с неорганическими катализаторами
Ферменты –как и неорганические катализаторы :

Субстратная специфичность ферментов
Абсолютная (фермент катализирует реакцию только с одним субстратом):аргиназа, уреаза,
Субстратная специфичность ферментов
Абсолютная (фермент катализирует реакцию только с одним субстратом):аргиназа, уреаза,
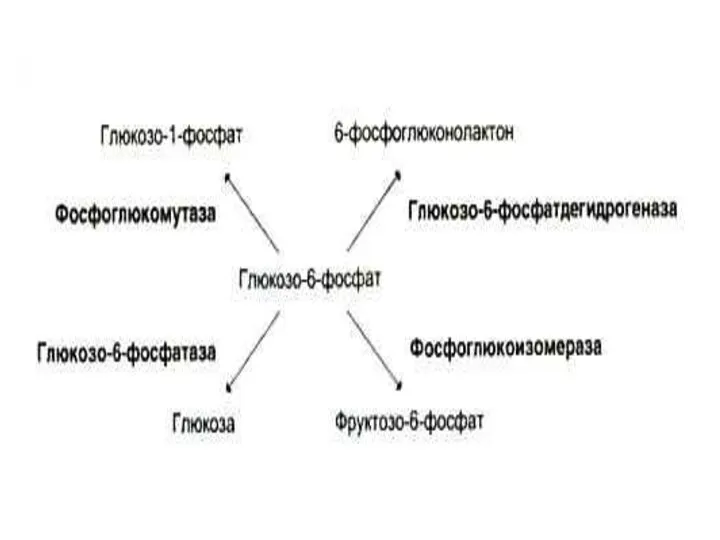

Ферменты
Конститутивные (синтезируются постоянно): ферменты гликолиза, окисления жирных кислот ит.д.
Адаптивные, индуцибельные (синтезируются
Ферменты
Конститутивные (синтезируются постоянно): ферменты гликолиза, окисления жирных кислот ит.д.
Адаптивные, индуцибельные (синтезируются

Организация и структурированность ферментов
Ферменты экскретируемые (работают во внешней среде, вне клеток):
Организация и структурированность ферментов
Ферменты экскретируемые (работают во внешней среде, вне клеток):

Ферменты клеточного метаболизма
Растворимые (в цитозоле, матриксе лизосом, митохондрий): фосфофруктокиназа, кислая фосфатаза,
Ферменты клеточного метаболизма
Растворимые (в цитозоле, матриксе лизосом, митохондрий): фосфофруктокиназа, кислая фосфатаза,

СТРОЕНИЕ ФЕРМЕНТОВ
Активный центр (субстратный + каталитический участки)
Аллостерические ферменты имеют еще регуляторный
СТРОЕНИЕ ФЕРМЕНТОВ
Активный центр (субстратный + каталитический участки)
Аллостерические ферменты имеют еще регуляторный

СТРОЕНИЕ ФЕРМЕНТОВ
Каталитический центр «отвечает» за механизм катализа, большие группы ферментов могут
СТРОЕНИЕ ФЕРМЕНТОВ
Каталитический центр «отвечает» за механизм катализа, большие группы ферментов могут
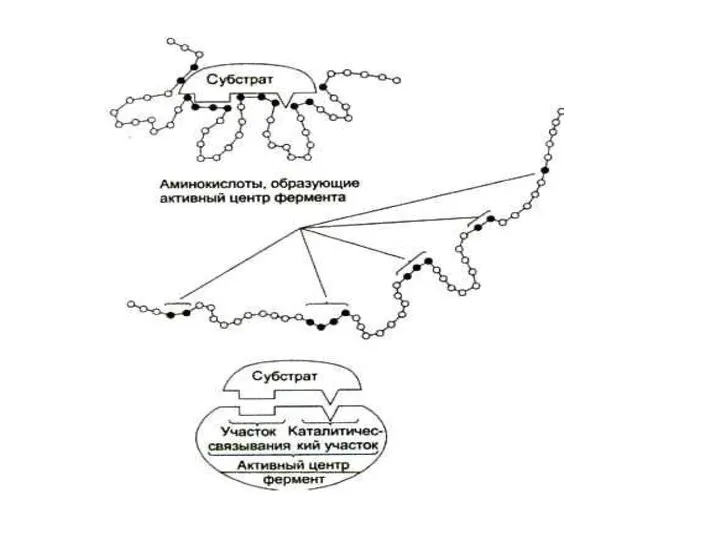

КАТАЛИТИЧЕСКИЙ УЧАСТОК ФЕРМЕНТОВ
Каталитический центр простых ферментов состоит из реакционоспособных аминокислот:
КАТАЛИТИЧЕСКИЙ УЧАСТОК ФЕРМЕНТОВ
Каталитический центр простых ферментов состоит из реакционоспособных аминокислот:

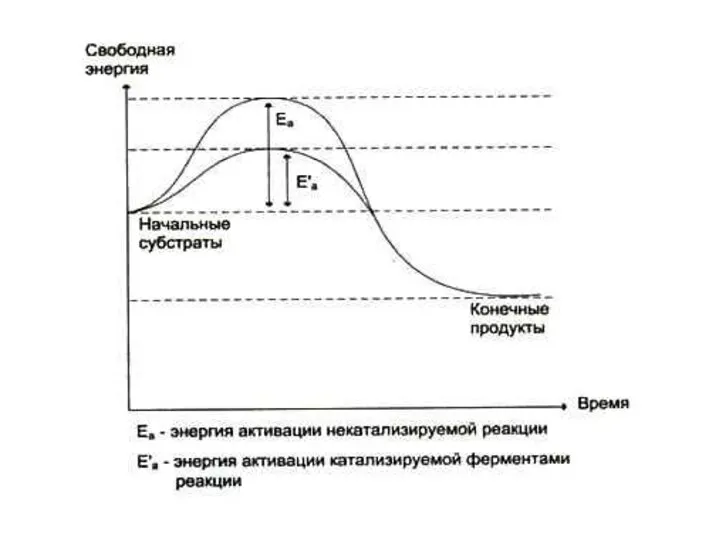
Как работают ферменты?
Снижают энергию активации молекул субстрата. Скорость реакции пропорциональна количеству
Как работают ферменты?
Снижают энергию активации молекул субстрата. Скорость реакции пропорциональна количеству
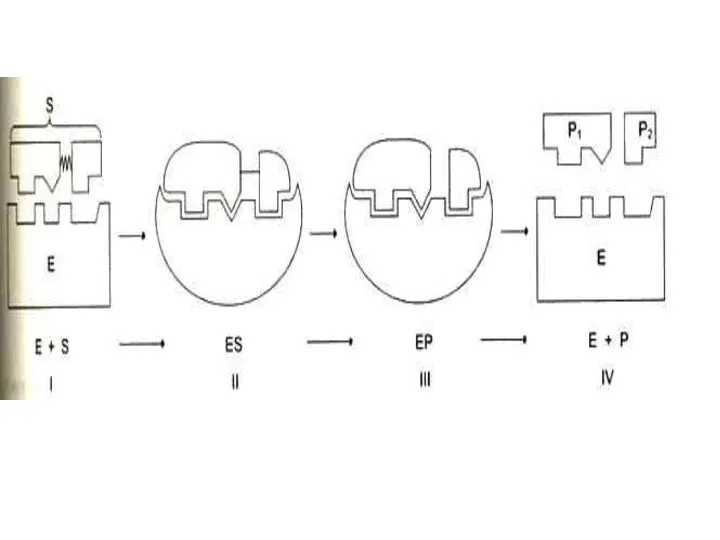

