- Фізіологія збудження(ССО)

Содержание
- 2. Передача їнформації – серія нервових імпульсів (ПД), що поширюються по нервовим волокнам Нервові волокна – відростки
- 3. БУДОВА НЕМІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН Складається з 7 – 12 тонких аксонів, які оточені шваннівською клітиною Аксоплазма
- 4. БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН Мієлінова оболонка – результат багаторазового обертання шваннівської клітини навколо осьового циліндра і
- 5. БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН Ділянки мембрани, позбавлені мієліну (1мкм) – перехвати Ранвьє Відстань між перехватами пропорційна
- 6. Формування мієлінової оболонки навколо аксона на різних стадіях йго развитку (А – Г) ( Судаков, 2000)
- 7. БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН 1. ЯДРО Мієлінова оболонка Перехват Ранвьє АКСОН Шванівська клітина
- 8. БУДОВА НЕРВОВИХ ВОЛОКОН
- 9. ПОШИРЕННЯ ПОТЕНЦІАЛУ ДІЇ (ЗБУДЖЕННЯ) Механізм поширення збудження вздовж нервового або м’язового волокна - утворення місцевих струмів,
- 10. МЕХАНІЗМ ПОШИРЕННЯ ЗБУДЖЕННЯ У БЕЗМІЄЛІНОВОМУ ВОЛОКНІ – ПОСЛІДОВНА ДЕПОЛЯРИЗАЦІЯ ДІЛЯНОК МЕМБРАНИ В кожній ділянці волокна: виникає
- 11. збуджені раніше ділянки мембрани повертаються в стан спокою місцевий струм знижує мембранний потенціал незбудженої ділянки внаслідок
- 12. В кожному перехваті послідовно виникають: порогова деполяризація вхід Na в аксон виникнення зони позитивного заряду МЕХАНІЗМ
- 13. В кожному перехваті послідовно виникають: МЕХАНІЗМ ПРОВЕДЕНЯ ЗБУДЖЕННЯ В МІЄЛІНИЗОВАНОМУ ВОЛОКНІ виникнення місцевих деполяризуючих струмів, що
- 14. Імпульс переміщується стрибкоподібно від одного перехвату до іншого – сальтоторне проведення в стан збудження може перейти
- 15. Закони розповсюдження збудження по нервових волокнах: Анатомічної та фізіологічної безперервності волокна Проведення збудження можливо лише за
- 16. Закони розповсюдження збудження по нервових волокнах 2. Двохстороннього проведення збудження Збудження, що виникло у якій-небудь області
- 17. Закони розповсюдження збудження по нервових волокнах Ізольованого проведення збудження Збудження, що виникло у нервовому волокні, не
- 18. Розповсюдження збудження по нервових волокнах:
- 19. Типи нервових волокон, їх властивості та функціональне призначення Загальні властивості нервових волокон • практично невтомні •
- 20. ФІЗІОЛОГІЯ М’ЯЗІВ
- 21. ВИДИ М’ЯЗІВ Посмуговані (скелетні) Непосмуговані (гладенькі) СЕРЦЕВИЙ М'ЯЗ
- 22. забезпечення пози переміщення тіла у просторі переміщення окремих частин тіла джерело тепла – терморегуляція збудливість провідність
- 24. БУДОВА М’ЯЗА: 1 — м’яз; 2 — пучки; 3 — перимізій; 4 — капіляр; 5 —
- 25. Схема будови відрізка м'язового волокна 1 — саркоплазматичний ретикулум 2 — термінальні цистерни саркоплазматичного ретикулуму 3
- 26. . Саркомер складається із Т-телофрагми — лінії Z (для двох сусідніх саркомерів), половин І - та
- 28. ОСНОВНІ БІЛКИ МІОФІБРІЛ
- 29. АКТИН
- 31. Міозиновий філамент Молекула міозина 1 - фібрилярний стержень 2 - головки МІОЗИН
- 35. РЕЖИМИ М’ЯЗОВИХ СКОРОЧЕНЬ Одиночне скорочення – відповідь на одиночний імпульс. Швидкість з якою відбувається скорочення пропорційна
- 36. МЕХАНІЗМ М’ЯЗОВОГО СКОРОЧЕННЯ: Подразнення → Виникнення ПД → проведення ПД уздовж клітинної мембрани та усередину волокна
- 37. Механізм м’язового скорочення Подразнення. Електричне збудження в нервово - м’язовому cинапсі – Виникнення ПД м’яза Проведення
- 38. Механізм м’язового скорочення Під дією енергії АТФ головки міозину змінюють положення у просторі, що викликає ковзання
- 40. ВИТРАТИ АТФ: На роботу Na+-К+ насоса; На роботу Са++ насоса; На розрив містків актину та міозину.
- 41. МЕХАНІЗМ М'ЯЗОВОГО СКОРОЧЕННЯ
- 42. ТИПИ М’ЯЗОВИХ СКОРОЧЕНЬ
- 43. Співвідношення між збудженням і скороченням
- 44. СУММАЦІЯ СКОРОЧЕНЬ Тетанічне скорочення – відповідь на ряд послідовних імпульсів – сумація одиночних скорочень При відносно
- 45. Латентний період Укорочення або розвиток напруги Розслаблення а. Одиночне скорочення б. Зубчатий тетанус в. Перехід зубчатого
- 46. Синапс. Будова та механізм передачі збудження в хімічному синапсі. Національний фармацевтичний університет Кафедра фізіології та анатомії
- 47. КЛАСИФІКАЦІЯ СИНАПСІВ ЗА МІСЦЕЗНАХОДЖЕННЯМ нервово- м’язові Синапс місце функціонального контакту двох збудливих тканин, одна з яких
- 48. НЕЙРО-НЕЙРОНАЛЬНІ СИНАПСИ
- 50. ЗА ДІЄЮ збуджуючі гальмівні під дією медіатора відчиняються Na - та K – канали, відбувається деполяризація
- 51. Електричний синапс
- 52. БУДОВА ХІМІЧНОГО СИНАПСА Пресинаптична частина: Кінцева гілочка аксона, яка втрачає мієлінову оболонку і розширюється – цибулеподібна
- 53. Хемозбудливі іонні канали
- 54. БУДОВА ХІМІЧНОГО СИНАПСА
- 57. медіатори Медіатор (посередник) - хімічна речовина, яка забезпечує однобічну передачу збудження в хімічному синапсі Ферменти, необхідні
- 58. медіатори За хімічною структурою • моноаміни (адреналін, норадреналін, ацетилхолін та інш) • амінокислоти (гамма-аміномасляна кислота (ГАМК),
- 59. Етапи синаптичної передачі Медіатор, що міститься у синаптичних пухирцях (везикулах) утворюється або в тілі нейрона або
- 60. Етапи синаптичної передачі Дифузія молекул медіатора через синаптичну щілину 6. Молекули медіатора зв’язуються з рецепторами постсинаптичної
- 61. Хемозбудливі іонні канали
- 62. Етапи синаптичної передачі 7. Молекули медіатора після дії на рецептор вилучаються із синаптичної щілини шляхом реабсорбції
- 63. Властивості хімічного синапса Однобічне проведення збудження від пресинаптичної мембрани до посинаптичної Висока втома пов’язана з виснаженням
- 66. Скачать презентацию

