- Эволюционная биология развития. Введение и общие понятия. (Тема 1)

Содержание
- 2. Эволюционная биология развития - область знаний, изучающая механизмы эволюции и возникшая из сочетания трех наук: Эмбриология
- 3. Термин “развитие” (development) в данном контексте обозначает только индивидуальное развитие, т.е. онтогенез.
- 4. Два подхода к развитию: Изучение судьбы и взаимодействия зачатков в развивающемся зародыше (подход механики развития). Изучение
- 5. Пример первого подхода: пересадка зачатка хорды привела к появлению полного дополнительного комплекса осевых органов (Spemann, Mangold,
- 6. Пример второго подхода: области экспрессии семи генов семейства Hox, показанные в эмбрионе дрозофилы методом гибридизации in
- 7. Август Вейсман - создатель первой теории генетической регуляции развития (задолго до возникновения понятий “ген” и “генетика”).
- 8. По Вейсману, клетки многоклеточного организма отличаются друг от друга набором ядерных детерминантов (= генов). Полный набор
- 9. Такой механизм дифференцировки клеток вполне реален. Например, он встречается у нематод: на схеме - диминуция хроматина
- 10. Рисунок самого Теодора Бовери: зародыш аскариды на стадии дробления. Видно, как в некоторых клетках частично разрушаются
- 11. Карта развития аскариды. Красный - клетки зародышевого пути с полным геномом, желтый - пресоматические клетки, прошедшие
- 12. Диминуция хроматина - достаточно распространенное (но не всеобщее!) явление. Кроме нематод, она описана у насекомых, ракообразных,
- 13. Тем не менее у большинства животных соматические клетки содержат тот же набор генов, что и половые
- 14. Типичное число генов многоклеточного животного - около 20 тысяч. Какие из них особенно важны с точки
- 15. Характерные (но не всеобщие) свойства генов развития (developmental genes): Их области экспрессии часто охватывают целые отделы,
- 16. Под фактором транскрипции здесь понимается белок, который «умеет» включать (усиливать) или выключать (ослаблять) транскрипцию других генов,
- 17. Белки, которые связываются с энхансером и сайленсером - это и есть факторы транскрипции. Любой фактор транскрипции
- 18. Довольно часто проявлением мутаций генов развития служит гомеозис (букв. «уподобление») - тип изменчивости, при котором один
- 19. Уильям Бэтсон (1861-1925). Слева - парадный портрет, справа - карикатура: Бэтсон изучает изменчивость дятлов.
- 20. Пример гомеозиса: химическое воздействие на регенерирующий после ампутации хвост головастика вызвало избыточную экспрессию одного из генов
- 21. Ретиноевая кислота - вещество, использованное в опыте Мюллера и Вагнера. Это пример морфогена - сигнального вещества,
- 22. В организме часто возникает градиент концентрации морфогена, т.е. ее направленное изменение. Например, BMP и Wnt -
- 23. Рассмотрим модель, в которой: Концентрация вещества-морфогена линейно растет вдоль тела животного. Есть гены А и Б,
- 24. Модель французского флага (Гилберт, 1995).
- 25. Позиционная информация - комплекс физико-химических факторов, действующих на клетку, зависящих от ее положения и определяющих ее
- 26. Более сложный вариант позиционной информации: формирование волоска на листе растения. Морфоген-активатор (обычно ауксин) стимулирует рост волоска,
- 27. Гены развития часто группируются в семейства. Семейство генов - это группа генов, происходящих от одного гена-предшественника
- 28. Еще один пример гомеозиса: мутация bithorax, превращающая третий грудной сегмент дрозофилы во второй (T3→T2). Получается муха
- 29. При отсутствии в области будущего сегмента T3 продукта определенного гена (он называется Ubx) вместо сегмента T3
- 30. Не кроется ли здесь механизм резких эволюционных изменений? Веерокрылые - отряд насекомых, у которых на 2-м
- 31. Молекулярная филогения, поддерживающая объединение двукрылых и веерокрылых в группу Halteria (Whiting, 2001).
- 32. Молекулярная филогения, отвергающая гипотезу Halteria (Rokas et al., 1999). Здесь ближайшими родственниками двукрылых оказываются бабочки.
- 33. Еще одна молекулярная филогения, на которой веерокрылые оказываются родственниками жуков (Wiegmann et al., 2009).
- 34. Гипотеза Halteria на первый взгляд выглядит убедительной. Однако: Филогенетические исследования не подтверждают существования ветви, включающей только
- 35. В предельно редуцированном виде все развитие может рассматриваться как последовательность взаимосвязанных актов экспрессии генов. На схеме
- 37. Скачать презентацию

