- Нуклеиновые кислоты. Строение и функции

Содержание
- 2. Функции нуклеиновых кислот Основная функция нуклеиновых кислот - хранение и передача наследственнойнаследственной информации, а также непосредствен-ное
- 3. Составные элементы ДНК и РНК Нуклеиновые кислоты построены из трех элементов: 1)гетероциклические азотистые основания; 2) пятичленный
- 4. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ Фридрих Мишер (1869) (Швейцария) Нуклеиновые основания Пиримидин Пурин рКв = 2,3 рКв= 1,3
- 5. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ Нуклеиновые основания (в лактамной форме) Пиримидиновые Урацил Ura (2,4-диоксопиримидин) Тимин Thy (5-метил-2,4-диоксопиримидин, 5-метилурацил
- 6. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ Нуклеиновые основания (в лактамной форме) Пуриновые Аденин Ade (6-аминопурин) Гуанин Gua (2-амино-6-оксопурин)
- 7. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ Нуклеиновые основания (в лактамной форме) Лактим-лактамная таутомерия
- 8. Лактам - лактимная таутомерия нуклеиновых оснований
- 9. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ Нуклеиновые основания (в лактамной форме) Плоское строение молекул пиримидина и пурина
- 10. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ R=OH β-D-рибофураноза R=H 2-Дезокси-β-D-рибофураноза
- 11. Углеводы в структуре нуклеиновых кислот (рибоза и дезоксирибоза) D- рибоза β- D-рибофураноза D-2-дезоксирибоза β- D-2-дезоксирибоза
- 12. НУКЛЕОЗИДЫ Общая структура нуклеозида R=OH Рибонуклеозид R=H Дезоксирибонуклеозид
- 13. НУКЛЕОЗИДЫ НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
- 14. НУКЛЕОЗИДЫ НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
- 15. НУКЛЕОЗИДЫ НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
- 16. НУКЛЕОЗИДЫ НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
- 17. НУКЛЕОТИДЫ
- 18. АТФ – аденозинтрифосфат- является нуклеотидом и представляет собой одну из наиболее важных биологически активных молекул, вовлеченных
- 19. АТФ АДФ + Н3РО4 АТФ является универсальным энергоносителем для всех биологических процессов, которые идут с затратой
- 20. ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ Циклофосфаты
- 21. Первооткрывателем цАМФ считается американский биохимик Эрл Сазерленд (1957 г.) . Нобелевская премия 1971 года за «за
- 22. В тканях животных и человека цАМФ и цГМФ осуществляют функции внутриклеточного посредника (вторичного мессенджера) в осуществлении
- 23. Механизм передачи информации в клетку ПРОТЕИНКИНАЗЫ
- 24. Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путем фосфори-лирования остатков аминокис-лот, имеющих гидроксильные
- 25. Механизм действия протеинкиназ Протеинкиназа АТФ -АДФ Протеинкиназа АТФ -АДФ Протеинкиназа АТФ -АДФ Серин Треонин Тироксин
- 26. СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ Первичная структура нуклеиновых кислот
- 27. Расшифровка структуры ДНК и РНК связана с именами американца Френсиса Крига,, англичанина Мориса Уилкинса и Джеймса
- 28. Фрагмент структуры ДНК
- 29. На одном конце полинуклеотидной цепи находится свободная фосфатная группа при 5/ атоме дезоксирибофу-ранозы, на другом –
- 31. Вторичная структура нуклеиновых кислот Образование водородных связей между парами нуклеиновых оснований (комплементарные пары оснований) тимин-аденин, цитозин-гуанин
- 35. Вторичная СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
- 36. Структура фермента ДНК-полимеразы
- 37. Четыре принципа строения молекул ДНК Нерегулярность чередования К регулярному сахарофосфатному оставу нерегулярно присоединены азотистые основания. Азотистые
- 38. «Это слишком красиво, чтобы быть правдой» - Френсис Криг –первооткрыватель структуры ДНК (соавторы Джеймс Уитсон и
- 39. Розалинд Франклин была в двух шагах от открытия структуры ДНК
- 40. Диаметр ДЕК в сотни раз меньше длины волны света (диаметр двойной спирали – 2 нм, длина
- 41. 1. Структура одной цепи может быть полностью и однозначно восстановлена по другой, за счет того, что
- 42. По сути ДНК- очень длинное наследственное письмо, в котором в строчку в определенном порядке записаны буквы
- 43. ДНК находится в хромосомах. Каждая хромосома представлена одной линейной молекулой ДНК. В самой большой хромосоме человека
- 44. БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ В форме ДНК хранится генетическая информация, затем она переписывается в форму мРНК.
- 45. БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ 3. Транспортная РНК (тРНК) переносит аминокислоты к рибосомам, где они соединяются пептидными
- 46. СТРУКТУРА РИБОСОМАЛЬНОЙ РНК C. Трехмерная структура рРНК малой субъединицы. Домены (разного цвета) образуют отдельные блоки укладки.
- 47. Транскрипция (переписывание), как и репликация (считывание), включает денатурацию - расплетание двойной цепочки ДНК и построение по
- 48. Вид рибосомы бактериальной клетки
- 49. Процесс перевода с языка нуклеотидов на язык аминокислот называется трансляцией и происходит в рибосомах. Рибосома –,
- 50. Между матричной РНК и синтезируемым белком существуют посредники – транспортные РНК, или тРНК. Они транспортируют аминокислоты
- 51. Флавиннуклеотиды
- 52. Флавиннуклеотиды
- 53. Никотинамиднуклеотиды Энантиотропные атомы Hа (про-R) и Нб (про-S) в молекуле НАДН
- 54. Последовательность из трех букв (нуклеиновых оснований) называется триплетом, поэтому генетический код является триплетным. .
- 55. Генетический код –вырожденный, поскольку одна аминокислота может быть закодирована несколькими триплетами
- 56. Френсис Криг - первооткрыватель структуры ДНК- 1953 г. Нобелевские премии по физиологии и медицине присуждена в
- 57. Гипотеза мутаций и теория химического канцерогенеза
- 58. Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей среды
- 59. Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей среды
- 61. Производные пиримидина и пурина Тимин, урацил, цитозин – производные пиримидина разрушаются в организме с образованием низкомолекулярных
- 62. Превращение пуринов. Мочевая кислота
- 63. Функции мочевой кислоты Является мощным стимулятором центральной нервной системы, ингибируя фосфодиэстеразу, которая служит посредником действия гормонов
- 64. Антиметаболиты Антиметаболиты в химическом отношении напоминают естественные метаболиты, но не идентичны им; тем самым они нарушают
- 65. Формулы антагонистов нуклеиновых оснований 6-меркапто пурина и 5-фторурацила как пример молекулярной модификации с целью получения соединений,
- 67. Скачать презентацию

Функции нуклеиновых кислот
Основная функция нуклеиновых кислот - хранение и передача наследственнойнаследственной
Функции нуклеиновых кислот
Основная функция нуклеиновых кислот - хранение и передача наследственнойнаследственной