Передача їнформації – серія нервових імпульсів (ПД), що поширюються по нервовим
Передача їнформації – серія нервових імпульсів (ПД), що поширюються по нервовим

БУДОВА НЕМІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Складається з 7 – 12 тонких аксонів, які
БУДОВА НЕМІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Складається з 7 – 12 тонких аксонів, які
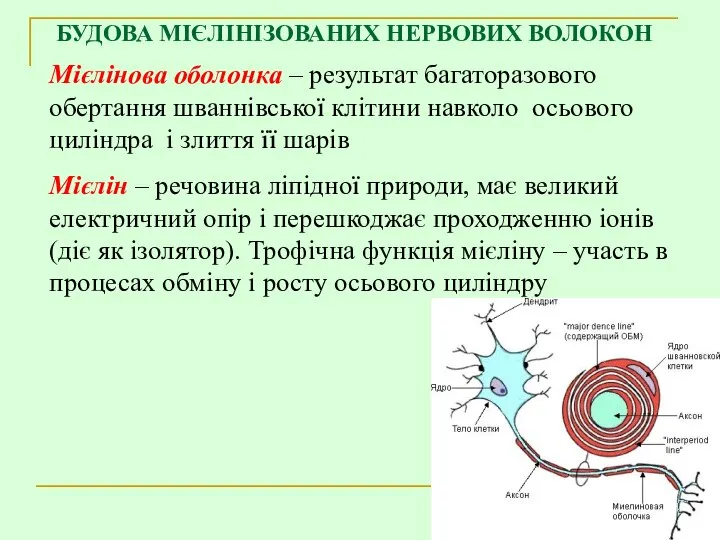
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Мієлінова оболонка – результат багаторазового обертання шваннівської клітини
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Мієлінова оболонка – результат багаторазового обертання шваннівської клітини
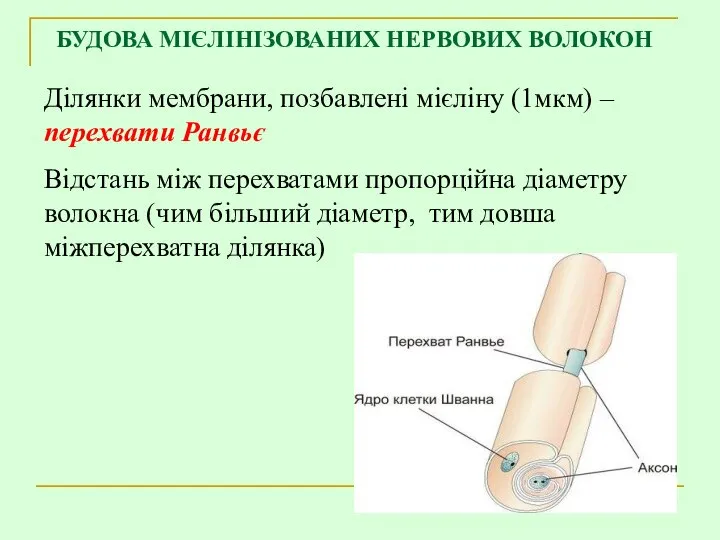
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Ділянки мембрани, позбавлені мієліну (1мкм) – перехвати Ранвьє
Відстань
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
Ділянки мембрани, позбавлені мієліну (1мкм) – перехвати Ранвьє
Відстань

Формування мієлінової оболонки навколо аксона на різних стадіях йго развитку (А – Г)
Формування мієлінової оболонки навколо аксона на різних стадіях йго развитку (А – Г)
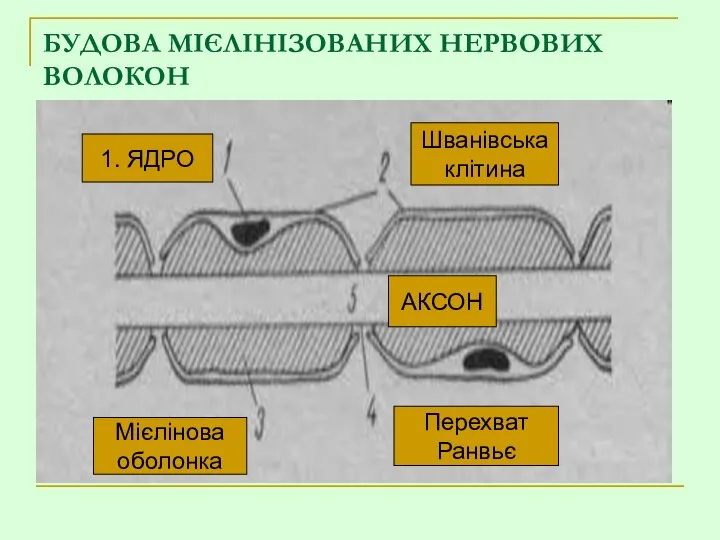
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
1. ЯДРО
Мієлінова
оболонка
Перехват
Ранвьє
АКСОН
Шванівська
клітина
БУДОВА МІЄЛІНІЗОВАНИХ НЕРВОВИХ ВОЛОКОН
1. ЯДРО
Мієлінова
оболонка
Перехват
Ранвьє
АКСОН
Шванівська
клітина

