- Основні проблеми макроеволюції

Содержание
- 2. Морські лілії
- 3. Основні проблеми макроеволюції Закономірності еволюції груп (таксонів) Адаптаціогенез Закономірності еволюції онтогенезу Прогрес і еволюція Спрямованість еволюційного
- 4. Закономірності еволюції груп Правило незворотності еволюції (закон Долло) Правило походження нових груп від неспеціалізованих предків(закон Копа)
- 5. Правило незворотності еволюції Форма тіла швидкоплава-ючих тварин набуває конвергентної схожості, проте повернення до предкових форм у
- 6. Спеціалізація не потребує змін у всіх органах системи, проте може бути досягнута за умов збереження у
- 7. Еволюція шляхом гіперморфозу: Закон Копа-Депере Предком могутнього плейстоценового південного слона був невеличкий ссавець мерітерій.
- 8. Засоби філогенетичних змін органів і функцій Збільшення (розширення) кількості функцій Посилення (інтенсифікація) головної функції Послаблення головної
- 9. Гетеробатмія (А.Л. Тахтаджян, 1954) – це неоднаковий еволюційний рівень розвитку різних органів, що виникає внаслідок неоднакового
- 10. Форми еволюційних змін онтогенезу Ембріонізація онтогенеза Перебудови кореляційних взаємозв’язків Неотенізація Автономізація онтогенеза Філембріогенези Анаболія Девіація Архалаксис
- 11. Біогенетичний закон та закон зародкової подібності
- 12. Рекапітуляція будови задньої кінцівки стегоцефалів у онтогенезі сучасних амфібій (за Северцовим, 2005)
- 13. Листкова жаба Паразитичні оси
- 14. Моделі еволюції онтогенезу (філембріогенези та ембріоадаптація). A-І - фази онтогенезу; N- народження; Z- зигота. Індексами позначено
- 15. Прогрес в еволюції Необмежений Біологічний Морфофізіологічний Біотехнічний Груповий
- 16. Необмежений прогрес Терипетна тенденція Цефалізація Поява ноосфери Зростання середньої тривалості існування родів у процесі еволюції Збільшення
- 17. Зростання світимості Сонця +30% 4,0млрд.р. до сучасності Поява ноосфери Зростання об’єму світового океану (4,0-0,5млрд.р.) Террес-тризація Зростання
- 18. Зростання чисельності родів морських організмів Кількість видів морських тварин в різні періоди фанерозоя
- 19. Ріст середньої тривалості існування родів у процесі еволюції Середня тривалість існування родів у млн. років
- 20. Просторова структура ареалу соболя (Martes zibellina). 1 – вихідні межі ареалу, 2 – острівки населення, що
- 21. Різниця у чисельності видів, що входять до різних родів родини білячих: 1 – Callosciurus, 2 –
- 22. Зростання ступеня різноманіття в межах певного таксону при захопленні їм нової адаптивної зони отримало назву адаптивної
- 23. Ароморфоз - еволюційні перебудови організмів, що ведуть до морфофізіологічного прогресу. Катаморфоз - регресивні зміни організмів. Епектоморфози
- 24. Співвідношення між основними формами еволюційного процеса
- 25. Посилення легеневого дихання (за рахунок появи діафрагми та більш досконалої реберної мускулатури)
- 26. Паралельна еволюція. Ссавці. Процес набуття ознак маммального рівня організації йшов паралельно в кількох споріднених групах теріоморфних
- 27. Паралельна еволюція. Птахи. Рептилії Тріас Юра Крейда Палеоген Protoavis Hesperornis Ichtyornis Chaoyangia Сучасні віялохвості птахи (Ornithurae)
- 28. Деяки опірені динозаври-протоптахи Авімім Конфуціусорніс, рахонавіс, енанціорніс Крипто- воланс
- 29. Асинхронний паралелізм на прикладі розвитку шаблезубості: Махайродонтові Справжні кішки Сумчасті Теріодонти Hopiophoneus Smilоdon Dinictus Pseudaelurus Thylacosmilus
- 30. Монофілія та поліфілія: гради та клади Риби Амфібії Рептилії Птахи та ссавці
- 31. Розвиток носових "рогів" у еволюції титанотеріїв: 1 - Eotitanops, 2 - Lymnohyops, 3 - Manteoceras, 4
- 32. Найбільш стійкими до глобальної екологічної кризи на границі крейди та палеогену виявились старі роди.
- 33. Філогенетичний цикл Кількість видів Середній вік видів Об’єм вільного простору в адаптивній зоні таксону Середній об’єм
- 34. Філогенетичний цикл на прикладі археоциатів Кількість видів Середня площа ареала Середня тривалість існування роду Середній об’єм
- 35. Деякі еволюційні зміни у родині Equidae: 1-5 передні кінцівки представників родів Eohippus (1), Miohippus (2), Parahippus
- 36. Адаптивний компроміс
- 38. Інадаптації та евадаптації Інадаптація - неглибоке, однобічне пристосування, яке здійснюється за рахунок порушення координаційних зв’язків у
- 39. Евадаптація - глибоке, досконале пристосування, яке досягається за рахунок оптимізації координаційних зв’язків в організмі.
- 40. Група креодонтів досягла швидкого успіху, проте згодом поступилася справжнім хижим. Оксієна
- 41. Темпи еволюційних перетворень 1 Дарвін : зміна розмірів органа на 0,001 його величини за 1000 років
- 42. Вік напівсучасної фауни родів деяких таксонів Ссавці - 4 млн р. Птахі - 10 млн. р.
- 43. Вік напівсучасної фауни деяких таксонів та середня генетична дистанція між видами (за Расніциним, зі скороченнями та
- 44. Живі викопні - роди, які існують на Землі десятки-сотні міліонів років
- 46. Види – ізоляти (мешканці рефугіумів) Типи живих викопних Види – мешканці реліктових біогеоценозів Види, що займають
- 47. Популяції виду Delamarentulus tristani Silv. з групи Protura мешкають у Коста-Ріці та у Західній Африці. Ізоляція
- 48. Фактори, що сприяють швидкій еволюції груп: Статеве розмноження Можливість дії статевого добору Складність будови Великі розміри
- 50. Скачать презентацию