Эволюционная биология развития - область знаний, изучающая механизмы эволюции и возникшая
Эволюционная биология развития - область знаний, изучающая механизмы эволюции и возникшая
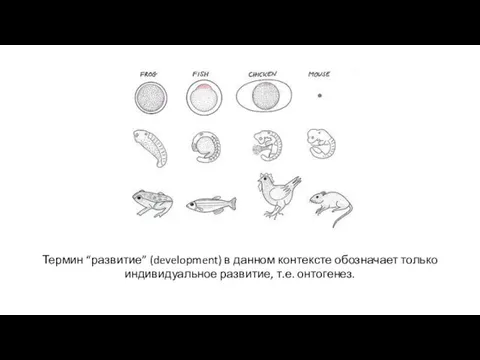
Термин “развитие” (development) в данном контексте обозначает только индивидуальное развитие, т.е.
Термин “развитие” (development) в данном контексте обозначает только индивидуальное развитие, т.е.

Два подхода к развитию:
Изучение судьбы и взаимодействия зачатков в развивающемся зародыше
Два подхода к развитию:
Изучение судьбы и взаимодействия зачатков в развивающемся зародыше
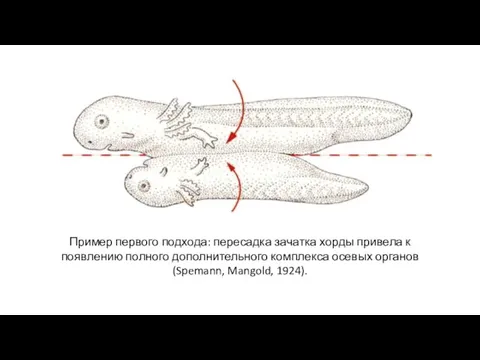
Пример первого подхода: пересадка зачатка хорды привела к появлению полного дополнительного
Пример первого подхода: пересадка зачатка хорды привела к появлению полного дополнительного
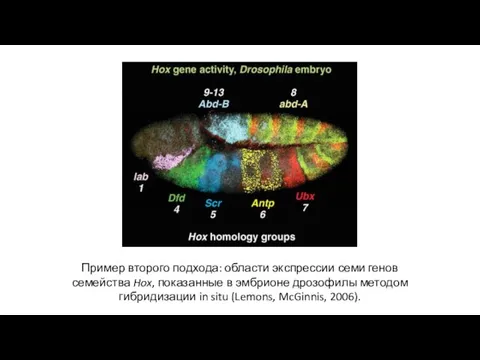
Пример второго подхода: области экспрессии семи генов семейства Hox, показанные в
Пример второго подхода: области экспрессии семи генов семейства Hox, показанные в

Август Вейсман - создатель первой теории генетической регуляции развития (задолго до
Август Вейсман - создатель первой теории генетической регуляции развития (задолго до
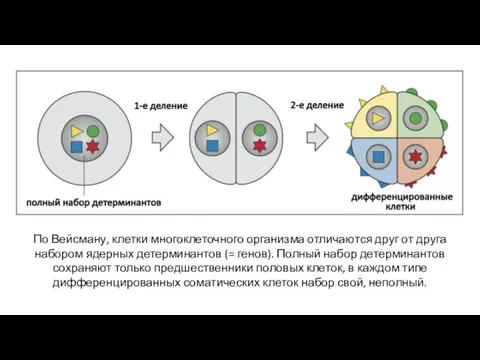
По Вейсману, клетки многоклеточного организма отличаются друг от друга набором ядерных
По Вейсману, клетки многоклеточного организма отличаются друг от друга набором ядерных
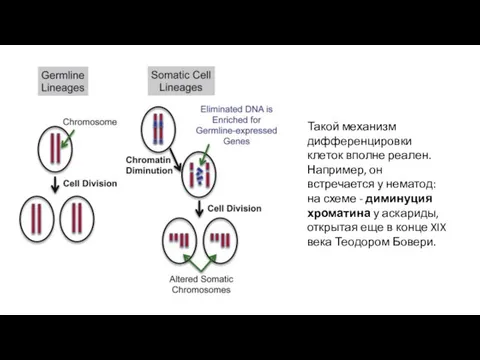
Такой механизм дифференцировки клеток вполне реален. Например, он встречается у нематод:
Такой механизм дифференцировки клеток вполне реален. Например, он встречается у нематод:

Рисунок самого Теодора Бовери: зародыш аскариды на стадии дробления. Видно, как
Рисунок самого Теодора Бовери: зародыш аскариды на стадии дробления. Видно, как

Карта развития аскариды. Красный - клетки зародышевого пути с полным геномом,
Карта развития аскариды. Красный - клетки зародышевого пути с полным геномом,

Диминуция хроматина - достаточно распространенное (но не всеобщее!) явление. Кроме нематод,
Диминуция хроматина - достаточно распространенное (но не всеобщее!) явление. Кроме нематод,
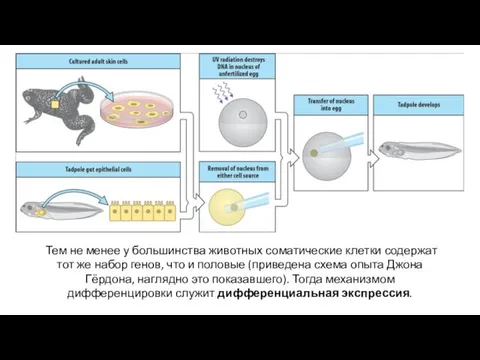
Тем не менее у большинства животных соматические клетки содержат тот же
Тем не менее у большинства животных соматические клетки содержат тот же
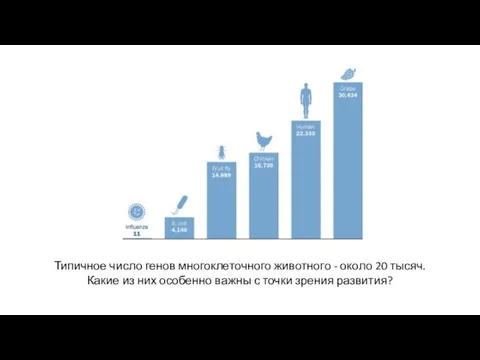
Типичное число генов многоклеточного животного - около 20 тысяч.
Какие из
Типичное число генов многоклеточного животного - около 20 тысяч.
Какие из

Характерные (но не всеобщие) свойства генов развития (developmental genes):
Их области экспрессии
Характерные (но не всеобщие) свойства генов развития (developmental genes):
Их области экспрессии
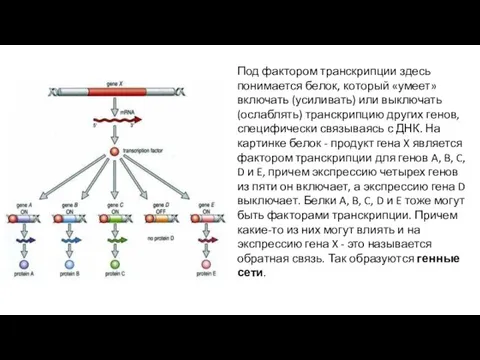
Под фактором транскрипции здесь понимается белок, который «умеет» включать (усиливать) или
Под фактором транскрипции здесь понимается белок, который «умеет» включать (усиливать) или
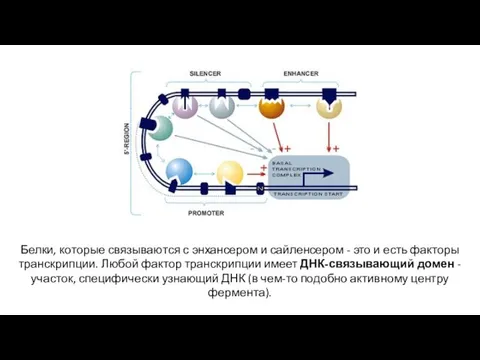
Белки, которые связываются с энхансером и сайленсером - это и есть
Белки, которые связываются с энхансером и сайленсером - это и есть

Довольно часто проявлением мутаций генов развития служит гомеозис (букв. «уподобление») -
Довольно часто проявлением мутаций генов развития служит гомеозис (букв. «уподобление») -