БУДОВА НЕРВОВИХ ВОЛОКОН
БУДОВА НЕРВОВИХ ВОЛОКОН

ПОШИРЕННЯ ПОТЕНЦІАЛУ ДІЇ
(ЗБУДЖЕННЯ)
Механізм поширення збудження вздовж нервового або м’язового волокна
ПОШИРЕННЯ ПОТЕНЦІАЛУ ДІЇ
(ЗБУДЖЕННЯ)
Механізм поширення збудження вздовж нервового або м’язового волокна

МЕХАНІЗМ ПОШИРЕННЯ ЗБУДЖЕННЯ У БЕЗМІЄЛІНОВОМУ ВОЛОКНІ – ПОСЛІДОВНА ДЕПОЛЯРИЗАЦІЯ ДІЛЯНОК МЕМБРАНИ
В
МЕХАНІЗМ ПОШИРЕННЯ ЗБУДЖЕННЯ У БЕЗМІЄЛІНОВОМУ ВОЛОКНІ – ПОСЛІДОВНА ДЕПОЛЯРИЗАЦІЯ ДІЛЯНОК МЕМБРАНИ
В
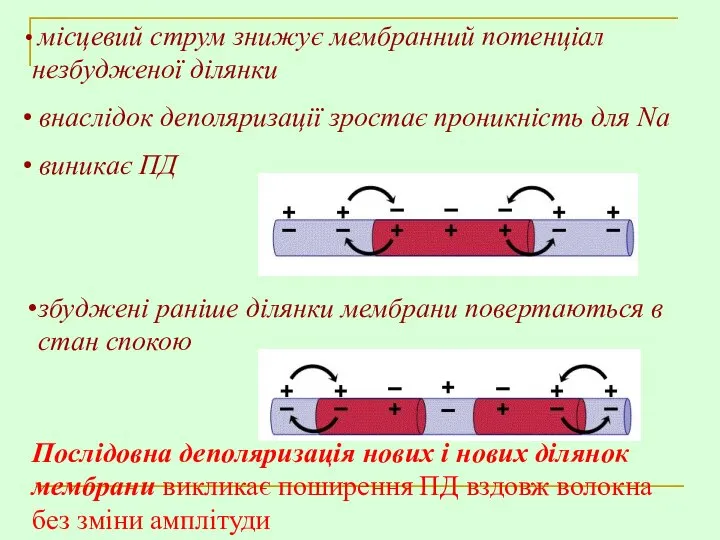
збуджені раніше ділянки мембрани повертаються в стан спокою
місцевий струм знижує
збуджені раніше ділянки мембрани повертаються в стан спокою
місцевий струм знижує
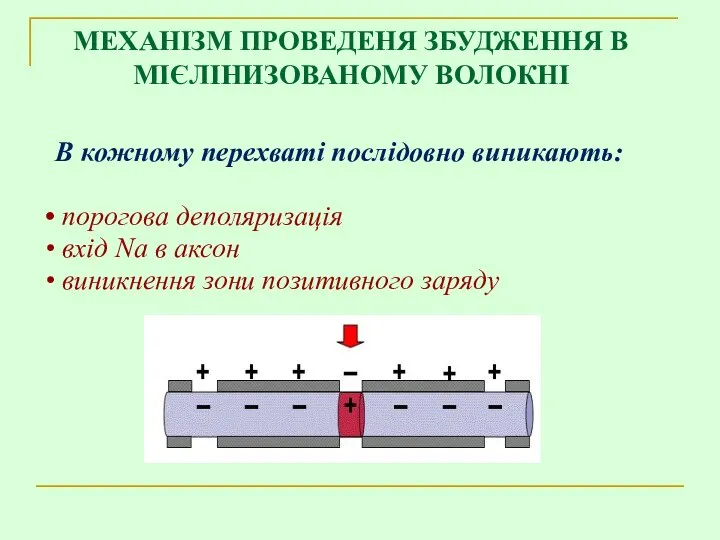
В кожному перехваті послідовно виникають:
порогова деполяризація
вхід Na в аксон
В кожному перехваті послідовно виникають:
порогова деполяризація
вхід Na в аксон
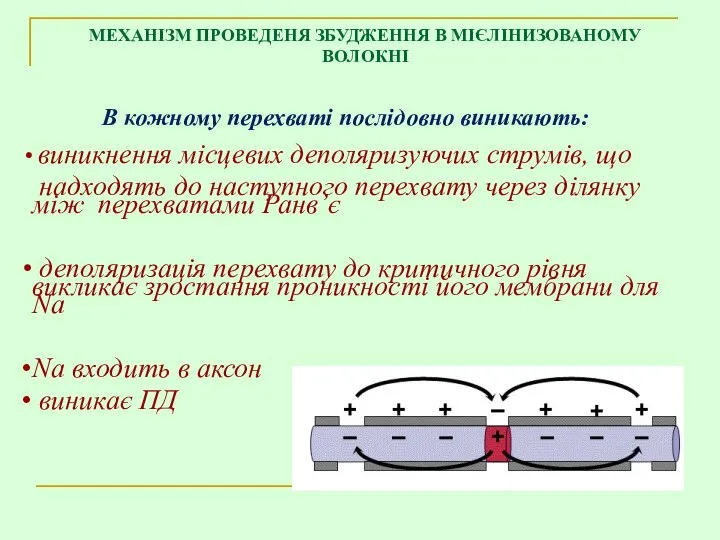
В кожному перехваті послідовно виникають:
МЕХАНІЗМ ПРОВЕДЕНЯ ЗБУДЖЕННЯ В МІЄЛІНИЗОВАНОМУ ВОЛОКНІ
виникнення
В кожному перехваті послідовно виникають:
МЕХАНІЗМ ПРОВЕДЕНЯ ЗБУДЖЕННЯ В МІЄЛІНИЗОВАНОМУ ВОЛОКНІ
виникнення
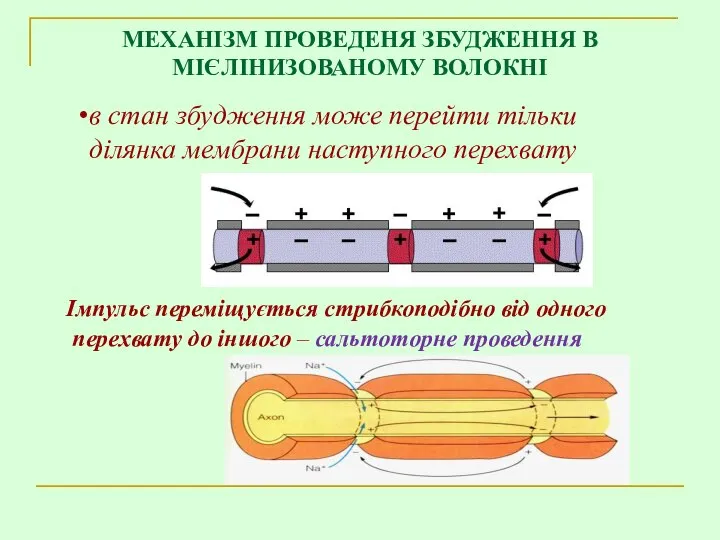
Імпульс переміщується стрибкоподібно від одного
перехвату до іншого – сальтоторне проведення
в
Імпульс переміщується стрибкоподібно від одного
перехвату до іншого – сальтоторне проведення
в