Морські лілії
Морські лілії

Основні проблеми макроеволюції
Закономірності еволюції груп (таксонів)
Адаптаціогенез
Закономірності еволюції онтогенезу
Прогрес і еволюція
Спрямованість еволюційного
Основні проблеми макроеволюції
Закономірності еволюції груп (таксонів)
Адаптаціогенез
Закономірності еволюції онтогенезу
Прогрес і еволюція
Спрямованість еволюційного

Закономірності еволюції груп
Правило незворотності еволюції (закон Долло)
Правило походження нових груп від
Закономірності еволюції груп
Правило незворотності еволюції (закон Долло)
Правило походження нових груп від

Правило незворотності еволюції
Форма тіла швидкоплава-ючих тварин набуває конвергентної схожості, проте повернення
Правило незворотності еволюції
Форма тіла швидкоплава-ючих тварин набуває конвергентної схожості, проте повернення
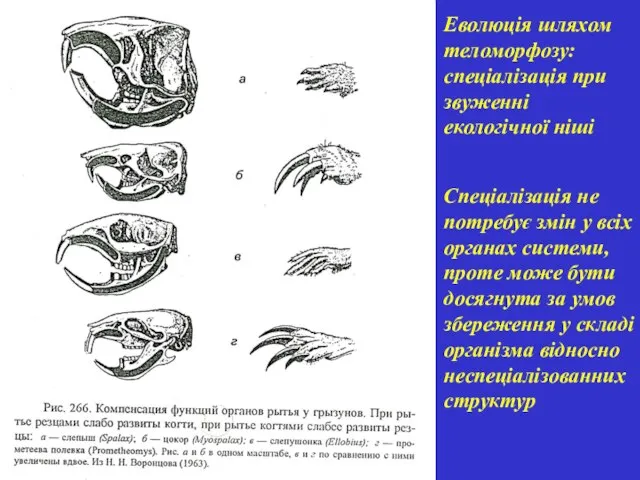
Спеціалізація не потребує змін у всіх органах системи, проте може бути
Спеціалізація не потребує змін у всіх органах системи, проте може бути

Еволюція шляхом гіперморфозу: Закон Копа-Депере
Предком могутнього плейстоценового південного слона був невеличкий
Еволюція шляхом гіперморфозу: Закон Копа-Депере
Предком могутнього плейстоценового південного слона був невеличкий
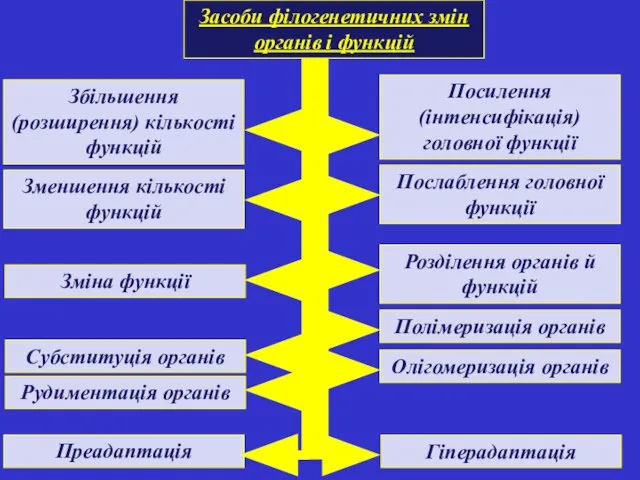
Засоби філогенетичних змін органів і функцій
Збільшення (розширення) кількості функцій
Посилення (інтенсифікація) головної
Засоби філогенетичних змін органів і функцій
Збільшення (розширення) кількості функцій
Посилення (інтенсифікація) головної