Составные элементы ДНК и РНК
Нуклеиновые кислоты построены из трех элементов:
1)гетероциклические
Составные элементы ДНК и РНК
Нуклеиновые кислоты построены из трех элементов:
1)гетероциклические
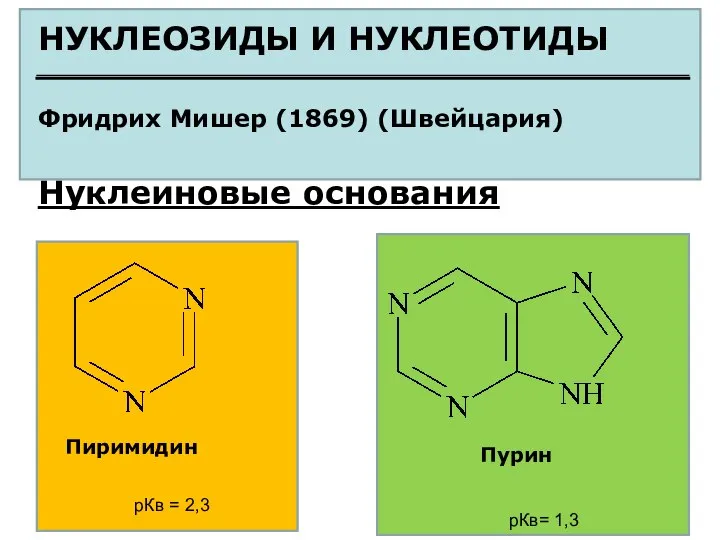
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Фридрих Мишер (1869) (Швейцария)
Нуклеиновые основания
Пиримидин
Пурин
рКв =
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Фридрих Мишер (1869) (Швейцария)
Нуклеиновые основания
Пиримидин
Пурин
рКв =
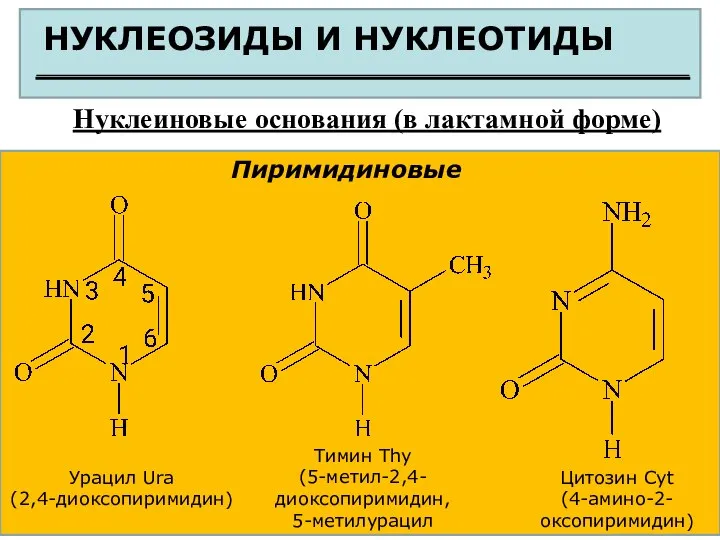
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Пиримидиновые
Урацил Ura
(2,4-диоксопиримидин)
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Пиримидиновые
Урацил Ura
(2,4-диоксопиримидин)
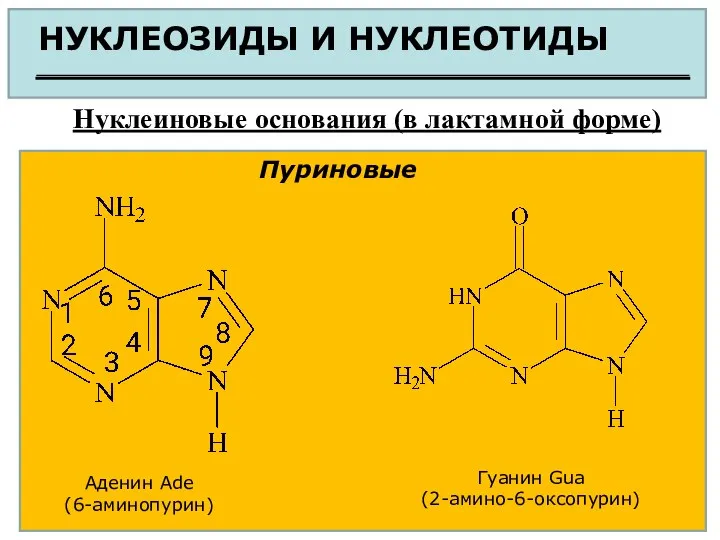
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Пуриновые
Аденин Ade
(6-аминопурин)
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Пуриновые
Аденин Ade
(6-аминопурин)
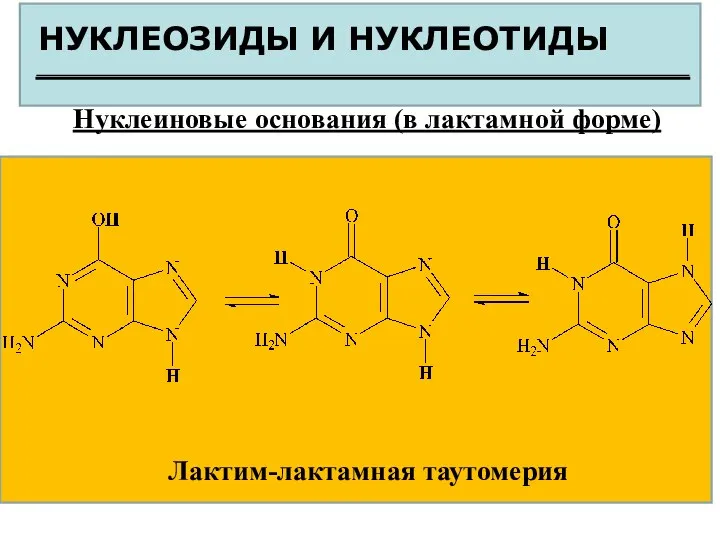
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Лактим-лактамная таутомерия
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Лактим-лактамная таутомерия
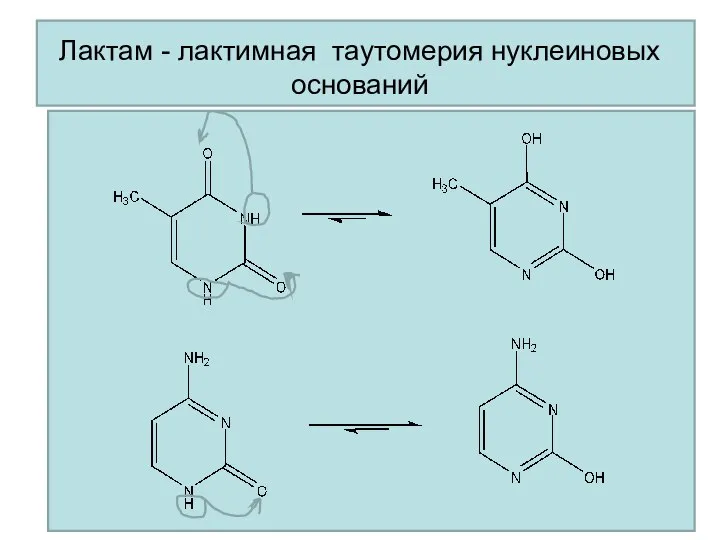
Лактам - лактимная таутомерия нуклеиновых оснований
Лактам - лактимная таутомерия нуклеиновых оснований
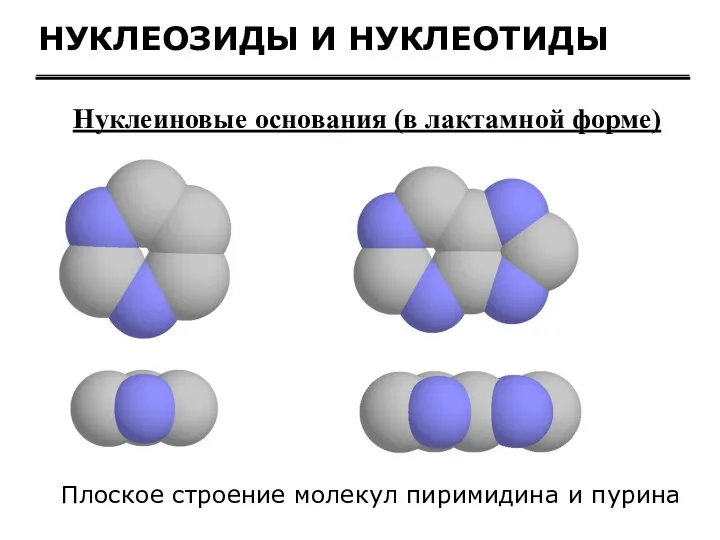
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Плоское строение молекул
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеиновые основания (в лактамной форме)
Плоское строение молекул
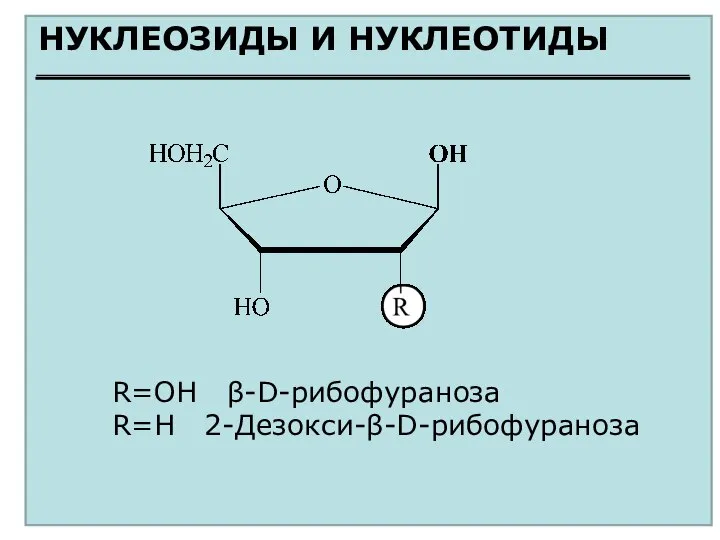
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
R=OH β-D-рибофураноза
R=H 2-Дезокси-β-D-рибофураноза
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
R=OH β-D-рибофураноза
R=H 2-Дезокси-β-D-рибофураноза
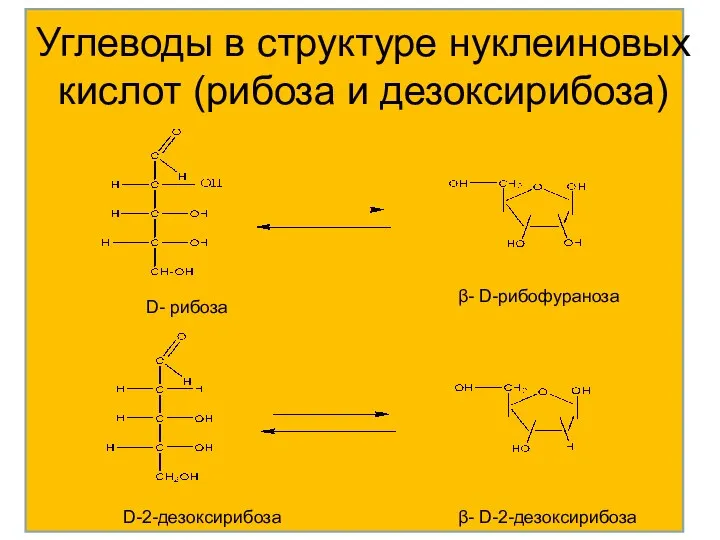
Углеводы в структуре нуклеиновых кислот (рибоза и дезоксирибоза)
D- рибоза
β- D-рибофураноза
D-2-дезоксирибоза
β- D-2-дезоксирибоза
Углеводы в структуре нуклеиновых кислот (рибоза и дезоксирибоза)
D- рибоза
β- D-рибофураноза
D-2-дезоксирибоза
β- D-2-дезоксирибоза
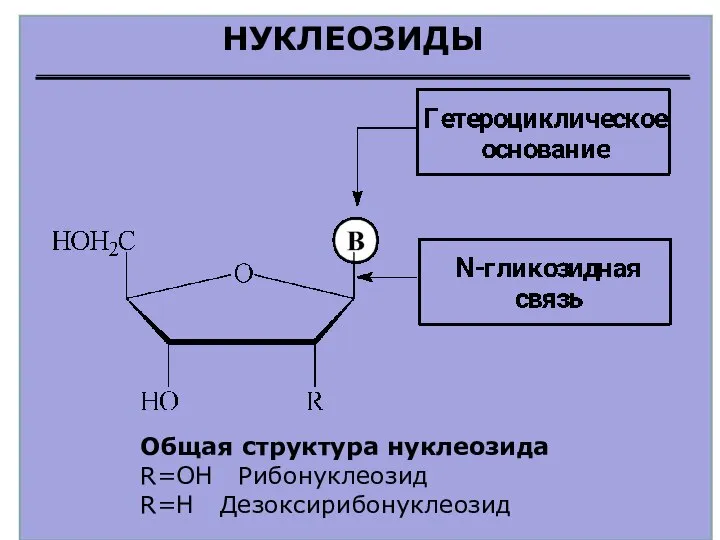
НУКЛЕОЗИДЫ
Общая структура нуклеозида
R=OH Рибонуклеозид
R=H Дезоксирибонуклеозид
НУКЛЕОЗИДЫ
Общая структура нуклеозида
R=OH Рибонуклеозид
R=H Дезоксирибонуклеозид
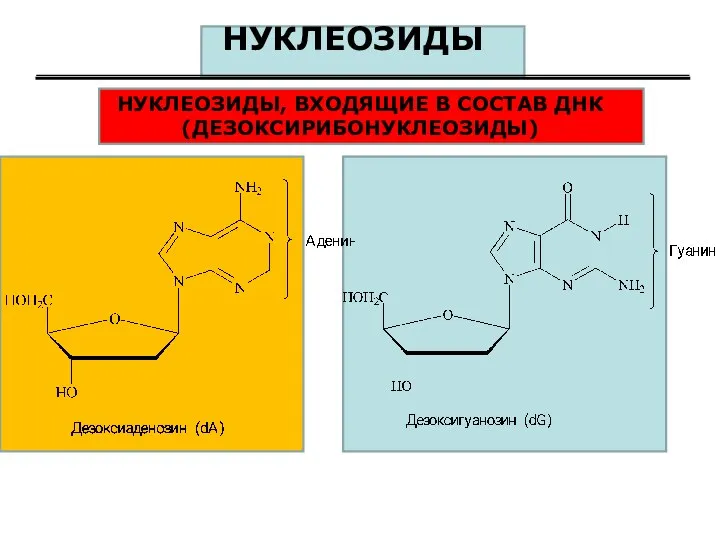
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
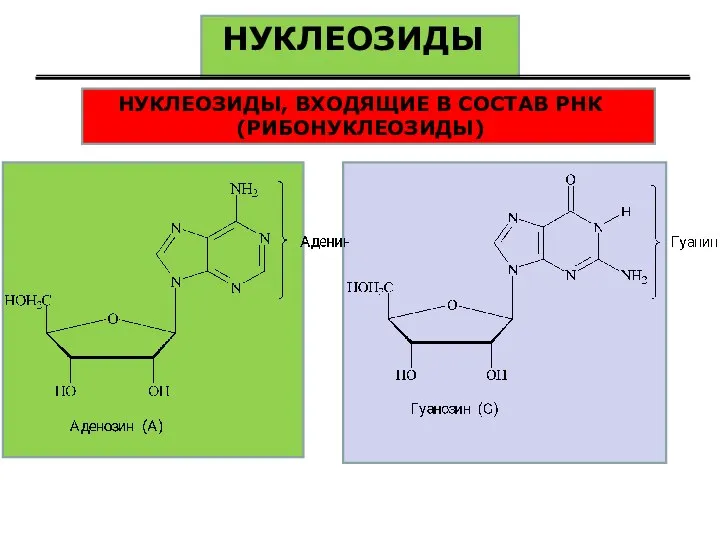
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
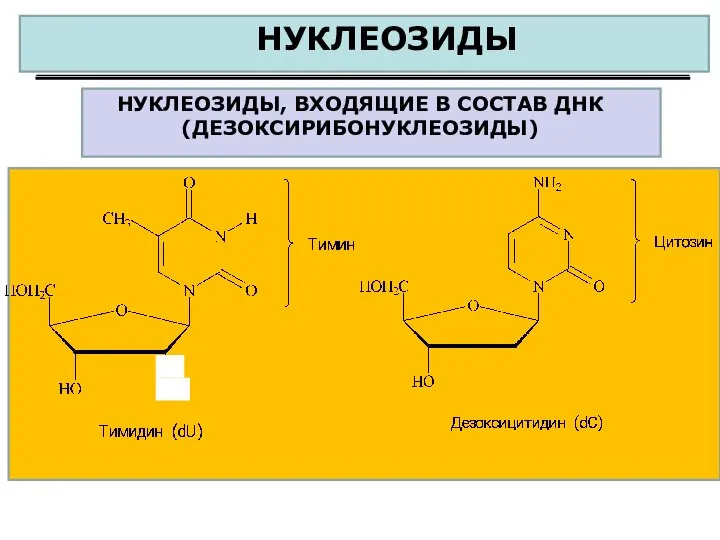
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ ДНК (ДЕЗОКСИРИБОНУКЛЕОЗИДЫ)
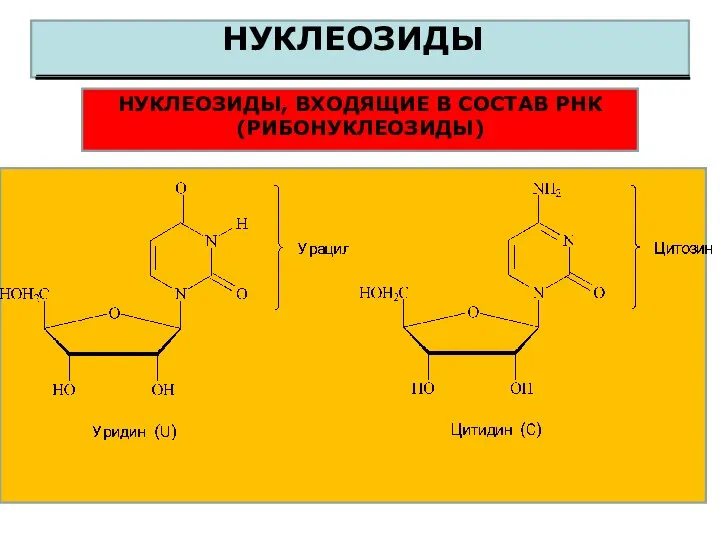
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
НУКЛЕОЗИДЫ
НУКЛЕОЗИДЫ, ВХОДЯЩИЕ В СОСТАВ РНК (РИБОНУКЛЕОЗИДЫ)
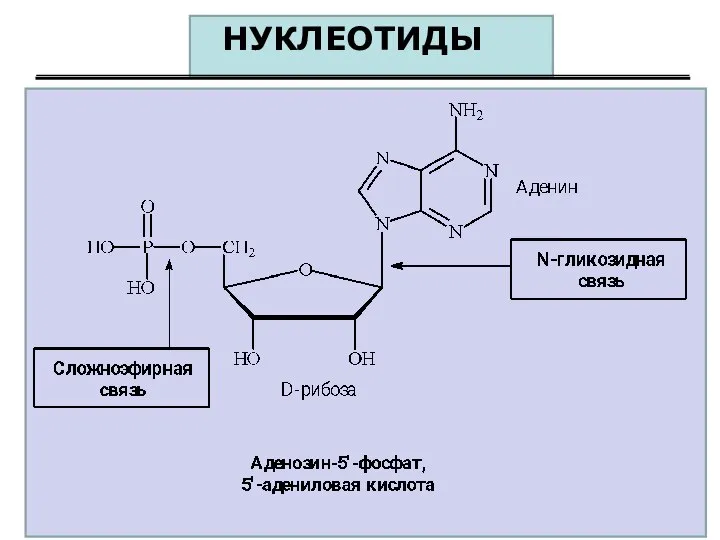
НУКЛЕОТИДЫ
НУКЛЕОТИДЫ
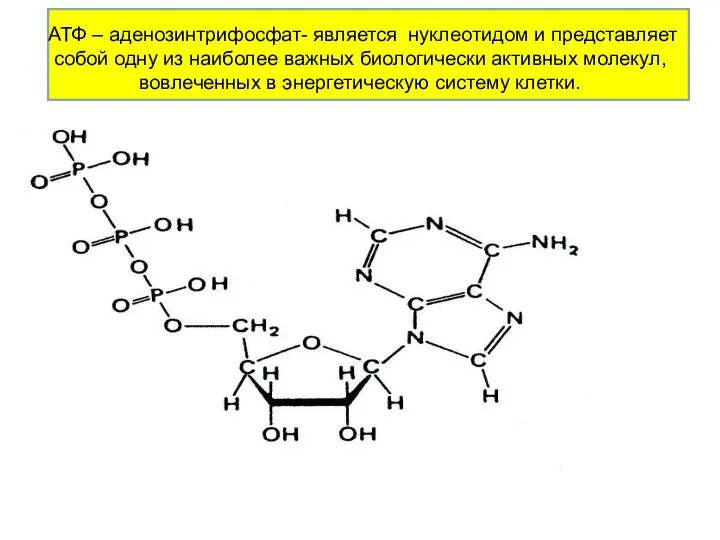
АТФ – аденозинтрифосфат- является нуклеотидом и представляет собой одну из наиболее
АТФ – аденозинтрифосфат- является нуклеотидом и представляет собой одну из наиболее

