- Построение филогенетических деревьев

Содержание
- 2. Особенности молекулярной эволюции 1. Скорость эволюции любого белка, выраженная через число аминокислотных замен на сайт в
- 3. Что такое филогенетические деревья? Дерево — это граф, в котором два соседних узла соединены только одним
- 4. Кладограммы и филограммы Кладограммы отражают только порядок ветвления, филограммы — ещё и длину ветвей
- 5. Сколько здесь разных кладограмм? a b c d e a e c d b a b
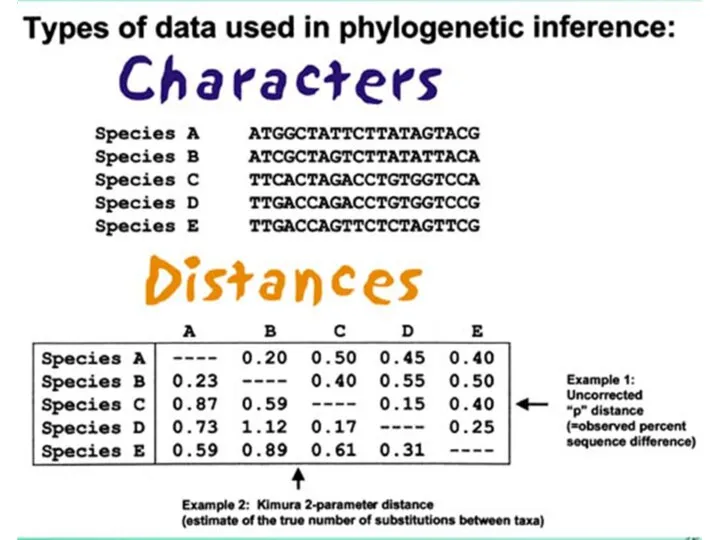
- 6. Выбор последовательностей Последовательности должны быть гомологичны! Программа выровняет любые последовательности => нужно проверить с помощью Blast
- 7. «Эффект тыквенного пирога» Рецепт тыквенного пирога на филогенетическом дереве креветок.
- 8. Выбор последовательностей
- 9. Особенности молекулярной эволюции 2. Функционально менее важные молекулы или их части эволюционируют (накапливая эволюционные замены) быстрее,
- 10. Различия между деревом генов и деревом видов Проблема: ортологи и паралоги
- 11. Молекулярная конвергенция
- 12. Филогенетические маркёры Свойства: Гены, которые представлены одной копией в геноме лучше, чем те, у которых множество
- 14. Филогенетические маркёры Рибосомальные гены Митохондриальные гены (COI/II, 12s RNA, cyt b) Хлоропластные гены Гены домашнего хозяйства
- 15. Выбор модели замен Результаты вычисления эволюционных дистанций будут отличаться в зависимости от выбранной модели замен
- 16. Выбор модели замен AIC — Akaike’s Information Criterion. Быстрее BIC — Bayesian information criteria. Не «любит»
- 18. Методы реконструкции филогении
- 19. Дистанционные методы Neghbor-joining Начинаем с пары ветвей, которые меньше всего отличаются между собой