Гетеробатмія (А.Л. Тахтаджян, 1954) – це неоднаковий еволюційний рівень розвитку різних органів,
Гетеробатмія (А.Л. Тахтаджян, 1954) – це неоднаковий еволюційний рівень розвитку різних органів,
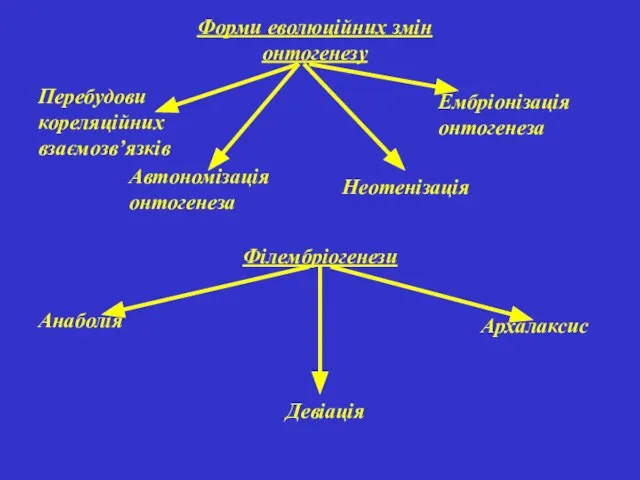
Форми еволюційних змін онтогенезу
Ембріонізація онтогенеза
Перебудови кореляційних взаємозв’язків
Неотенізація
Автономізація онтогенеза
Філембріогенези
Анаболія
Девіація
Архалаксис
Форми еволюційних змін онтогенезу
Ембріонізація онтогенеза
Перебудови кореляційних взаємозв’язків
Неотенізація
Автономізація онтогенеза
Філембріогенези
Анаболія
Девіація
Архалаксис
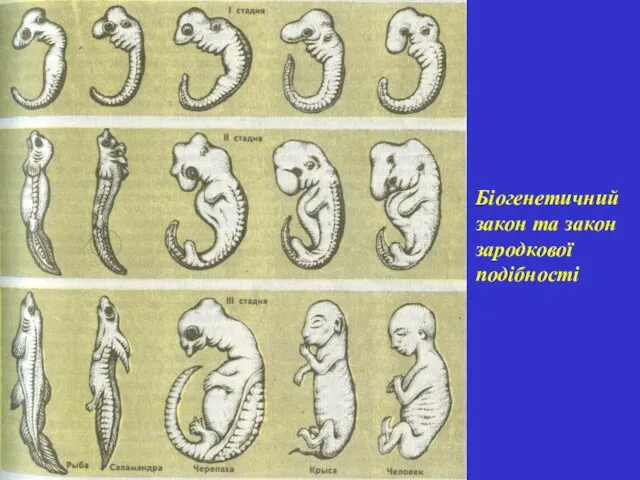
Біогенетичний закон та закон зародкової подібності
Біогенетичний закон та закон зародкової подібності
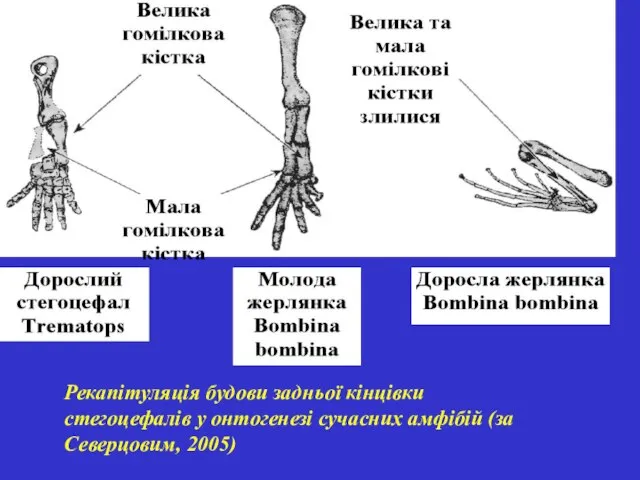
Рекапітуляція будови задньої кінцівки стегоцефалів у онтогенезі сучасних амфібій (за Северцовим,
Рекапітуляція будови задньої кінцівки стегоцефалів у онтогенезі сучасних амфібій (за Северцовим,