АТФ АДФ + Н3РО4
АТФ является универсальным энергоносителем для всех биологических процессов,
АТФ АДФ + Н3РО4
АТФ является универсальным энергоносителем для всех биологических процессов,
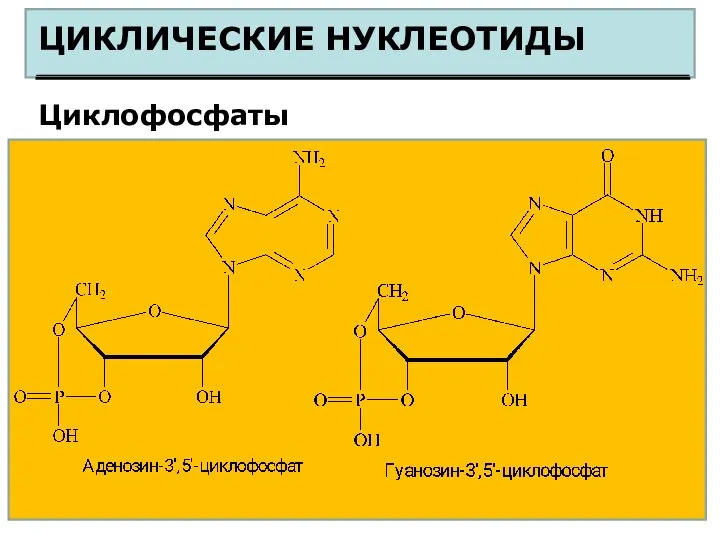
ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ
Циклофосфаты
ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ
Циклофосфаты