Закони розповсюдження збудження по нервових волокнах:
Анатомічної та фізіологічної безперервності волокна
Проведення збудження
Закони розповсюдження збудження по нервових волокнах:
Анатомічної та фізіологічної безперервності волокна
Проведення збудження

Закони розповсюдження збудження по нервових волокнах
2. Двохстороннього проведення збудження
Збудження, що виникло
Закони розповсюдження збудження по нервових волокнах
2. Двохстороннього проведення збудження
Збудження, що виникло

Закони розповсюдження збудження по нервових волокнах
Ізольованого проведення збудження
Збудження, що виникло у
Закони розповсюдження збудження по нервових волокнах
Ізольованого проведення збудження
Збудження, що виникло у
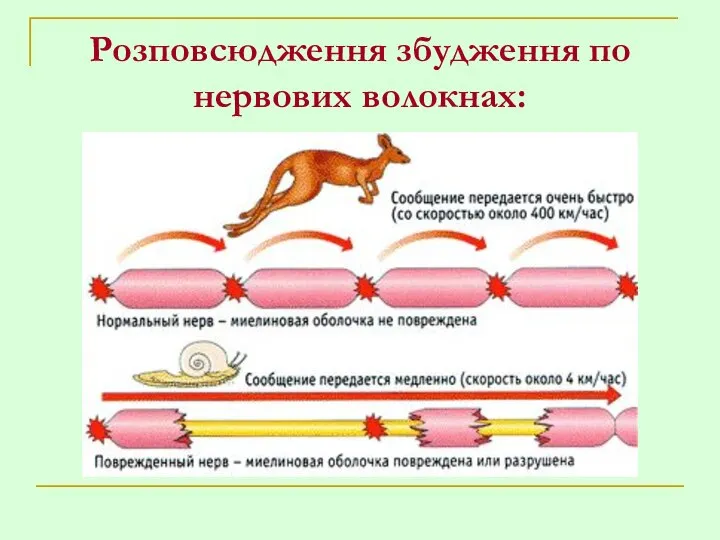
Розповсюдження збудження по нервових волокнах:
Розповсюдження збудження по нервових волокнах:

Типи нервових волокон, їх властивості та функціональне призначення
Загальні властивості нервових волокон
•
Типи нервових волокон, їх властивості та функціональне призначення
Загальні властивості нервових волокон
•

ФІЗІОЛОГІЯ М’ЯЗІВ
ФІЗІОЛОГІЯ М’ЯЗІВ

ВИДИ М’ЯЗІВ
Посмуговані
(скелетні)
Непосмуговані
(гладенькі)
СЕРЦЕВИЙ
М'ЯЗ
ВИДИ М’ЯЗІВ
Посмуговані
(скелетні)
Непосмуговані
(гладенькі)
СЕРЦЕВИЙ
М'ЯЗ

забезпечення пози
переміщення тіла у просторі
переміщення окремих частин тіла
забезпечення пози
переміщення тіла у просторі
переміщення окремих частин тіла
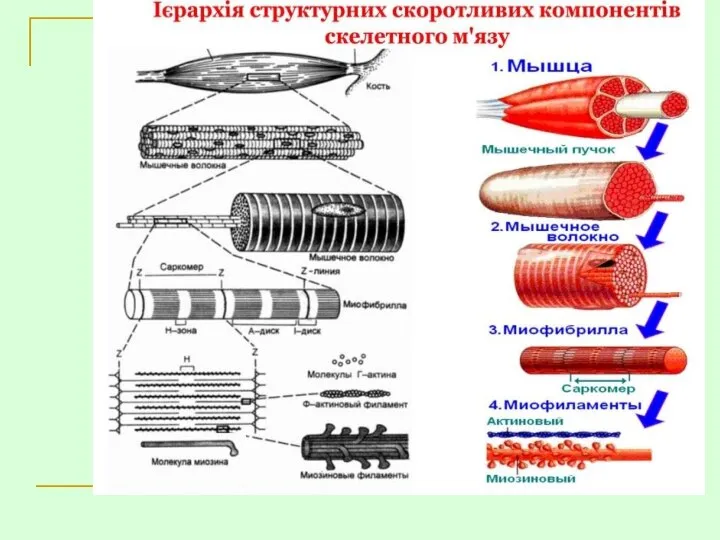
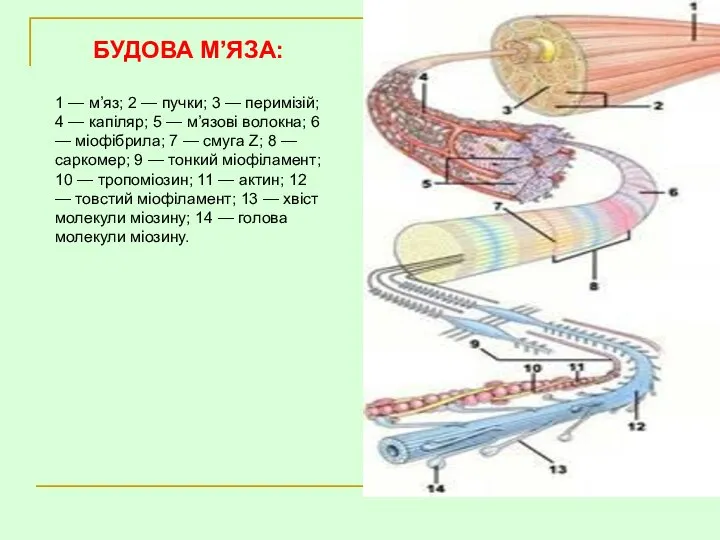
БУДОВА М’ЯЗА:
1 — м’яз; 2 — пучки; 3 — перимізій;
БУДОВА М’ЯЗА:
1 — м’яз; 2 — пучки; 3 — перимізій;

