- Физиология слуха и равновесия

Содержание
- 5. Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в перилимфе полукружные каналы, маточка,
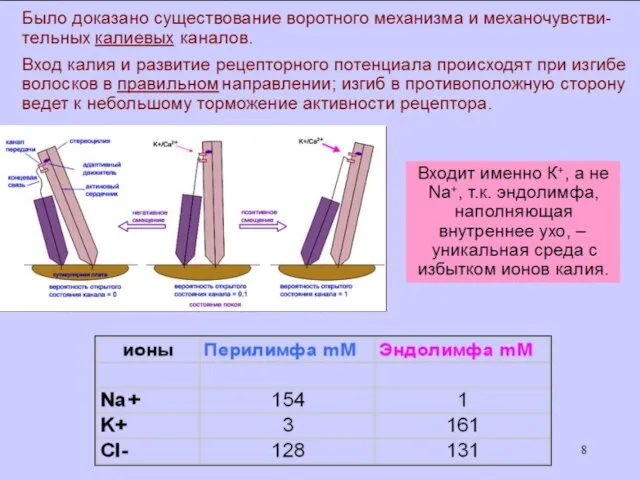
- 6. Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и перилимфе; Концентрация Na+ в
- 7. Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц. Источник эндокохлеарного потенциала — сосудистая
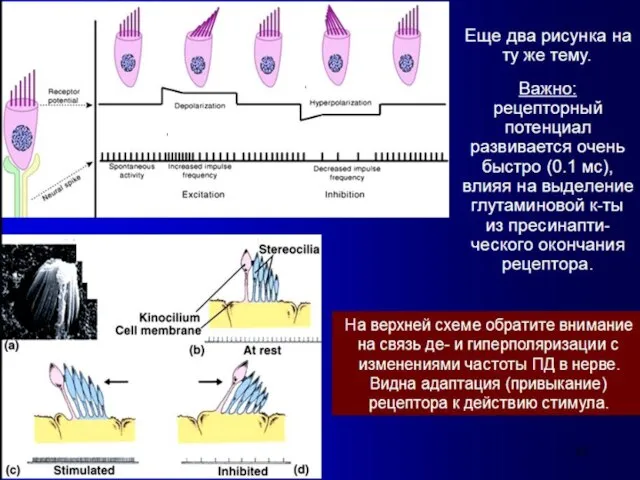
- 8. Последовательность сенсорного преобразования в органе слуха: Собственно рецепторами являются внутренние волосковые клетки. Наружные волосковые клетки, обладая
- 10. Сканирующая электронная микрофотография волосковых клеток кортиева органа Покровная мембрана удалена, а микроскоп направлен на базиллярную мембрану.
- 11. Волосковые клетки
- 15. Высота звука - это субъективное восприятие частоты звуковых колебаний. Человеческое ухо воспринимает частоты в диапазоне от
- 16. 2. Временным способом (20 – 200 Гц) Информация кодируется разной частотой импульсации в одних и тех
- 17. Острота слуха Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев
- 18. Сила звука Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше порога
- 19. Слуховые пути и центры кохлеарное ядро (нейроны 2-го порядка), комплекс ядер верхней оливы и трапецевидного тела
- 20. Направление источника звука определяется 2 путями: При звуке высокой частоты (звук отражается от головы) источник оценивается
- 21. Слуховая кора Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела, в то время как
- 22. Чувство равновесия
- 24. Структуры вестибулярного аппарата
- 25. Полукружные каналы работают в парах (билатерально) При вращение головы вправо: Волосковые клетки с правой стороны гиперполяризуются
- 26. Определение углового ускорения Полукружные каналы Примерно ортогональны друг другу Оба конца заканчиваются в овальном мешочке (место
- 27. Определение линейного ускорения Отолитовые органы: Активный участок – макула – представлена волосковыми клетками, отолитовой мембраной и
- 29. Волосковые клетки При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс ацетилхолина, который и
- 30. Гребень волосков каждого рецептора настроен на «свое» направление ускорения Темная точка - киноцилия. Какой рецептор из
- 31. Передача сигнала в ЦНС Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны которых
- 32. Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений): в спинной мозг в мозжечок в средний
- 33. Вестибулярная информация необходима: Спинному мозгу (от ядра Дейтерса и медиального ядер по латеральному и медиальному вестибулоспинальным
- 35. Укачивание Укачивание - иррадиация возбуждения по центрам ромбовидной ямки. Субъективные ощущения: головокружение, тошнота и другие реакции,
- 36. Вестибулярная система как источник эмоций (новизна ощущений!)
- 37. Обонятельная сенсорная система
- 38. Классификация запахов по Эймуэру Первичные или основные: - камфорный - (камфора, 1,8-цинеол) - острый или едкий
- 39. Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле),
- 40. Втягивание воздуха Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается принюхиванием. Втягивание воздуха —
- 41. Рецепторный аппарат Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно 100 млн рецепторных
- 42. От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек. В волоски встроены обонятельные
- 43. ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток,
- 44. В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы. Эти синаптические
- 45. Регистрация обонятельного сигнала взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков активация G‑белка повышение активности аденилатциклазы
- 46. Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы, но и Ca2+, связывающийся