Механизмы катализа (гипотезы)
Э.Фишер, 1890г. – гипотеза «шаблона», (ключ-замок). Объясняет только абсолютную
Механизмы катализа (гипотезы)
Э.Фишер, 1890г. – гипотеза «шаблона», (ключ-замок). Объясняет только абсолютную
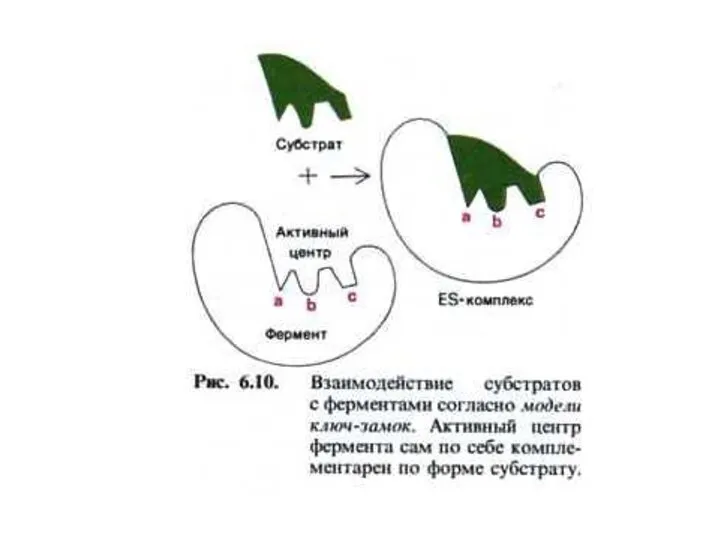
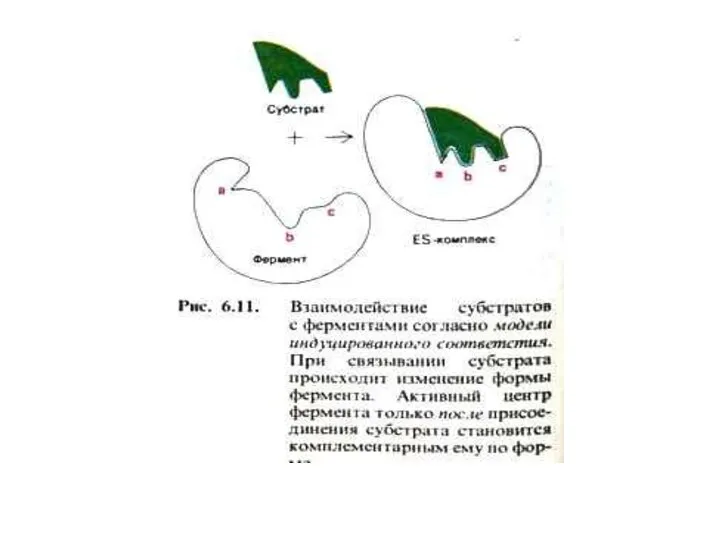

Химизм катализа
Кислотно-основной: молекулы ферментов в активном центре содержат функциональные группы, служащие
Химизм катализа
Кислотно-основной: молекулы ферментов в активном центре содержат функциональные группы, служащие

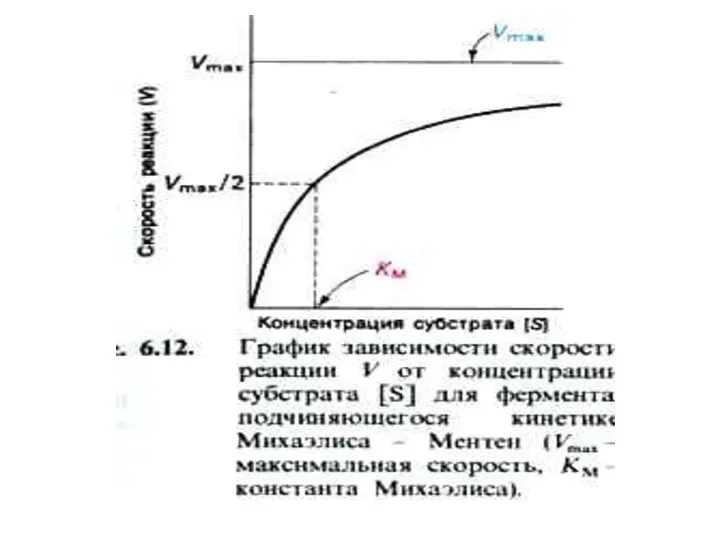
КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
L.Michaelis, M.Menten,1913г.
k1 k3
S + E ES E
КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
L.Michaelis, M.Menten,1913г.
k1 k3
S + E ES E

КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Л. Михаэлис и М. Ментен решают уравнение относительно [ES],
КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Л. Михаэлис и М. Ментен решают уравнение относительно [ES],
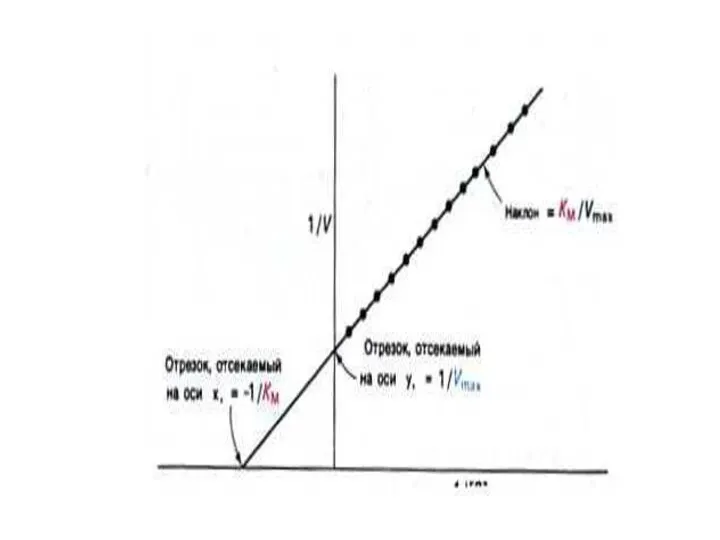
![КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА [E] [S] [ES] = [S] + (k3 +](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1302977/slide-23.jpg)
КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
[E] [S]
[ES] =
[S] + (k3 + k2)
КИНЕТИКА ФЕРМЕНТАТИВНОГО КАТАЛИЗА
[E] [S]
[ES] =
[S] + (k3 + k2)

Классификация ферментов
JUBMB ввел международную классификацию ферментов . Шифр каждого фермента состоит
Классификация ферментов
JUBMB ввел международную классификацию ферментов . Шифр каждого фермента состоит

Классификация ферментов.
1.ОКСИДОРЕДУКТАЗЫ
1.1 Дегидрогеназы
SH2 + S* = S + S*H2 (ЛДГ,МДГ,АДГ)
1.2 Оксидазы
SH2
Классификация ферментов.
1.ОКСИДОРЕДУКТАЗЫ
1.1 Дегидрогеназы
SH2 + S* = S + S*H2 (ЛДГ,МДГ,АДГ)
1.2 Оксидазы
SH2

Классификация ферментов
2.ТРАНСФЕРАЗЫ
2.1. Аминотрансферазы
2.2. Фосфотрансферазы
2.3. Метилтрансферазы
2.4. Глюкуронилтрансферазы
2.5. Сульфотрансферазы
2.7. РНК- и ДНК- полимеразы
Классификация ферментов
2.ТРАНСФЕРАЗЫ
2.1. Аминотрансферазы
2.2. Фосфотрансферазы
2.3. Метилтрансферазы
2.4. Глюкуронилтрансферазы
2.5. Сульфотрансферазы
2.7. РНК- и ДНК- полимеразы

Классификация ферментов
3. ГИДРОЛАЗЫ
3.1.Эстеразы
3.2 Гликозидазы
3.3. Пептидазы
3.4. Тиолазы
3.5. Рибонуклеазы
3.6. Амидазы
3.7. Дезаминазы
Классификация ферментов
3. ГИДРОЛАЗЫ
3.1.Эстеразы
3.2 Гликозидазы
3.3. Пептидазы
3.4. Тиолазы
3.5. Рибонуклеазы
3.6. Амидазы
3.7. Дезаминазы

Классификация ферментов
4. ЛИАЗЫ
Отщепление каких-либо групп негидролитическим и неокислительным путем или присоединение
Классификация ферментов
4. ЛИАЗЫ
Отщепление каких-либо групп негидролитическим и неокислительным путем или присоединение

Классификация ферментов
5. ИЗОМЕРАЗЫ
Внутримолекулярные превращения:
внутримолекулярный перенос групп; перенос связей.
Изомеразы,
Мутазы,
Рацемазы.
Классификация ферментов
5. ИЗОМЕРАЗЫ
Внутримолекулярные превращения:
внутримолекулярный перенос групп; перенос связей.
Изомеразы,
Мутазы,
Рацемазы.