- 20. Дистанционные методы Neghbor-joining
- 21. Дистанционные методы Neghbor-joining
- 22. Зачем нужна аутгруппа Молекулярно-филогенетические методы используют информацию о последовательностях внешней группы (контроля), дистанция от которой для
- 23. Дистанционные методы Neghbor-joining Не учитываются обратные и параллельные замены => Мы считаем не настоящую дистанцию (расстояние),
- 24. Методы максимальной экономии Минимизация числа замен символов Всегда реконструируют предковые последовательности Лучше работает на небольших наборах
- 25. Методы максимальной экономии B C A B C A B C A B C A D
- 27. Методы максимальной вероятности Так же, как и в случае с методами максимальной экономии, генерирует все возможные
- 28. Методы максимальной вероятности В позиции j для каждого внутреннего узла допустимы все четыре нуклеотида, значит всего
- 29. Оценка поддержки дерева Bootstrap
- 31. Скачать презентацию

Особенности молекулярной эволюции
1. Скорость эволюции любого белка, выраженная через число аминокислотных
Особенности молекулярной эволюции
1. Скорость эволюции любого белка, выраженная через число аминокислотных
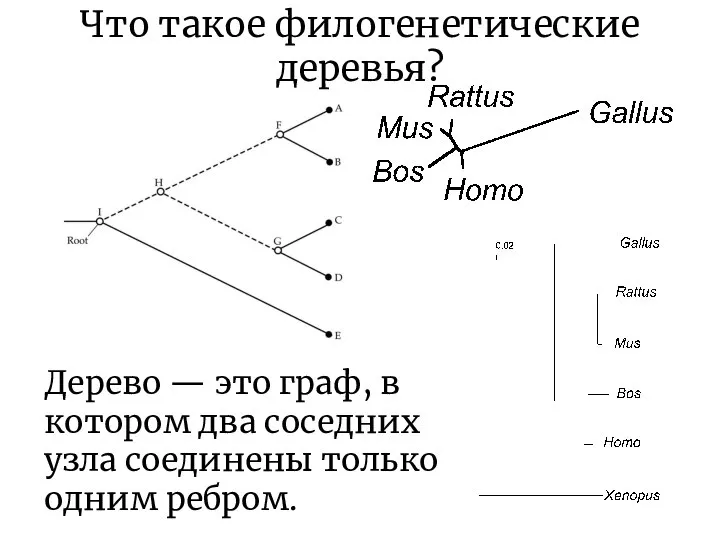
Что такое филогенетические деревья?
Дерево — это граф, в котором два соседних
Что такое филогенетические деревья?
Дерево — это граф, в котором два соседних
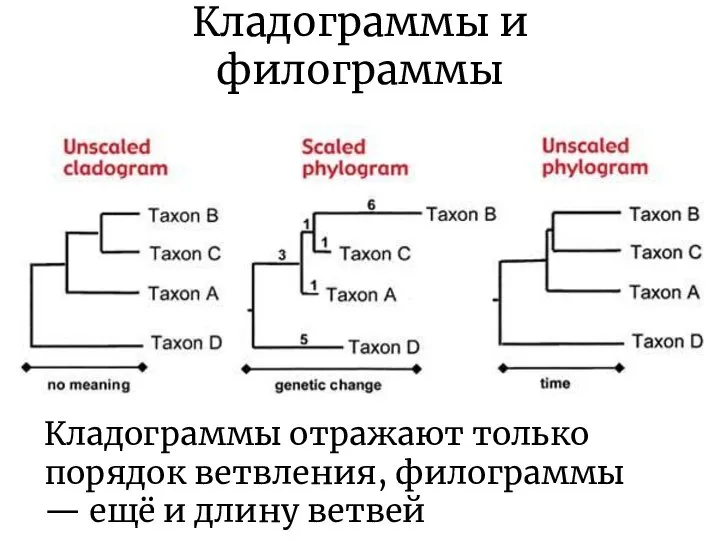
Кладограммы и филограммы
Кладограммы отражают только порядок ветвления, филограммы — ещё и
Кладограммы и филограммы
Кладограммы отражают только порядок ветвления, филограммы — ещё и
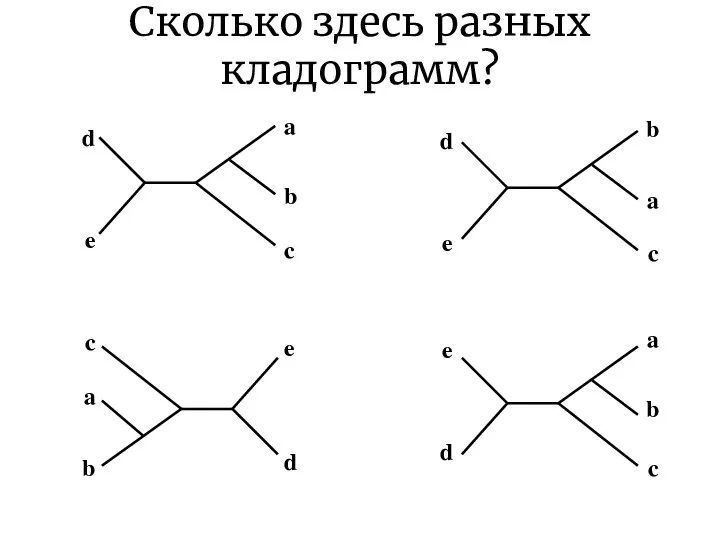
Сколько здесь разных кладограмм?
