- Популяционно – статистический метод изучения генетики человека

Содержание
- 2. Методы изучения генетики человека Клинико – генеалогический Близнецовый Популяционно – статистический Цитогенетический Метод генетики соматических клеток
- 3. Определение популяции (научное эволюционно-генетическое, Н.В. Тимофеева - Ресовским) Это совокупность особей определенного вида, обладающих общим генофондом
- 4. Определение популяции (продолжение) внутри которого практически осуществляется та или иная степень панмиксии (случайного, не зависящего от
- 5. Определение популяции (окончание). эта совокупность особей отделена от соседних таких же совокупностей особей данного вида той
- 6. Задачи популяционно -статистического метода: 1. Изучить генетическую структуру популяций, их генофонд, взаимодействие факторов, обусловливающих постоянство и
- 7. Задачи популяционно -статистического метода: 2. Изучить наследственные болезни населения, частоту нормальных и патологических генов, генотипов, фенотипов
- 8. Задачи популяционно -статистического метода: 3. Изучает закономерности распространения наследственных болезней в разных по строению популяциях и
- 9. Задачи популяционно -статистического метода: 4. Изучает частоты генов в популя-ции, включая наследственные болезни 5. Изучает закономерности
- 10. Задачи популяционно -статистического метода: 6.Роль наследственности и среды в возникновении болезней с наслед-ственной предрасположенностью (эпидемические заболевания,
- 11. Задачи популяционно -статистического метода: 7. Влияния наследственных и средовых факторов в создании фенотипического полиморфизма человека по
- 12. Значение популяционно -статистического метода Дает возможность определить генетическую структуру популяции, рассчитать в популяции человека частоту нормальных
- 13. Значение популяционно -статистического метода Изучает генетическую структуру популяций, их генофонд, взаимодействие факторов, обусловливающих постоянство и изменение
- 14. Табл. Наследуемость некоторых признаков человека, определенная близнецовым методом
- 15. Популяционно-статистический метод включает: 1) правильный выбор популяции, 2) сбор материала и 3) статистический анализ полученных результатов.
- 16. Основа популяционно- статистического метода В основе метода лежит закономерность, установленная в 1908 г. английским математиком Дж.
- 17. Черты идеальной популяции: большая численность, свободное скрещивание (панмиксия) организмов, отсутствие отбора и мутационного процесса, отсутствие миграций
- 18. Основной смысл закона Харди - Вайнберга В идеальной популяции соотношение частоты доминантных гомозигот (АД), гетерозигот (Аа)
- 19. Факторы, стимулирующие сдвиг равновесия в популяции: родственные браки, мутации, дрейф генов, отбор, миграции и другие.
- 20. Значение закона Харди – Вайнберга. Является основой при рассмотрении генетических преобразований, происходящих в естественных и искусственно
- 21. Соотношение численности разных генотипов и фенотипов в панмиктической популяции Определяется по формуле бинома Ньютона: (p +q)
- 22. Обозначения в формуле бинома Ньютона р — частота доминантного аллеля А, q — частота рецессивного аллеля
- 23. Пояснение закона Харди Вейнберга Частота доминантных гомозигот (АА) равна квадрату вероятности встречае-мости доминантного аллеля, частота гетерозигот
- 24. Пример наследования Rh - фактора В одном из городов 16 % людей оказались резус-отрицательными и 84
- 25. Разбор задачи: Носители резус-положительного фактора могут иметь генотип СС или Сс. Чтобы определить, какая часть из
- 26. Расчет задачи: Гомозиготы по рецессивному аллелю составляют 16% или 0,16; отсюда q = Ѵ16 = 0,40
- 27. Расчет задачи: Процент в популяции зигот СС и Сс вычисляем следующим образом: СС = р2 =
- 28. Заключение по задаче: Следовательно, среди обследованного населения положительный резус-фактор имели 36 % с генотипом СС и
- 29. Особенности закона Харди – Вайнберга: 1. Описывает популяции в состоянии покоя. В этом он аналогичен первому
- 30. Особенности закона Харди – Вайнберга: 2. Закон Харди – Вайнберга гласит: при отсутствии возмущающих процессов частоты
- 31. Особенности закона Харди – Вайнберга: 3. Задает точку отсчета, по отношению к которой анализируются изменения, вызванные
- 32. Особенности закона Харди – Вайнберга: 4. В малочисленных популяциях человека закон Харди - Вайнберга не применим,
- 33. Эволюционные процессы в генетике человека А) мутации Б) миграции В) дрейф генов Г) естественный отбор
- 34. Определение мутации. Мутации (изменения)- источники генетической изменчивости, но частота крайне низка. Это процесс чрезвычайно медленный, поэтому
- 35. Пример расчета мутаций 1 Существуют два аллеля одного локуса (т. е. два варианта одного гена) —
- 36. Пример расчета мутаций2 Допустим также, что в начальный момент времени (до начала процесса мутации) частота аллеля
- 37. Пример расчета мутаций3 Во втором поколении доля V оставшихся аллелей а (частота встречаемости которых в популяции
- 38. Пример расчета мутаций4 По прошествии t поколений частота аллеля а будет равна: р0 ( 1 -
- 39. Пример расчета мутаций5 Однако все определяется величиной V. В естественных условиях она чрезвычайно мала и составляет