Классификация ферментов
6. ЛИГАЗЫ (СИНТЕТАЗЫ)
Соединение 2-х молекул с использованием знергии гидролиза макроэргической
Классификация ферментов
6. ЛИГАЗЫ (СИНТЕТАЗЫ)
Соединение 2-х молекул с использованием знергии гидролиза макроэргической

Определение активности ферментов
Содержание (количество) фермента определить невозможно. Определяют активность фермента =
Определение активности ферментов
Содержание (количество) фермента определить невозможно. Определяют активность фермента =

Единицы ферментативной активности
Общая активность : количество молей субстрата (или продукта) за
Единицы ферментативной активности
Общая активность : количество молей субстрата (или продукта) за

Единицы ферментативной активности
1МЕ соответствует количеству фермента, катализирующего превращение 1 мкмоля субстрата
Единицы ферментативной активности
1МЕ соответствует количеству фермента, катализирующего превращение 1 мкмоля субстрата

Регуляция активности ферментов
Скорость ферментативных реакций (активность ферментов) зависит от:
[ E ],
Регуляция активности ферментов
Скорость ферментативных реакций (активность ферментов) зависит от:
[ E ],

Регуляция активности ферментов
Количество фермента находится под генетическим контролем, определяется скоростью синтеза
Регуляция активности ферментов
Количество фермента находится под генетическим контролем, определяется скоростью синтеза

Регуляция активности ферментов
Зависимость от рН среды –специфическое свойство биокатализаторов. Каждый фермент
Регуляция активности ферментов
Зависимость от рН среды –специфическое свойство биокатализаторов. Каждый фермент

Регуляция активности ферментов
Зависимость активности ферментов от t0 среды подчиняется законам термодинамики.
Регуляция активности ферментов
Зависимость активности ферментов от t0 среды подчиняется законам термодинамики.
![Регуляция активности ферментов Зависимость скорости реакции от [S] различна для «Михаэлисовских»](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1302977/slide-39.jpg)
Регуляция активности ферментов
Зависимость скорости реакции от [S] различна для «Михаэлисовских» ферментов
Регуляция активности ферментов
Зависимость скорости реакции от [S] различна для «Михаэлисовских» ферментов
![Регуляция активности ферментов Зависимость скорости реакции от [P]: На начальных этапах](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1302977/slide-40.jpg)
Регуляция активности ферментов
Зависимость скорости реакции от [P]:
На начальных этапах реакции, когда
Регуляция активности ферментов
Зависимость скорости реакции от [P]:
На начальных этапах реакции, когда

Регуляция активности ферментов
Профермент – активный фермент:
Лимитированный протеолиз
Ковалентная модификация (фосфорилирование, ацетилирование, аденилирование
Регуляция активности ферментов
Профермент – активный фермент:
Лимитированный протеолиз
Ковалентная модификация (фосфорилирование, ацетилирование, аденилирование

Регуляция активности ферментов
Лимитированный (ограниченный) протеолиз:
Профермент синтезируется в виде крупной молекулы –предшественника
Подвергается
Регуляция активности ферментов
Лимитированный (ограниченный) протеолиз:
Профермент синтезируется в виде крупной молекулы –предшественника
Подвергается
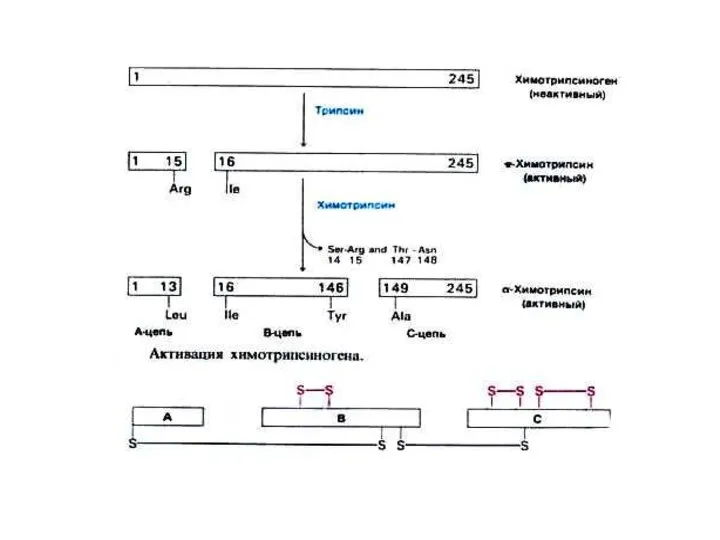

Регуляция активности ферментов
Ковалентная модификация:
Е – ОН ? Е – О-РО3 (протеинфосфотранфераза
Регуляция активности ферментов
Ковалентная модификация:
Е – ОН ? Е – О-РО3 (протеинфосфотранфераза
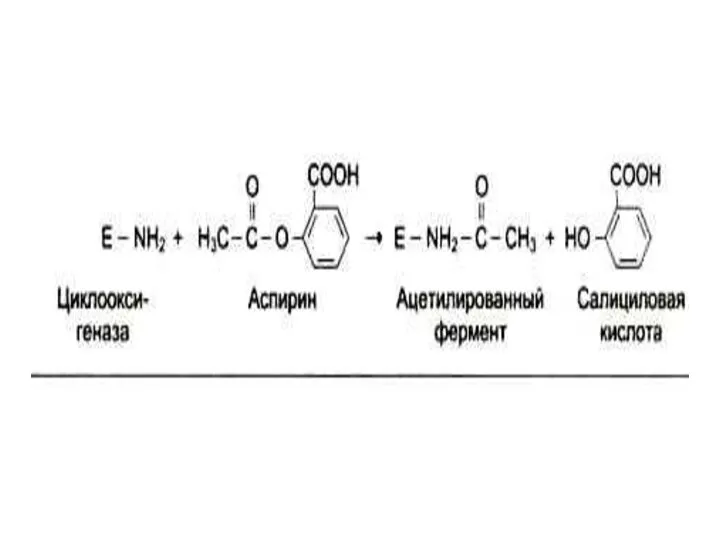

Регуляция активности ферментов
Белок –белковые взаимодействия:
Ассоциация – диссоциация белковых субъединиц:
G –
Регуляция активности ферментов
Белок –белковые взаимодействия:
Ассоциация – диссоциация белковых субъединиц:
G –

Регуляция активности ферментов
Действие активаторов и ингибиторов:
Ингибиторы по прочности связывания с ферментом
Регуляция активности ферментов
Действие активаторов и ингибиторов:
Ингибиторы по прочности связывания с ферментом


Регуляция активности ферментов
Ингибиторы по механизму действия:
Конкурентные (ингибитор- структурный аналог субстрата,
Регуляция активности ферментов
Ингибиторы по механизму действия:
Конкурентные (ингибитор- структурный аналог субстрата,
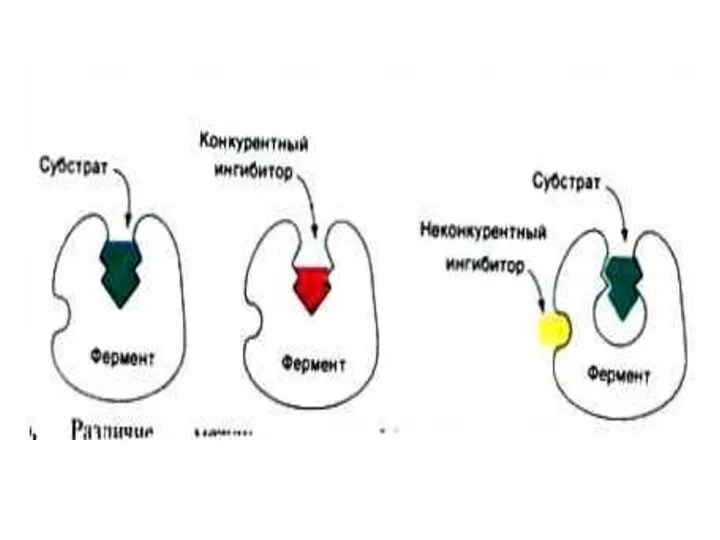

