- Онковіруси. Історія. Загальні положення

Содержание
- 2. Вперше вірусну природу пухлин продемонстрували: Еритробластоз курей (Ellerman та Bang, 1908) саркома курчат (Rous, 1911) Rockefeller
- 3. РАК – інфекційна хвороба? 1909р. Пейтон Раус відкрив саркомо-індукуючий агент у курчат
- 4. Fujinami тa Inamoto в 1914р. показали, що віруси індукують саркоми у птахів; Через ~30 років у
- 5. Основні положення Зільбер Л.О. 1945 Віруси можуть викликати пухлини у ссавців Онкогенні віруси частіше встановлюють персистентну
- 6. Віруси рідко бувають повністю канцерогенними Між первинною інфекцією та появою пухлин – тривалий латентний період Штами
- 7. Інфекційний Oнкогенний кореляція між високою захворюваністю певним типом раку та високою інфікованістю населення певним типом вірусу
- 8. Загальні властивості онкогенних вірусів 1) віруси лише ініціюють патологічний процес, посилюючі проліферацію та генетичну нестабільність інфікованих
- 9. Як віруси трансформують клітини ? Вірусна інфекція підвищує ризик розвитку раку Діє як “мутаген” Інші ко-фактори
- 10. Докази вірусного походження пухлин Присутність частини вірусного геному в пухлині та експресія деяких вірусних генів Інфекція
- 11. Фактори, що сприяють трансформації клітин вірусами Абортивна інфекція (переривання повного циклу репродукції) інфекція повним інфекційним вірусом
- 12. На тепер відомо більше 150 онкогенних вірусів, які поділяються на великі групи: РНК-геномні віруси активують онкогени
- 13. Таксономія онкогенних вірусів DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses RNA viruses: retroviruses flaviviruses
- 14. Віруси людей , що асоціюються з трансформацією HPV 16, 18, 31, 33, 45 Рак шийки матки
- 15. ОНКОГЕННІ РНК ВІРУСИ
- 16. Alpharetrovirus: вірус лейкозу птахів, морфологічний тип-C; (B)Betaretrovirus:вірус раку молочних залоз мишей морфологічний тип-B; (C) Gammaretrovirus: вірус
- 17. Морфологічні типи тип А – розмір 70-75 нм, ендогенні, дефектні, мають вигляд електроннощільних шарів локалізуються всередині
- 18. "Молоді" ендогенні ретровіруси Ендогенізація екзогенних ретровірусів – через проникнення в зародкову лінію. Процес має випадковий характер
- 19. •більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази • Вони мають ген env , вірусоподіні частки
- 20. тип В - розміром 100-160 нм, мають ексцентрично розміщений електроннощільний нуклеокапсид, досить довгі пепломери, типовий представник
- 21. Біологічні властивості РВ 1.Різний ступінь онкогенності для природних хазяїв Високий - для вірусів саркоматозно-лейкозного комплексу; помірний
- 22. За ступенем онкогенності: Гостротрансформуючі ретровіруси (віруси гострих лейкозів у птахів і мишей та саркоми Рауса у
- 23. Біологічні властивості РВ 2.Здатність долати тканинні, видові та класові бар’єри, що особливо притаманно вірусам типу С
- 24. Онкогенні РНК віруси
- 25. Формування віріонів
- 26. Будова віріону
- 27. Онкогенні віруси РНК : Диплоїдна має cap- та poly-А Позитивної полярності Вірусна РНК не може транслюватись,
- 28. Вірусний геном (oncornaviruses) LTR LTR GAG POL ENV Long terminal repeat Group specific antigens (нуклеокапсид) РНК-залежна
- 29. Зворотна Транскриптаза 1970 р. Д. Балтімор, С. Міцутані та Г. Темін виявили зворотну транскриптазу в частинках
- 30. Вірусна РНК Зворотна транскриптаза R U5 U3 R U3 R U5 U3 R U5 Утворились LTR
- 31. Онкогенні РНК віруси головне: відмінність двох форм РНК R U5 GAG POL ENV U3 R
- 32. Для вирішення питання про природу включення геному вірусу в ДНК клітини необхідно відповісти на питання: коли,
- 33. Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий 3’вкорочений кінець З”єднання 3’ кінців
- 34. Онкогенні РНК віруси Батьківська РНК РНК/ДНК гібрид Лінійний DНК/DНК дублекс Циркулярний дублекс DНК Інтеграція Реплікація Транскрипція
- 35. Replicative cycle
- 36. РАК та РЕТРОВІРУСИ
- 37. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 38. http://www.blc.arizona.edu/marty/411/Modules/Lectures/Figures/Trans_Retros.GIF Більшість природних онкогенних вірусів дефектні: Без вірусу-помічника не можуть розмножуватись (не дефектний) (дефектний, без Pol
- 39. R U5 GAG POL ENV U3 R Деякі Ретровіруси мають додатковий ген “типовий ретровірус” SRC
- 40. Вірус саркоми птахів (FSV) R U5 dGAG FMS dENV U3 R Вірус мієлоцитозу птахів (MC29) R
- 41. Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу Клітинні онкогени можуть індукувати трансформацію тільки
- 42. вірус саркоми Рауса є результатом рекомбінацій між c-src і давнім стандартним ретровірусом птахів. функції нормальних генів
- 43. Структура дефектного РВ
- 44. Гостро трансформуючі ретровіруси src Rous sarcoma virus Chicken myc Avian myelocytomatosis virus Chicken erb A, erb
- 46. Трансдукуючі ретровіруси Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей (виняток RSV= src- oнкоген)
- 47. Будова та локалізація pp60-src
- 48. Активація src-гену
- 49. SRC Окисний стрес Мітоз ФР АГ Цитокіни Позаклітинний матрикс Мітотичні функції Стрес. механізм RAS MYC Реорганізація
- 50. Наслідки ретровірусної трансдукції “Простий удар” канцерогенезу (одна подія) Поліклональна : пухлинний ріст починається в кожній інфікованій
- 51. Характеристика вірусних онкогенів (Virology 2000)
- 52. Трансдуковні гени – це гени сигнального шляху Зовнішні сигнальні молекули або фактори росту (sis) Клітинні рецептори
- 53. C-SRC V-SRC
- 54. Мутації протоонкогенів Ras характерні для пухлин людини Мутації в кодонах 12, 13, 61 генів K-RAS, N-RAS,
- 55. Мутації протоонкогенів Ras викликають перманентну стимуляцію розмноження неопластичних клітин Ras Ras ГДФ ГТФ SOS GAP Ras
- 56. Erb B Epidermal growth factor receptor Трансдукований вірусом еритробластозу птахів та фібросаркоми птахів P P P
- 57. 1. Sis – фактор росту тромбоцитів (PDGF) кДНК диких мавп та котів викликає саркому Трансформовані клітини
- 58. інтегрини (a5b1 и др) Рецептори мітогенів/мотогенів (Met.) PI3K Raf ERK Rho Ras MEK Rac PAK MEKK
- 59. За функціональною активністю онкогени можна розділити на групи 1.а) з тирозинкіназною активністю : abl, erb B,
- 60. Онкогени як сигнали трансдукції Фактор росту v-sis, int-1, int-2, hst, fgf-5 Рецептори фактору росту v-erb-B, v-fms,
- 61. Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою онкогенів: -«кількісна» - збільшення кількості
- 63. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 64. Cis-активуючі ретровіруси Не мають онкогена Зберігають всі вірусні гени Компетентні за реплікацією
- 65. Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду Немає онкогена! – Яким чином утворюється
- 66. Пухлина походить з однієї клітини - моноклональна ALV може інтегруватись в геном клітини БAГАТЬОХ місцях Але
- 67. Чи специфічні ці сайти? Вірус Myelocytoma виділений від різних птахів завжди знаходять в одному й тому
- 68. Як C-onc залучаються в не вірусний канцерогенез?
- 69. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 70. Протоонкогени та перебудова хромомсом Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc. інсерція промотора Ig тяжкого
- 71. Механізм клітинної трансфорації cis-активуючих ретровірусів Випадкова ретровірусна інтеграція в клітинний геном Інсерційна активація (або дезактивація) с-onc
- 73. Наслідки активації онкогену при інсерції Ретровірусів Клітинна трансформація відбувається завдяки інсерції поблизу потенційного онкогена Пухлини моноклональні:
- 74. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 75. Human T cell Leukemia Virus type I (HTLV-I) Асоційований з 2 фатальними хворобами людей Т-клітинний лейкоз
- 76. HTLV-I та ATL 1980 Галло ізолював тип C retrovirus (HTLV1) від хворого з “різкою T клітинною
- 77. Міжклітинний контакт Статевий (60% від чоловіка до жінки проти 1% від жінки до чоловіка) Продуктами крові
- 78. Онкогенез, опосередкований важливим ретровірусним білком Виключення з парадігми ретровірусного онкогенезу (HTLV-1) HTLV-1 не має послідовностей клітинного
- 79. HTLV-I genome 9 kilobase РНК геном Унікальні регуляторні білки Tax та Rev Важливі для вірусної реплікації
- 80. Tax тa Oнкогенез Tax необхідний для вірусної реплікації 40kda фосфопротеїн Tранскрипційний активатор геному HTLV-I Взаємодія з
- 81. Tax – множинний трансактиватор Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію з клітинними промоторами Дисоціює
- 82. Білки регулятори клітинного циклу – мішені дії Tax Iнактивація p53 (G1/S контроль рестрикції) Активує циклін D,
- 83. Tax репресує DНК -репарацію Репресія DНК pol β , яка залучена в репарацію основ та нуклеотидів
- 84. Механізми клітинної трансформації ретровірусами
- 85. Онкогенні РНК- віруси
- 87. Скачать презентацию