- 40. Пример расчета мутаций6 В таком темпе, для того чтобы изменить частоту аллеля а от 1 до
- 41. Значение мутаций в генетике Для того чтобы мутации сами по себе привели к сколько-нибудь значительному изменению
- 42. Виды мутаций по измеению генотипа Генные (или точковые), Внутрихромосомные, Межхромосомные, Геномные (изменение числа хромосом.).
- 43. Виды генных мутаций Прямые (А>а) и обратные (а>А). Частота возникновения прямых му-таций значительно выше обратных. Одни
- 44. Частота мутаций, обусловливающих тяжелые болезни Частота возникновения патологи-ческих мутаций отдельного гена (гемофилия, ретинобластома, пигментная ксеродерма и
- 45. Чем обусловлен рост мутаций? Химические мутагены : (канцерогены) - промышленные яды, инсектициды, гербициды, пищевые добавки и
- 46. Чем обусловлен рост мутаций? Сильные мутагены различные виды излучений (рентгеновские лучи, гамма — лучи, а и
- 47. Естественный отбор Вызывает направленное изменение генофонда путем элиминации из популяции менее приспособленных особей или снижения их
- 48. Доминантная ахондроплазия (карликовость) Эта болезнь хорошо изучена в популяциях Дании. Больные имеют пониженную жизнеспособность и умирают
- 49. Доминантная ахондроплазия (карликовость) Выжившие карлики редко вступают в брак и имеют мало детей. Анализ показывает, что
- 50. Отбор мутантных генов по данным В. Маккьюсика (1968г) Большинство мутантов: 15 % плодов - погибают до
- 51. Значение мутированного гена В ряде случаев патологический ген в гетерозиготном состоянии может повышать жизнеспособность особи (серповидноклеточная
- 52. Наследование серповидно клеточной анемии Заболевание распространено в странах Африки и Азии. У людей, гомозиготных по аллелю
- 53. Наследование серповидно клеточной анемии Гетерозиготы НbАНbS более устойчивы к малярии, чем нормальные гомозиготы НbАНbА и НbSНbS.
- 54. Наследование серповидно клеточной анемии. Отбор работает в пользу гетерозигот. В районах, где не было малярии, гомозиготы
- 55. Наследование серповидно клеточной анемии. В некоторых районах Африки гетерозиготы составляют до 70 %. населения. "Платой" за
- 56. 2. Миграция генов Определение: процесс перемещения особей из одной популяции в другую и последующее скрещивание представителей
- 57. Значение миграции генов Миграция обеспечивает «поток генов», т.е. изменение генетического состава популяции, обусловленное поступлением новых генов.
- 58. Локальная популяция А, членов которой будем называть старожилами, и популяцию В, членов которой назовем мигрантами. Допустим,
- 59. Частота встречаемости аллеля а в следующем (смешанном) поколении в локальной популяции (популяции-реципиенте) составит: р 1 =
- 60. Применив элементарные алгебраические приемы и перегруппировав члены уравнения, находим, что новая частота аллеля равна исходной частоте
- 61. Т.е. чем больше доля пришельцев в популя-ции и чем больше различие в частотах алле-ля а у
- 62. Реальные популяции редко бывают полностью изолированными. Всегда происходит некоторое передвижение, которое может быть не только активным,
- 63. Случайный дрейф генов называется изменение частот аллелей в ряду поколений, являющееся результатом действия случайных причин, например
- 64. Общее правило случайных процессов таково: величина стандартного отклонения частот генов в популяции всегда находится в обратной
- 65. Так, в небольшом изоляте (дункеры в штате Пенсильвания, США, выходцы из Германии) частота генов групп крови
- 66. Примером действия дрейфа генов в человеческих популяциях может служить "эффект родоначальника". Он наблюдается, если структура популяции
- 67. Г) Естественный отбор. Большое значение имеет процесс ествест-венного отбора – т.е. процесс дифферен-циального воспроизводства потомства генетически
- 68. Выделяют 3 основных типа мутации: * вредоносные * нейтральные благоприятствующие. Большинство новых мутаций, возникающих в популяции,
- 69. Однако существуют мутации, появление которых не нарушает функционирование организма. Приспосабливаемость таких мутантов может быть даже высока,
- 70. Третий тип мутантов – благоприятствующий появляется крайне редко: такие мутации могут повысить приспособленность организма. В этом
- 71. Закон Харди – Вайнберга действует лищь тогда, когда скрещивание случайно и вероятность скрещивания двух генотиопов равно
- 72. Поскольку родственные особи в генетичес-ком отношении более сходны между собой, чем не стоящие в родствен организмы,
- 73. Общая закономерность заключается в том, что в популяциях инбридинг повышает частоту фенотипического проявления вредных рецессивных аллелей,
- 74. Мерой генетических последствий инбридинга служит коэффициент инбридинга, представляющий собой вероятность того, что у какой либо особи
- 75. Таблица Коэффициент инбридинга (F) в потомстве от родственных скрещиваний
- 76. У растений при самоопылении коэффициент самый высокий (F = 1/2). У человека супружеские отношения между родителями
- 77. Ассортивностью называется не случайное заключение браков на основе сходства по любым факторам. Ассортативность практически всегда осуществляется
- 78. Ассортивность, как и инбридинг, снижает гетерозиготность. В результате ассортивности, гомозиготы заключают браки с гомозиготами, а гетерозиготы
- 80. Скачать презентацию