a
b
c
d
e
a
e
c
d
b
a
b
c
e
d
b
a
c
d
e
Сколько здесь разных кладограмм?
a
b
c
d
e
a
e
c
d
b
a
b
c
e
d
b
a
c
d
e

Выбор последовательностей
Последовательности должны быть гомологичны! Программа выровняет любые последовательности => нужно
Выбор последовательностей
Последовательности должны быть гомологичны! Программа выровняет любые последовательности => нужно
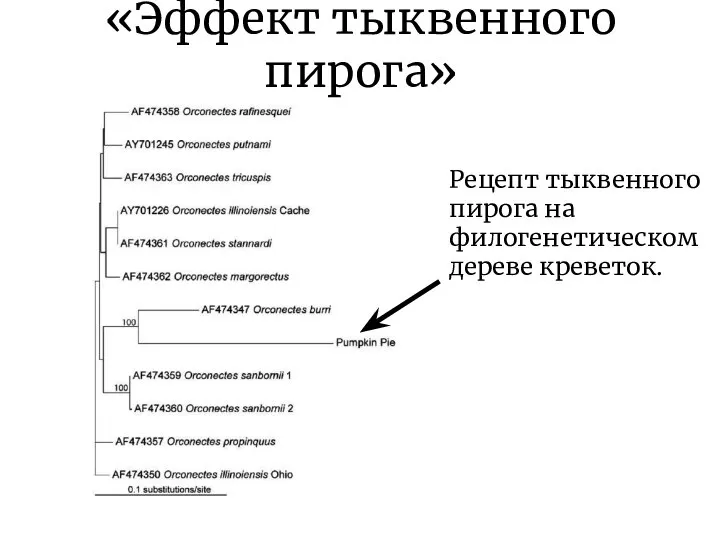
«Эффект тыквенного пирога»
Рецепт тыквенного пирога на филогенетическом дереве креветок.
«Эффект тыквенного пирога»
Рецепт тыквенного пирога на филогенетическом дереве креветок.
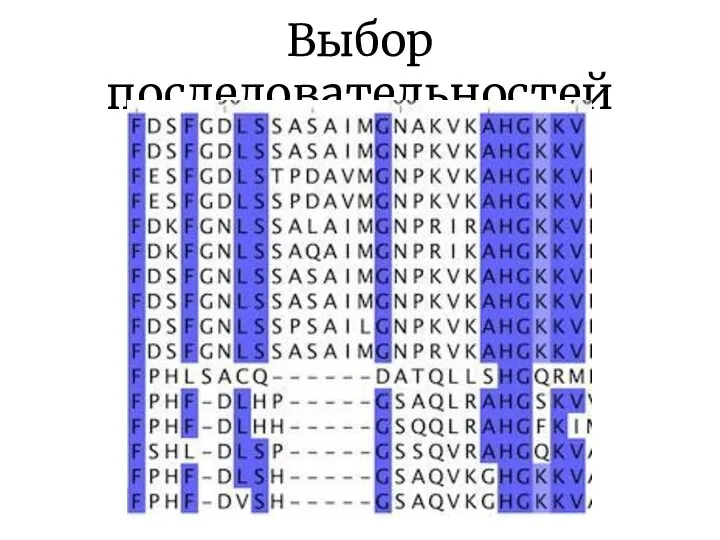
Выбор последовательностей
Выбор последовательностей

Особенности молекулярной эволюции
2. Функционально менее важные молекулы или их части эволюционируют
Особенности молекулярной эволюции
2. Функционально менее важные молекулы или их части эволюционируют
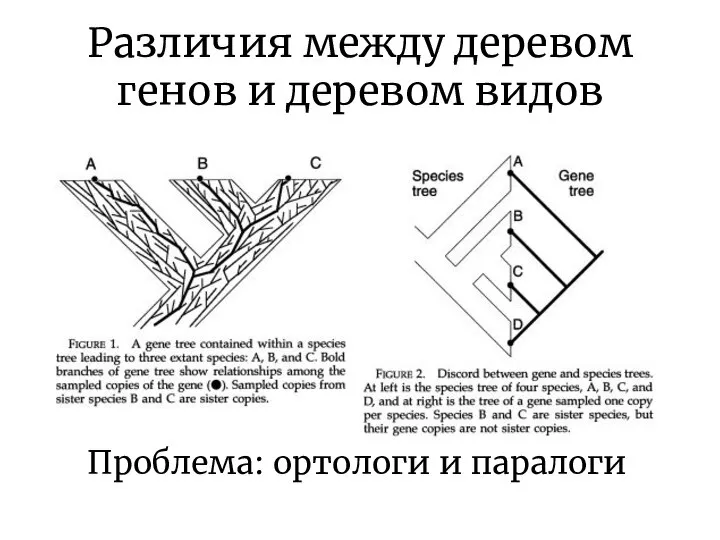
Различия между деревом генов и деревом видов
Проблема: ортологи и паралоги
Различия между деревом генов и деревом видов
Проблема: ортологи и паралоги
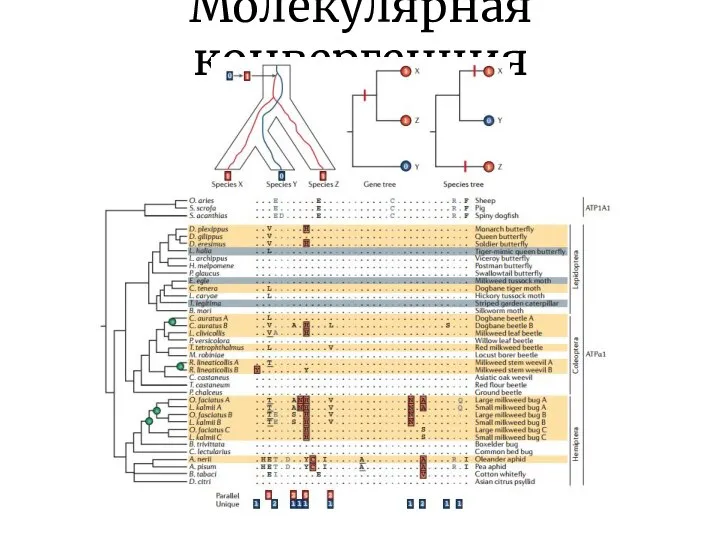
Молекулярная конвергенция