Первооткрывателем цАМФ считается американский биохимик Эрл Сазерленд (1957 г.) . Нобелевская
Первооткрывателем цАМФ считается американский биохимик Эрл Сазерленд (1957 г.) . Нобелевская

В тканях животных и человека цАМФ и цГМФ осуществляют функции внутриклеточного
В тканях животных и человека цАМФ и цГМФ осуществляют функции внутриклеточного
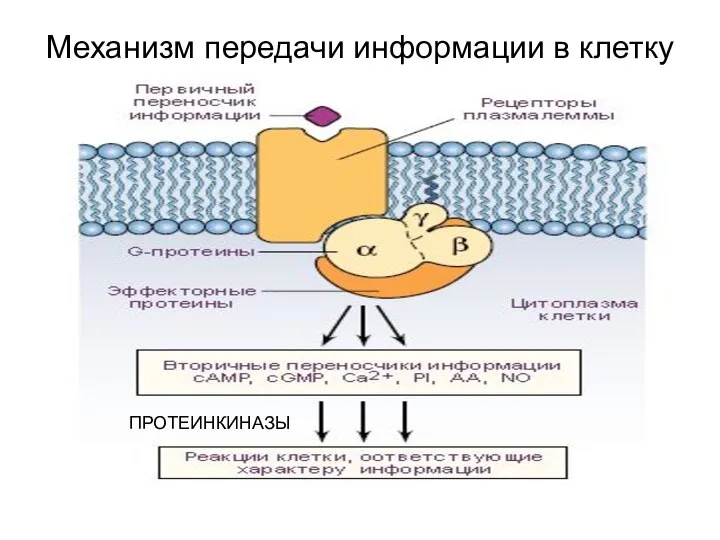
Механизм передачи информации в клетку
ПРОТЕИНКИНАЗЫ
Механизм передачи информации в клетку
ПРОТЕИНКИНАЗЫ

Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз).
Протеинкиназы модифицируют другие белки путем
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз).
Протеинкиназы модифицируют другие белки путем

Механизм действия протеинкиназ
Протеинкиназа
АТФ
-АДФ
Протеинкиназа
АТФ
-АДФ
Протеинкиназа
АТФ
Механизм действия протеинкиназ
Протеинкиназа
АТФ
-АДФ
Протеинкиназа
АТФ
-АДФ
Протеинкиназа
АТФ
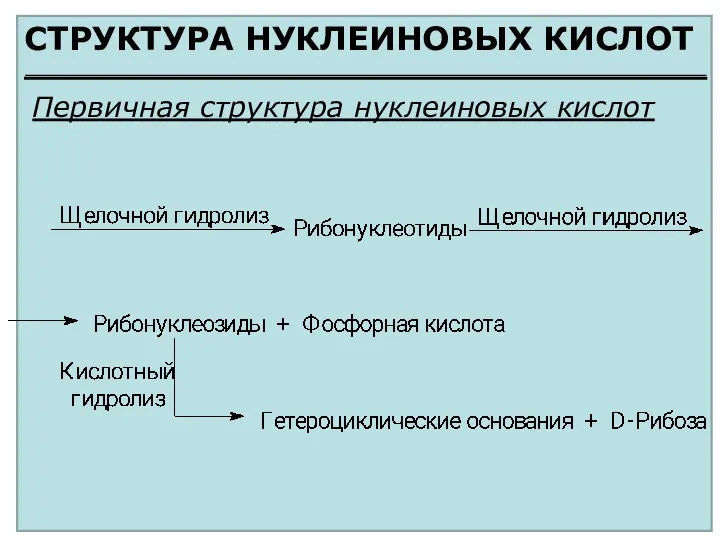
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Первичная структура нуклеиновых кислот
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Первичная структура нуклеиновых кислот

Расшифровка структуры ДНК и РНК
связана с именами американца Френсиса Крига,,
Расшифровка структуры ДНК и РНК
связана с именами американца Френсиса Крига,,
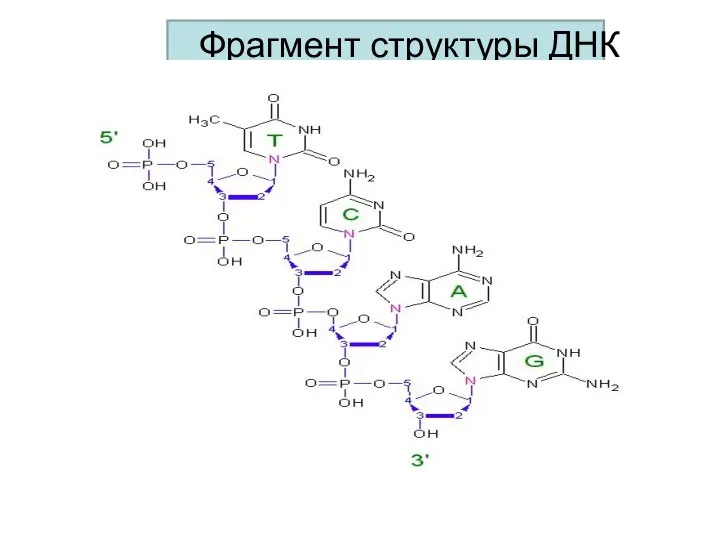
Фрагмент структуры ДНК
Фрагмент структуры ДНК