Листкова жаба
Паразитичні оси
Листкова жаба
Паразитичні оси
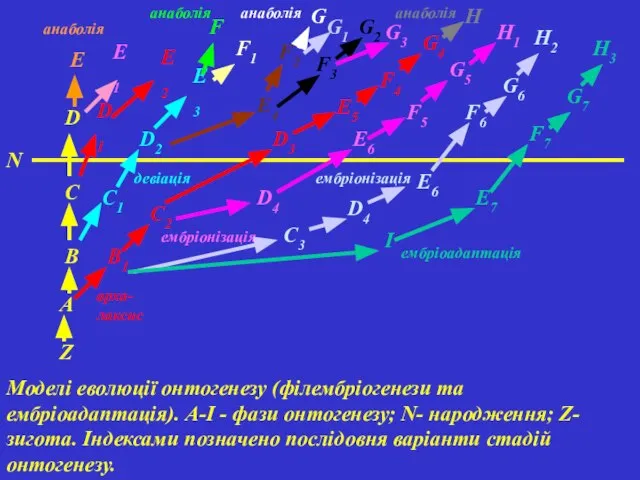
Моделі еволюції онтогенезу (філембріогенези та ембріоадаптація). A-І - фази онтогенезу; N-
Моделі еволюції онтогенезу (філембріогенези та ембріоадаптація). A-І - фази онтогенезу; N-
Прогрес в еволюції
Необмежений
Біологічний
Морфофізіологічний
Біотехнічний
Груповий
Прогрес в еволюції
Необмежений
Біологічний
Морфофізіологічний
Біотехнічний
Груповий
Необмежений прогрес
Терипетна тенденція
Цефалізація
Поява ноосфери
Зростання середньої тривалості існування родів у процесі еволюції
Збільшення
Необмежений прогрес
Терипетна тенденція
Цефалізація
Поява ноосфери
Зростання середньої тривалості існування родів у процесі еволюції
Збільшення
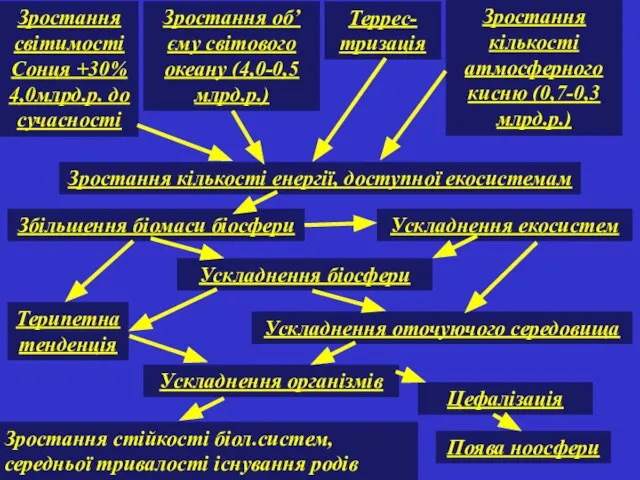
Зростання світимості Сонця +30% 4,0млрд.р. до сучасності
Поява ноосфери
Зростання об’єму світового океану
Зростання світимості Сонця +30% 4,0млрд.р. до сучасності
Поява ноосфери
Зростання об’єму світового океану
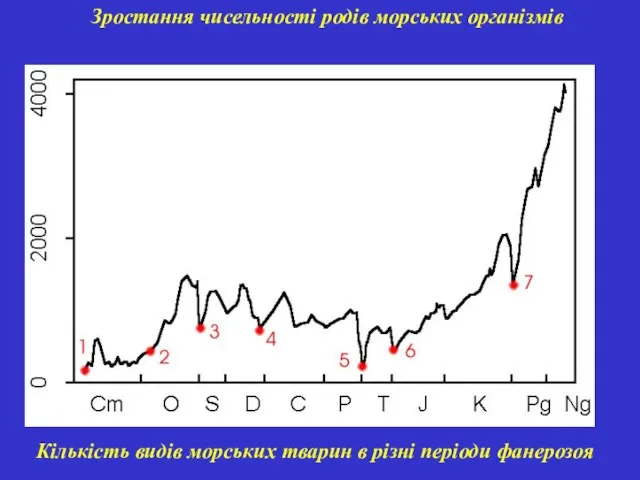
Зростання чисельності родів морських організмів
Кількість видів морських тварин в різні періоди
Зростання чисельності родів морських організмів
Кількість видів морських тварин в різні періоди
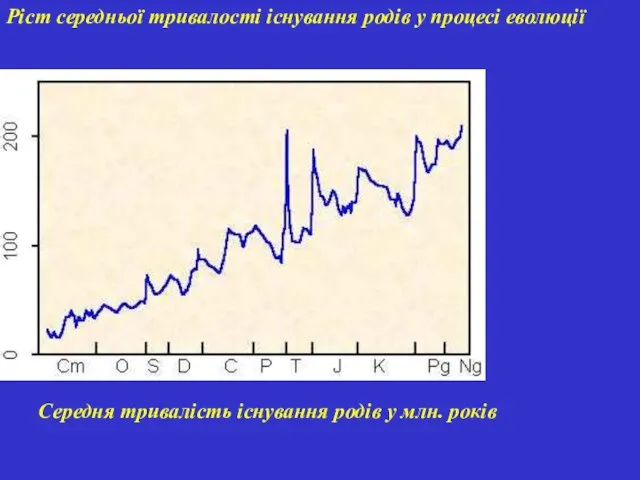
Ріст середньої тривалості існування родів у процесі еволюції
Середня тривалість існування родів
Ріст середньої тривалості існування родів у процесі еволюції
Середня тривалість існування родів
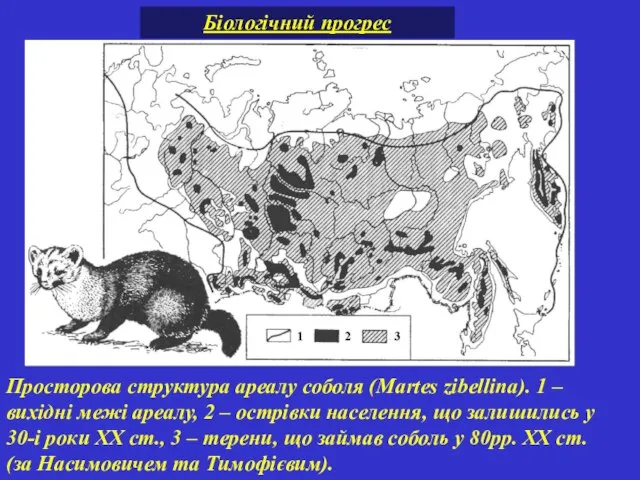
Просторова структура ареалу соболя (Martes zibellina). 1 – вихідні межі ареалу,
Просторова структура ареалу соболя (Martes zibellina). 1 – вихідні межі ареалу,
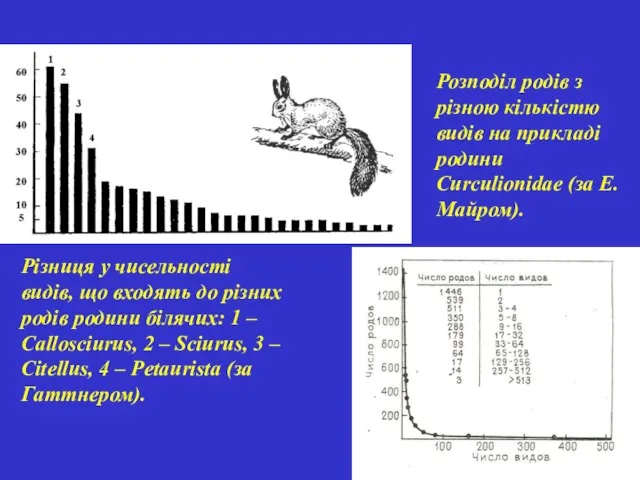
Різниця у чисельності видів, що входять до різних родів родини білячих:
Різниця у чисельності видів, що входять до різних родів родини білячих:

Зростання ступеня різноманіття в межах певного таксону при захопленні їм нової
Зростання ступеня різноманіття в межах певного таксону при захопленні їм нової

Ароморфоз - еволюційні перебудови організмів, що ведуть до морфофізіологічного прогресу.
Катаморфоз
Ароморфоз - еволюційні перебудови організмів, що ведуть до морфофізіологічного прогресу.
Катаморфоз

Співвідношення між основними формами еволюційного процеса
Співвідношення між основними формами еволюційного процеса
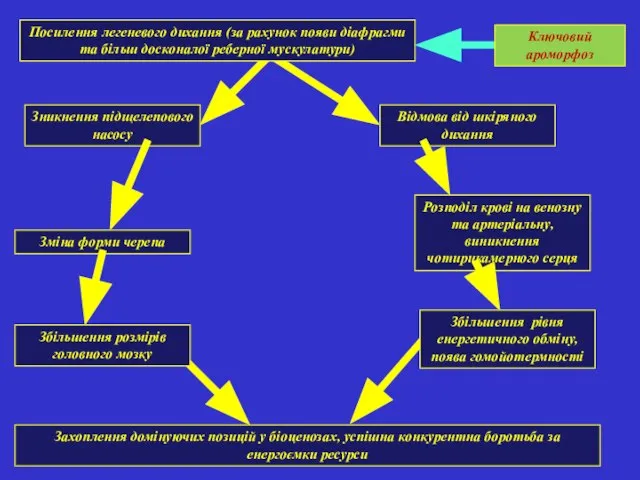
Посилення легеневого дихання (за рахунок появи діафрагми та більш досконалої реберної
Посилення легеневого дихання (за рахунок появи діафрагми та більш досконалої реберної
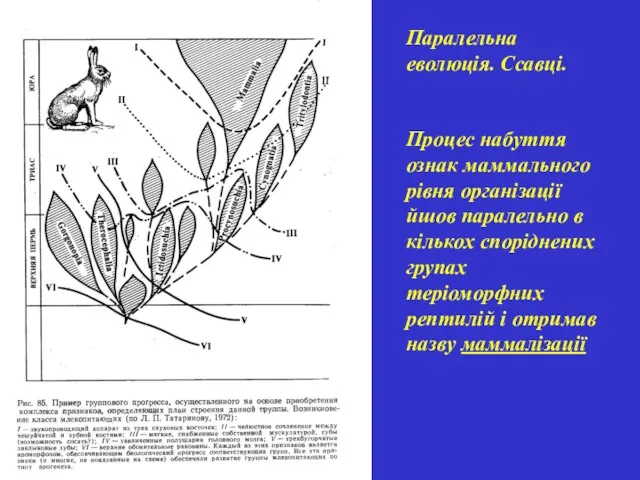
Паралельна еволюція. Ссавці.
Процес набуття ознак маммального рівня організації йшов паралельно в
Паралельна еволюція. Ссавці.
Процес набуття ознак маммального рівня організації йшов паралельно в
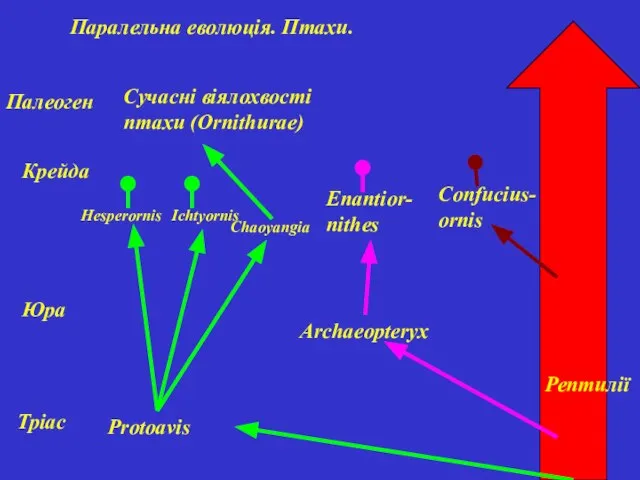
Паралельна еволюція. Птахи.
Рептилії
Тріас
Юра
Крейда
Палеоген
Protoavis
Hesperornis
Ichtyornis
Chaoyangia
Сучасні віялохвості птахи (Ornithurae)
Archaeopteryx
Enantior-nithes
Confucius-ornis
Паралельна еволюція. Птахи.
Рептилії
Тріас
Юра
Крейда
Палеоген
Protoavis
Hesperornis
Ichtyornis
Chaoyangia
Сучасні віялохвості птахи (Ornithurae)
Archaeopteryx
Enantior-nithes
Confucius-ornis