Молекулярная конвергенция

Филогенетические маркёры
Свойства:
Гены, которые представлены одной копией в геноме лучше, чем
Филогенетические маркёры
Свойства:
Гены, которые представлены одной копией в геноме лучше, чем

Филогенетические маркёры
Рибосомальные гены
Митохондриальные гены (COI/II, 12s RNA, cyt b)
Хлоропластные гены
Гены
Филогенетические маркёры
Рибосомальные гены
Митохондриальные гены (COI/II, 12s RNA, cyt b)
Хлоропластные гены
Гены

Выбор модели замен
Результаты вычисления эволюционных дистанций будут отличаться в зависимости от
Выбор модели замен
Результаты вычисления эволюционных дистанций будут отличаться в зависимости от
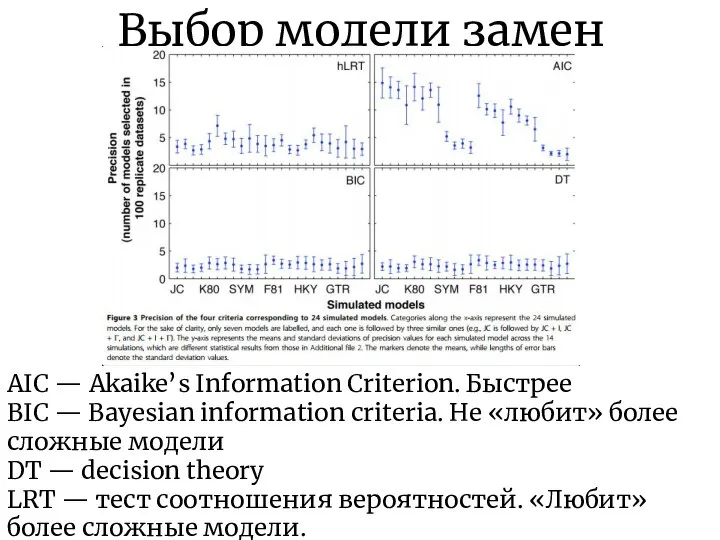
Выбор модели замен
AIC — Akaike’s Information Criterion. Быстрее
BIC — Bayesian information
Выбор модели замен
AIC — Akaike’s Information Criterion. Быстрее
BIC — Bayesian information
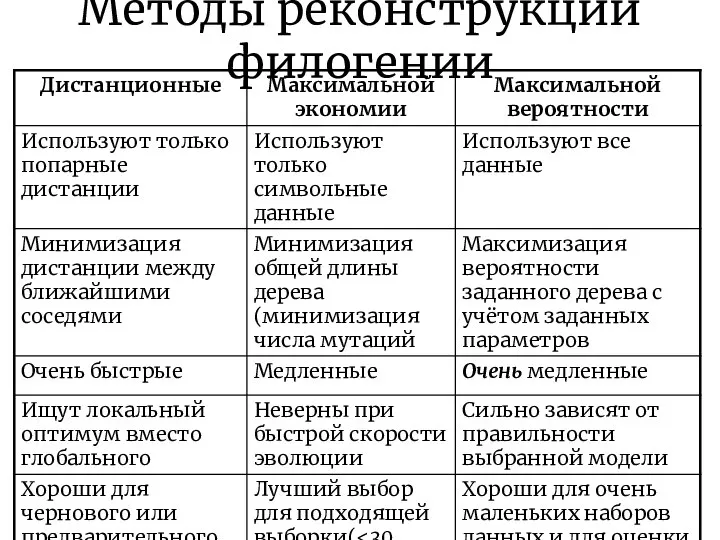
Методы реконструкции филогении
Методы реконструкции филогении
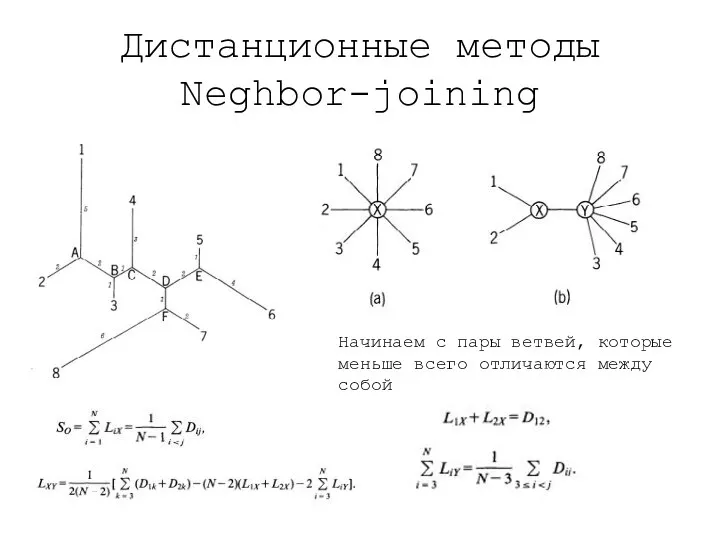
Дистанционные методы
Neghbor-joining
Начинаем с пары ветвей, которые меньше всего отличаются между собой
Дистанционные методы
Neghbor-joining