Схема будови відрізка м'язового волокна
1 — саркоплазматичний ретикулум
2 — термінальні
Схема будови відрізка м'язового волокна
1 — саркоплазматичний ретикулум
2 — термінальні
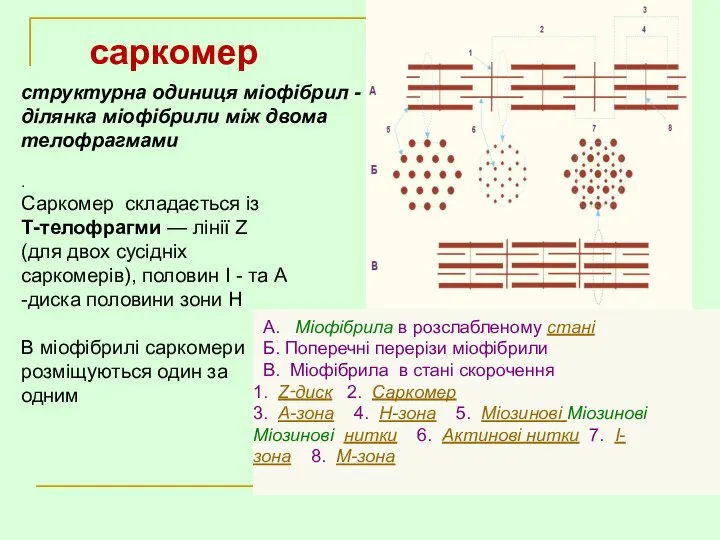
.
Саркомер складається із
Т-телофрагми — лінії Z (для двох сусідніх саркомерів), половин
.
Саркомер складається із
Т-телофрагми — лінії Z (для двох сусідніх саркомерів), половин


ОСНОВНІ БІЛКИ МІОФІБРІЛ
ОСНОВНІ БІЛКИ МІОФІБРІЛ
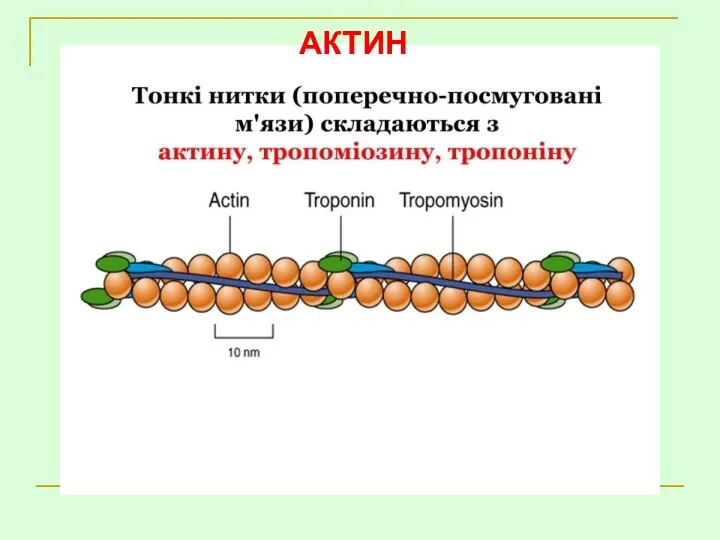
АКТИН
АКТИН


Міозиновий філамент
Молекула міозина
1 - фібрилярний стержень
2 - головки
МІОЗИН
Міозиновий філамент
Молекула міозина
1 - фібрилярний стержень
2 - головки
МІОЗИН




РЕЖИМИ М’ЯЗОВИХ СКОРОЧЕНЬ
Одиночне скорочення – відповідь на одиночний імпульс. Швидкість з
РЕЖИМИ М’ЯЗОВИХ СКОРОЧЕНЬ
Одиночне скорочення – відповідь на одиночний імпульс. Швидкість з

МЕХАНІЗМ М’ЯЗОВОГО СКОРОЧЕННЯ:
Подразнення → Виникнення ПД → проведення ПД уздовж клітинної
МЕХАНІЗМ М’ЯЗОВОГО СКОРОЧЕННЯ:
Подразнення → Виникнення ПД → проведення ПД уздовж клітинної

Механізм м’язового скорочення
Подразнення. Електричне збудження в нервово - м’язовому cинапсі –
Механізм м’язового скорочення
Подразнення. Електричне збудження в нервово - м’язовому cинапсі –

Механізм м’язового скорочення
Під дією енергії АТФ головки міозину змінюють положення у
Механізм м’язового скорочення
Під дією енергії АТФ головки міозину змінюють положення у