Конкурентное и неконкурентное ингибирование активности ферментов
Кинетика конкурентного и неконкурентного ингибирования различна:
Конкурентный
Конкурентное и неконкурентное ингибирование активности ферментов
Кинетика конкурентного и неконкурентного ингибирования различна:
Конкурентный
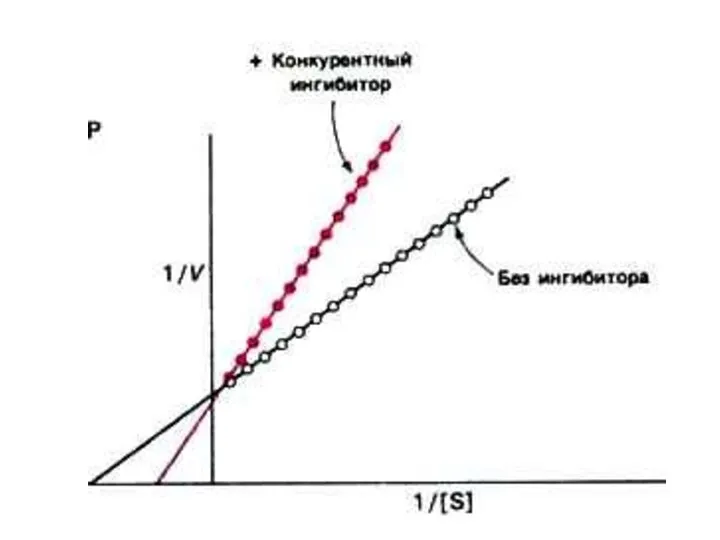

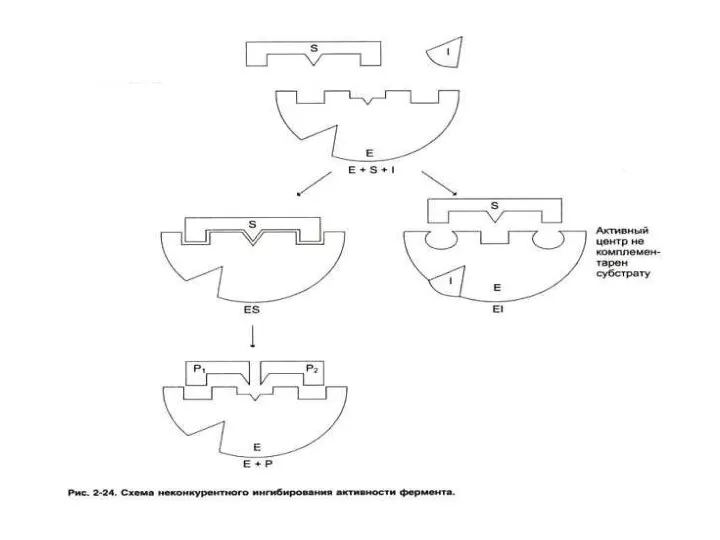
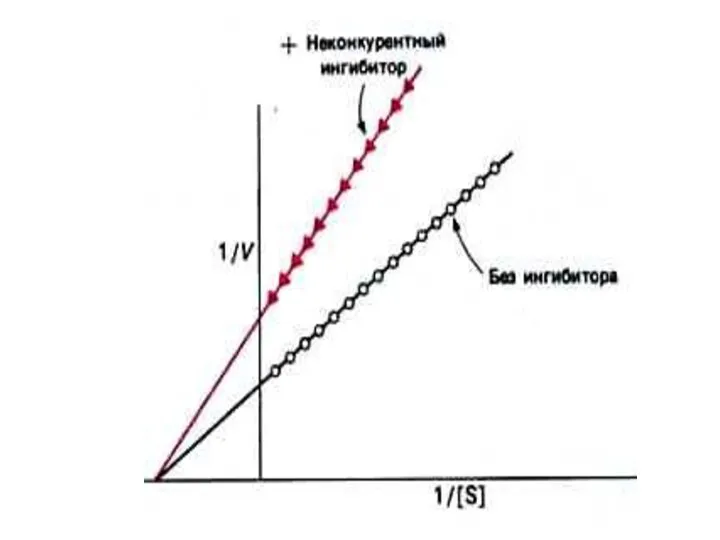
Конкурентное и неконкурентное ингибирование активности ферментов
Неконкурентный ингибитор снижает скорость реакции до
Конкурентное и неконкурентное ингибирование активности ферментов
Неконкурентный ингибитор снижает скорость реакции до

Изоферменты –молекулярные формы ферментов
Разные молекулярные формы одного фермента (катализируют одну и
Изоферменты –молекулярные формы ферментов
Разные молекулярные формы одного фермента (катализируют одну и

Компартментализация
Важный момент регуляции скорости метаболических путей в эукариотической клетке.
Участие мембран заключается
Компартментализация
Важный момент регуляции скорости метаболических путей в эукариотической клетке.
Участие мембран заключается

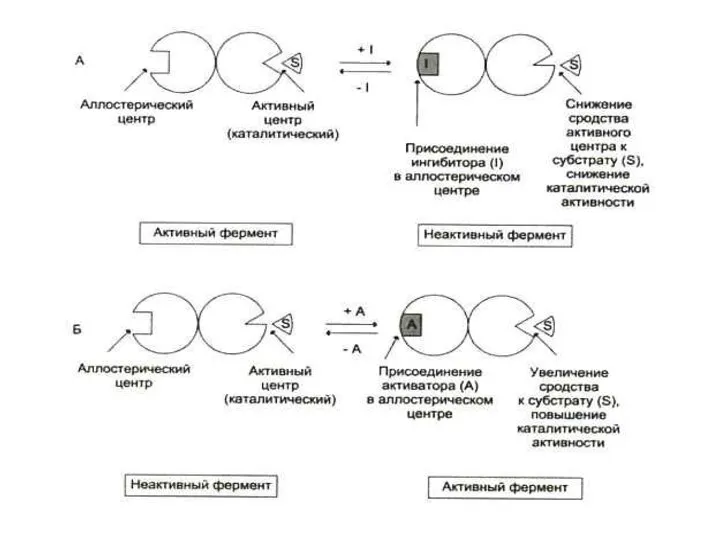
Аллостерические ферменты
«ИНЫЕ»! Не подчиняются кинетике Михаэлиса – Ментен.
Кроме каталитического центра имеют
Аллостерические ферменты
«ИНЫЕ»! Не подчиняются кинетике Михаэлиса – Ментен.
Кроме каталитического центра имеют

Аллостерические ферменты
Эффекторами чаще всего служат метаболиты данного ферментативного процесса. Неспецифическими регуляторами
Аллостерические ферменты
Эффекторами чаще всего служат метаболиты данного ферментативного процесса. Неспецифическими регуляторами

Аллостерические ферменты
Как правило, аллостерические ферменты катализируют ключевые (лимитирующие скорость) реакции метаболических
Аллостерические ферменты
Как правило, аллостерические ферменты катализируют ключевые (лимитирующие скорость) реакции метаболических

Кофакторы и коферменты
Холофермент = кофермент (кофактор) +
+ апофермент.
Коферменты (кофакторы) –
Кофакторы и коферменты
Холофермент = кофермент (кофактор) +
+ апофермент.
Коферменты (кофакторы) –