Вперше вірусну природу пухлин продемонстрували:
Еритробластоз курей (Ellerman та Bang,
Вперше вірусну природу пухлин продемонстрували:
Еритробластоз курей (Ellerman та Bang,
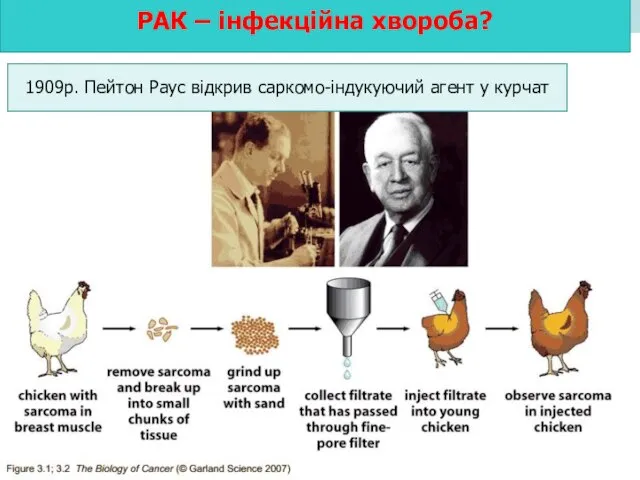
РАК – інфекційна хвороба?
1909р. Пейтон Раус відкрив саркомо-індукуючий агент у
РАК – інфекційна хвороба?
1909р. Пейтон Раус відкрив саркомо-індукуючий агент у

Fujinami тa Inamoto в 1914р. показали, що віруси індукують саркоми
Fujinami тa Inamoto в 1914р. показали, що віруси індукують саркоми

Основні положення
Зільбер Л.О. 1945
Віруси можуть викликати пухлини у ссавців
Онкогенні віруси
Основні положення
Зільбер Л.О. 1945
Віруси можуть викликати пухлини у ссавців
Онкогенні віруси

Віруси рідко бувають повністю канцерогенними
Між первинною інфекцією та появою пухлин –
Віруси рідко бувають повністю канцерогенними
Між первинною інфекцією та появою пухлин –

Інфекційний Oнкогенний
кореляція між високою захворюваністю певним типом раку та високою інфікованістю
Інфекційний Oнкогенний
кореляція між високою захворюваністю певним типом раку та високою інфікованістю

Загальні властивості онкогенних вірусів
1) віруси лише ініціюють патологічний процес, посилюючі проліферацію
Загальні властивості онкогенних вірусів
1) віруси лише ініціюють патологічний процес, посилюючі проліферацію

Як віруси трансформують клітини ?
Вірусна інфекція підвищує ризик розвитку раку
Як віруси трансформують клітини ?
Вірусна інфекція підвищує ризик розвитку раку

Докази вірусного походження пухлин
Присутність частини вірусного геному в пухлині та
Докази вірусного походження пухлин
Присутність частини вірусного геному в пухлині та

Фактори, що сприяють трансформації клітин вірусами
Абортивна інфекція (переривання повного циклу репродукції)
інфекція
Фактори, що сприяють трансформації клітин вірусами
Абортивна інфекція (переривання повного циклу репродукції)
інфекція

На тепер відомо більше 150 онкогенних вірусів, які поділяються на великі
На тепер відомо більше 150 онкогенних вірусів, які поділяються на великі

Таксономія онкогенних вірусів
DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses
RNA
Таксономія онкогенних вірусів
DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses
RNA

Віруси людей , що асоціюються з трансформацією
HPV 16, 18, 31,
Віруси людей , що асоціюються з трансформацією
HPV 16, 18, 31,