ВИТРАТИ АТФ:
На роботу Na+-К+ насоса;
На роботу Са++ насоса;
На розрив містків актину
ВИТРАТИ АТФ:
На роботу Na+-К+ насоса;
На роботу Са++ насоса;
На розрив містків актину
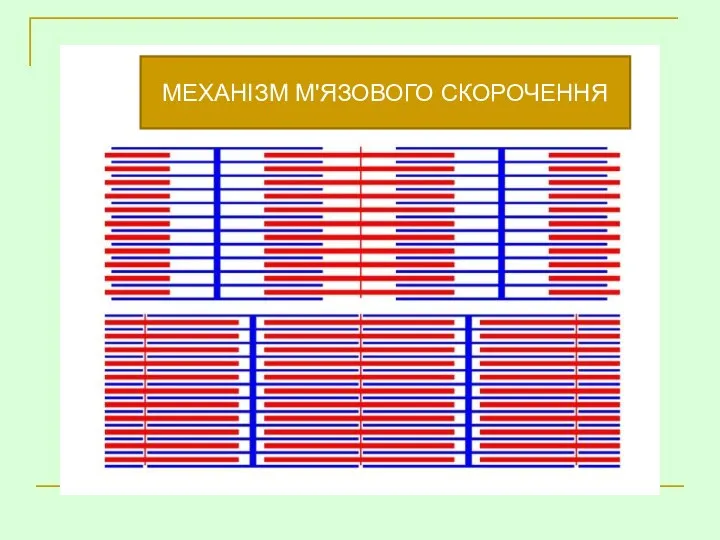
МЕХАНІЗМ М'ЯЗОВОГО СКОРОЧЕННЯ
МЕХАНІЗМ М'ЯЗОВОГО СКОРОЧЕННЯ

ТИПИ М’ЯЗОВИХ СКОРОЧЕНЬ
ТИПИ М’ЯЗОВИХ СКОРОЧЕНЬ

Співвідношення між збудженням і скороченням
Співвідношення між збудженням і скороченням
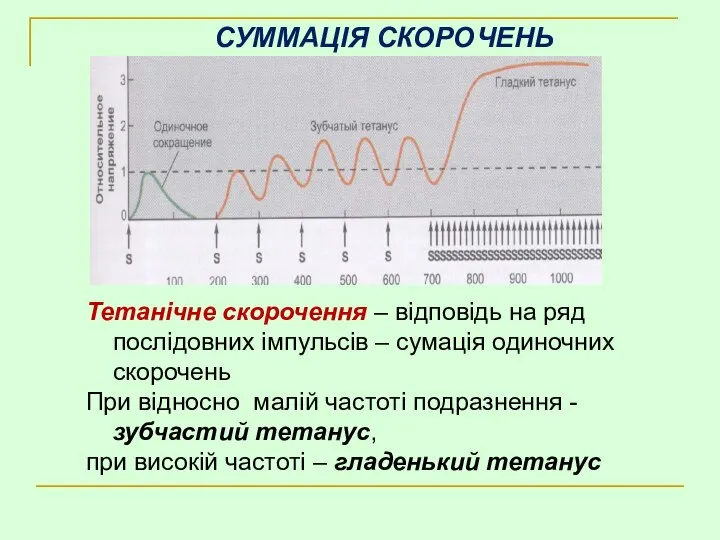
СУММАЦІЯ СКОРОЧЕНЬ
Тетанічне скорочення – відповідь на ряд послідовних імпульсів – сумація
СУММАЦІЯ СКОРОЧЕНЬ
Тетанічне скорочення – відповідь на ряд послідовних імпульсів – сумація

Латентний період
Укорочення або розвиток напруги
Розслаблення
а. Одиночне скорочення
б. Зубчатий тетанус
Латентний період
Укорочення або розвиток напруги
Розслаблення
а. Одиночне скорочення
б. Зубчатий тетанус
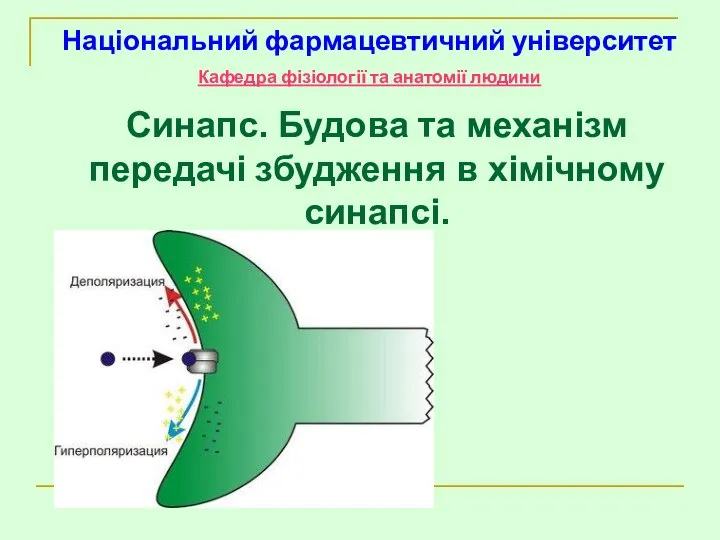
Синапс. Будова та механізм передачі збудження в хімічному синапсі.
Національний фармацевтичний університет
Кафедра
Синапс. Будова та механізм передачі збудження в хімічному синапсі.
Національний фармацевтичний університет
Кафедра
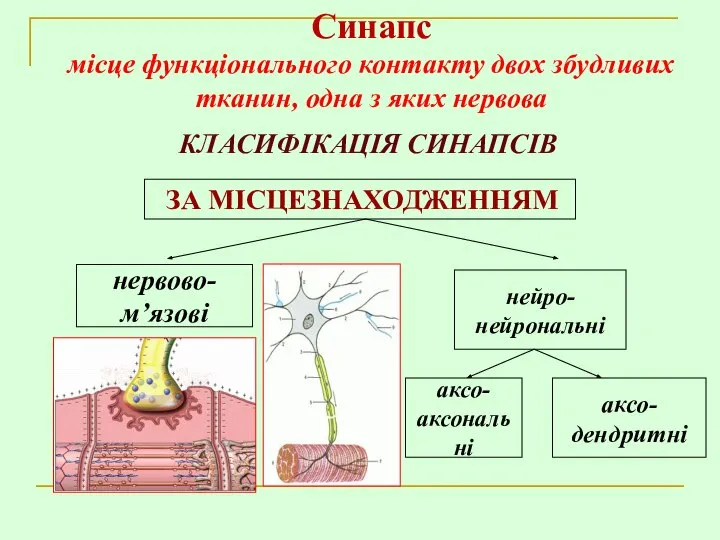
КЛАСИФІКАЦІЯ СИНАПСІВ
ЗА МІСЦЕЗНАХОДЖЕННЯМ
нервово-
м’язові
Синапс
місце функціонального контакту двох збудливих тканин, одна
КЛАСИФІКАЦІЯ СИНАПСІВ
ЗА МІСЦЕЗНАХОДЖЕННЯМ
нервово-
м’язові
Синапс
місце функціонального контакту двох збудливих тканин, одна
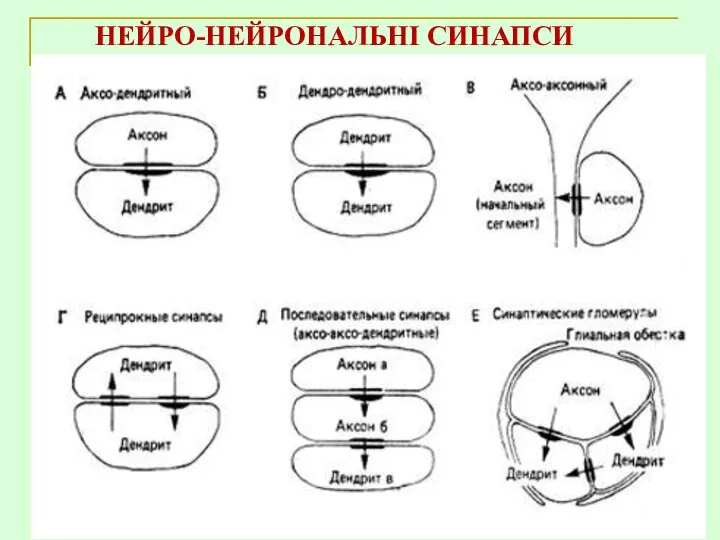
НЕЙРО-НЕЙРОНАЛЬНІ СИНАПСИ
НЕЙРО-НЕЙРОНАЛЬНІ СИНАПСИ