Кофакторы) (неорганической природы)
Ионы металлов:
Стабилизируют субстрат (Е – S – Me), Mg2+-
Кофакторы) (неорганической природы)
Ионы металлов:
Стабилизируют субстрат (Е – S – Me), Mg2+-
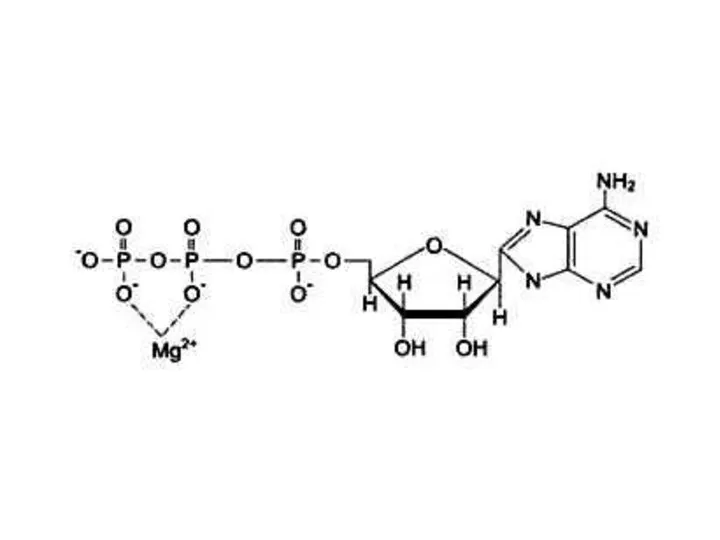
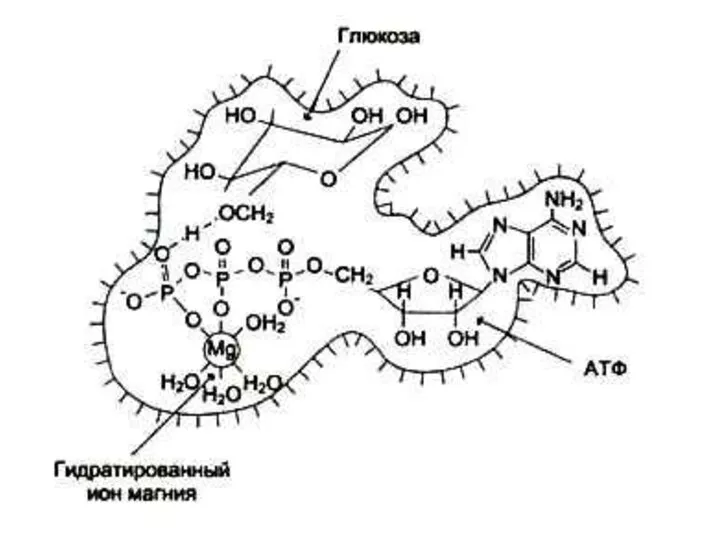
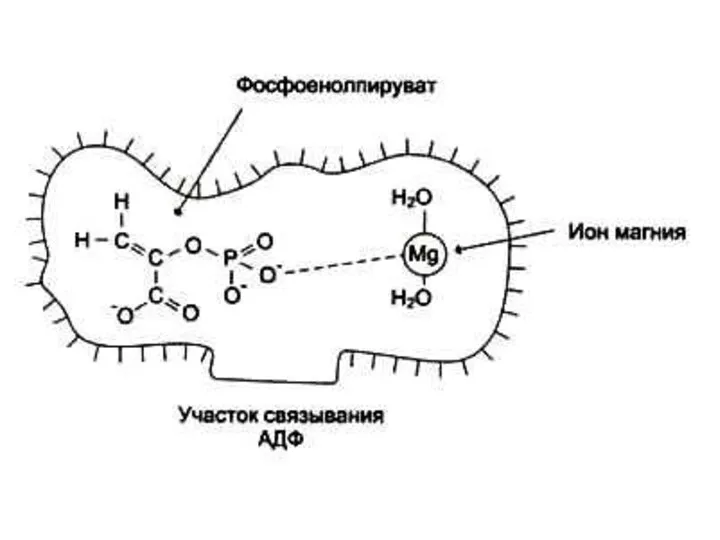
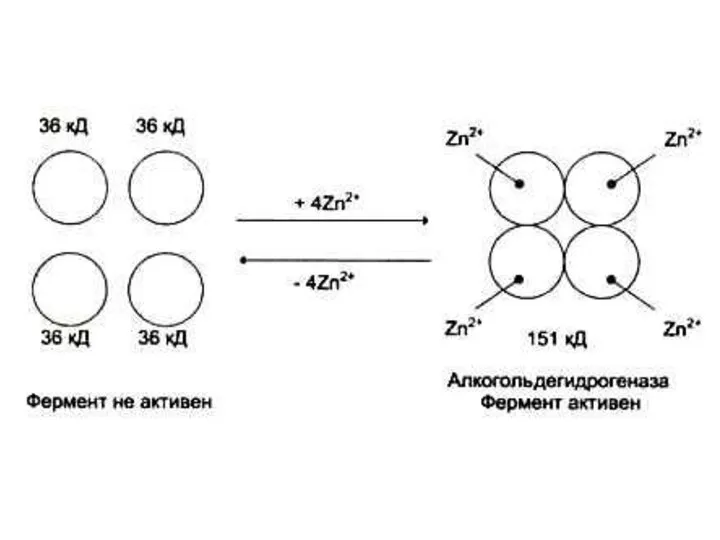
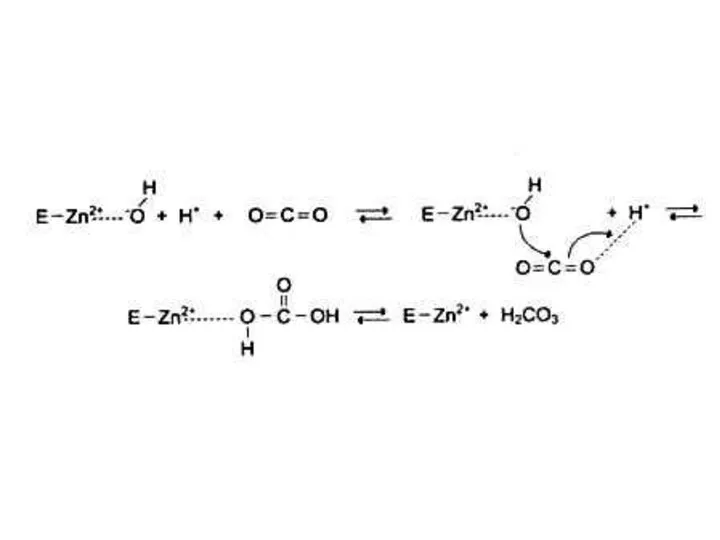
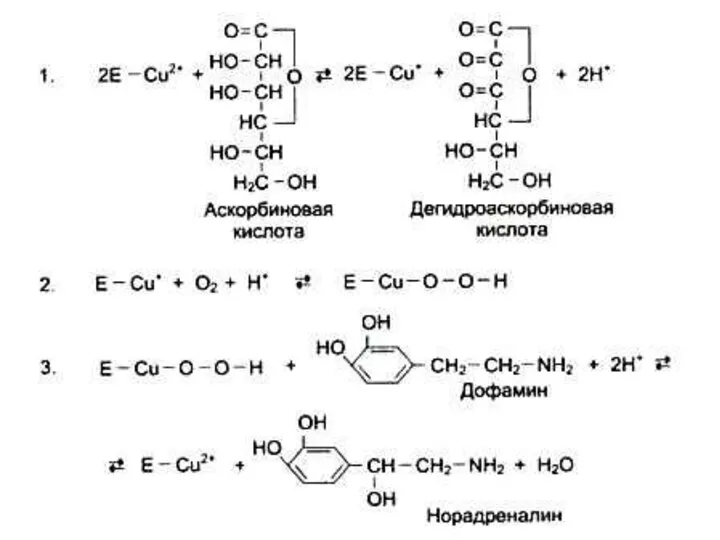

Коферменты
Производные витаминов (фосфорилированные: пиридоксальфосфат, тиаминпирофосфат или аденилированные: НАД, ФМН, ФАД)
Гем
Коферменты
Производные витаминов (фосфорилированные: пиридоксальфосфат, тиаминпирофосфат или аденилированные: НАД, ФМН, ФАД)
Гем

ВИТАМИНЫ
Низкомолекулярные органические вещества различного строения, которые животные должны получать извне полностью
ВИТАМИНЫ
Низкомолекулярные органические вещества различного строения, которые животные должны получать извне полностью

Из истории витаминологии
Научные основы витаминологии – начало 19 века:
Н.И. Лунин, Ф.
Из истории витаминологии
Научные основы витаминологии – начало 19 века:
Н.И. Лунин, Ф.