ОНКОГЕННІ РНК ВІРУСИ
ОНКОГЕННІ РНК ВІРУСИ
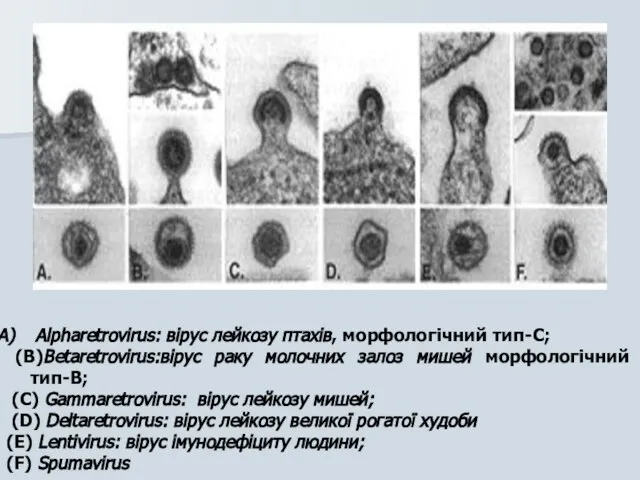
Alpharetrovirus: вірус лейкозу птахів, морфологічний тип-C;
(B)Betaretrovirus:вірус раку молочних залоз мишей
(B)Betaretrovirus:вірус раку молочних залоз мишей

Морфологічні типи
тип А –
розмір 70-75 нм,
ендогенні,
дефектні,
мають
Морфологічні типи
тип А –
розмір 70-75 нм,
ендогенні,
дефектні,
мають

"Молоді" ендогенні ретровіруси
Ендогенізація екзогенних ретровірусів – через проникнення в зародкову лінію.
"Молоді" ендогенні ретровіруси
Ендогенізація екзогенних ретровірусів – через проникнення в зародкову лінію.
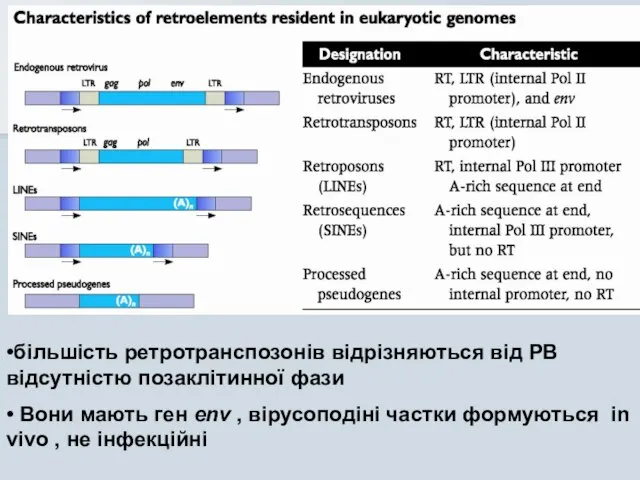
•більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази
• Вони мають ген
•більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази
• Вони мають ген

тип В - розміром 100-160 нм, мають ексцентрично розміщений електроннощільний нуклеокапсид,
тип В - розміром 100-160 нм, мають ексцентрично розміщений електроннощільний нуклеокапсид,

Біологічні властивості РВ
1.Різний ступінь онкогенності для природних хазяїв Високий - для
Біологічні властивості РВ
1.Різний ступінь онкогенності для природних хазяїв Високий - для

За ступенем онкогенності:
Гостротрансформуючі ретровіруси
(віруси гострих лейкозів у птахів і мишей
За ступенем онкогенності:
Гостротрансформуючі ретровіруси
(віруси гострих лейкозів у птахів і мишей

Біологічні властивості РВ
2.Здатність долати тканинні, видові та класові бар’єри, що особливо
Біологічні властивості РВ
2.Здатність долати тканинні, видові та класові бар’єри, що особливо
Онкогенні РНК віруси
Онкогенні РНК віруси
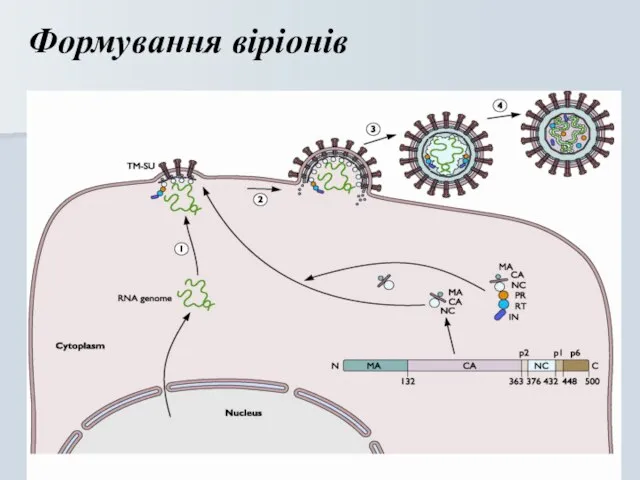
Формування віріонів
Формування віріонів
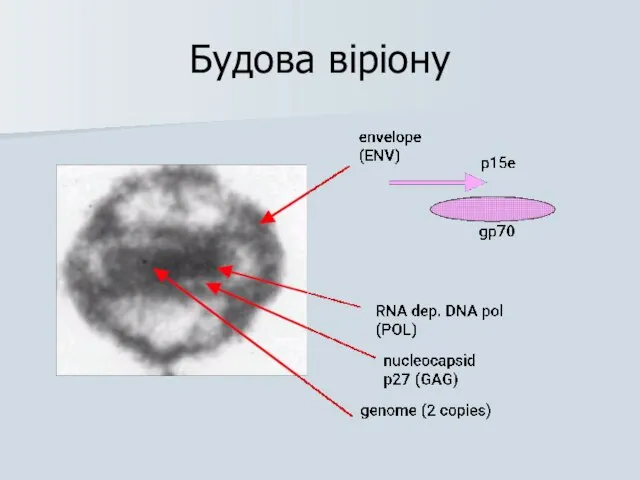
Будова віріону
Будова віріону

Онкогенні віруси
РНК :
Диплоїдна має cap- та poly-А
Позитивної полярності
Вірусна РНК
Онкогенні віруси
РНК :
Диплоїдна має cap- та poly-А
Позитивної полярності
Вірусна РНК
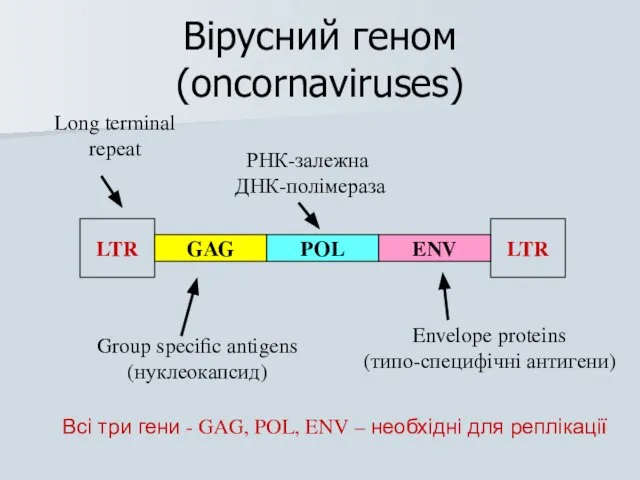
Вірусний геном (oncornaviruses)
LTR
LTR
GAG
POL
ENV
Long terminal
repeat
Group specific antigens
(нуклеокапсид)
РНК-залежна
ДНК-полімераза
Envelope proteins
(типо-специфічні антигени)
Всі три гени
Вірусний геном (oncornaviruses)
LTR
LTR
GAG
POL
ENV
Long terminal
repeat
Group specific antigens
(нуклеокапсид)
РНК-залежна
ДНК-полімераза
Envelope proteins
(типо-специфічні антигени)
Всі три гени