Уильям Бэтсон (1861-1925). Слева - парадный портрет, справа - карикатура: Бэтсон
Уильям Бэтсон (1861-1925). Слева - парадный портрет, справа - карикатура: Бэтсон
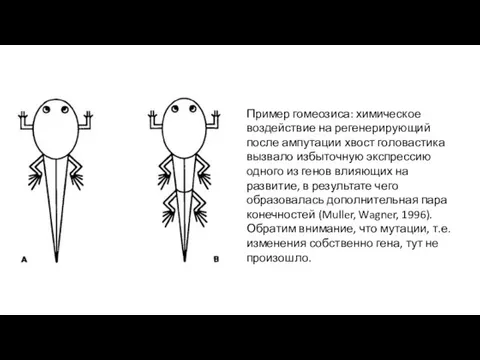
Пример гомеозиса: химическое воздействие на регенерирующий после ампутации хвост головастика вызвало
Пример гомеозиса: химическое воздействие на регенерирующий после ампутации хвост головастика вызвало
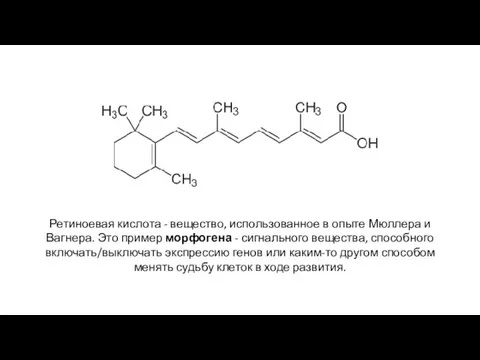
Ретиноевая кислота - вещество, использованное в опыте Мюллера и Вагнера. Это
Ретиноевая кислота - вещество, использованное в опыте Мюллера и Вагнера. Это

В организме часто возникает градиент концентрации морфогена, т.е. ее направленное изменение.
В организме часто возникает градиент концентрации морфогена, т.е. ее направленное изменение.

Рассмотрим модель, в которой:
Концентрация вещества-морфогена линейно растет вдоль тела животного.
Есть гены
Рассмотрим модель, в которой:
Концентрация вещества-морфогена линейно растет вдоль тела животного.
Есть гены
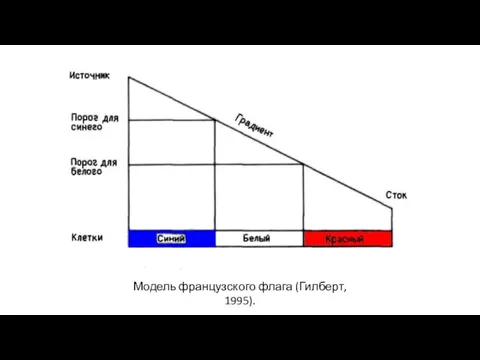
Модель французского флага (Гилберт, 1995).
Модель французского флага (Гилберт, 1995).
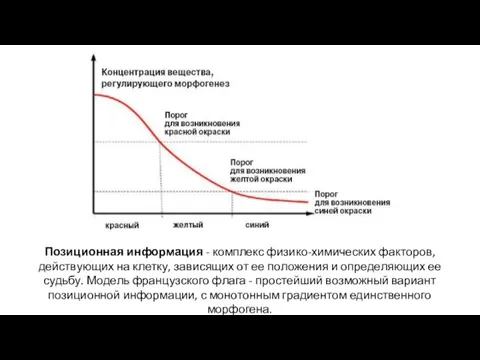
Позиционная информация - комплекс физико-химических факторов, действующих на клетку, зависящих от
Позиционная информация - комплекс физико-химических факторов, действующих на клетку, зависящих от
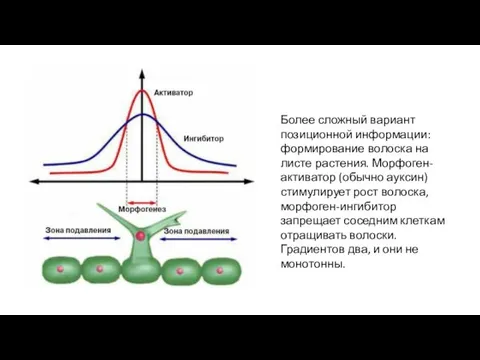
Более сложный вариант позиционной информации: формирование волоска на листе растения. Морфоген-активатор
Более сложный вариант позиционной информации: формирование волоска на листе растения. Морфоген-активатор