История витаминологии
А.Виндаус, 1928 г. – открытие витамина Д и холестерола.
У.Хеворс и
История витаминологии
А.Виндаус, 1928 г. – открытие витамина Д и холестерола.
У.Хеворс и

Номенклатура витаминов
Буквенные символы (А, В,С…)
Химические названия (тиамин,рибофлавин, никотинамид….)
Биологическая классификация (антирахитический,
Номенклатура витаминов
Буквенные символы (А, В,С…)
Химические названия (тиамин,рибофлавин, никотинамид….)
Биологическая классификация (антирахитический,

Биохимические функции витаминов
1. Обеспечивают активность ферментов (кофакторная ф –ция): В1,В2.,В6 ,РР,
Биохимические функции витаминов
1. Обеспечивают активность ферментов (кофакторная ф –ция): В1,В2.,В6 ,РР,
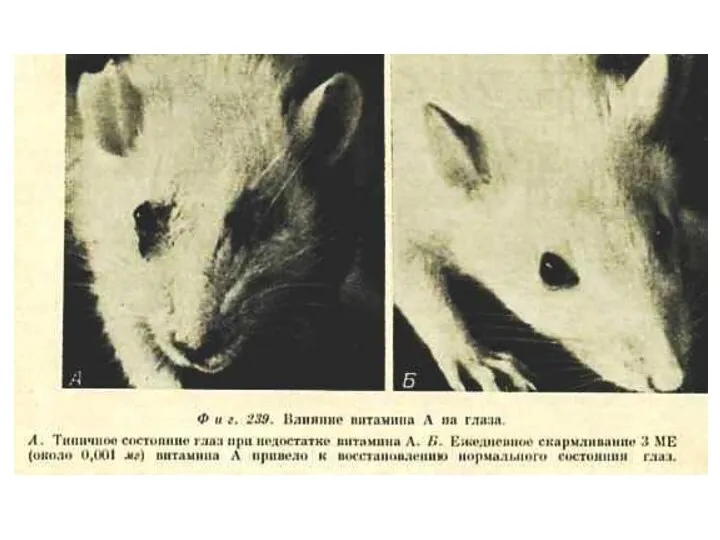
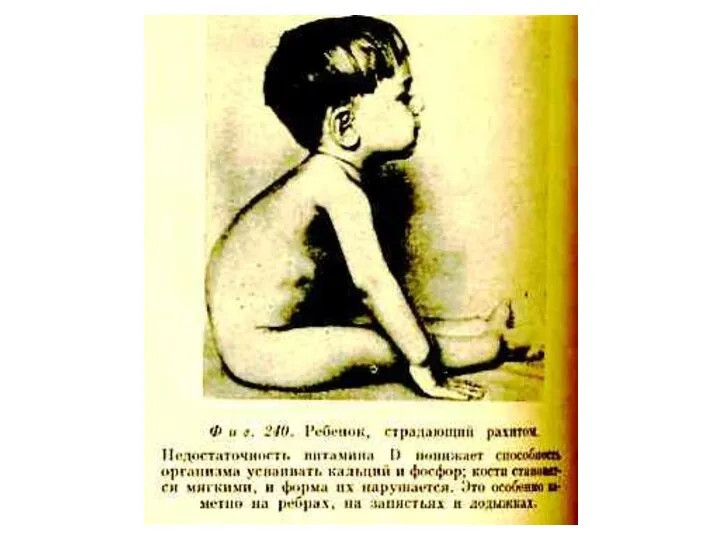
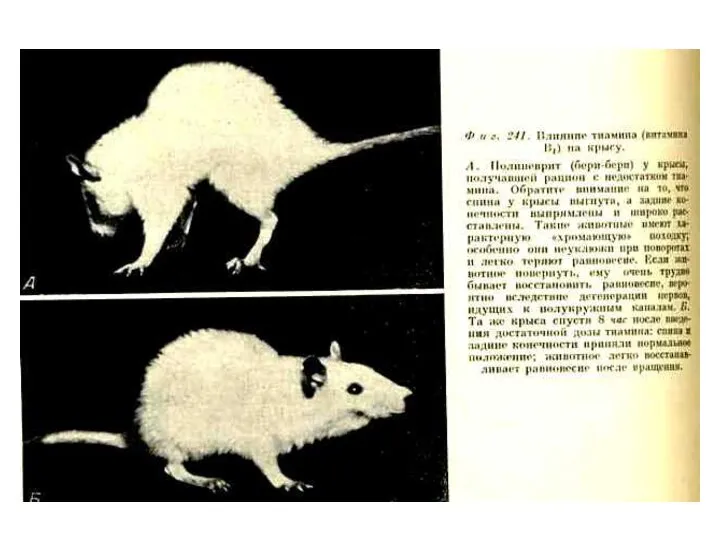
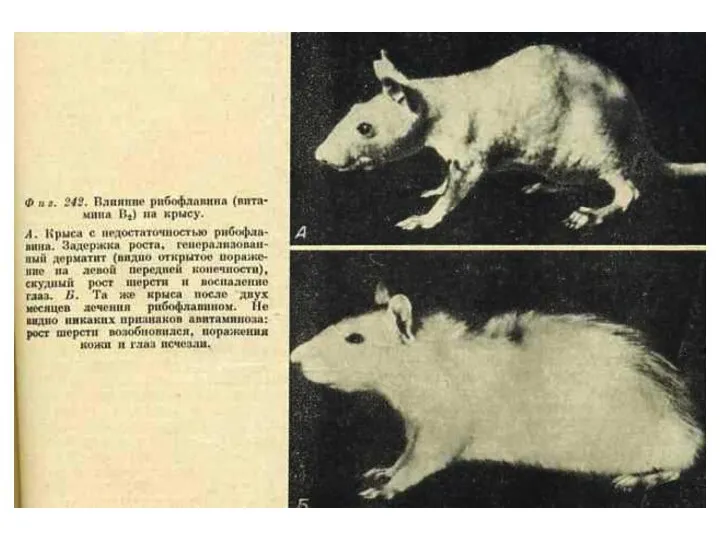
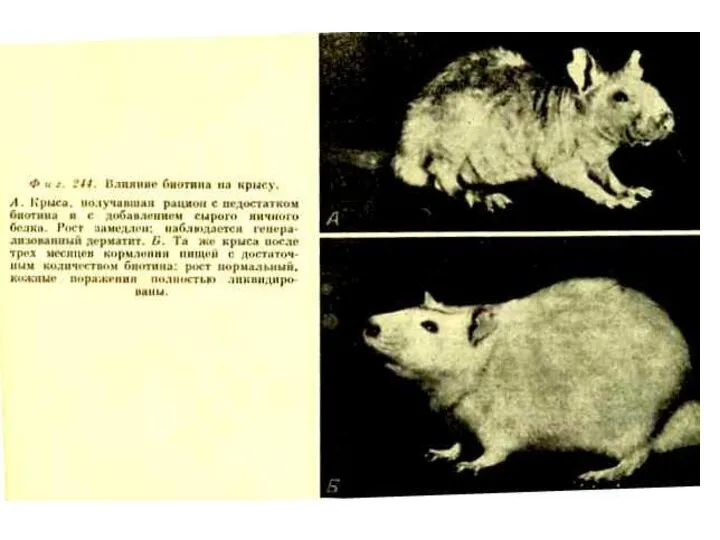