Зворотна Транскриптаза
1970 р.
Д. Балтімор, С. Міцутані та Г. Темін виявили
Зворотна Транскриптаза
1970 р.
Д. Балтімор, С. Міцутані та Г. Темін виявили
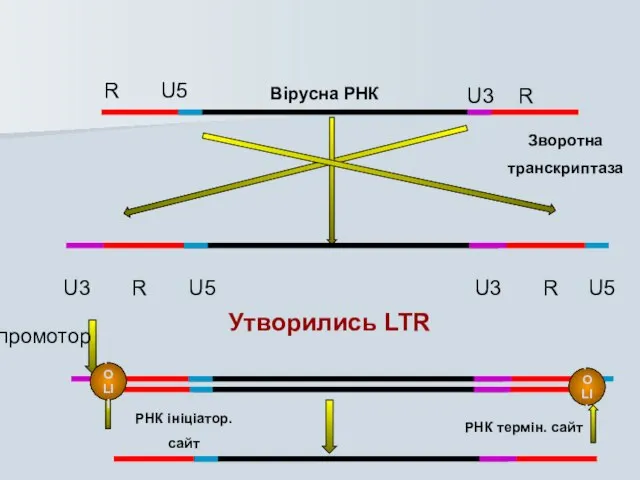
Вірусна РНК
Зворотна
транскриптаза
R U5
U3 R
U3 R U5
U3 R U5
Утворились LTR
POLII
Вірусна РНК
Зворотна
транскриптаза
R U5
U3 R
U3 R U5
U3 R U5
Утворились LTR
POLII
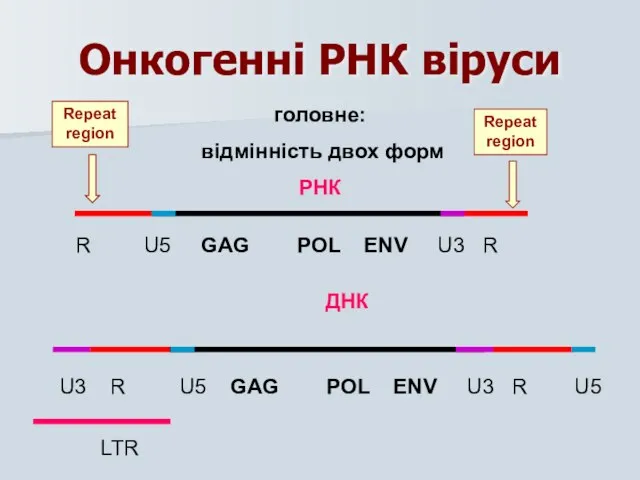
Онкогенні РНК віруси
головне:
відмінність двох форм
РНК
R U5 GAG POL ENV U3
Онкогенні РНК віруси
головне:
відмінність двох форм
РНК
R U5 GAG POL ENV U3

Для вирішення питання про природу включення геному вірусу в ДНК клітини
Для вирішення питання про природу включення геному вірусу в ДНК клітини
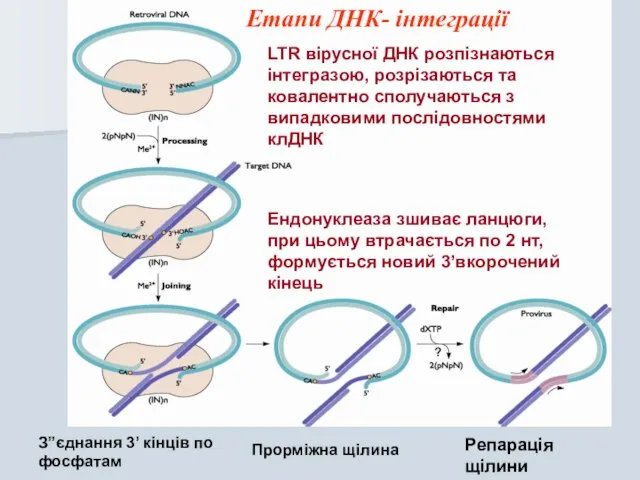
Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий
Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий
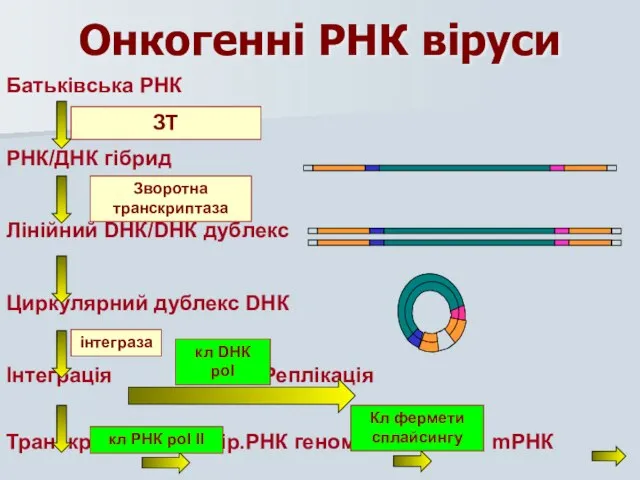
Онкогенні РНК віруси
Батьківська РНК
РНК/ДНК гібрид
Лінійний DНК/DНК дублекс
Циркулярний дублекс DНК
Інтеграція Реплікація
Транскрипція
Онкогенні РНК віруси
Батьківська РНК
РНК/ДНК гібрид
Лінійний DНК/DНК дублекс
Циркулярний дублекс DНК
Інтеграція Реплікація
Транскрипція
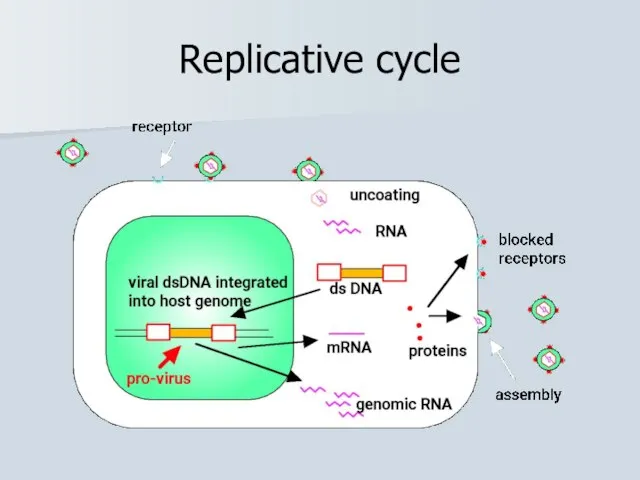
Replicative cycle
Replicative cycle
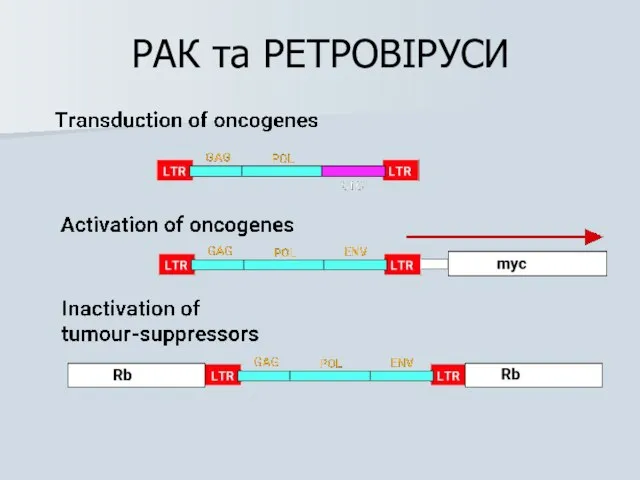
РАК та РЕТРОВІРУСИ
РАК та РЕТРОВІРУСИ

Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена
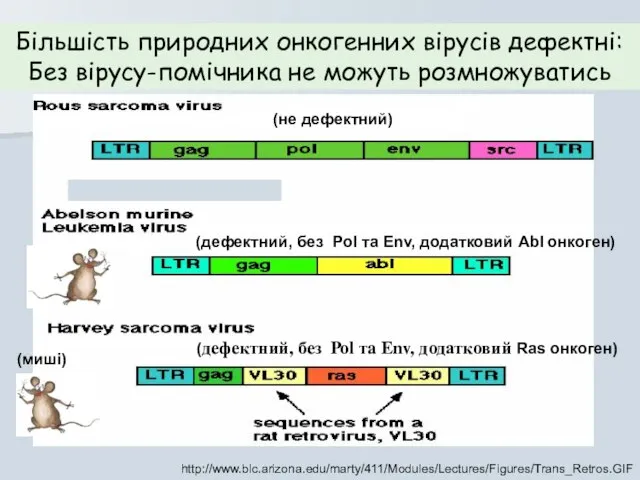
http://www.blc.arizona.edu/marty/411/Modules/Lectures/Figures/Trans_Retros.GIF
Більшість природних онкогенних вірусів дефектні:
Без вірусу-помічника не можуть розмножуватись
(не дефектний)
(дефектний,
http://www.blc.arizona.edu/marty/411/Modules/Lectures/Figures/Trans_Retros.GIF
Більшість природних онкогенних вірусів дефектні:
Без вірусу-помічника не можуть розмножуватись
(не дефектний)
(дефектний,

R U5 GAG POL ENV U3 R
Деякі Ретровіруси мають додатковий
R U5 GAG POL ENV U3 R
Деякі Ретровіруси мають додатковий

Вірус саркоми птахів (FSV)
R U5 dGAG FMS dENV U3 R
Вірус
Вірус саркоми птахів (FSV)
R U5 dGAG FMS dENV U3 R
Вірус

Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу
Клітинні онкогени
Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу
Клітинні онкогени

вірус саркоми Рауса є результатом рекомбінацій між c-src і давнім стандартним
вірус саркоми Рауса є результатом рекомбінацій між c-src і давнім стандартним

Структура дефектного РВ
Структура дефектного РВ

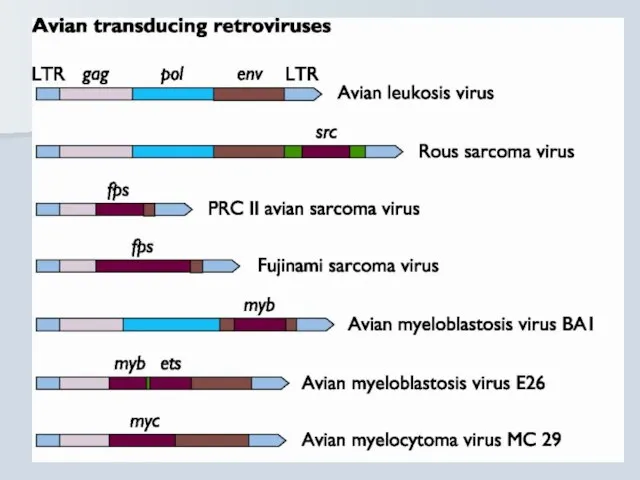
Гостро трансформуючі ретровіруси
src Rous sarcoma virus Chicken
myc Avian myelocytomatosis virus Chicken
erb A, erb B
Гостро трансформуючі ретровіруси
src Rous sarcoma virus Chicken
myc Avian myelocytomatosis virus Chicken
erb A, erb B
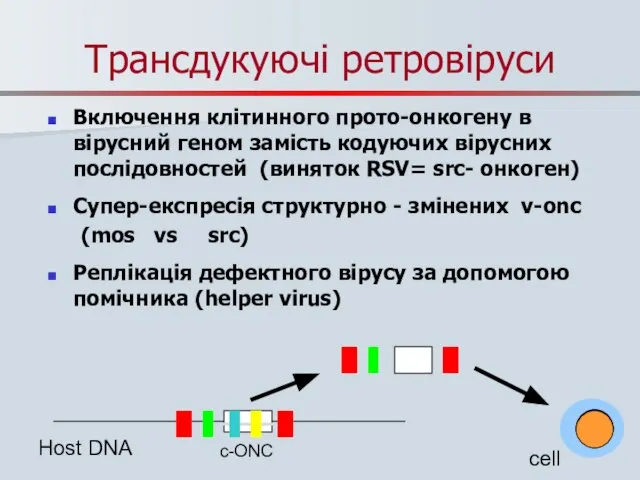
Трансдукуючі ретровіруси
Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей
Трансдукуючі ретровіруси
Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей
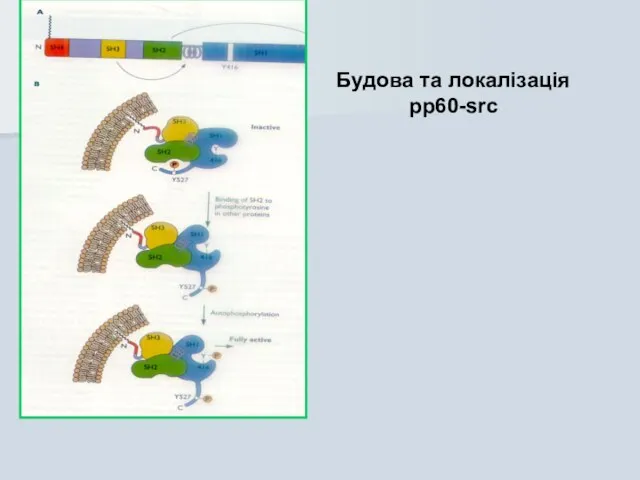
Будова та локалізація pp60-src
Будова та локалізація pp60-src