Деяки опірені динозаври-протоптахи
Авімім
Конфуціусорніс, рахонавіс, енанціорніс
Крипто- воланс
Деяки опірені динозаври-протоптахи
Авімім
Конфуціусорніс, рахонавіс, енанціорніс
Крипто- воланс
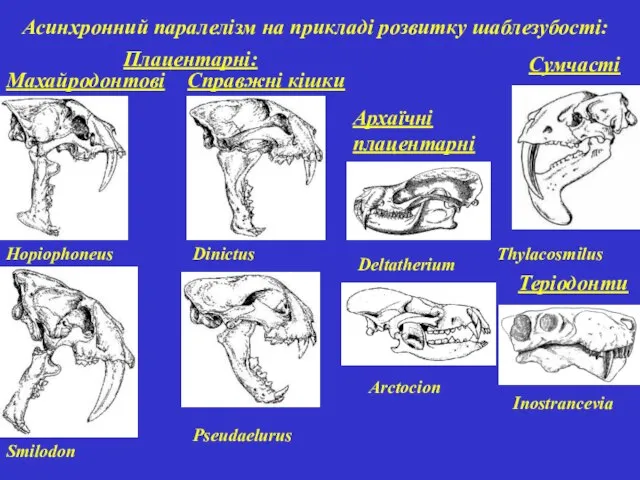
Асинхронний паралелізм на прикладі розвитку шаблезубості:
Махайродонтові
Справжні кішки
Сумчасті
Теріодонти
Hopiophoneus
Smilоdon
Dinictus
Pseudaelurus
Thylacosmilus
Inostrancevia
Плацентарні:
Архаїчні плацентарні
Deltatherium
Arctocion
Асинхронний паралелізм на прикладі розвитку шаблезубості:
Махайродонтові
Справжні кішки
Сумчасті
Теріодонти
Hopiophoneus
Smilоdon
Dinictus
Pseudaelurus
Thylacosmilus
Inostrancevia
Плацентарні:
Архаїчні плацентарні
Deltatherium
Arctocion