Метаболизм витаминов
Обмен очень интенсивный. Не накапливаются! (Исключение – жирорастворимые витамины, частично
Метаболизм витаминов
Обмен очень интенсивный. Не накапливаются! (Исключение – жирорастворимые витамины, частично

Источники витаминов:
Пищевые: желток яиц, печень, икра, проростки злаков, дрожжи.
Микрофлора кишечника.
КАК СОХРАНИТЬ
Источники витаминов:
Пищевые: желток яиц, печень, икра, проростки злаков, дрожжи.
Микрофлора кишечника.
КАК СОХРАНИТЬ

Причины гиповитаминозов:
Первичный (экзогенный) гипо- или авитаминоз: алиментарная недостаточность. Голодание, «однобокая» диета,
Причины гиповитаминозов:
Первичный (экзогенный) гипо- или авитаминоз: алиментарная недостаточность. Голодание, «однобокая» диета,

Причины гиповитаминозов:
Нарушение всасывания в ЖКТ жирорастворимых витаминов (снижение желчеобразования); В12 (недостаток
Причины гиповитаминозов:
Нарушение всасывания в ЖКТ жирорастворимых витаминов (снижение желчеобразования); В12 (недостаток

Взаимодействие витаминов
Витамин влияет на метаболизм другого витамина (Токоферол препятствует переокислению витамина
Взаимодействие витаминов
Витамин влияет на метаболизм другого витамина (Токоферол препятствует переокислению витамина

ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
А (ретинол, ретиналь), антиксерофтальмический.
Источник: продукты животного происхождения (печень рыб);растительные
ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
А (ретинол, ретиналь), антиксерофтальмический.
Источник: продукты животного происхождения (печень рыб);растительные

ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
Д (кальциферол), антирахитический.
Источники: эндогенный синтез (УФ), продукты животного происхождения, дрожжи
ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
Д (кальциферол), антирахитический.
Источники: эндогенный синтез (УФ), продукты животного происхождения, дрожжи

ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
Е (токоферол), антистерильный
Источники: растительные масла.
Обеспечивает стабильность клеточных мембран,
ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ.
Е (токоферол), антистерильный
Источники: растительные масла.
Обеспечивает стабильность клеточных мембран,

ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. К (филлохиноны), антигеморрагический
Источники: зелень, печень, дрожжи, синтез микрофлорой.
Является кофактором
ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ. К (филлохиноны), антигеморрагический
Источники: зелень, печень, дрожжи, синтез микрофлорой.
Является кофактором

Водорастворимые витамины.
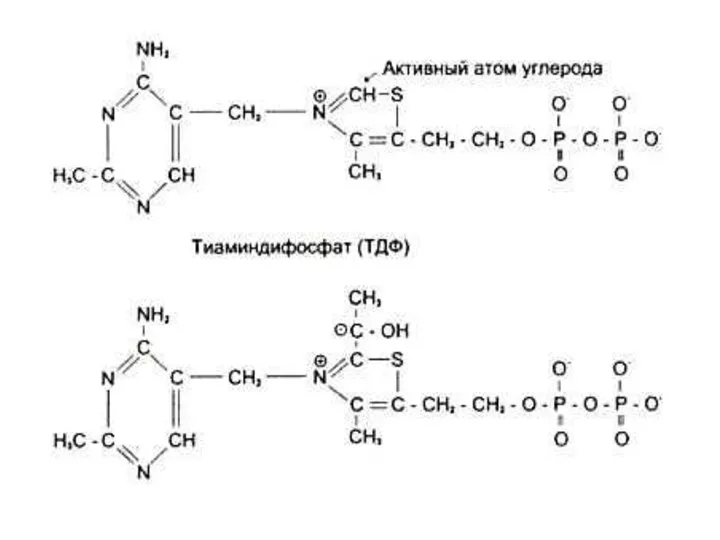
В1 (тиамин), антиневритный
Кофакторная форма – ТДФ (тиаминдифосфат)
Работает в составе дегидрогеназных
Водорастворимые витамины.
В1 (тиамин), антиневритный
Кофакторная форма – ТДФ (тиаминдифосфат)
Работает в составе дегидрогеназных

Водорастворимые витамины.
В2 (рибофлавин), витамин роста
Образует кофакторы ФМН, ФАД.
Работает в составе анаэробных
Водорастворимые витамины.
В2 (рибофлавин), витамин роста
Образует кофакторы ФМН, ФАД. Работает в составе анаэробных
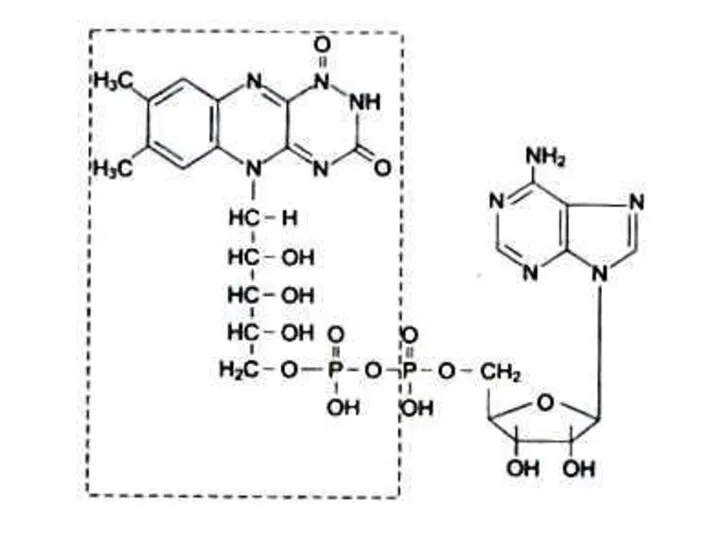
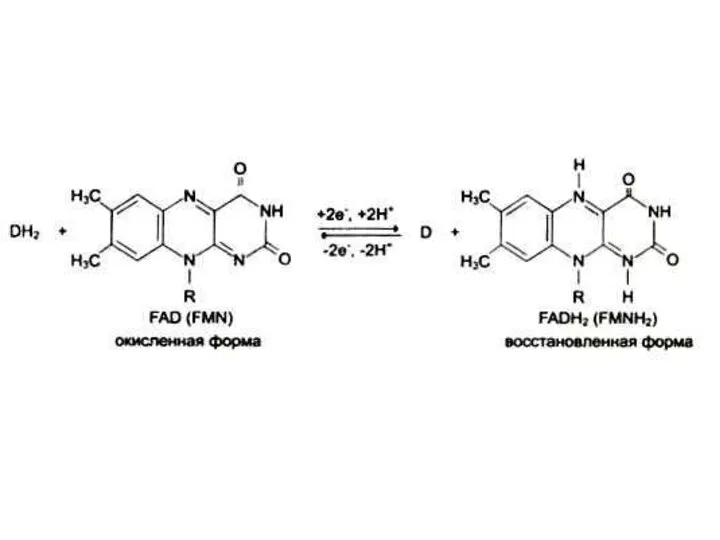

Водорастворимые витамины.
В3 (пантотеновая кислота), универсальный витамин.
Входит в состав КоА, активатора и
Водорастворимые витамины.
В3 (пантотеновая кислота), универсальный витамин.
Входит в состав КоА, активатора и