Начинаем с пары ветвей, которые меньше всего отличаются между собой

Дистанционные методы
Neghbor-joining
Дистанционные методы
Neghbor-joining
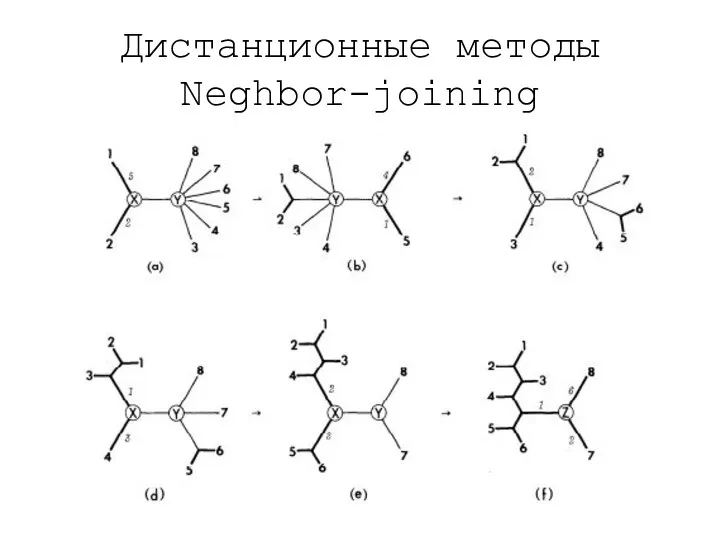
Дистанционные методы
Neghbor-joining
Дистанционные методы
Neghbor-joining
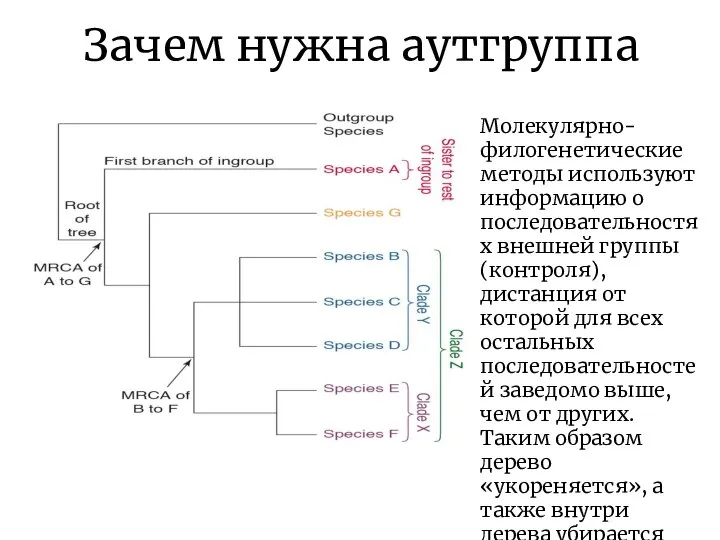
Зачем нужна аутгруппа
Молекулярно-филогенетические методы используют информацию о последовательностях внешней группы (контроля),
Зачем нужна аутгруппа
Молекулярно-филогенетические методы используют информацию о последовательностях внешней группы (контроля),
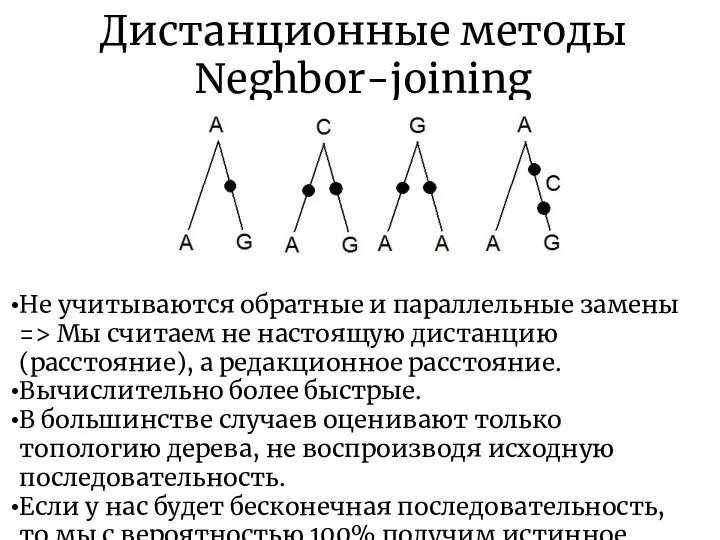
Дистанционные методы
Neghbor-joining
Не учитываются обратные и параллельные замены
=> Мы считаем не настоящую
Дистанционные методы
Neghbor-joining
Не учитываются обратные и параллельные замены
=> Мы считаем не настоящую

Методы максимальной экономии
Минимизация числа замен символов
Всегда реконструируют предковые последовательности
Лучше работает на
небольших
Методы максимальной экономии
Минимизация числа замен символов
Всегда реконструируют предковые последовательности
Лучше работает на
небольших
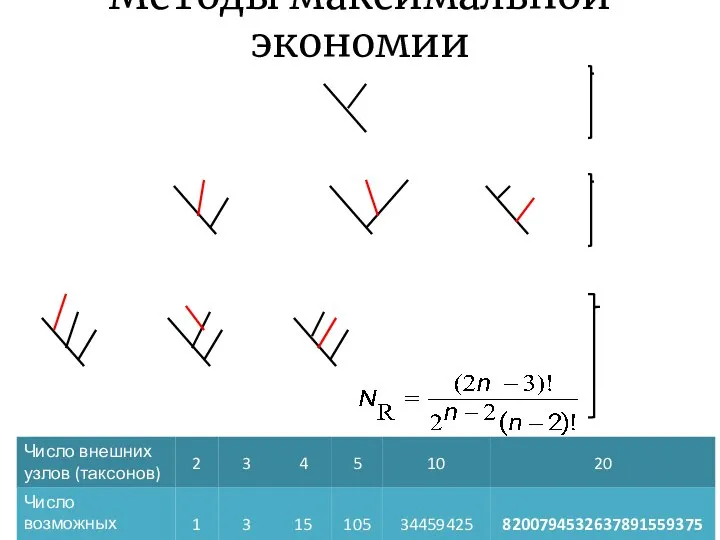