Монофілія та поліфілія: гради та клади
Риби
Амфібії
Рептилії
Птахи та ссавці
Монофілія та поліфілія: гради та клади
Риби
Амфібії
Рептилії
Птахи та ссавці
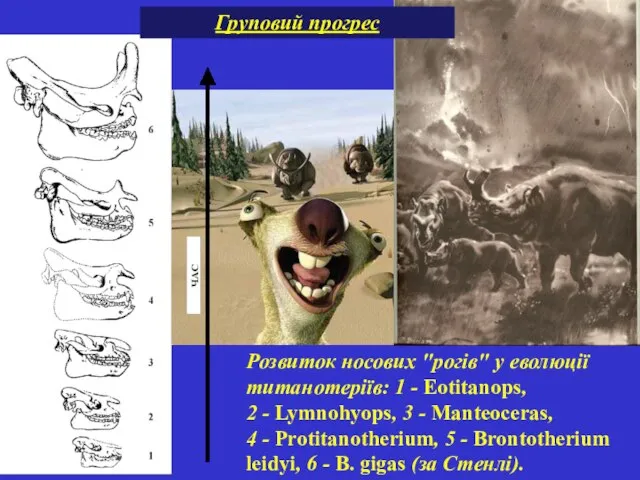
Розвиток носових "рогів" у еволюції титанотеріїв: 1 - Eotitanops, 2 - Lymnohyops, 3 - Manteoceras, 4 - Protitanotherium, 5 - Brontotherium
Розвиток носових "рогів" у еволюції титанотеріїв: 1 - Eotitanops, 2 - Lymnohyops, 3 - Manteoceras, 4 - Protitanotherium, 5 - Brontotherium
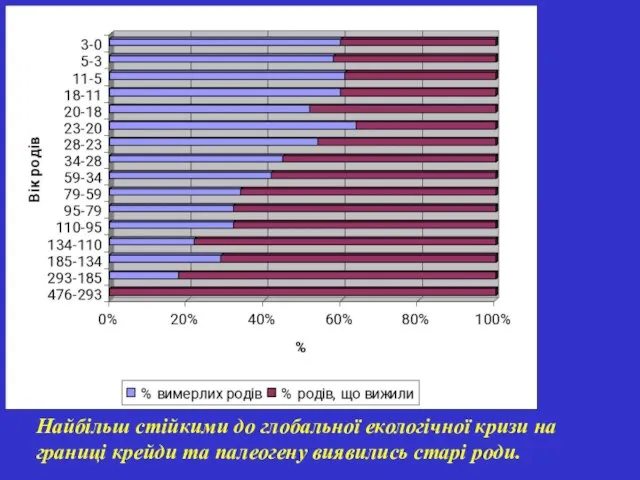
Найбільш стійкими до глобальної екологічної кризи на границі крейди та палеогену
Найбільш стійкими до глобальної екологічної кризи на границі крейди та палеогену
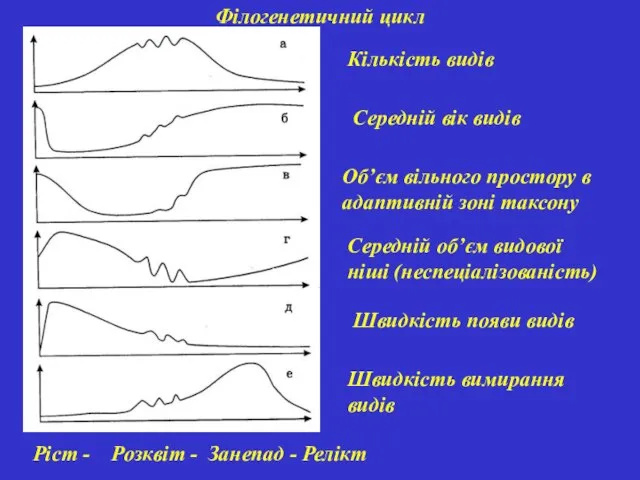
Філогенетичний цикл
Кількість видів
Середній вік видів
Об’єм вільного простору в адаптивній зоні таксону
Середній
Філогенетичний цикл
Кількість видів
Середній вік видів
Об’єм вільного простору в адаптивній зоні таксону
Середній
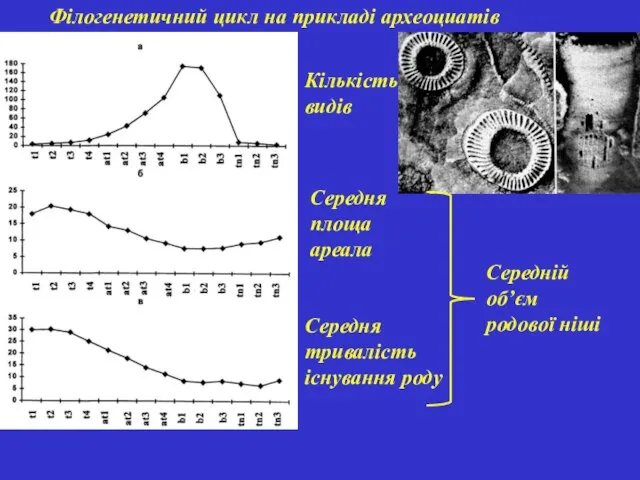
Філогенетичний цикл на прикладі археоциатів
Кількість видів
Середня площа ареала
Середня тривалість існування роду
Середній
Філогенетичний цикл на прикладі археоциатів
Кількість видів
Середня площа ареала
Середня тривалість існування роду
Середній
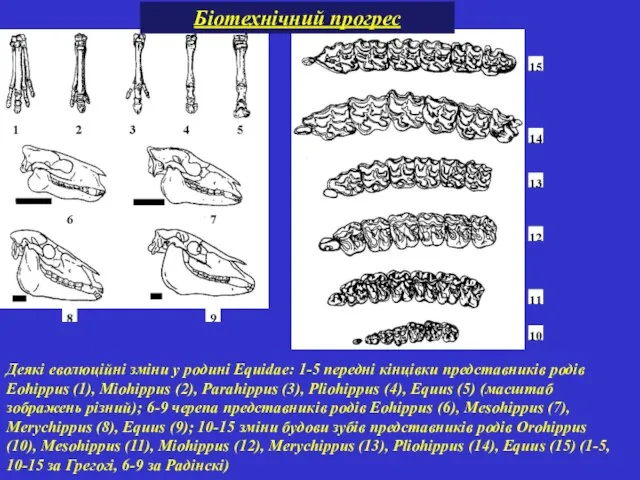
Деякі еволюційні зміни у родині Equidae: 1-5 передні кінцівки представників родів Eohippus
Деякі еволюційні зміни у родині Equidae: 1-5 передні кінцівки представників родів Eohippus