Методы изучения генетики человека
Клинико – генеалогический
Близнецовый
Популяционно – статистический
Цитогенетический
Метод генетики соматических клеток
Биохимический
Методы изучения генетики человека
Клинико – генеалогический
Близнецовый
Популяционно – статистический
Цитогенетический
Метод генетики соматических клеток
Биохимический

Определение популяции (научное эволюционно-генетическое, Н.В. Тимофеева - Ресовским)
Это совокупность особей
Определение популяции (научное эволюционно-генетическое, Н.В. Тимофеева - Ресовским)
Это совокупность особей

Определение популяции
(продолжение)
внутри которого практически осуществляется та или иная степень панмиксии
Определение популяции
(продолжение)
внутри которого практически осуществляется та или иная степень панмиксии

Определение популяции (окончание).
эта совокупность особей отделена от соседних таких же
Определение популяции (окончание).
эта совокупность особей отделена от соседних таких же

Задачи популяционно -статистического метода:
1. Изучить генетическую структуру популяций, их генофонд, взаимодействие
Задачи популяционно -статистического метода:
1. Изучить генетическую структуру популяций, их генофонд, взаимодействие

Задачи популяционно -статистического метода:
2. Изучить наследственные болезни населения, частоту нормальных и
Задачи популяционно -статистического метода:
2. Изучить наследственные болезни населения, частоту нормальных и

Задачи популяционно -статистического метода:
3. Изучает закономерности распространения наследственных болезней в разных
Задачи популяционно -статистического метода:
3. Изучает закономерности распространения наследственных болезней в разных

Задачи популяционно -статистического метода:
4. Изучает частоты генов в популя-ции, включая наследственные
Задачи популяционно -статистического метода:
4. Изучает частоты генов в популя-ции, включая наследственные

Задачи популяционно -статистического метода:
6.Роль наследственности и среды в возникновении болезней с
Задачи популяционно -статистического метода:
6.Роль наследственности и среды в возникновении болезней с

Задачи популяционно -статистического метода:
7. Влияния наследственных и средовых факторов в создании
Задачи популяционно -статистического метода:
7. Влияния наследственных и средовых факторов в создании

Значение популяционно -статистического метода
Дает возможность определить генетическую структуру популяции, рассчитать
Значение популяционно -статистического метода
Дает возможность определить генетическую структуру популяции, рассчитать