Методы максимальной экономии
B
C
A
B
C
A
B
C
A
B
C
A
D
D
D
B
C
A
D
B
C
A
D
E
E
B
C
A
D
E
…………………
Step 1
Step 2
Step 3
Методы максимальной экономии
B
C
A
B
C
A
B
C
A
B
C
A
D
D
D
B
C
A
D
B
C
A
D
E
E
B
C
A
D
E
…………………
Step 1
Step 2
Step 3
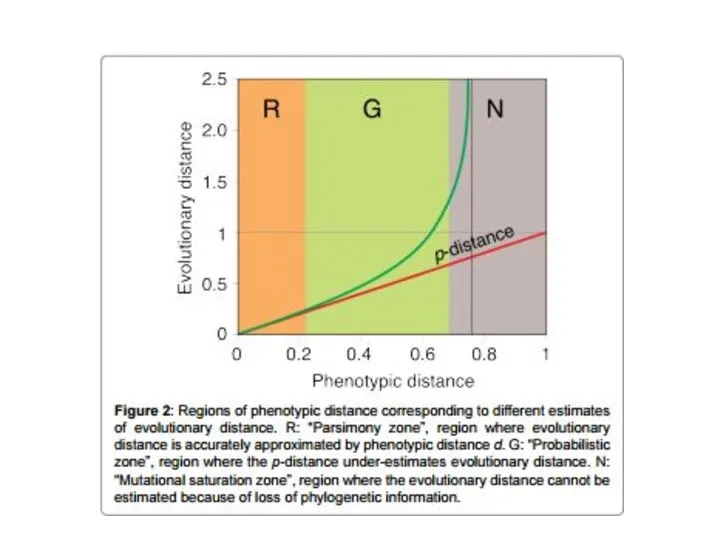

Методы максимальной вероятности
Так же, как и в случае с методами максимальной
Методы максимальной вероятности
Так же, как и в случае с методами максимальной

Методы максимальной вероятности
В позиции j для каждого внутреннего узла допустимы все
Методы максимальной вероятности
В позиции j для каждого внутреннего узла допустимы все
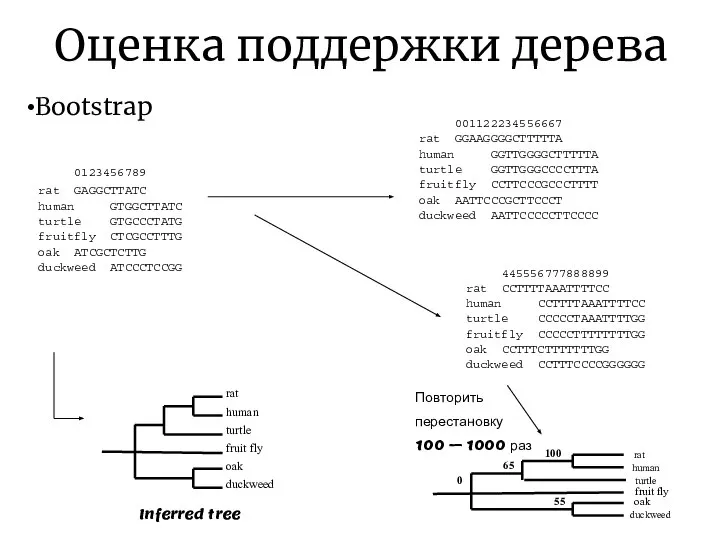
Оценка поддержки дерева
Bootstrap
Оценка поддержки дерева
Bootstrap
 Sport in France
Sport in France Введение в язык программирования Python
Введение в язык программирования Python Культура и традиции народов Якутии
Культура и традиции народов Якутии Тип запись. Структурный тип
Тип запись. Структурный тип Генетический код
Генетический код Эволюция технологии разработки ПО. Процедурная и объектная декомпозиция
Эволюция технологии разработки ПО. Процедурная и объектная декомпозиция Физиология пищеварения