Водорастворимые витамины.
В5, РР (никотинамид), антипеллагрический.
Образует кофакторы НАД, НАДФ.
Входит в состав
Водорастворимые витамины.
В5, РР (никотинамид), антипеллагрический.
Образует кофакторы НАД, НАДФ.
Входит в состав
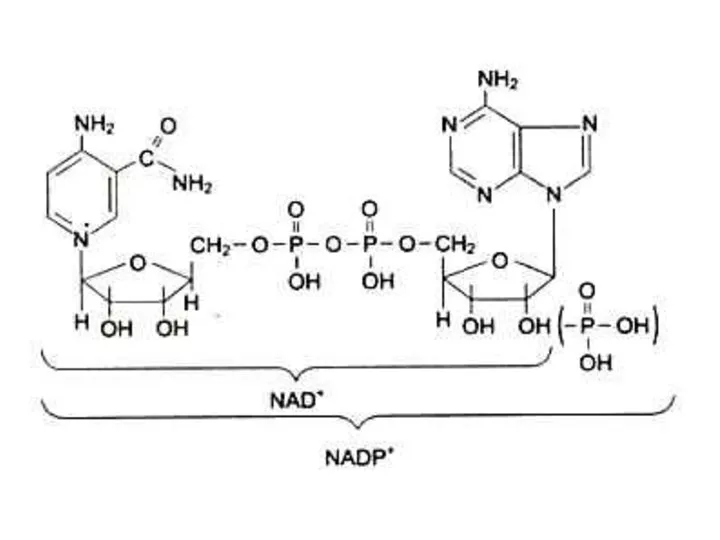
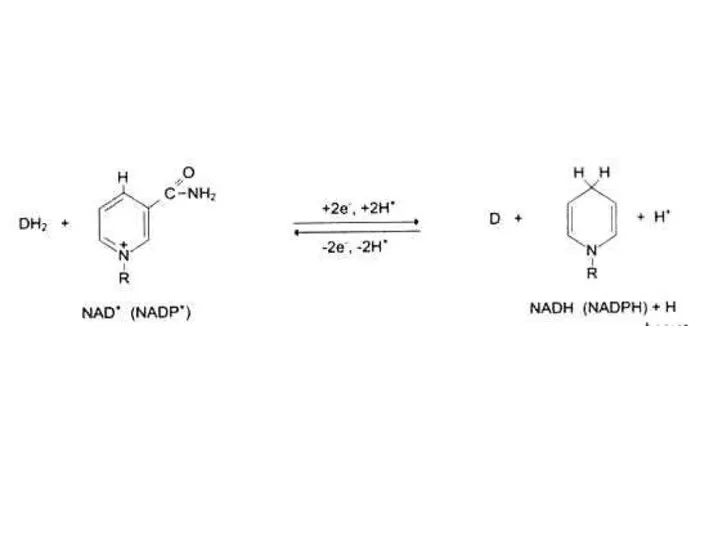

Водорастворимые витамины.
В6(пиридоксаль), антидерматитный
Образует кофактор пиридоксальфосфат.
Входит в состав трансфераз и декарбоксилаз аминокислот,
Водорастворимые витамины.
В6(пиридоксаль), антидерматитный
Образует кофактор пиридоксальфосфат.
Входит в состав трансфераз и декарбоксилаз аминокислот,

Водорастворимые витамины.
Н (биотин), антисеборейный.
Водорастворимые витамины.
Н (биотин), антисеборейный.
 Факторы, влияющие на потребность организации в персонале
Факторы, влияющие на потребность организации в персонале Оптимизация полномочий субъектов Российской Федерации и муниципальных образований
Оптимизация полномочий субъектов Российской Федерации и муниципальных образований Программирование на алгоритмическом языке. Часть III (9 класс)
Программирование на алгоритмическом языке. Часть III (9 класс) Организация технического обслуживания и ремонта автомобиля Тойота Ленд Крузер 200
Организация технического обслуживания и ремонта автомобиля Тойота Ленд Крузер 200 Экономика Франции
Экономика Франции Поэзия второй половины XIX века
Поэзия второй половины XIX века  Механизмы ввода и вывода информации. Понятие сериализации
Механизмы ввода и вывода информации. Понятие сериализации Спид и спид- индикаторные болезни человека
Спид и спид- индикаторные болезни человека Кафедра «Финансы и налоги» Преподаватель: Просалова Вероника Сергеевна, к.э.н., доцент «БЮДЖЕТНОЕ ПЛАНИРОВАНИЕ И ПРОГНОЗИРОВА
Кафедра «Финансы и налоги» Преподаватель: Просалова Вероника Сергеевна, к.э.н., доцент «БЮДЖЕТНОЕ ПЛАНИРОВАНИЕ И ПРОГНОЗИРОВА Аттестационная работа. Индивидуальная карта развития в области культуры здоровья
Аттестационная работа. Индивидуальная карта развития в области культуры здоровья Презентация Источники финансирования инвестиций
Презентация Источники финансирования инвестиций Расчет количества обоев для спальни и коридора
Расчет количества обоев для спальни и коридора 100 шагов Президента
100 шагов Президента Проект «За страницами учебника» Творческое название проекта – ПТИЦЫ В КАРТИНАХ РУССКИХ ХУДОЖНИКОВ. Выполнил ученик 11 б класса
Проект «За страницами учебника» Творческое название проекта – ПТИЦЫ В КАРТИНАХ РУССКИХ ХУДОЖНИКОВ. Выполнил ученик 11 б класса  ВВЕДЕНИЕ В ВЫЧИСЛИТЕЛЬНУЮ МАТЕМАТИКУ Лекция 3 22 сентября 2009 ВЫЧИСЛИТЕЛЬНАЯ ЛИНЕЙНАЯ АЛГЕБРА
ВВЕДЕНИЕ В ВЫЧИСЛИТЕЛЬНУЮ МАТЕМАТИКУ Лекция 3 22 сентября 2009 ВЫЧИСЛИТЕЛЬНАЯ ЛИНЕЙНАЯ АЛГЕБРА  Компенсация реактивной мощности. (БСК). Лекция 04
Компенсация реактивной мощности. (БСК). Лекция 04 Международные конфликты: причины и сущность
Международные конфликты: причины и сущность Бизнес-план компьютерной мастерской
Бизнес-план компьютерной мастерской Кодирование
Кодирование Биохимия ферментов-2
Биохимия ферментов-2 Поступление в БПОУ ВО «Грязовецкий политехнический техникум»
Поступление в БПОУ ВО «Грязовецкий политехнический техникум»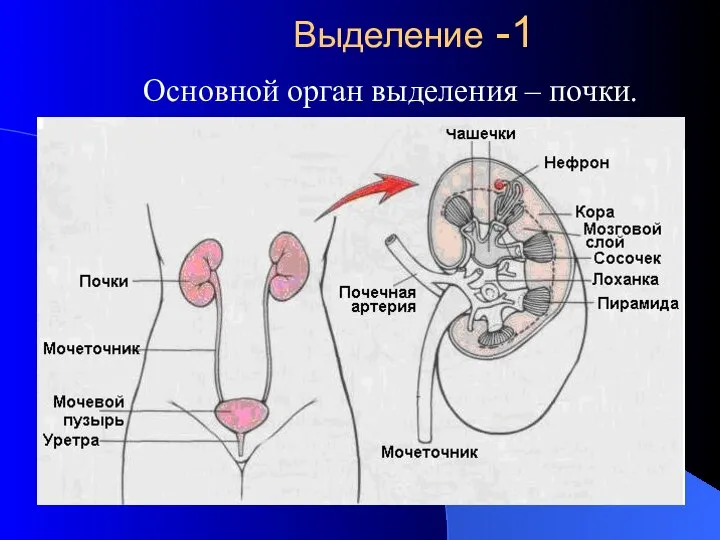 Выделение
Выделение  Городецкая роспись как средство арт терапии и цветотерапии Выполнили ученицы 6Б класса МОУ Школы №55 Бочкарёва Анна Мельникова
Городецкая роспись как средство арт терапии и цветотерапии Выполнили ученицы 6Б класса МОУ Школы №55 Бочкарёва Анна Мельникова  Развитие территориального общественного самоуправления в Юсьвинском сельском поселении
Развитие территориального общественного самоуправления в Юсьвинском сельском поселении Общие сведения о зубчатых передачах
Общие сведения о зубчатых передачах Информация о NOVADAN (www.NOVADAN.DK) NOVADAN (Дания) является одним из европейских лидеров среди производителей моющих средств, дезинфектантов
Информация о NOVADAN (www.NOVADAN.DK) NOVADAN (Дания) является одним из европейских лидеров среди производителей моющих средств, дезинфектантов  Продвижение интернет-магазина: бюджетный маркетинг
Продвижение интернет-магазина: бюджетный маркетинг Сахарный диабет
Сахарный диабет