Значение популяционно -статистического метода
Изучает генетическую структуру популяций, их генофонд, взаимодействие факторов,
Значение популяционно -статистического метода
Изучает генетическую структуру популяций, их генофонд, взаимодействие факторов,
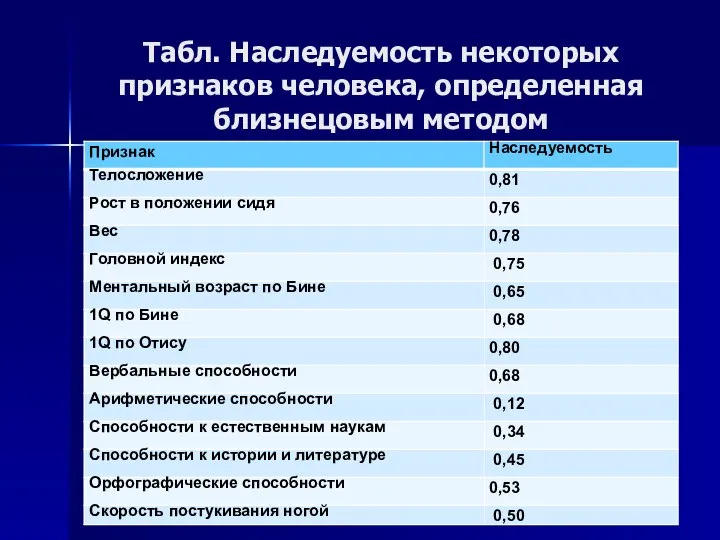
Табл. Наследуемость некоторых признаков человека, определенная близнецовым методом
Табл. Наследуемость некоторых признаков человека, определенная близнецовым методом

Популяционно-статистический метод включает:
1) правильный выбор популяции,
2) сбор материала и
3) статистический
Популяционно-статистический метод включает:
1) правильный выбор популяции,
2) сбор материала и
3) статистический

Основа популяционно- статистического метода
В основе метода лежит закономерность, установленная в 1908
Основа популяционно- статистического метода
В основе метода лежит закономерность, установленная в 1908

Черты идеальной популяции:
большая численность,
свободное скрещивание (панмиксия) организмов,
отсутствие отбора и
Черты идеальной популяции:
большая численность,
свободное скрещивание (панмиксия) организмов,
отсутствие отбора и

Основной смысл закона Харди - Вайнберга
В идеальной популяции соотношение частоты доминантных
Основной смысл закона Харди - Вайнберга
В идеальной популяции соотношение частоты доминантных

Факторы, стимулирующие сдвиг равновесия
в популяции:
родственные браки,
мутации,
дрейф генов,
Факторы, стимулирующие сдвиг равновесия
в популяции:
родственные браки,
мутации,
дрейф генов,

Значение закона Харди – Вайнберга.
Является основой при рассмотрении генетических преобразований, происходящих
Значение закона Харди – Вайнберга.
Является основой при рассмотрении генетических преобразований, происходящих

Соотношение численности разных генотипов и фенотипов в панмиктической популяции
Определяется по
Соотношение численности разных генотипов и фенотипов в панмиктической популяции
Определяется по

Обозначения в формуле бинома Ньютона
р — частота доминантного аллеля А,
Обозначения в формуле бинома Ньютона
р — частота доминантного аллеля А,

Пояснение закона Харди Вейнберга
Частота доминантных гомозигот (АА) равна квадрату вероятности встречае-мости
Пояснение закона Харди Вейнберга
Частота доминантных гомозигот (АА) равна квадрату вероятности встречае-мости

Пример наследования Rh - фактора
В одном из городов 16 % людей
Пример наследования Rh - фактора
В одном из городов 16 % людей

Разбор задачи:
Носители резус-положительного фактора могут иметь генотип СС или Сс. Чтобы
Разбор задачи:
Носители резус-положительного фактора могут иметь генотип СС или Сс. Чтобы

Расчет задачи:
Гомозиготы по рецессивному аллелю составляют 16% или 0,16; отсюда q
Расчет задачи:
Гомозиготы по рецессивному аллелю составляют 16% или 0,16; отсюда q

Расчет задачи:
Процент в популяции зигот СС и Сс вычисляем следующим образом:
СС
Расчет задачи:
Процент в популяции зигот СС и Сс вычисляем следующим образом:
СС

Заключение по задаче:
Следовательно, среди обследованного населения положительный резус-фактор имели 36 %
Заключение по задаче:
Следовательно, среди обследованного населения положительный резус-фактор имели 36 %