На одном конце полинуклеотидной цепи находится свободная фосфатная группа при 5/
На одном конце полинуклеотидной цепи находится свободная фосфатная группа при 5/
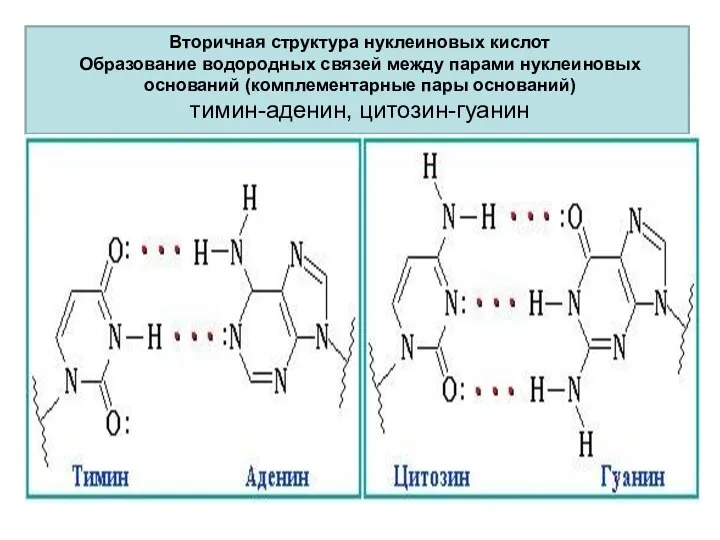
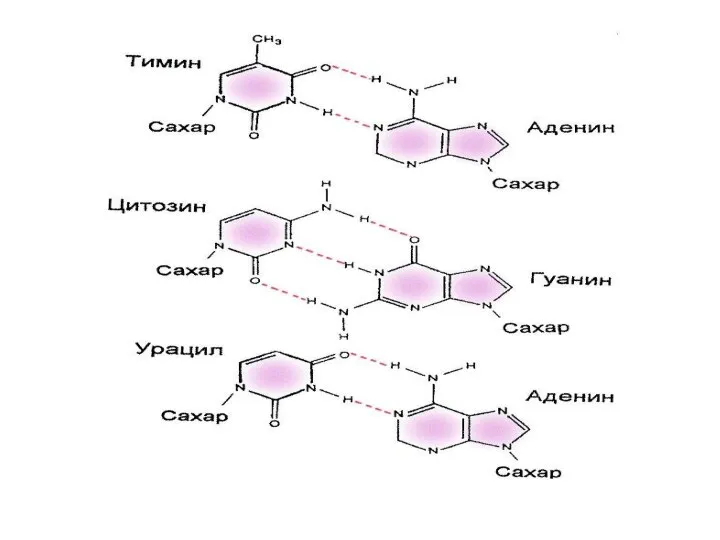
Вторичная структура нуклеиновых кислот
Образование водородных связей между парами нуклеиновых оснований (комплементарные
Вторичная структура нуклеиновых кислот Образование водородных связей между парами нуклеиновых оснований (комплементарные
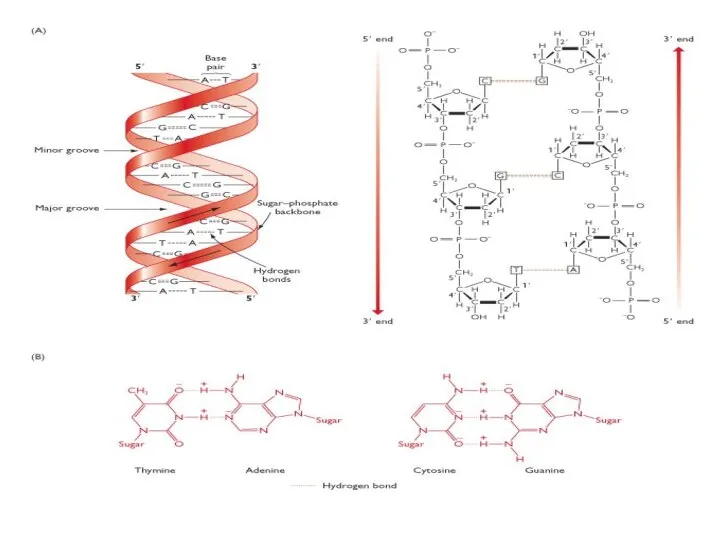
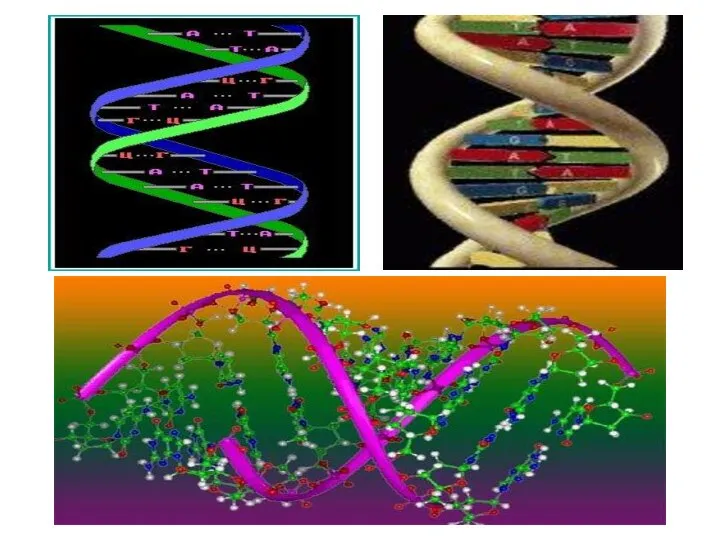
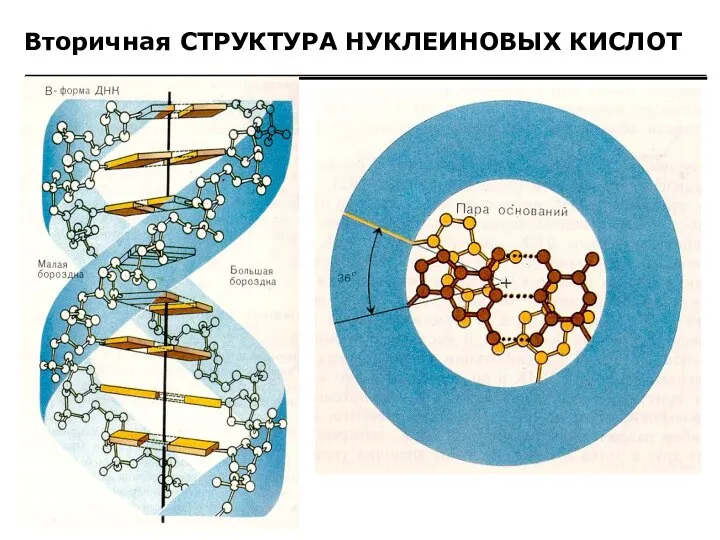
Вторичная СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Вторичная СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
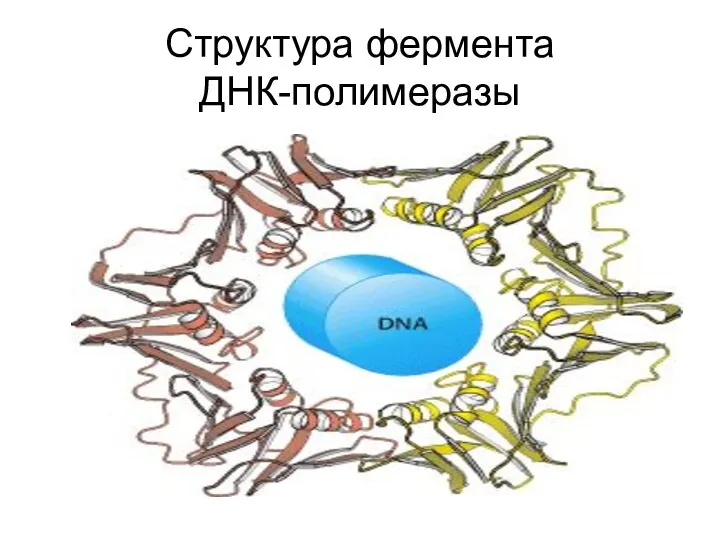
Структура фермента
ДНК-полимеразы
Структура фермента
ДНК-полимеразы

Четыре принципа строения молекул ДНК
Нерегулярность чередования
К регулярному сахарофосфатному оставу нерегулярно
Четыре принципа строения молекул ДНК
Нерегулярность чередования
К регулярному сахарофосфатному оставу нерегулярно

«Это слишком красиво, чтобы быть правдой» - Френсис Криг –первооткрыватель структуры
«Это слишком красиво, чтобы быть правдой» - Френсис Криг –первооткрыватель структуры