Активація src-гену
Активація src-гену
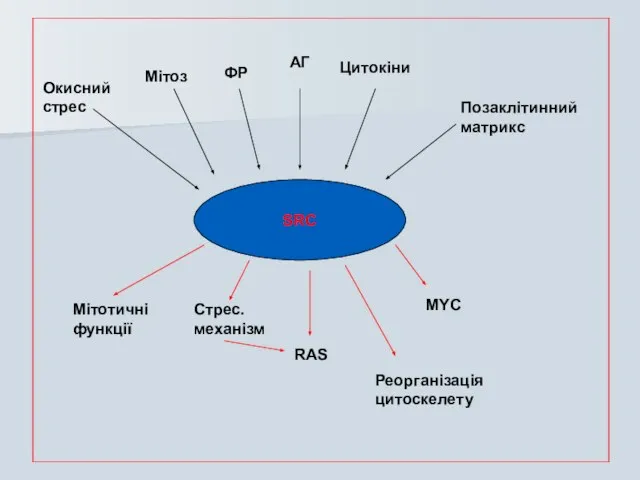
SRC
Окисний
стрес
Мітоз
ФР
АГ
Цитокіни
Позаклітинний
матрикс
Мітотичні
функції
Стрес.
механізм
RAS
MYC
Реорганізація
цитоскелету
SRC
Окисний
стрес
Мітоз
ФР
АГ
Цитокіни
Позаклітинний
матрикс
Мітотичні
функції
Стрес.
механізм
RAS
MYC
Реорганізація
цитоскелету

Наслідки ретровірусної трансдукції
“Простий удар” канцерогенезу (одна подія)
Поліклональна : пухлинний ріст починається
Наслідки ретровірусної трансдукції
“Простий удар” канцерогенезу (одна подія)
Поліклональна : пухлинний ріст починається

Характеристика вірусних онкогенів (Virology 2000)
Характеристика вірусних онкогенів (Virology 2000)

Трансдуковні гени – це гени сигнального шляху
Зовнішні сигнальні молекули або
Трансдуковні гени – це гени сигнального шляху
Зовнішні сигнальні молекули або
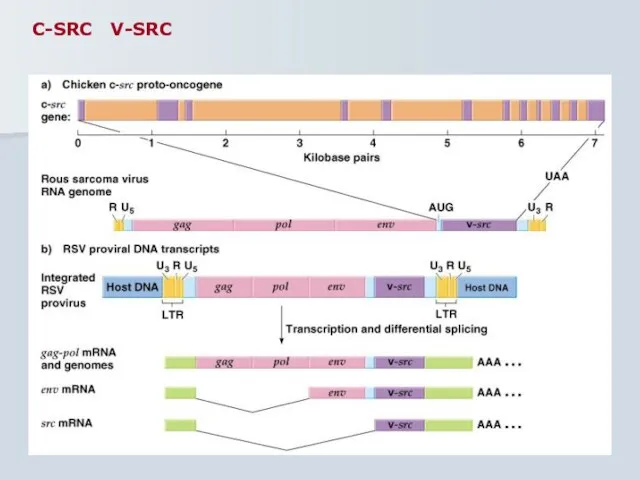
C-SRC V-SRC
C-SRC V-SRC

Мутації протоонкогенів Ras
характерні для пухлин людини
Мутації в кодонах 12,
Мутації протоонкогенів Ras
характерні для пухлин людини
Мутації в кодонах 12,
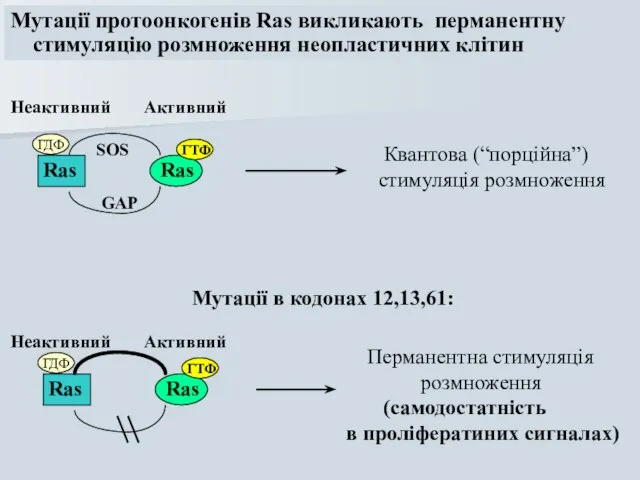
Мутації протоонкогенів Ras викликають перманентну
стимуляцію розмноження неопластичних клітин
Ras
Ras
ГДФ
ГТФ
SOS
GAP
Ras
Ras
ГДФ
ГТФ
Мутації в кодонах
Мутації протоонкогенів Ras викликають перманентну
стимуляцію розмноження неопластичних клітин
Ras
Ras
ГДФ
ГТФ
SOS
GAP
Ras
Ras
ГДФ
ГТФ
Мутації в кодонах
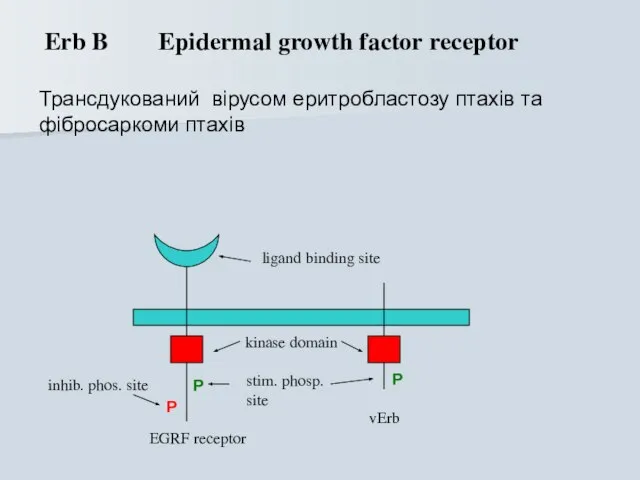
Erb B Epidermal growth factor receptor
Трансдукований вірусом еритробластозу птахів
Erb B Epidermal growth factor receptor
Трансдукований вірусом еритробластозу птахів
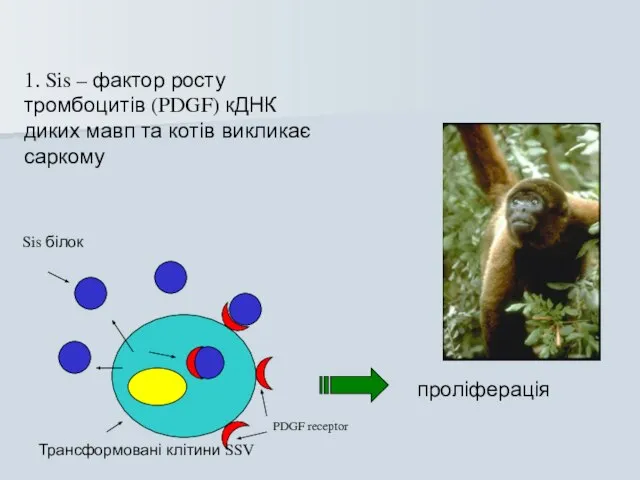
1. Sis – фактор росту тромбоцитів (PDGF) кДНК диких мавп та
1. Sis – фактор росту тромбоцитів (PDGF) кДНК диких мавп та
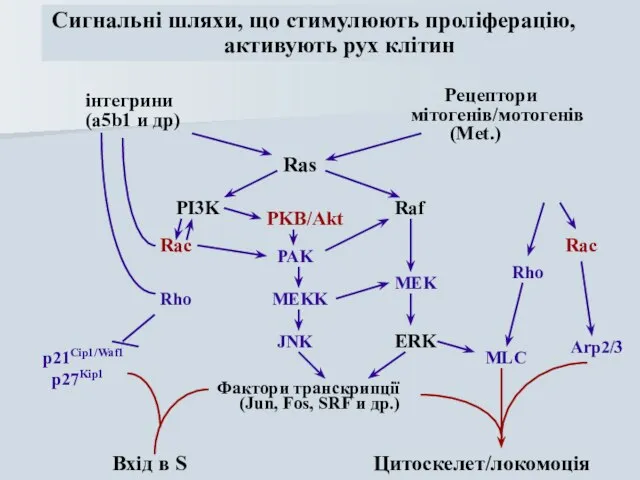
інтегрини
(a5b1 и др)
Рецептори
мітогенів/мотогенів
(Met.)
PI3K
Raf
ERK
Rho
Ras
MEK
Rac
PAK
MEKK
JNK
Фактори транскрипції
(Jun, Fos, SRF и
інтегрини
(a5b1 и др)
Рецептори
мітогенів/мотогенів
(Met.)
PI3K
Raf
ERK
Rho
Ras
MEK
Rac
PAK
MEKK
JNK
Фактори транскрипції
(Jun, Fos, SRF и