Особенности закона Харди – Вайнберга:
1. Описывает популяции в состоянии покоя.
В
Особенности закона Харди – Вайнберга:
1. Описывает популяции в состоянии покоя.
В

Особенности закона Харди – Вайнберга:
2. Закон Харди – Вайнберга гласит: при
Особенности закона Харди – Вайнберга:
2. Закон Харди – Вайнберга гласит: при

Особенности закона Харди – Вайнберга:
3. Задает точку отсчета, по отношению к
Особенности закона Харди – Вайнберга:
3. Задает точку отсчета, по отношению к

Особенности закона Харди – Вайнберга:
4. В малочисленных популяциях человека закон Харди
Особенности закона Харди – Вайнберга:
4. В малочисленных популяциях человека закон Харди

Эволюционные процессы в генетике человека
А) мутации
Б) миграции
В) дрейф генов
Г) естественный отбор
Эволюционные процессы в генетике человека
А) мутации
Б) миграции
В) дрейф генов
Г) естественный отбор

Определение мутации.
Мутации (изменения)- источники генетической изменчивости, но частота крайне низка.
Это
Определение мутации.
Мутации (изменения)- источники генетической изменчивости, но частота крайне низка.
Это

Пример расчета мутаций 1
Существуют два аллеля одного локуса (т. е. два
Пример расчета мутаций 1
Существуют два аллеля одного локуса (т. е. два

Пример расчета мутаций2
Допустим также, что в начальный момент времени (до начала
Пример расчета мутаций2
Допустим также, что в начальный момент времени (до начала

Пример расчета мутаций3
Во втором поколении доля V оставшихся аллелей а (частота
Пример расчета мутаций3
Во втором поколении доля V оставшихся аллелей а (частота

Пример расчета мутаций4
По прошествии t поколений частота аллеля а будет
Пример расчета мутаций4
По прошествии t поколений частота аллеля а будет

Пример расчета мутаций5
Однако все определяется величиной V.
В естественных условиях она
Пример расчета мутаций5
Однако все определяется величиной V.
В естественных условиях она

Пример расчета мутаций6
В таком темпе, для того чтобы изменить частоту аллеля
Пример расчета мутаций6
В таком темпе, для того чтобы изменить частоту аллеля

Значение мутаций в генетике
Для того чтобы мутации сами по себе привели
Значение мутаций в генетике
Для того чтобы мутации сами по себе привели

Виды мутаций по измеению генотипа
Генные (или точковые),
Внутрихромосомные,
Межхромосомные,
Виды мутаций по измеению генотипа
Генные (или точковые),
Внутрихромосомные,
Межхромосомные,

Виды генных мутаций
Прямые (А>а) и обратные (а>А). Частота возникновения прямых му-таций
Виды генных мутаций
Прямые (А>а) и обратные (а>А). Частота возникновения прямых му-таций

Частота мутаций, обусловливающих тяжелые болезни
Частота возникновения патологи-ческих мутаций отдельного гена (гемофилия,
Частота мутаций, обусловливающих тяжелые болезни
Частота возникновения патологи-ческих мутаций отдельного гена (гемофилия,

Чем обусловлен рост мутаций?
Химические мутагены : (канцерогены) - промышленные яды,
Чем обусловлен рост мутаций?
Химические мутагены : (канцерогены) - промышленные яды,

Чем обусловлен рост мутаций?
Сильные мутагены различные виды излучений (рентгеновские лучи,
Чем обусловлен рост мутаций?
Сильные мутагены различные виды излучений (рентгеновские лучи,

Естественный отбор
Вызывает направленное изменение генофонда путем элиминации из популяции менее приспособленных
Естественный отбор
Вызывает направленное изменение генофонда путем элиминации из популяции менее приспособленных

Доминантная ахондроплазия (карликовость)
Эта болезнь хорошо изучена в популяциях Дании. Больные имеют
Доминантная ахондроплазия (карликовость)
Эта болезнь хорошо изучена в популяциях Дании. Больные имеют

Доминантная ахондроплазия (карликовость)
Выжившие карлики редко вступают в брак и имеют мало
Доминантная ахондроплазия (карликовость)
Выжившие карлики редко вступают в брак и имеют мало

Отбор мутантных генов по данным В. Маккьюсика (1968г)
Большинство мутантов:
15 %
Отбор мутантных генов по данным В. Маккьюсика (1968г)
Большинство мутантов:
15 %