Розалинд Франклин была в двух шагах от открытия структуры ДНК
Розалинд Франклин была в двух шагах от открытия структуры ДНК
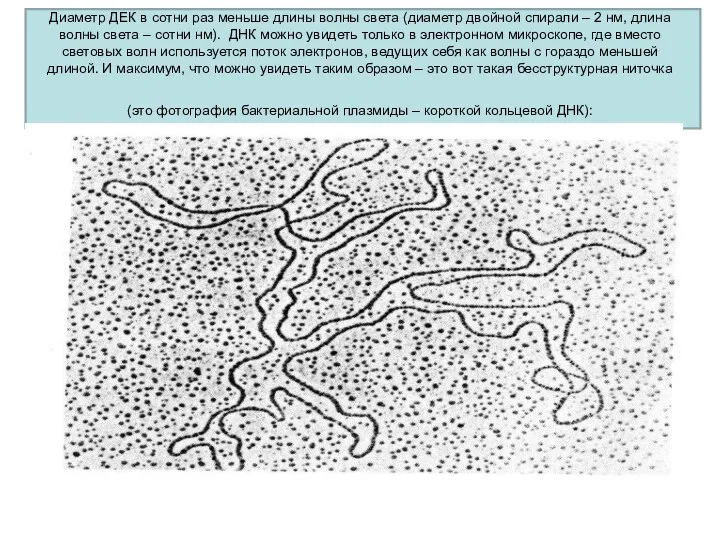
Диаметр ДЕК в сотни раз меньше длины волны света (диаметр двойной
Диаметр ДЕК в сотни раз меньше длины волны света (диаметр двойной
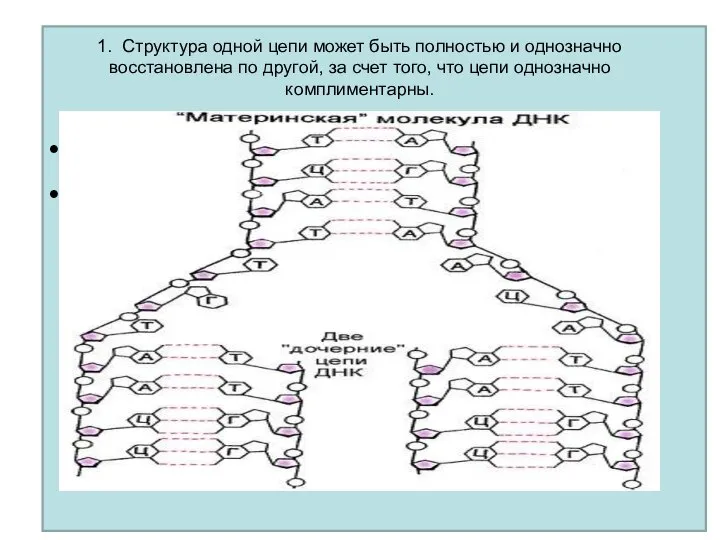
1. Структура одной цепи может быть полностью и однозначно восстановлена по
1. Структура одной цепи может быть полностью и однозначно восстановлена по

По сути ДНК- очень длинное наследственное письмо, в котором в строчку
По сути ДНК- очень длинное наследственное письмо, в котором в строчку

ДНК находится в хромосомах. Каждая хромосома представлена одной линейной молекулой
ДНК находится в хромосомах. Каждая хромосома представлена одной линейной молекулой

БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ
В форме ДНК хранится генетическая информация, затем она
БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ
В форме ДНК хранится генетическая информация, затем она

БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ
3. Транспортная РНК (тРНК) переносит аминокислоты к рибосомам,
БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ
3. Транспортная РНК (тРНК) переносит аминокислоты к рибосомам,
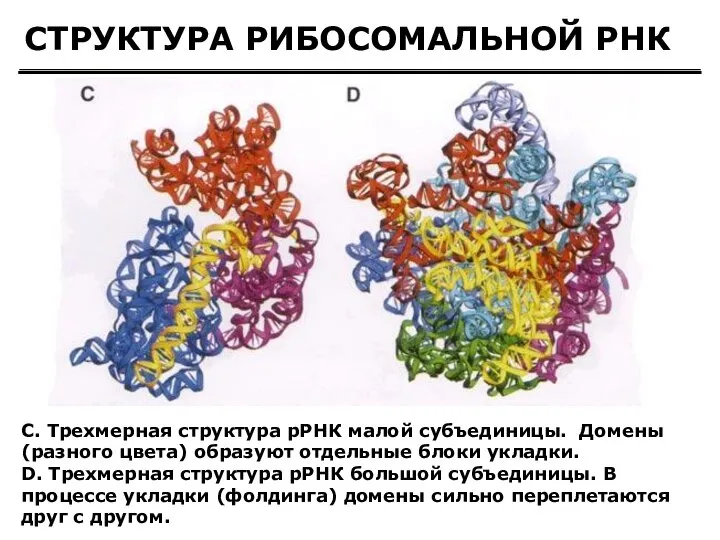
СТРУКТУРА РИБОСОМАЛЬНОЙ РНК
C. Трехмерная структура рРНК малой субъединицы. Домены (разного
СТРУКТУРА РИБОСОМАЛЬНОЙ РНК
C. Трехмерная структура рРНК малой субъединицы. Домены (разного
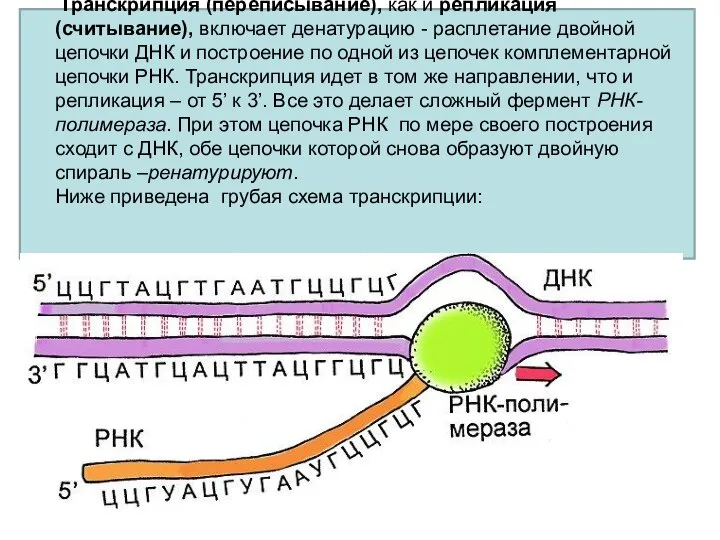
Транскрипция (переписывание), как и репликация (считывание), включает денатурацию - расплетание двойной
Транскрипция (переписывание), как и репликация (считывание), включает денатурацию - расплетание двойной
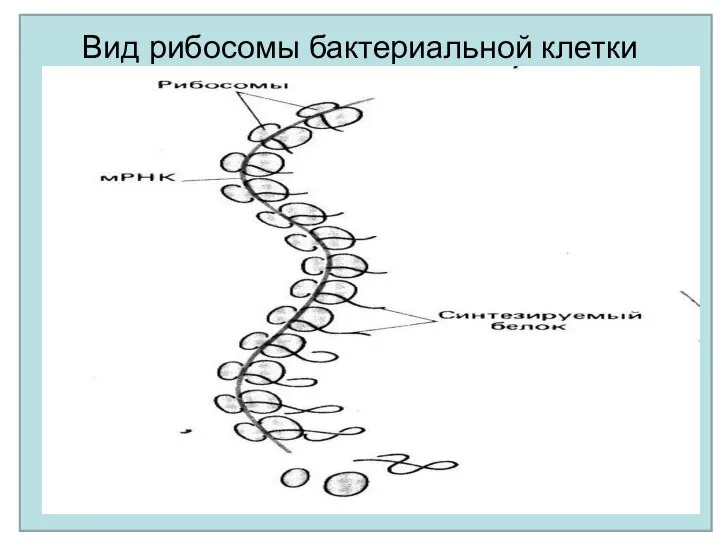
Вид рибосомы бактериальной клетки
Вид рибосомы бактериальной клетки