Физиология пищеварения  Генеральный план микрорайона «Улитка»
Генеральный план микрорайона «Улитка» Я и другие люди. Вежливость - презентация для начальной школы
Я и другие люди. Вежливость - презентация для начальной школы Лечка. Аускультация шумов сердца
Лечка. Аускультация шумов сердца Костюм Древнего Рима (V век до н. э. — V век н. э.)
Костюм Древнего Рима (V век до н. э. — V век н. э.) Специфика перевода
Специфика перевода Проблема защиты авторских прав. Подготовила студентка группы Юб 03/1303 Селезнева М.
Проблема защиты авторских прав. Подготовила студентка группы Юб 03/1303 Селезнева М.  Петушок – золотой гребешок - презентация для начальной школы
Петушок – золотой гребешок - презентация для начальной школы Травмы магистральных сосудов
Травмы магистральных сосудов Flüchtlinge in Deutschland
Flüchtlinge in Deutschland Weihnachten in Deutschland
Weihnachten in Deutschland Возможности системы DELO
Возможности системы DELO Презентация Microsoft Office PowerPoint (2)
Презентация Microsoft Office PowerPoint (2) Дорожная безопасность Материалы к занятиям по ПДД для учащихся 4 класса
Дорожная безопасность Материалы к занятиям по ПДД для учащихся 4 класса Мировая телеология и холизм. Мировой контроль. Тайное знание. Заговорщики. Тезис о заговоре
Мировая телеология и холизм. Мировой контроль. Тайное знание. Заговорщики. Тезис о заговоре Мультиплексор и демультиплексор
Мультиплексор и демультиплексор Собеседование по найму
Собеседование по найму Презентация "Архитектура западноевропейского Средневековья. Романский стиль" - скачать презентации по МХК
Презентация "Архитектура западноевропейского Средневековья. Романский стиль" - скачать презентации по МХК Кроки вступу до Запорізького Національного Унівкрситету
Кроки вступу до Запорізького Національного Унівкрситету Основы религиозной культуры и светской этики
Основы религиозной культуры и светской этики Тема занятия: Патофизиология энергетического и углеводного обмена
Тема занятия: Патофизиология энергетического и углеводного обмена  Сравнительная характеристика понятий «Управление Человеческими Ресурсами» и «Управление Персоналом» Выполнили: Епифанова Евге
Сравнительная характеристика понятий «Управление Человеческими Ресурсами» и «Управление Персоналом» Выполнили: Епифанова Евге