Значение мутированного гена
В ряде случаев патологический ген в гетерозиготном состоянии может
Значение мутированного гена
В ряде случаев патологический ген в гетерозиготном состоянии может

Наследование серповидно клеточной анемии
Заболевание распространено в странах Африки и Азии.
У
Наследование серповидно клеточной анемии
Заболевание распространено в странах Африки и Азии.
У

Наследование серповидно клеточной анемии
Гетерозиготы НbАНbS более устойчивы к малярии, чем нормальные
Наследование серповидно клеточной анемии
Гетерозиготы НbАНbS более устойчивы к малярии, чем нормальные

Наследование серповидно клеточной анемии.
Отбор работает в пользу гетерозигот. В районах, где
Наследование серповидно клеточной анемии.
Отбор работает в пользу гетерозигот. В районах, где

Наследование серповидно клеточной анемии.
В некоторых районах Африки гетерозиготы составляют до 70
Наследование серповидно клеточной анемии.
В некоторых районах Африки гетерозиготы составляют до 70

2. Миграция генов
Определение:
процесс перемещения особей из одной популяции в другую
2. Миграция генов
Определение:
процесс перемещения особей из одной популяции в другую

Значение миграции генов
Миграция обеспечивает «поток генов», т.е. изменение генетического состава популяции,
Значение миграции генов
Миграция обеспечивает «поток генов», т.е. изменение генетического состава популяции,

Локальная популяция А, членов которой будем называть старожилами, и популяцию В,
Локальная популяция А, членов которой будем называть старожилами, и популяцию В,

Частота встречаемости аллеля а в следующем (смешанном) поколении в локальной популяции
Частота встречаемости аллеля а в следующем (смешанном) поколении в локальной популяции

Применив элементарные алгебраические приемы и перегруппировав члены уравнения, находим, что новая
Применив элементарные алгебраические приемы и перегруппировав члены уравнения, находим, что новая

Т.е. чем больше доля пришельцев в популя-ции и чем больше различие
Т.е. чем больше доля пришельцев в популя-ции и чем больше различие

Реальные популяции редко бывают полностью изолированными. Всегда происходит некоторое
Реальные популяции редко бывают полностью изолированными. Всегда происходит некоторое

Случайный дрейф генов
называется изменение частот аллелей в ряду поколений,
Случайный дрейф генов
называется изменение частот аллелей в ряду поколений,

Общее правило случайных процессов таково: величина стандартного отклонения частот генов в
Общее правило случайных процессов таково: величина стандартного отклонения частот генов в

Так, в небольшом изоляте (дункеры в штате Пенсильвания, США, выходцы из
Так, в небольшом изоляте (дункеры в штате Пенсильвания, США, выходцы из

Примером действия дрейфа генов в человеческих популяциях может служить "эффект родоначальника".
Примером действия дрейфа генов в человеческих популяциях может служить "эффект родоначальника".

Г) Естественный отбор.
Большое значение имеет процесс ествест-венного отбора –
Г) Естественный отбор.
Большое значение имеет процесс ествест-венного отбора –

Выделяют 3 основных типа мутации:
* вредоносные
* нейтральные
благоприятствующие.
Большинство новых мутаций,
Выделяют 3 основных типа мутации:
* вредоносные
* нейтральные
благоприятствующие.
Большинство новых мутаций,

Однако существуют мутации, появление которых не нарушает функционирование организма. Приспосабливаемость
Однако существуют мутации, появление которых не нарушает функционирование организма. Приспосабливаемость

Третий тип мутантов – благоприятствующий появляется крайне редко: такие мутации
Третий тип мутантов – благоприятствующий появляется крайне редко: такие мутации

Закон Харди – Вайнберга действует лищь тогда, когда скрещивание случайно и
Закон Харди – Вайнберга действует лищь тогда, когда скрещивание случайно и

Поскольку родственные особи в генетичес-ком отношении более сходны между собой, чем
Поскольку родственные особи в генетичес-ком отношении более сходны между собой, чем

Общая закономерность заключается в том, что в популяциях инбридинг повышает
Общая закономерность заключается в том, что в популяциях инбридинг повышает