Процесс перевода с языка нуклеотидов на язык аминокислот называется трансляцией и
Процесс перевода с языка нуклеотидов на язык аминокислот называется трансляцией и
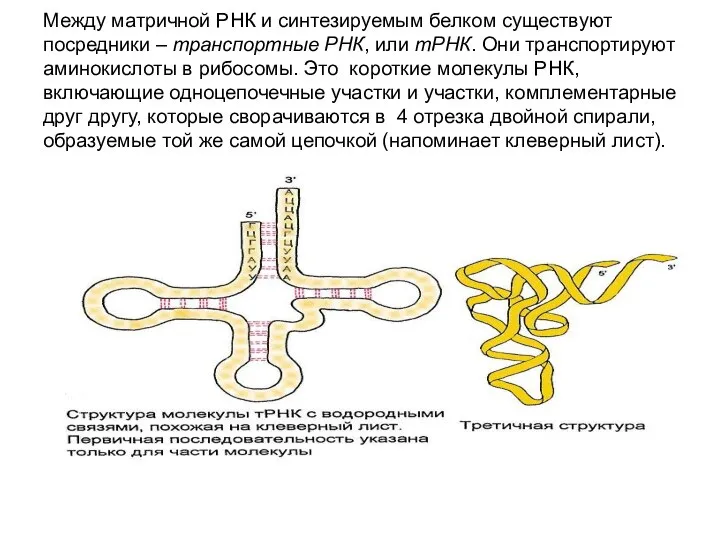
Между матричной РНК и синтезируемым белком существуют посредники – транспортные РНК,
Между матричной РНК и синтезируемым белком существуют посредники – транспортные РНК,
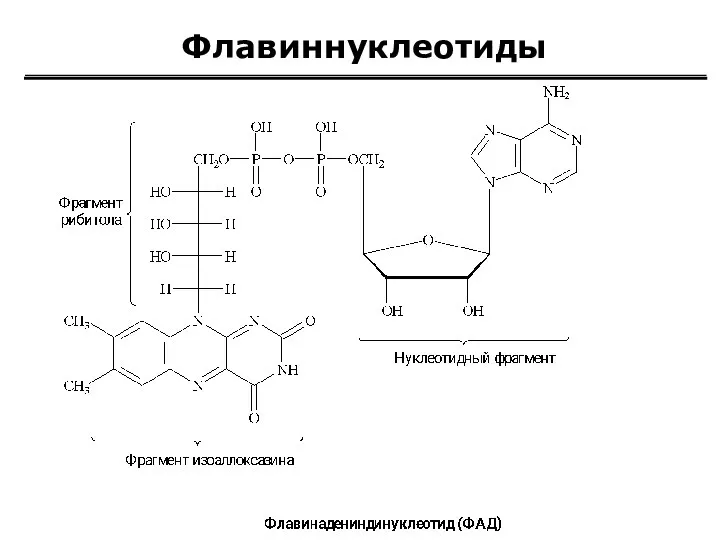
Флавиннуклеотиды
Флавиннуклеотиды
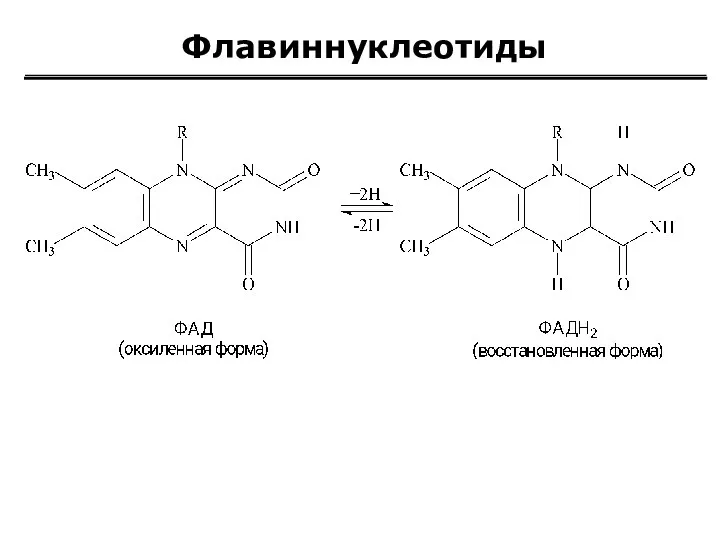
Флавиннуклеотиды
Флавиннуклеотиды
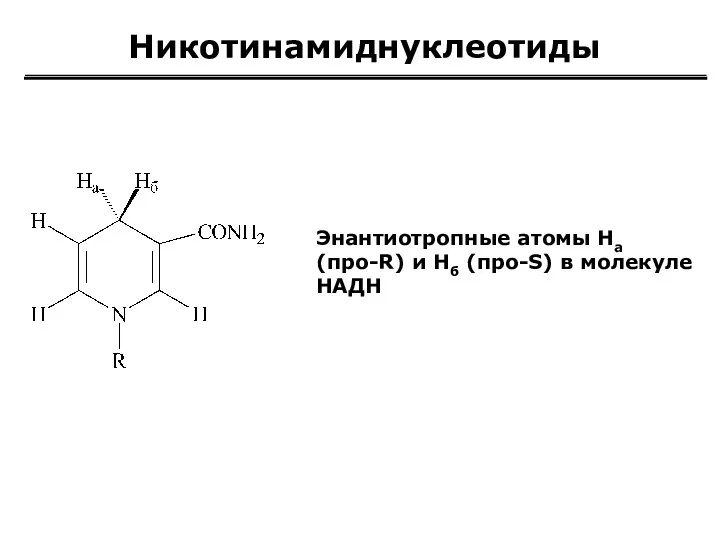
Никотинамиднуклеотиды
Энантиотропные атомы Hа
(про-R) и Нб (про-S) в молекуле НАДН
Никотинамиднуклеотиды
Энантиотропные атомы Hа
(про-R) и Нб (про-S) в молекуле НАДН
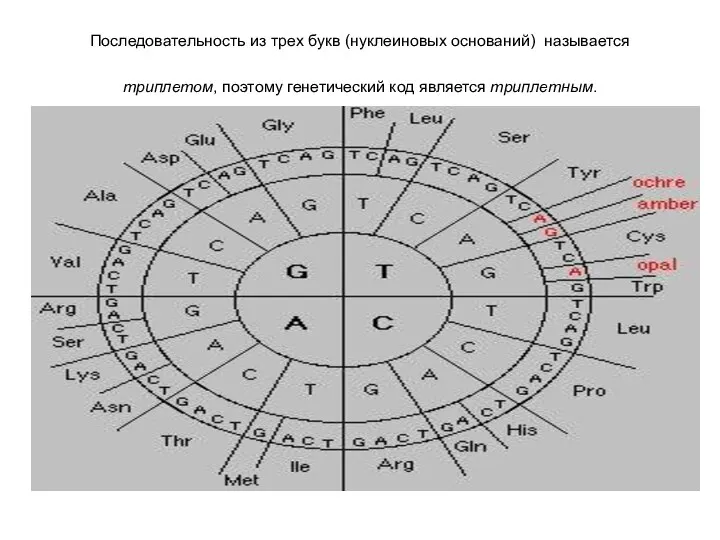
Последовательность из трех букв (нуклеиновых оснований) называется триплетом, поэтому генетический код
Последовательность из трех букв (нуклеиновых оснований) называется триплетом, поэтому генетический код
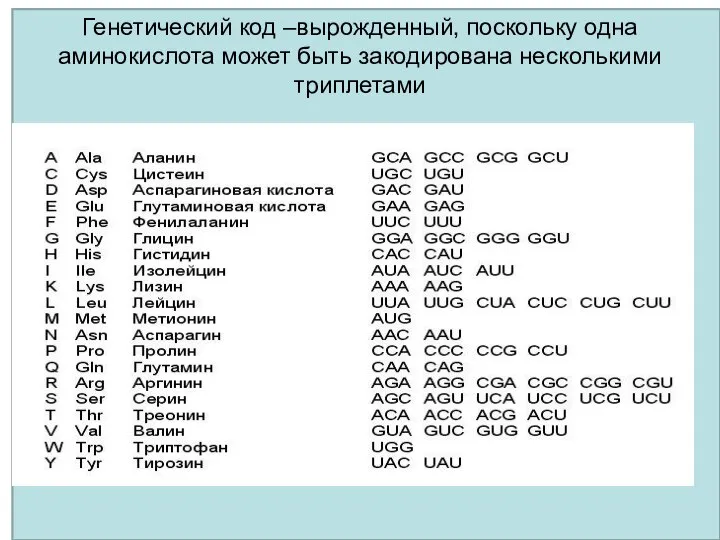
Генетический код –вырожденный, поскольку одна аминокислота может быть закодирована несколькими триплетами
Генетический код –вырожденный, поскольку одна аминокислота может быть закодирована несколькими триплетами

Френсис Криг - первооткрыватель структуры ДНК- 1953 г. Нобелевские премии по
Френсис Криг - первооткрыватель структуры ДНК- 1953 г. Нобелевские премии по

Гипотеза мутаций и
теория химического канцерогенеза
Гипотеза мутаций и
теория химического канцерогенеза
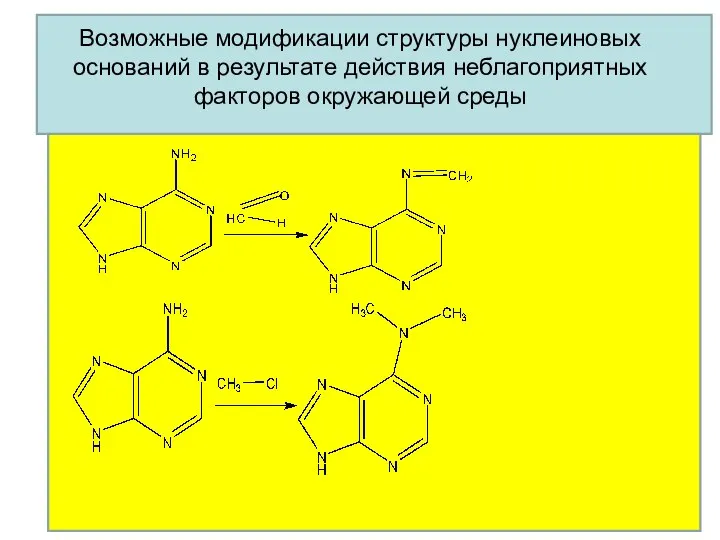
Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей
Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей
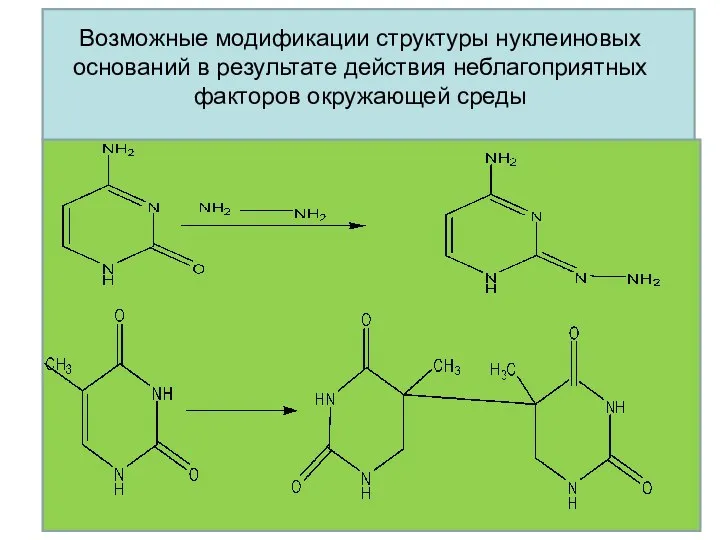
Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей
Возможные модификации структуры нуклеиновых оснований в результате действия неблагоприятных факторов окружающей
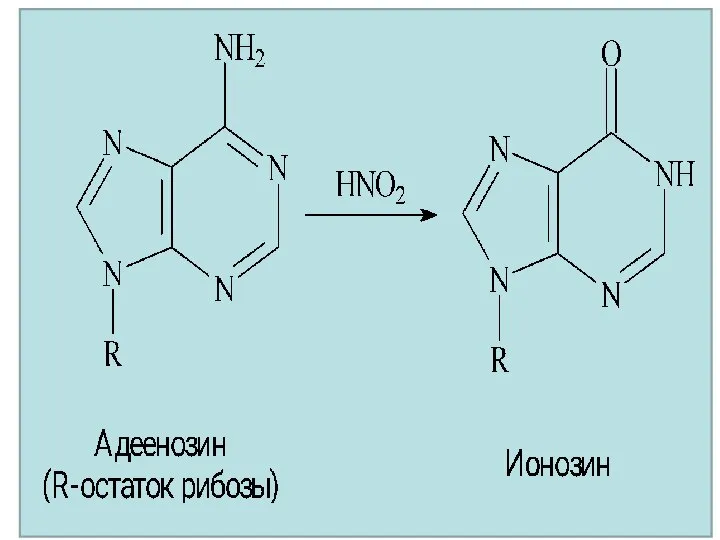