За функціональною активністю онкогени можна розділити на групи
1.а) з тирозинкіназною активністю
За функціональною активністю онкогени можна розділити на групи
1.а) з тирозинкіназною активністю

Онкогени як сигнали трансдукції
Фактор росту
v-sis, int-1, int-2, hst, fgf-5
Рецептори фактору росту
v-erb-B,
Онкогени як сигнали трансдукції
Фактор росту
v-sis, int-1, int-2, hst, fgf-5
Рецептори фактору росту
v-erb-B,

Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою
Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою


Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена

Cis-активуючі ретровіруси
Не мають онкогена
Зберігають всі вірусні гени
Компетентні за реплікацією
Cis-активуючі ретровіруси
Не мають онкогена
Зберігають всі вірусні гени
Компетентні за реплікацією

Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду
Немає онкогена!
Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду
Немає онкогена!

Пухлина походить з однієї клітини - моноклональна
ALV може інтегруватись в геном
Пухлина походить з однієї клітини - моноклональна
ALV може інтегруватись в геном

Чи специфічні ці сайти?
Вірус Myelocytoma виділений від різних птахів завжди
Чи специфічні ці сайти?
Вірус Myelocytoma виділений від різних птахів завжди

Як C-onc залучаються в не вірусний канцерогенез?
Як C-onc залучаються в не вірусний канцерогенез?
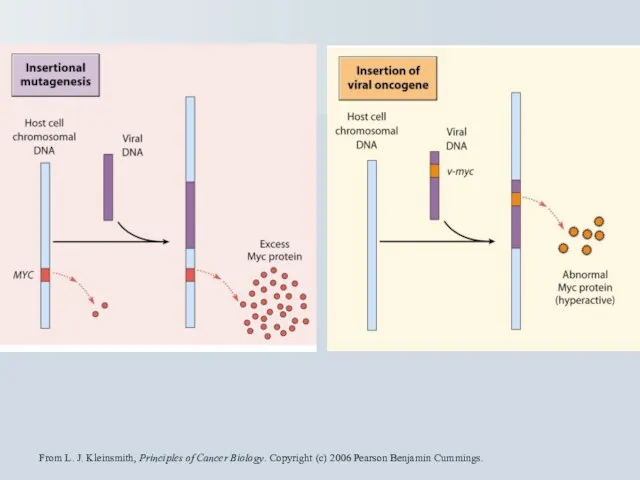
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006

Протоонкогени та перебудова хромомсом
Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc.
Протоонкогени та перебудова хромомсом
Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc.
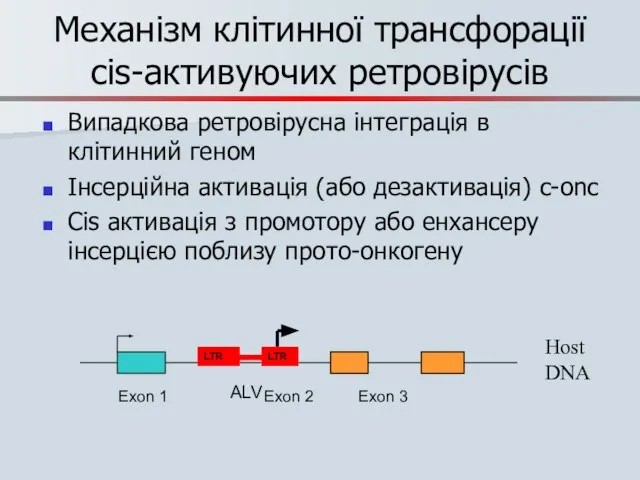
Механізм клітинної трансфорації cis-активуючих ретровірусів
Випадкова ретровірусна інтеграція в клітинний геном
Інсерційна активація
Механізм клітинної трансфорації cis-активуючих ретровірусів
Випадкова ретровірусна інтеграція в клітинний геном
Інсерційна активація
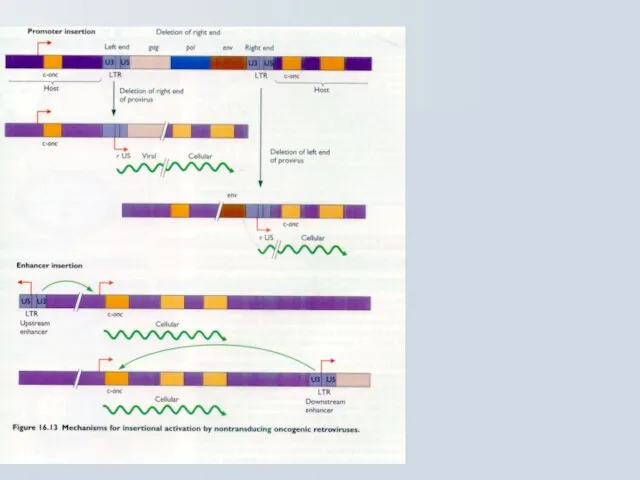

Наслідки активації онкогену при інсерції Ретровірусів
Клітинна трансформація відбувається завдяки
Наслідки активації онкогену при інсерції Ретровірусів
Клітинна трансформація відбувається завдяки

Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена

Human T cell Leukemia Virus type I (HTLV-I)
Асоційований з 2 фатальними
Human T cell Leukemia Virus type I (HTLV-I)
Асоційований з 2 фатальними