Мерой генетических последствий инбридинга служит коэффициент инбридинга, представляющий собой вероятность того,
Мерой генетических последствий инбридинга служит коэффициент инбридинга, представляющий собой вероятность того,
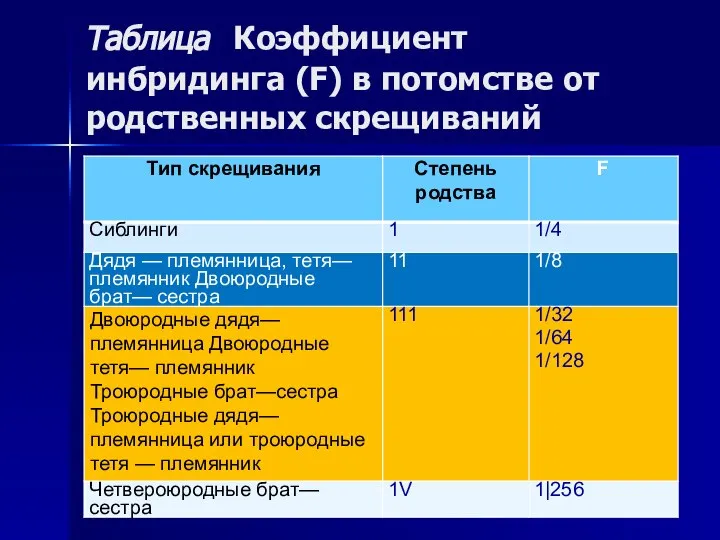
Таблица Коэффициент инбридинга (F) в потомстве от родственных скрещиваний
Таблица Коэффициент инбридинга (F) в потомстве от родственных скрещиваний

У растений при самоопылении коэффициент самый высокий (F = 1/2).
У
У растений при самоопылении коэффициент самый высокий (F = 1/2).
У

Ассортивностью называется не случайное заключение браков на основе сходства по любым
Ассортивностью называется не случайное заключение браков на основе сходства по любым

Ассортивность, как и инбридинг, снижает гетерозиготность. В результате ассортивности, гомозиготы заключают
Ассортивность, как и инбридинг, снижает гетерозиготность. В результате ассортивности, гомозиготы заключают
 Клуб исторического фехтования "Древняя Русь"
Клуб исторического фехтования "Древняя Русь" Презентация на тему "Мотивировать и обучать на расстоянии" - скачать презентации по Педагогике
Презентация на тему "Мотивировать и обучать на расстоянии" - скачать презентации по Педагогике Духовная культура
Духовная культура Нищете материальной нетрудно помочь, нищете души - невозможно.
Нищете материальной нетрудно помочь, нищете души - невозможно.  Йоль. Символика Йоля
Йоль. Символика Йоля Открытый кубок Санкт-Петербурга в классе яхт ORC
Открытый кубок Санкт-Петербурга в классе яхт ORC Moderne kommunikations mittel
Moderne kommunikations mittel Патология иммунитета
Патология иммунитета Организация видов работ при эксплуатации и реконструкции строительных объектов
Организация видов работ при эксплуатации и реконструкции строительных объектов Úvod do předmětu, právo a IT
Úvod do předmětu, právo a IT Синхронные машины
Синхронные машины Україна
Україна Das Roboterauto oder das fahrerlose Transportfahrzeug (Autonomus)
Das Roboterauto oder das fahrerlose Transportfahrzeug (Autonomus) Выполнили: Мельников Дмитрий, Лукин Максим
Выполнили: Мельников Дмитрий, Лукин Максим Цифровые элементы
Цифровые элементы Политическая элита и политическое лидерство
Политическая элита и политическое лидерство Формирования социального государства в России
Формирования социального государства в России Диаграмма деятельности языка UML 2 (Лекция 6)
Диаграмма деятельности языка UML 2 (Лекция 6) Neue Wörter
Neue Wörter Займись спортом! Участвуй в ГТО
Займись спортом! Участвуй в ГТО Смешанные и кочующие правовые системы
Смешанные и кочующие правовые системы Участие в программах поддержки малого и среднего бизнеса: возможности и риски
Участие в программах поддержки малого и среднего бизнеса: возможности и риски Комплектование ДОУ в 2017-2018 учебном году
Комплектование ДОУ в 2017-2018 учебном году Воскрешение Ваз 2107
Воскрешение Ваз 2107 Защита от ионизирующих излучений
Защита от ионизирующих излучений  ГБОУ ВПО РОСТОВСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ Кафедра детских инфекционных болезней
ГБОУ ВПО РОСТОВСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ Кафедра детских инфекционных болезней Promshum
Promshum Фальсификация и идентификация пищевых продуктов
Фальсификация и идентификация пищевых продуктов