Производные пиримидина и пурина
Тимин, урацил, цитозин – производные пиримидина разрушаются в
Производные пиримидина и пурина
Тимин, урацил, цитозин – производные пиримидина разрушаются в
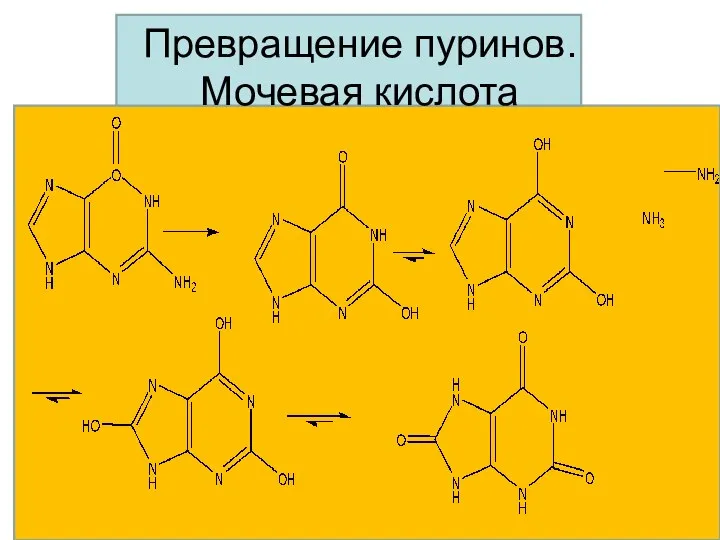
Превращение пуринов.
Мочевая кислота
Превращение пуринов.
Мочевая кислота

Функции мочевой кислоты
Является мощным стимулятором центральной нервной системы, ингибируя фосфодиэстеразу, которая
Функции мочевой кислоты
Является мощным стимулятором центральной нервной системы, ингибируя фосфодиэстеразу, которая

Антиметаболиты
Антиметаболиты в химическом отношении напоминают естественные метаболиты, но не идентичны
Антиметаболиты
Антиметаболиты в химическом отношении напоминают естественные метаболиты, но не идентичны
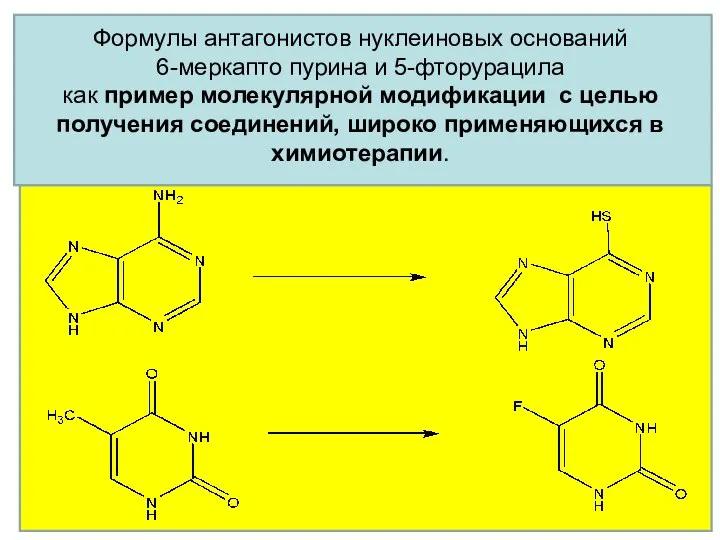
Формулы антагонистов нуклеиновых оснований
6-меркапто пурина и 5-фторурацила
как пример молекулярной модификации
Формулы антагонистов нуклеиновых оснований 6-меркапто пурина и 5-фторурацила как пример молекулярной модификации
 Прогнозирование остаточного ресурса объекта
Прогнозирование остаточного ресурса объекта Динамические и статические страницы. Лекция 1
Динамические и статические страницы. Лекция 1 Hardware printer
Hardware printer Презентация Пути совершенствования мотивации трудовой деятельности в таможенных органах
Презентация Пути совершенствования мотивации трудовой деятельности в таможенных органах Зимушка -зима
Зимушка -зима  Interesting facts about Scotland
Interesting facts about Scotland Сокращение алгебраических дробей - презентация по Алгебре
Сокращение алгебраических дробей - презентация по Алгебре Акробатические упражнения по «Комплексной программе физического воспитания для учащихся 1-11 классов». Методика обучения
Акробатические упражнения по «Комплексной программе физического воспитания для учащихся 1-11 классов». Методика обучения RedeX Plus - Your New Pace of Life
RedeX Plus - Your New Pace of Life Вормсский махзор
Вормсский махзор ПОДГОТОВИЛА ПРЕПОДАВАТЕЛЬ ПМ 04,05 АЗАРЦОВА А.М. ПОТРЕБНОСТИ ПАЦИЕНТА В СНЕ И ОТДЫХЕ
ПОДГОТОВИЛА ПРЕПОДАВАТЕЛЬ ПМ 04,05 АЗАРЦОВА А.М. ПОТРЕБНОСТИ ПАЦИЕНТА В СНЕ И ОТДЫХЕ Анализ установившихся и переходных режимов в системе электроснабжения 220/110 кВ
Анализ установившихся и переходных режимов в системе электроснабжения 220/110 кВ Основные источники современного российского конституционного права, их соотношение с источниками налогового права. Тема 2
Основные источники современного российского конституционного права, их соотношение с источниками налогового права. Тема 2 Частотно-керовані електроприводи змінного струму
Частотно-керовані електроприводи змінного струму Искусственные нейронные сети. Лекция 15-16
Искусственные нейронные сети. Лекция 15-16 Информационные технологии в электроэнергетике. SCADA-системы (Часть 3)
Информационные технологии в электроэнергетике. SCADA-системы (Часть 3) Технологическая карта урока как основа проектирования урока Если мы хотим достигнуть какой-нибудь цели воспитанием, то должны п
Технологическая карта урока как основа проектирования урока Если мы хотим достигнуть какой-нибудь цели воспитанием, то должны п Презентация на тему: Инжиниринг Выполнил: ученик 11а класса Журавлев Данил
Презентация на тему: Инжиниринг Выполнил: ученик 11а класса Журавлев Данил  Анализ видов, последствий и причин потенциальных несоответствий
Анализ видов, последствий и причин потенциальных несоответствий Задачи с использованием циклов
Задачи с использованием циклов Режимы работы электродвигателей
Режимы работы электродвигателей Схемы отвода конденсата греющего пара в регенеративных подогревателях поверхностного типа. Лекция 4
Схемы отвода конденсата греющего пара в регенеративных подогревателях поверхностного типа. Лекция 4 Химическая термодинамика: теоретическая основа биоэнергетики
Химическая термодинамика: теоретическая основа биоэнергетики Физиотерапевтические средства: дозирование, ошибки назначения, рецептура к.м.н., с.н.с. Жук Д.Д.
Физиотерапевтические средства: дозирование, ошибки назначения, рецептура к.м.н., с.н.с. Жук Д.Д.  Аналитик. Бизнес-аналитик
Аналитик. Бизнес-аналитик Актуальные проблемы физического воспитания в современных условиях образования
Актуальные проблемы физического воспитания в современных условиях образования Новелла первая Петрарка и Лаура
Новелла первая Петрарка и Лаура Non-State. Definition
Non-State. Definition