Гены развития часто группируются в семейства. Семейство генов - это группа
Гены развития часто группируются в семейства. Семейство генов - это группа

Еще один пример гомеозиса: мутация bithorax, превращающая третий грудной сегмент дрозофилы
Еще один пример гомеозиса: мутация bithorax, превращающая третий грудной сегмент дрозофилы

При отсутствии в области будущего сегмента T3 продукта определенного гена (он
При отсутствии в области будущего сегмента T3 продукта определенного гена (он

Не кроется ли здесь механизм резких эволюционных изменений? Веерокрылые - отряд
Не кроется ли здесь механизм резких эволюционных изменений? Веерокрылые - отряд
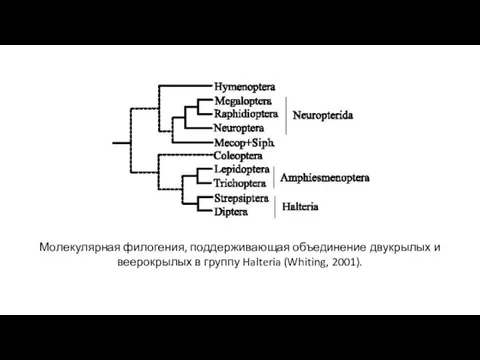
Молекулярная филогения, поддерживающая объединение двукрылых и веерокрылых в группу Halteria (Whiting,
Молекулярная филогения, поддерживающая объединение двукрылых и веерокрылых в группу Halteria (Whiting,
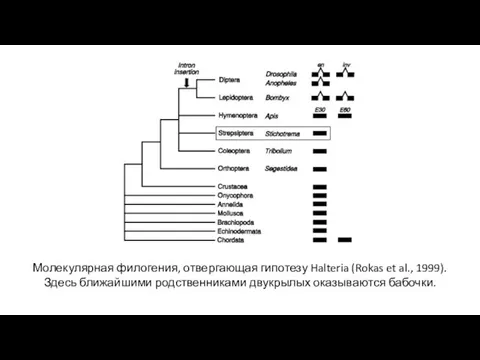
Молекулярная филогения, отвергающая гипотезу Halteria (Rokas et al., 1999). Здесь ближайшими
Молекулярная филогения, отвергающая гипотезу Halteria (Rokas et al., 1999). Здесь ближайшими

Еще одна молекулярная филогения, на которой веерокрылые оказываются родственниками жуков (Wiegmann
Еще одна молекулярная филогения, на которой веерокрылые оказываются родственниками жуков (Wiegmann