- 47. Концепция комбинаторного кодирования Каждый из миллионов одорантов имеет уникальный код; кодирование происходит на уровне органа обоняния,
- 48. Пути передачи обонятельной информации Аксоны митральных клеток в составе обонятельного тракта поднимаются в первичные обонятельные центры
- 49. Пути передачи обонятельной информации Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной коре и миндалевидном
- 51. Вкусовая сенсорная система
- 52. Основные вкусы Человек различает четыре первичных вкуса: сладкий, кислый, горький солёный, а также «умами» (от японского
- 53. Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и
- 54. Вкусовые области языка Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к кончику языка,
- 55. Вкусовая почка состоит из удлинённых светлых клеток различных типов. - На апикальной части микроворсинки, занимающие вкусовой
- 56. Различие вкусовых клеток При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь отдельные вкусовые клетки. Некоторые
- 57. Восприятие соленого и кислого Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC), открывая их для натрия.
- 58. Восприятие сладкого Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным к нему рецептором, связанным
- 59. Восприятие горького Горький стимул может деполяризовать мембрану тремя путями: закрытием калиевых каналов, путем взаимодействия с G-белком
- 60. Восприятие умами Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их. Это сопровождается деполяризацией и
- 61. ВКУСОВОЙ ПОРОГ Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов.
- 62. АДАПТАЦИЯ Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации. К сладкому и
- 63. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона - от передних двух
- 64. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 2. нейроны ядра одиночного пути расположены в ростральной
- 65. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 3. Нейроны вентромедиального ядра таламуса направляются к вентральной
- 66. Тактильная сенсорная система
- 67. Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но воспринимаются одними и теми
- 68. Рецепторы кожи Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой чувствительности. Тельца
- 69. Количество и плотность размещения рецепторов - Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных
- 70. Разрешающая способность и пороги ощущений: - Порог ощущения кожных рецепторов при надавливании: 10 мкм - Одновременный
- 71. Адаптация
- 72. Передача импульсации Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам со скоростью 30-70 м/с.
- 73. Пути передачи соматосенсорных сигналов Практически вся сенсорная информация от сегментов тела поступает в спинной мозг через
- 74. Войдя в спинной мозг они направляются: - либо прямо к продолговатому мозгу (лемнисковая система: тонкий, или
- 76. Скачать презентацию
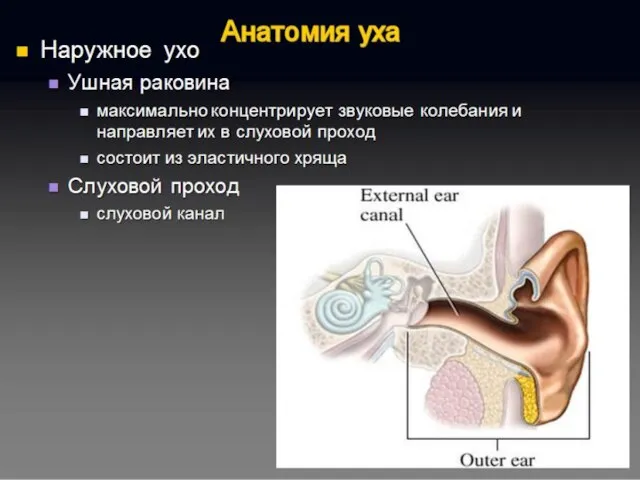
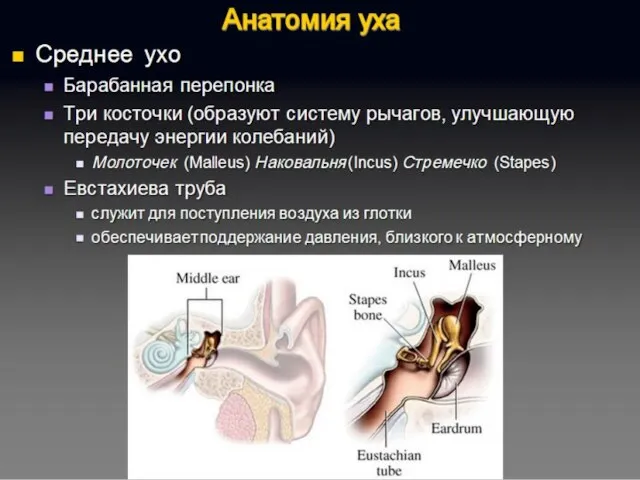
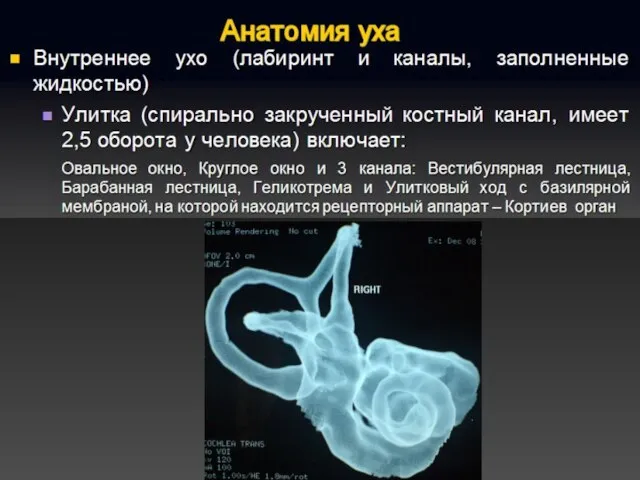
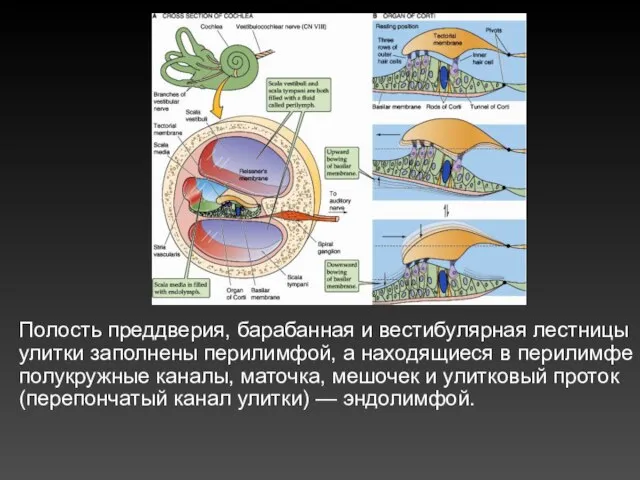
Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся
Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся
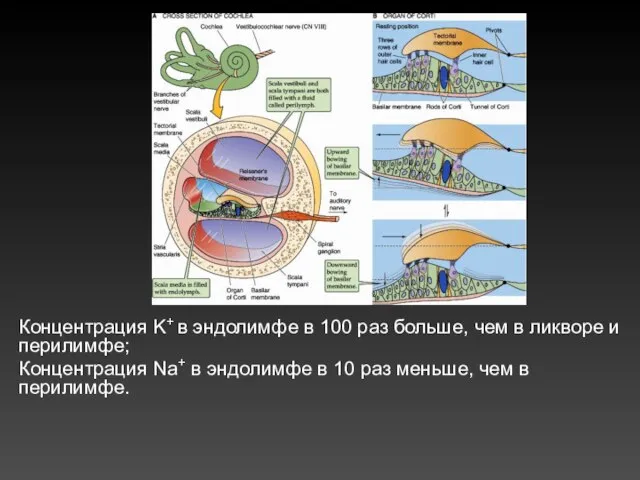
Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе
Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе
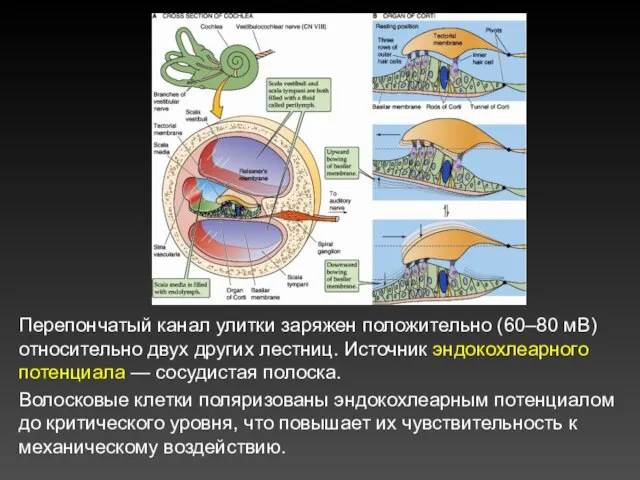
Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц.
Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц.
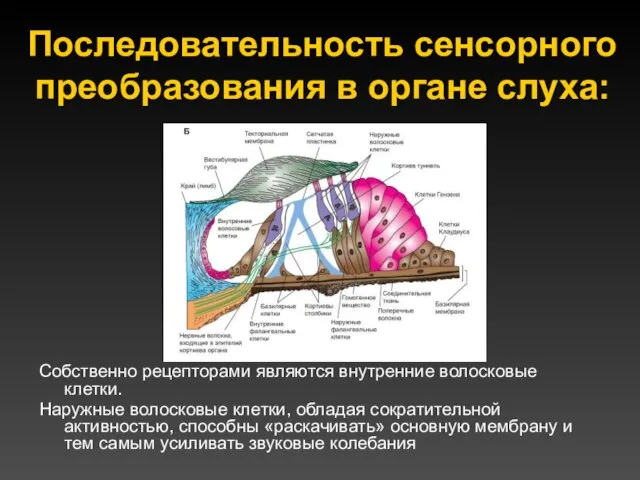
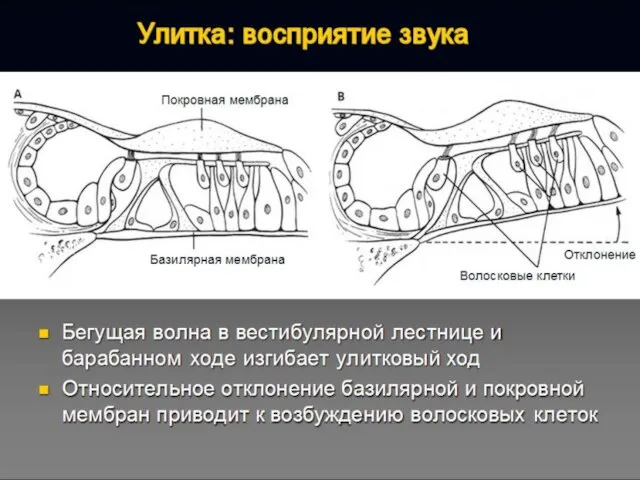
Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые
Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые
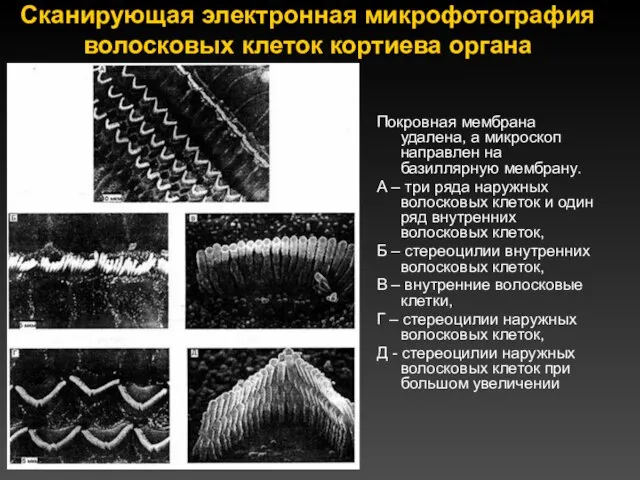
Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп
Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп

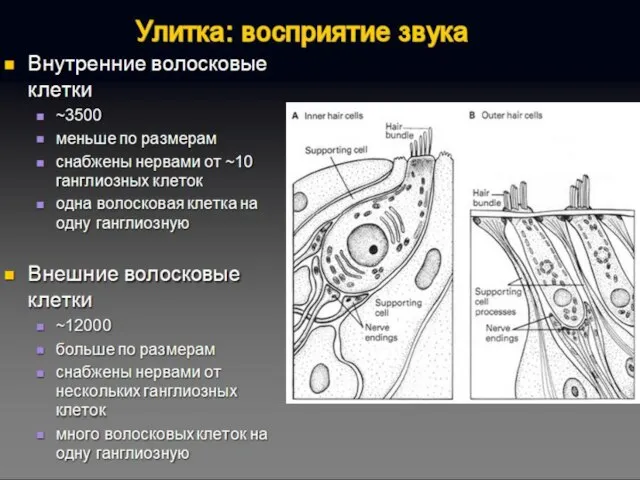
Волосковые клетки
Волосковые клетки

Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
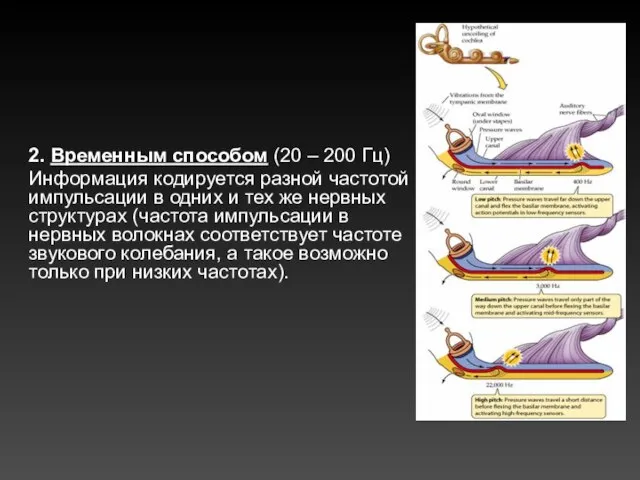
2. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в
2. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в

Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек
Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек

Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз
Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз
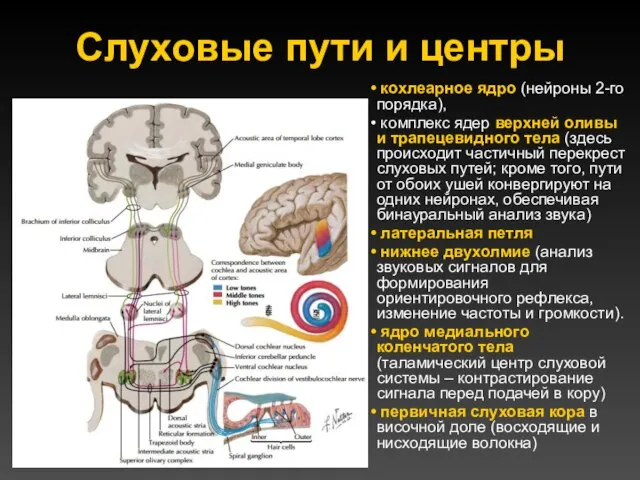
Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс
Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс

Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается
Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается
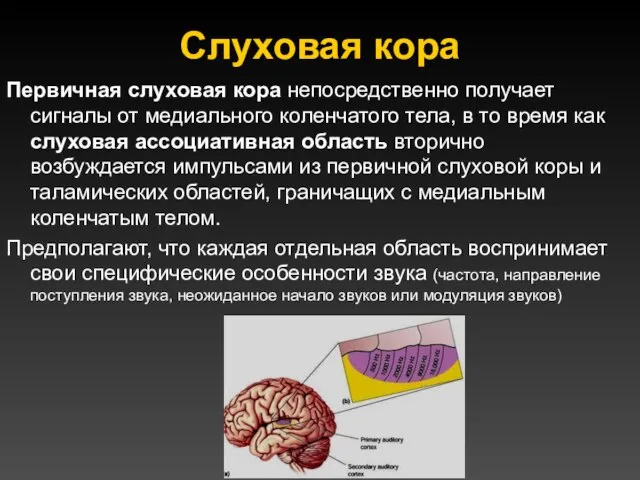
Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела,
Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела,

Чувство равновесия
Чувство равновесия

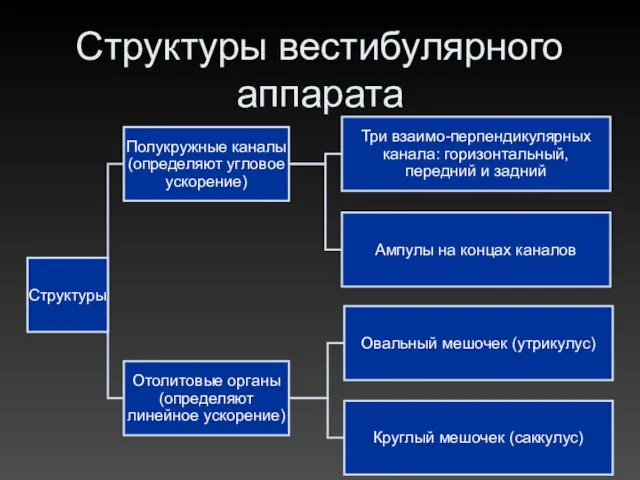
Структуры вестибулярного аппарата
Структуры вестибулярного аппарата

Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с
Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с

Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном
Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном
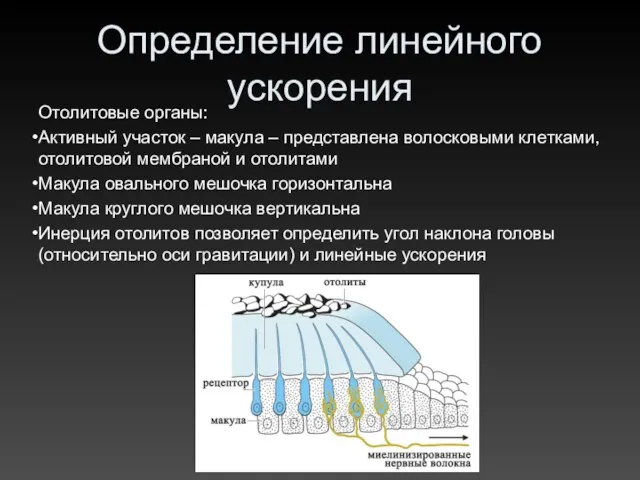
Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками,
Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками,
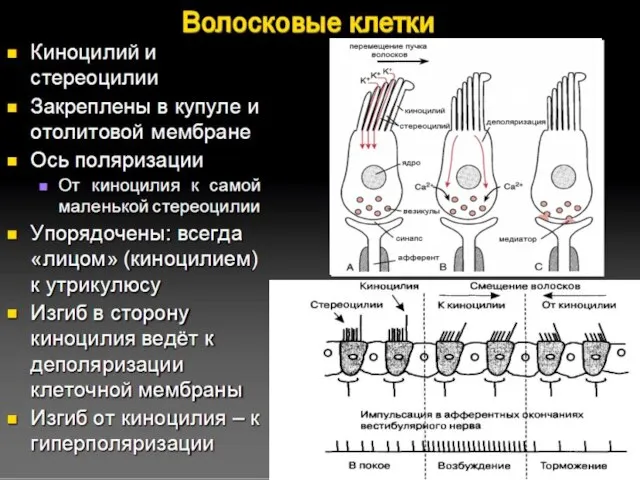
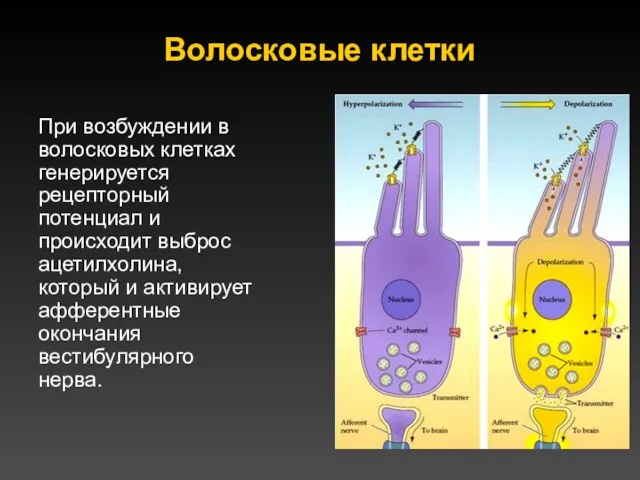
Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит
Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит
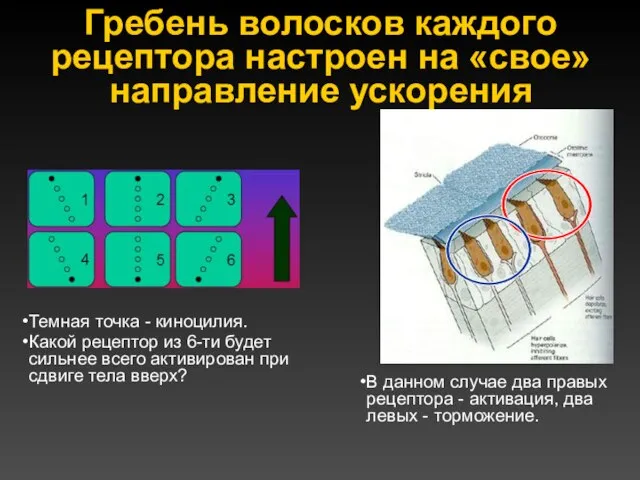
Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка -
Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка -
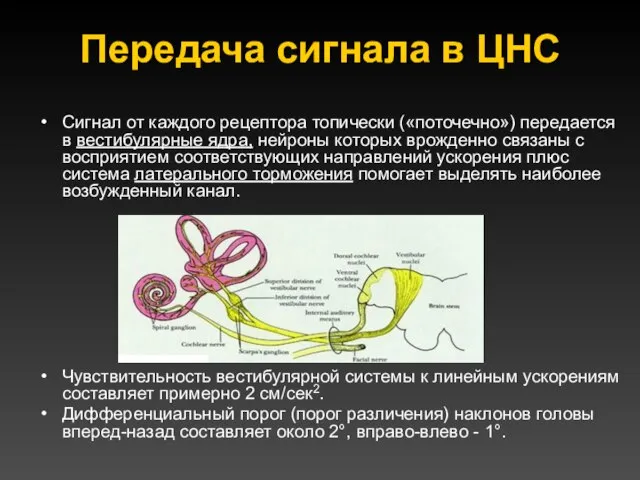
Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в
Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в
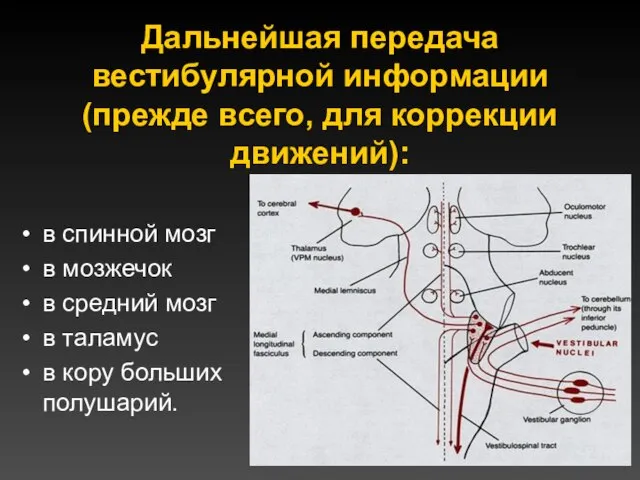
Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг
Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг

Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по
Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по
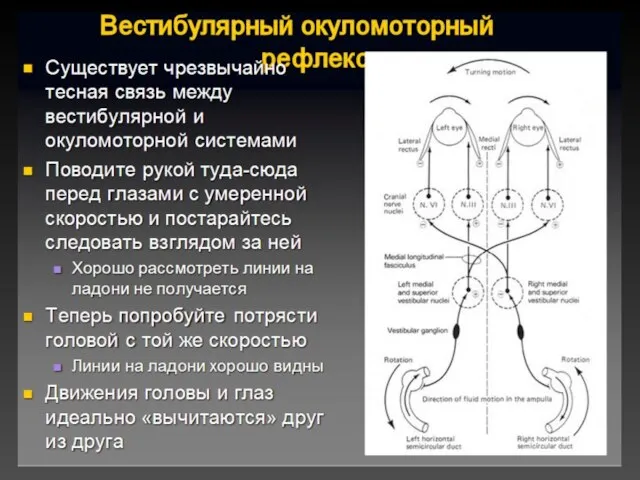
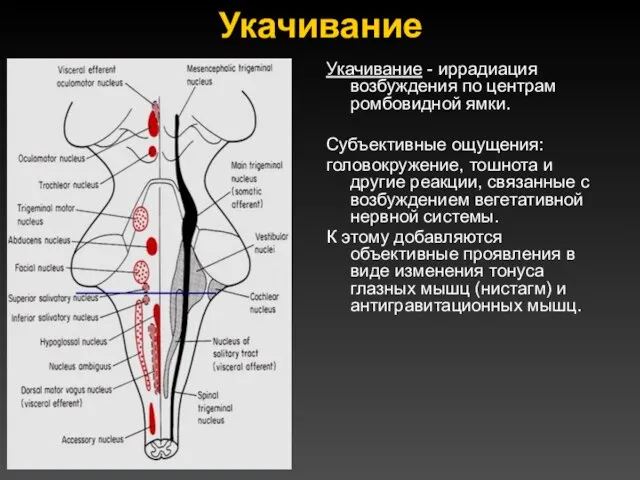
Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и
Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и

Вестибулярная система как источник эмоций
(новизна ощущений!)
Вестибулярная система как источник эмоций
(новизна ощущений!)
Обонятельная сенсорная система
Обонятельная сенсорная система

Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
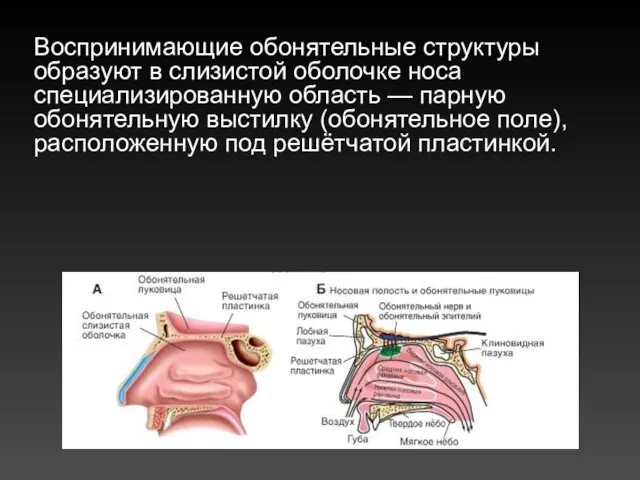
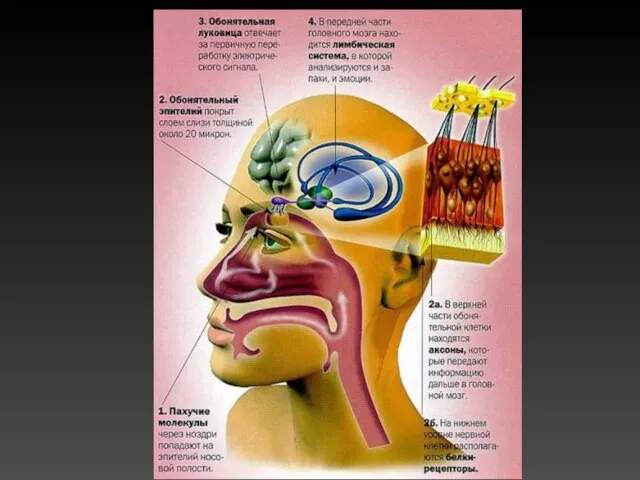
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную

Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается
Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается
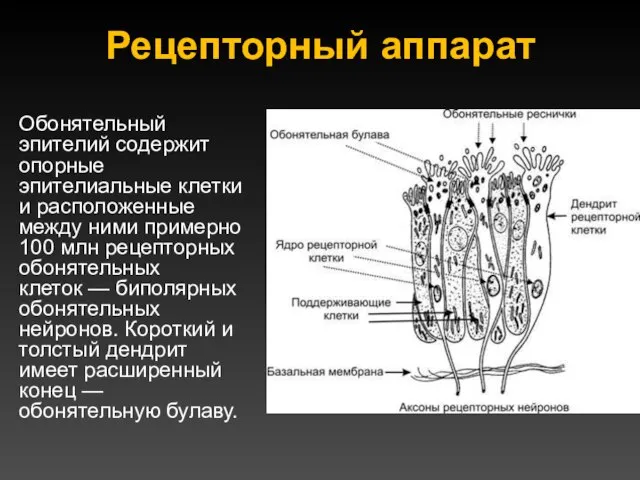
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
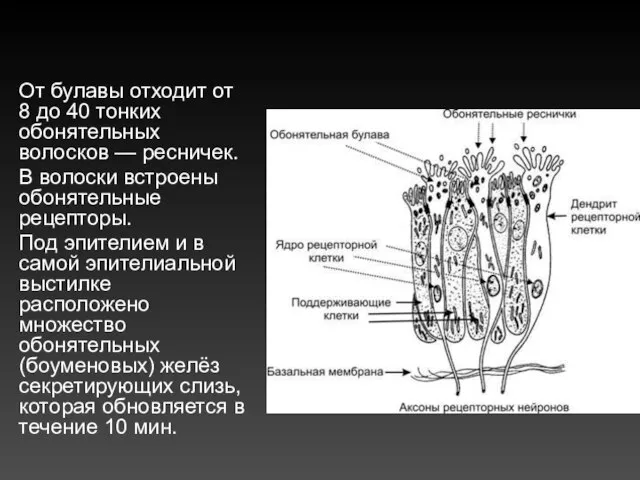
От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.
От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.

ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
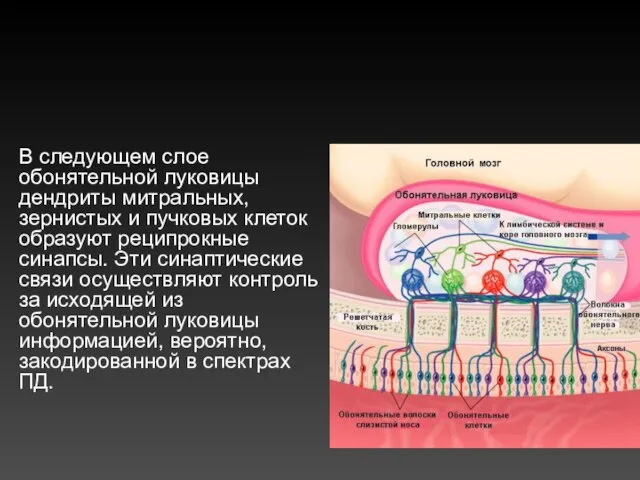
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток

Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация
Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация

Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные
Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные

Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит
Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит

Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются
Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются

Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в
Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в
Вкусовая сенсорная система
Вкусовая сенсорная система

Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также
Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также
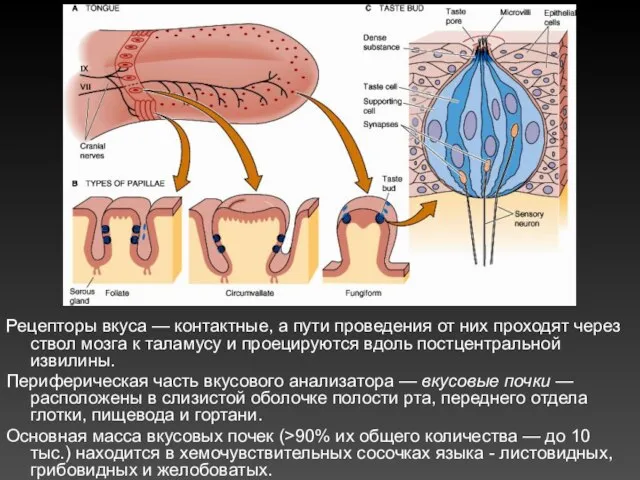
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол
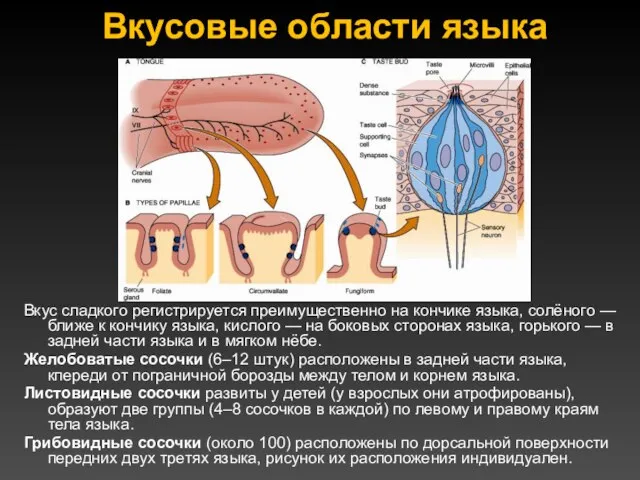
Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе
Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе
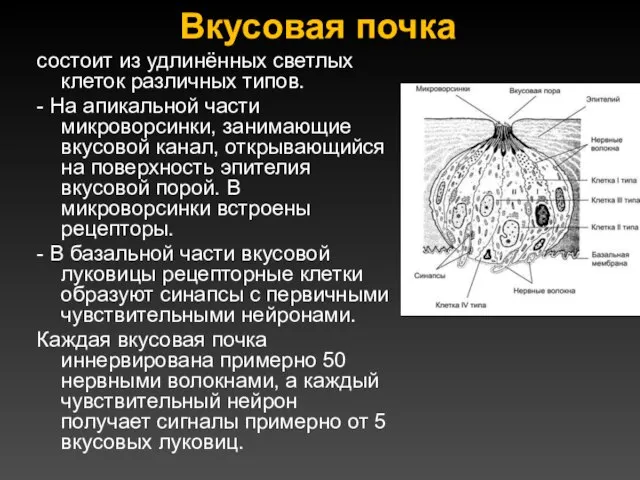
Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной
Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной

Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь
Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь
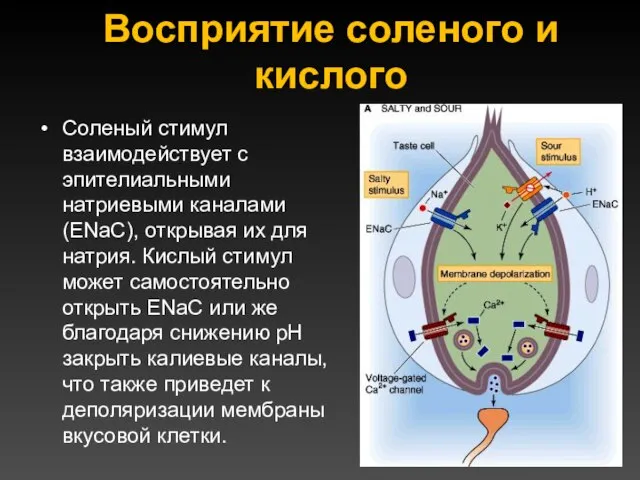
Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC),
Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC),
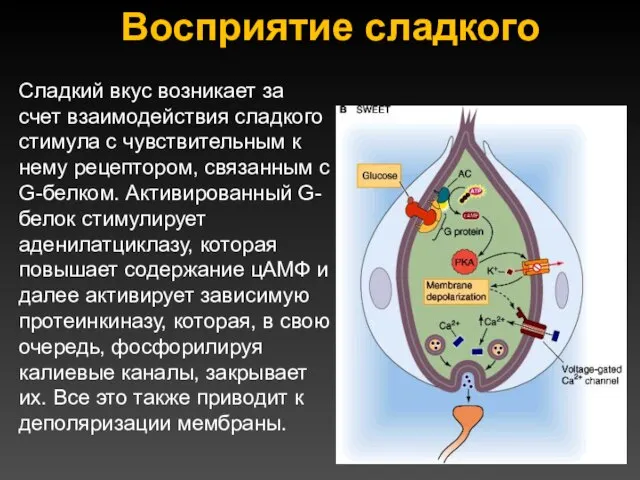
Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным
Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным
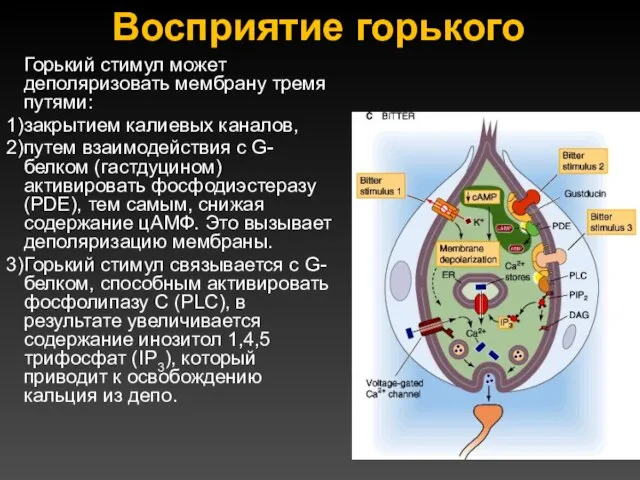
Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
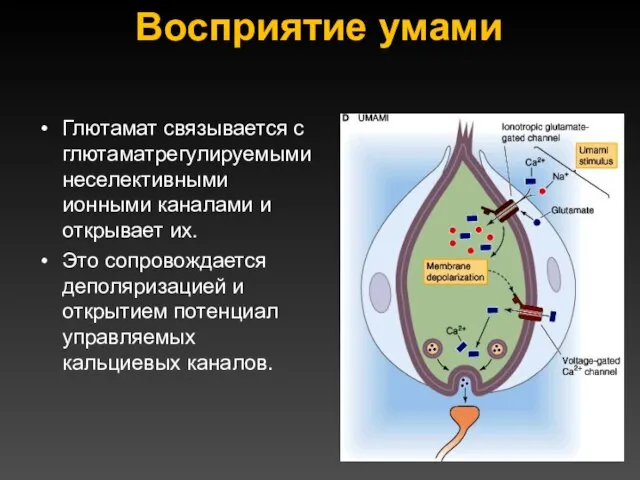
Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.
Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.

ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и
ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и

АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его
АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его
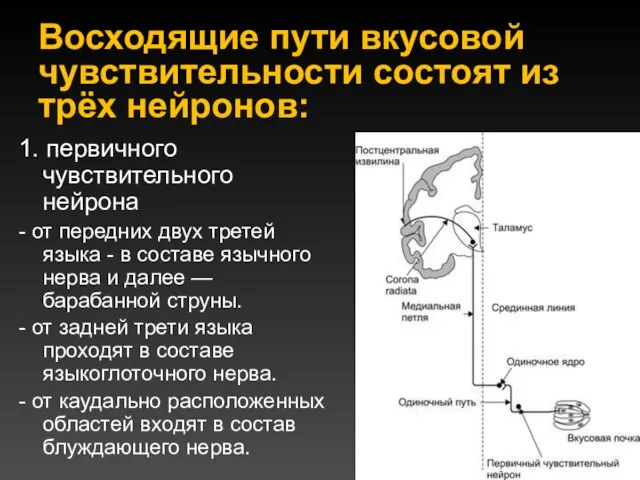
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
-
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
-
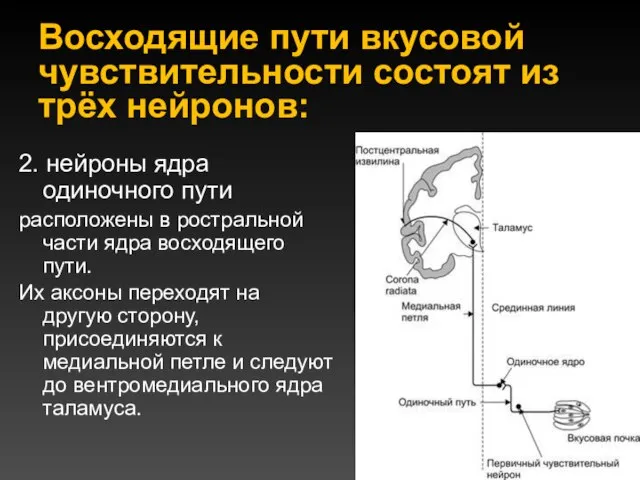
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра

Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра

Тактильная сенсорная система
Тактильная сенсорная система

Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений,
Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений,
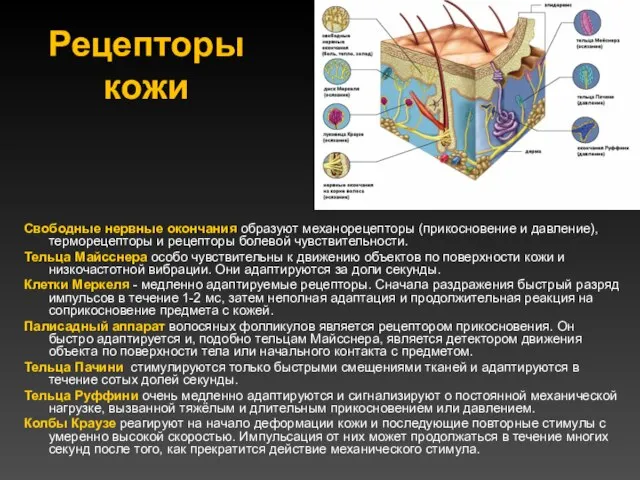
Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы
Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы

Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в
Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в
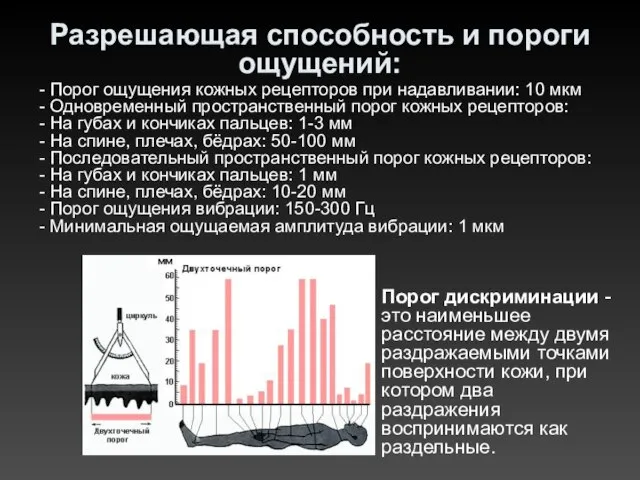
Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании:
Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании:

Адаптация
Адаптация

Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам
Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам
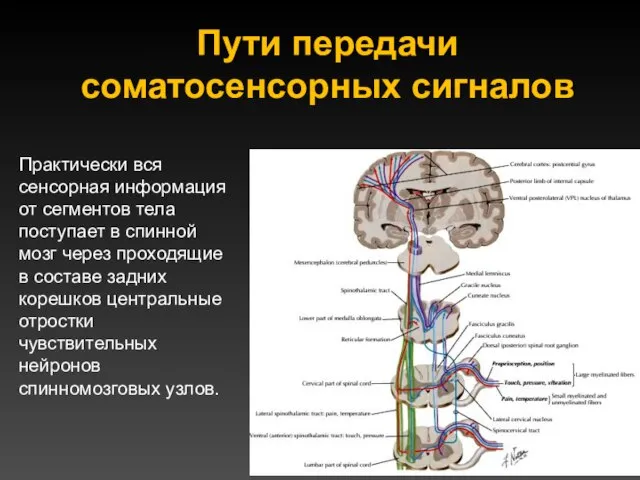
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает
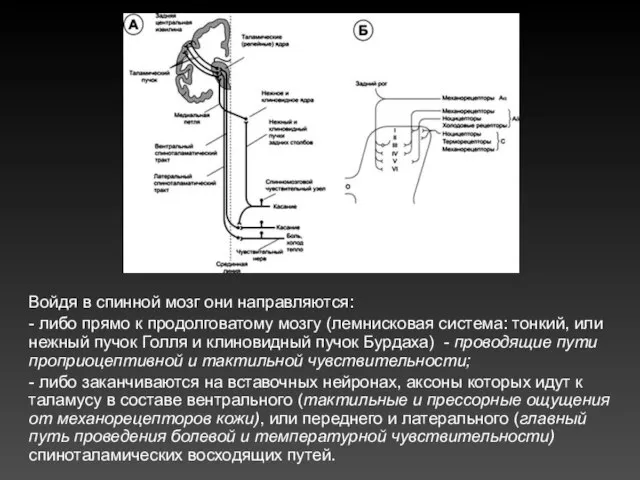
Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу
Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу
 Транспорт веществ в организме животных
Транспорт веществ в организме животных Функциональная система дыхания. Физиология внешнего дыхания
Функциональная система дыхания. Физиология внешнего дыхания Работа учеников 11 А класса Лалыко Екатерины Колесниковой Лизы
Работа учеников 11 А класса Лалыко Екатерины Колесниковой Лизы Влияние дождевых червей на образование почвы
Влияние дождевых червей на образование почвы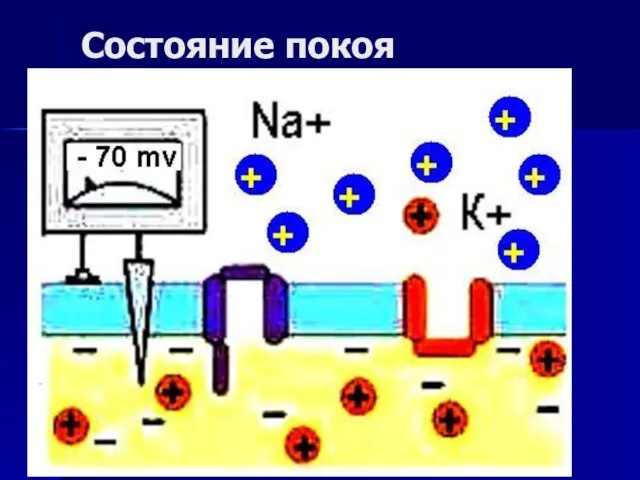 Возбуждение клетки. Переход от покоя к активному состоянию
Возбуждение клетки. Переход от покоя к активному состоянию Витамины и их роль в жизни людей
Витамины и их роль в жизни людей Наследственные болезни человека
Наследственные болезни человека Биохимия минерализованных тканей полости рта
Биохимия минерализованных тканей полости рта Neurology. Spinal cord compression
Neurology. Spinal cord compression Митоз
Митоз Изучение растений Красной книги Кубани
Изучение растений Красной книги Кубани Система виділення. Фізіологія нирки
Система виділення. Фізіологія нирки Мутации. Структура гена, общая характеристика генома человека
Мутации. Структура гена, общая характеристика генома человека Нервная система человека
Нервная система человека Путешествие в мир природы. Планета растений. Планета животных
Путешествие в мир природы. Планета растений. Планета животных Биогеоценоз и экосистема
Биогеоценоз и экосистема История развития биологии. Значение биологии в жизни человека и общества
История развития биологии. Значение биологии в жизни человека и общества Кто живёт в земле
Кто живёт в земле История
История Жануарлар психологиясының белсенділігін сипаттау
Жануарлар психологиясының белсенділігін сипаттау Общая характеристика цестод дифиллоботриоз плотоядных. Дипилидиоз плотоядных
Общая характеристика цестод дифиллоботриоз плотоядных. Дипилидиоз плотоядных СЕЛЕКЦИЯ Выполнила: Козодерова Анна Ученица 11 класса «А» Проверила: Шарова М.А.
СЕЛЕКЦИЯ Выполнила: Козодерова Анна Ученица 11 класса «А» Проверила: Шарова М.А. «Использование ИКТ на уроках биологии в целях повышения качества знаний обучающихся» Учитель биологии шл №27 г.Астана Матковск
«Использование ИКТ на уроках биологии в целях повышения качества знаний обучающихся» Учитель биологии шл №27 г.Астана Матковск Взаимодействие генов в генотипе
Взаимодействие генов в генотипе Минеральная вода Презентация для потребителей минеральной воды_
Минеральная вода Презентация для потребителей минеральной воды_ Урок бабочки
Урок бабочки Удивительный лимон
Удивительный лимон КОЛЬЧАТЫЕ ЧЕРВИ. ДОЖДЕВОЙ ЧЕРВЬ Нет сомнения, что вряд ли есть еще другие животные, которые сыграли бы столь важную роль в ист
КОЛЬЧАТЫЕ ЧЕРВИ. ДОЖДЕВОЙ ЧЕРВЬ Нет сомнения, что вряд ли есть еще другие животные, которые сыграли бы столь важную роль в ист