HTLV-I та ATL
1980 Галло ізолював тип C retrovirus (HTLV1) від
HTLV-I та ATL
1980 Галло ізолював тип C retrovirus (HTLV1) від

Міжклітинний контакт
Статевий (60% від чоловіка до жінки проти 1% від жінки
Міжклітинний контакт
Статевий (60% від чоловіка до жінки проти 1% від жінки

Онкогенез, опосередкований важливим ретровірусним білком
Виключення з парадігми ретровірусного онкогенезу (HTLV-1)
HTLV-1
Онкогенез, опосередкований важливим ретровірусним білком
Виключення з парадігми ретровірусного онкогенезу (HTLV-1)
HTLV-1
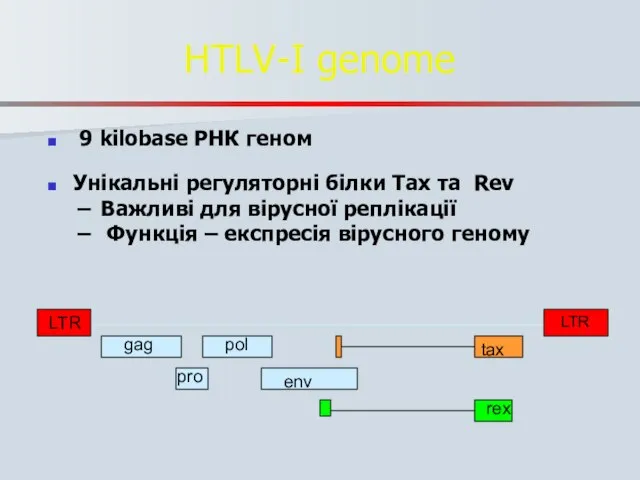
HTLV-I genome
9 kilobase РНК геном
Унікальні регуляторні білки Tax та Rev
HTLV-I genome
9 kilobase РНК геном
Унікальні регуляторні білки Tax та Rev

Tax тa Oнкогенез
Tax необхідний для вірусної реплікації
40kda фосфопротеїн
Tранскрипційний активатор геному HTLV-I
Взаємодія
Tax тa Oнкогенез
Tax необхідний для вірусної реплікації
40kda фосфопротеїн
Tранскрипційний активатор геному HTLV-I
Взаємодія

Tax – множинний трансактиватор
Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію
Tax – множинний трансактиватор
Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію

Білки регулятори клітинного циклу – мішені дії Tax
Iнактивація p53 (G1/S контроль
Білки регулятори клітинного циклу – мішені дії Tax
Iнактивація p53 (G1/S контроль
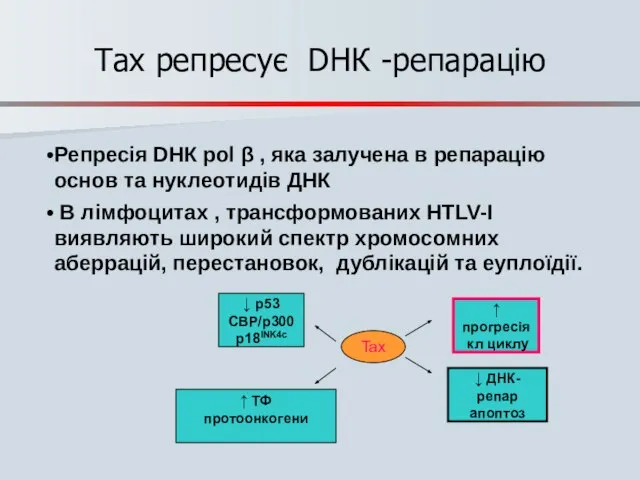
Tax репресує DНК -репарацію
Репресія DНК pol β , яка залучена в
Tax репресує DНК -репарацію
Репресія DНК pol β , яка залучена в

Механізми клітинної трансформації ретровірусами
Механізми клітинної трансформації ретровірусами

Онкогенні
РНК- віруси
Онкогенні
РНК- віруси
 «Эволюция человека»
«Эволюция человека» Эволюция человека
Эволюция человека Микрофлора окружающей среды. Санитарно-микробиологический контроль воды
Микрофлора окружающей среды. Санитарно-микробиологический контроль воды Эпителиальные ткани и железы. Ткани внутренней среды. Мышечные ткани. Нервные ткани
Эпителиальные ткани и железы. Ткани внутренней среды. Мышечные ткани. Нервные ткани …Е121…Е123..Е216…Е217…Е240… Пищевые добавки- полезные, вкусные и запрещённые Московских Анастасия 9 А класс МОУ СОШ № 27 города Бело
…Е121…Е123..Е216…Е217…Е240… Пищевые добавки- полезные, вкусные и запрещённые Московских Анастасия 9 А класс МОУ СОШ № 27 города Бело Endangered animals
Endangered animals Плесневые грибы и дрожжи (5 класс)
Плесневые грибы и дрожжи (5 класс) ТЕМА: Семейство Мальвовые ХЛОПЧАТНИК
ТЕМА: Семейство Мальвовые ХЛОПЧАТНИК  Сердечнососудистая, дыхательная и нервная системы
Сердечнососудистая, дыхательная и нервная системы Значение опорнодвигательной системы. Строение костей
Значение опорнодвигательной системы. Строение костей Культуральные свойства: Характеристика колоний микроорганизма
Культуральные свойства: Характеристика колоний микроорганизма Пингвины
Пингвины Технологии профайлинга и верификации
Технологии профайлинга и верификации Изменчивость. Мутации
Изменчивость. Мутации Мультимедийная презентация урока биологии в 7 классе Донская Ольга Демьяновна МБОУ «СОШ №14» г. Нефтеюганск
Мультимедийная презентация урока биологии в 7 классе Донская Ольга Демьяновна МБОУ «СОШ №14» г. Нефтеюганск Пескоройки. Стадия жизни миноги
Пескоройки. Стадия жизни миноги Русская берёза
Русская берёза Дыхание животных
Дыхание животных Життєвий цикл організації
Життєвий цикл організації Презентация на тему Клеточная инженерия
Презентация на тему Клеточная инженерия  Barības vielu veidošanās augos. Fotosintēze Dabaszinības 5.klase
Barības vielu veidošanās augos. Fotosintēze Dabaszinības 5.klase УРОК № 1.
УРОК № 1. Обмен веществ и энергии. Метаболизм
Обмен веществ и энергии. Метаболизм Исследование функций внешнего дыхания. Лекция 1
Исследование функций внешнего дыхания. Лекция 1 Історичний розвиток органічного світу
Історичний розвиток органічного світу Биохимия, предмет и задачи. Структурно-функциональная организация белковых молекул
Биохимия, предмет и задачи. Структурно-функциональная организация белковых молекул Тип кольчатые черви, класс малощетинковые
Тип кольчатые черви, класс малощетинковые Окислительное фосфорилирование
Окислительное фосфорилирование