Гипотеза Halteria на первый взгляд выглядит убедительной. Однако:
Филогенетические исследования не подтверждают
Гипотеза Halteria на первый взгляд выглядит убедительной. Однако:
Филогенетические исследования не подтверждают
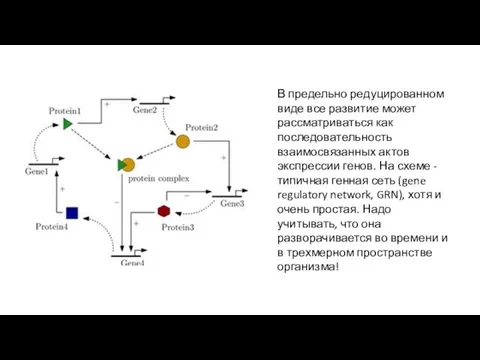
В предельно редуцированном виде все развитие может рассматриваться как последовательность взаимосвязанных
В предельно редуцированном виде все развитие может рассматриваться как последовательность взаимосвязанных
 Вода – колыбель жизни. Жизнь на земле зародилась в воде
Вода – колыбель жизни. Жизнь на земле зародилась в воде Презентация на тему "Классификация моллюсков" - скачать презентации по Биологии
Презентация на тему "Классификация моллюсков" - скачать презентации по Биологии Презентация на тему "Каркас нашего тела" - скачать презентации по Биологии
Презентация на тему "Каркас нашего тела" - скачать презентации по Биологии Рыба в питании человека
Рыба в питании человека Анатомия органов пищеварительного канала
Анатомия органов пищеварительного канала Презентация на тему "Вплив людини на біосферу" - скачать бесплатно презентации по Биологии
Презентация на тему "Вплив людини на біосферу" - скачать бесплатно презентации по Биологии Класс Паукообразные
Класс Паукообразные Презентация на тему "Божественный нектар" - скачать презентации по Биологии
Презентация на тему "Божественный нектар" - скачать презентации по Биологии Бұлшықет жиырылу биофизикасы
Бұлшықет жиырылу биофизикасы Опорно-двигательная система птиц
Опорно-двигательная система птиц Молекулярная биология на стыке веков: ген как структурная и функциональная единица
Молекулярная биология на стыке веков: ген как структурная и функциональная единица Виды нарушения зрения и заболевания глаз у детей
Виды нарушения зрения и заболевания глаз у детей Презентация на тему "Мир вокруг нас. Игровые моменты" - скачать презентации по Биологии
Презентация на тему "Мир вокруг нас. Игровые моменты" - скачать презентации по Биологии Аттестационная работа. Образовательная программа учебного курса в 6 классе Биологическое краеведение
Аттестационная работа. Образовательная программа учебного курса в 6 классе Биологическое краеведение Паразитические черви
Паразитические черви  Физиология дыхательной системы
Физиология дыхательной системы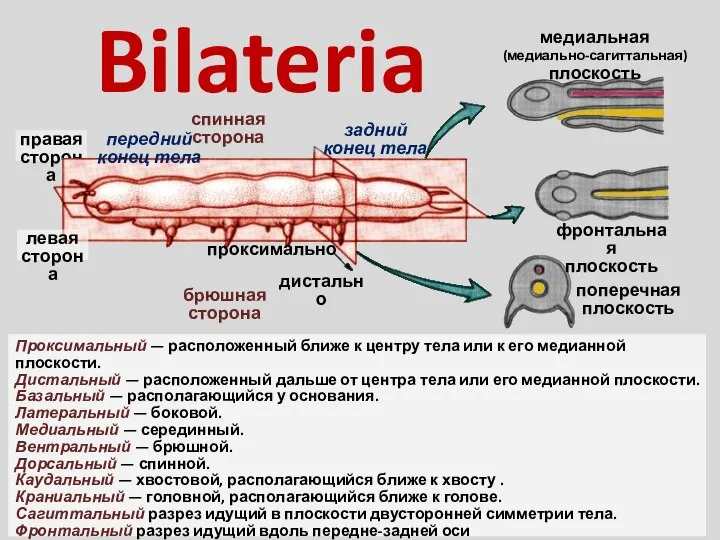 Bilateria
Bilateria Животные и растения, полезные для человека
Животные и растения, полезные для человека Леворукие дети. Некоторые факты из жизни левшей
Леворукие дети. Некоторые факты из жизни левшей Доказательства эволюции животных. 7 класс
Доказательства эволюции животных. 7 класс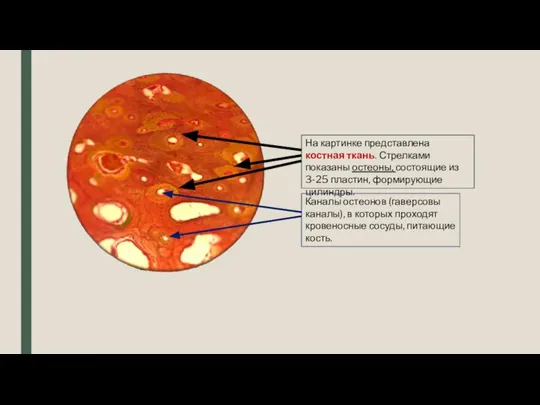 Костная ткань
Костная ткань Поиск и приготовление пищи
Поиск и приготовление пищи Остеология. Состав костной ткани. Строение и классификация костей
Остеология. Состав костной ткани. Строение и классификация костей Клещи
Клещи Цикл трикарбоновых кислот. Глюконеогенез. Обмен гликогена. Лекция № 5
Цикл трикарбоновых кислот. Глюконеогенез. Обмен гликогена. Лекция № 5 Вкусовая сенсорная система
Вкусовая сенсорная система Нервная ткань. Молекулярные механизмы синаптической передачи
Нервная ткань. Молекулярные механизмы синаптической передачи Липидттер. Негізгі сұрақтар
Липидттер. Негізгі сұрақтар