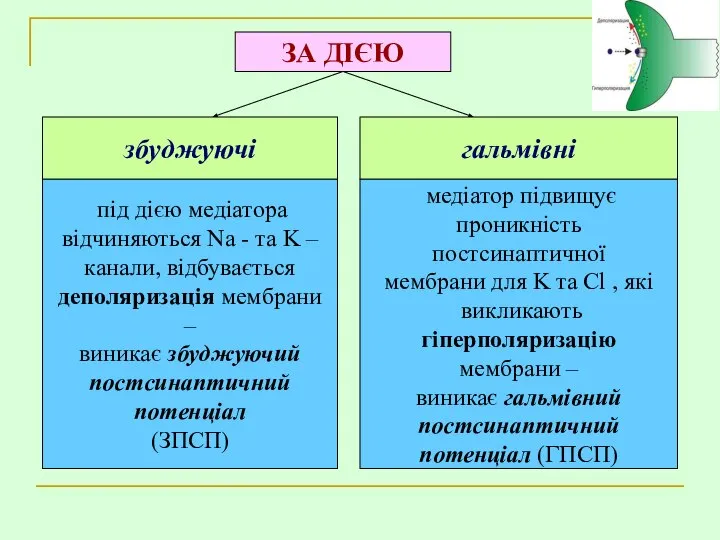
ЗА ДІЄЮ
збуджуючі
гальмівні
під дією медіатора
відчиняються Na - та K –
ЗА ДІЄЮ
збуджуючі
гальмівні
під дією медіатора
відчиняються Na - та K –

Електричний синапс
Електричний синапс

БУДОВА ХІМІЧНОГО СИНАПСА
Пресинаптична частина:
Кінцева гілочка аксона, яка втрачає мієлінову оболонку і
БУДОВА ХІМІЧНОГО СИНАПСА
Пресинаптична частина:
Кінцева гілочка аксона, яка втрачає мієлінову оболонку і

Хемозбудливі іонні канали
Хемозбудливі іонні канали
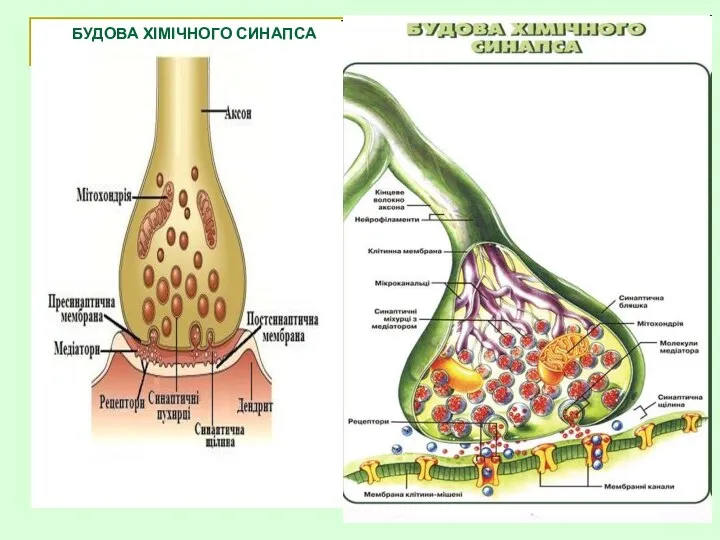
БУДОВА ХІМІЧНОГО СИНАПСА
БУДОВА ХІМІЧНОГО СИНАПСА



медіатори
Медіатор (посередник) - хімічна речовина, яка забезпечує однобічну передачу збудження в
медіатори
Медіатор (посередник) - хімічна речовина, яка забезпечує однобічну передачу збудження в

медіатори
За хімічною структурою
• моноаміни (адреналін, норадреналін, ацетилхолін та інш)
• амінокислоти (гамма-аміномасляна
медіатори
За хімічною структурою
• моноаміни (адреналін, норадреналін, ацетилхолін та інш)
• амінокислоти (гамма-аміномасляна

Етапи синаптичної передачі
Медіатор, що міститься у синаптичних пухирцях (везикулах) утворюється або
Етапи синаптичної передачі
Медіатор, що міститься у синаптичних пухирцях (везикулах) утворюється або

Етапи синаптичної передачі
Дифузія молекул медіатора
через синаптичну щілину
6. Молекули медіатора
Етапи синаптичної передачі
Дифузія молекул медіатора
через синаптичну щілину
6. Молекули медіатора
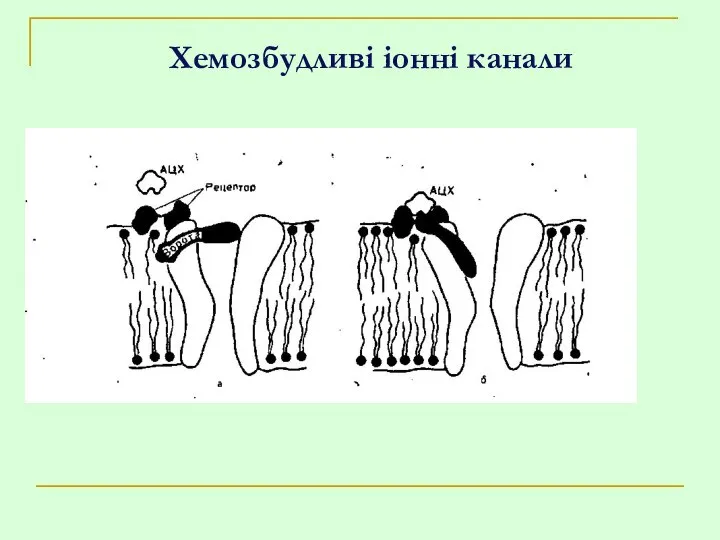
Хемозбудливі іонні канали
Хемозбудливі іонні канали

Етапи синаптичної передачі
7. Молекули медіатора після дії на рецептор вилучаються із
Етапи синаптичної передачі
7. Молекули медіатора після дії на рецептор вилучаються із

Властивості хімічного синапса
Однобічне проведення збудження від пресинаптичної мембрани до посинаптичної
Висока втома
Властивості хімічного синапса
Однобічне проведення збудження від пресинаптичної мембрани до посинаптичної
Висока втома

 Сердечно-сосудистая система Ангиология – наука о сосудах Сердце
Сердечно-сосудистая система Ангиология – наука о сосудах Сердце Ресайклеры. Курс базового уровня Б-005
Ресайклеры. Курс базового уровня Б-005 Презентация "Художественные открытия Серебряного века русской культуры" - скачать презентации по МХК
Презентация "Художественные открытия Серебряного века русской культуры" - скачать презентации по МХК Презентация "Применение спектрального анализа и различных видов излучения для определения подлинности произведений искусства
Презентация "Применение спектрального анализа и различных видов излучения для определения подлинности произведений искусства Проводники в электростатическом поле
Проводники в электростатическом поле Физиологическая основа внимания
Физиологическая основа внимания Акции на Одинцовских курганах 2019
Акции на Одинцовских курганах 2019 Самые выдающиеся запатентованные изобретения
Самые выдающиеся запатентованные изобретения  Направляющие прямолинейного движения с трением качения
Направляющие прямолинейного движения с трением качения Тема: ОТНОШЕНИЯ СОБСТВЕННОСТИ В РЫНОЧНОЙ ЭКОНОМИКЕ. Вопросы: Сущность собственности как экономической к
Тема: ОТНОШЕНИЯ СОБСТВЕННОСТИ В РЫНОЧНОЙ ЭКОНОМИКЕ. Вопросы: Сущность собственности как экономической к Интегрированные системы менеджмента в техносфере
Интегрированные системы менеджмента в техносфере ДОРОЖНЫЕ ЗНАКИ Воспитатель МБОУ «Кыновская СОШ №:65» Структурного подразделения «Детский сад ст. Кын» п. Кын, Лысьвенский район,
ДОРОЖНЫЕ ЗНАКИ Воспитатель МБОУ «Кыновская СОШ №:65» Структурного подразделения «Детский сад ст. Кын» п. Кын, Лысьвенский район,  Познание духовной природы мужчины и женщины - путь к гармонии
Познание духовной природы мужчины и женщины - путь к гармонии Магнийорганические соединения
Магнийорганические соединения  сметана
сметана Функции банка Подготовили: Шибанов Иван Титов Михаил
Функции банка Подготовили: Шибанов Иван Титов Михаил  Презентация на тему "Урок знаний 5 класс" - скачать презентации по Педагогике
Презентация на тему "Урок знаний 5 класс" - скачать презентации по Педагогике Счётчики. Лекция 4
Счётчики. Лекция 4 Истоки русского театра
Истоки русского театра VVED_Aминокислоты_Б19
VVED_Aминокислоты_Б19 Обучающий семинар для тьюторов проекта «Школьный стандарт профилактики» Ведущие: Екатерина Львовна Лашевская, начальник инфор
Обучающий семинар для тьюторов проекта «Школьный стандарт профилактики» Ведущие: Екатерина Львовна Лашевская, начальник инфор Свойства формовочных и стержневых смесей
Свойства формовочных и стержневых смесей Проф заболевания кожи (Тебенихин)
Проф заболевания кожи (Тебенихин) Паркур
Паркур Корпоративная культура (3). Семейная модель корпоративного управления
Корпоративная культура (3). Семейная модель корпоративного управления Тема 1. Финансы предприятий и финансовый менеджмент
Тема 1. Финансы предприятий и финансовый менеджмент  Частные вопросы гигиены труда
Частные вопросы гигиены труда  Трансформаторы. Трансформатор ОДЦЭ-5000/25Б
Трансформаторы. Трансформатор ОДЦЭ-5000/25Б