Адаптивний компроміс
Адаптивний компроміс

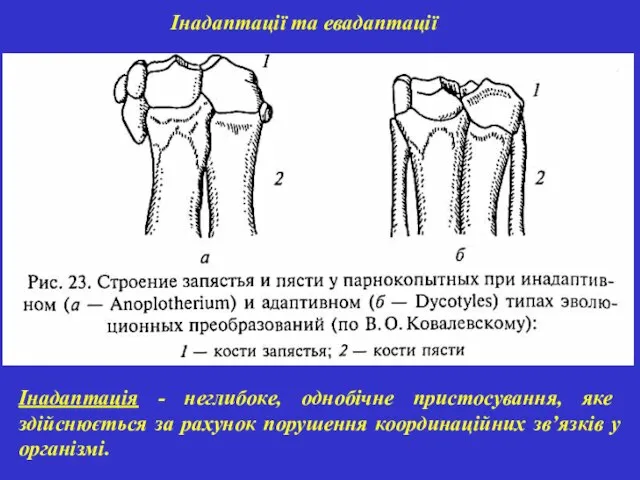
Інадаптації та евадаптації
Інадаптація - неглибоке, однобічне пристосування, яке здійснюється за рахунок
Інадаптації та евадаптації
Інадаптація - неглибоке, однобічне пристосування, яке здійснюється за рахунок
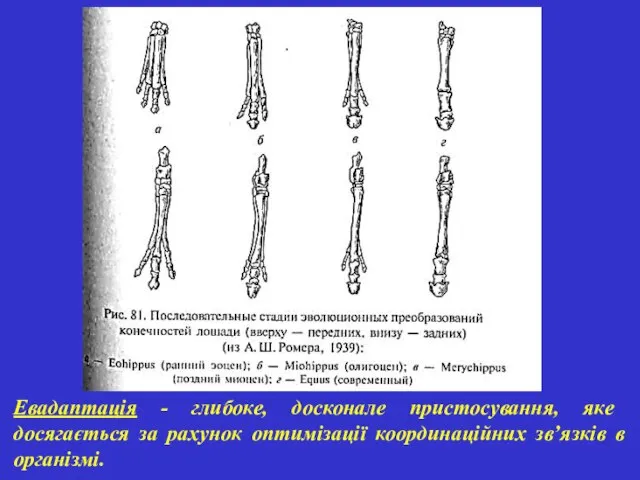
Евадаптація - глибоке, досконале пристосування, яке досягається за рахунок оптимізації координаційних
Евадаптація - глибоке, досконале пристосування, яке досягається за рахунок оптимізації координаційних

Група креодонтів досягла швидкого успіху, проте згодом поступилася справжнім хижим.
Оксієна
Група креодонтів досягла швидкого успіху, проте згодом поступилася справжнім хижим.
Оксієна

Темпи еволюційних перетворень
1 Дарвін : зміна розмірів органа на 0,001 його
Темпи еволюційних перетворень
1 Дарвін : зміна розмірів органа на 0,001 його

Вік напівсучасної фауни родів деяких таксонів
Ссавці - 4 млн р.
Птахі
Вік напівсучасної фауни родів деяких таксонів
Ссавці - 4 млн р.
Птахі
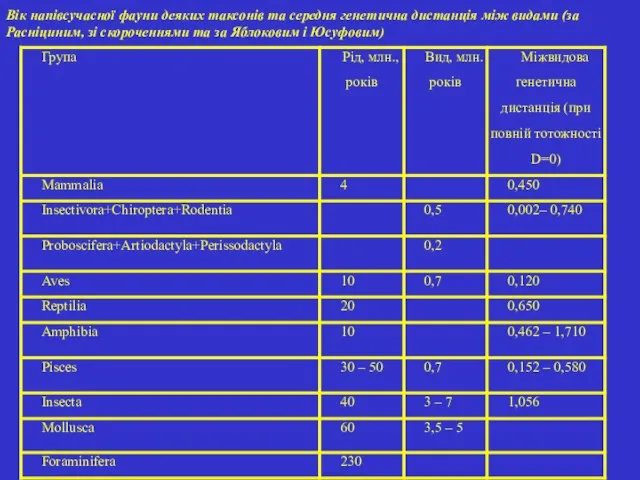
Вік напівсучасної фауни деяких таксонів та середня генетична дистанція між видами
Вік напівсучасної фауни деяких таксонів та середня генетична дистанція між видами

Живі викопні - роди, які існують на Землі десятки-сотні міліонів років
Живі викопні - роди, які існують на Землі десятки-сотні міліонів років


Види – ізоляти (мешканці рефугіумів)
Типи живих викопних
Види – мешканці реліктових біогеоценозів
Види,
Види – ізоляти (мешканці рефугіумів)
Типи живих викопних
Види – мешканці реліктових біогеоценозів
Види,

Популяції виду Delamarentulus tristani Silv. з групи Protura мешкають у Коста-Ріці
Популяції виду Delamarentulus tristani Silv. з групи Protura мешкають у Коста-Ріці

Фактори, що сприяють швидкій еволюції груп:
Статеве розмноження
Можливість дії статевого добору
Складність будови
Великі
Фактори, що сприяють швидкій еволюції груп:
Статеве розмноження
Можливість дії статевого добору
Складність будови
Великі
 Презентация на тему "Первая помощь при нарушении органов дыхания" - скачать презентации по Биологии
Презентация на тему "Первая помощь при нарушении органов дыхания" - скачать презентации по Биологии Обмен углеводов
Обмен углеводов Хищные птицы
Хищные птицы Метаболизм. Взаимосвязь различных путей обмена веществ
Метаболизм. Взаимосвязь различных путей обмена веществ Введение в фитотоксикологию
Введение в фитотоксикологию ГИГИЕНА СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ ЦЕЛИ УРОКА: Ознакомиться с основными заболеваниями ССС и правилами их профилактики. Научит
ГИГИЕНА СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ ЦЕЛИ УРОКА: Ознакомиться с основными заболеваниями ССС и правилами их профилактики. Научит Размножение живых организмов
Размножение живых организмов Знаменитые растения Крыма
Знаменитые растения Крыма Гормоны. Классификация гормонов
Гормоны. Классификация гормонов Предмет и задачи микробиологии
Предмет и задачи микробиологии Класс Насекомые
Класс Насекомые Анатомия сердца
Анатомия сердца Тип Членистоногие (Уроки РК по биологии 7 класс) Многообразие ракообразных. Паукообразные и их роль в природе
Тип Членистоногие (Уроки РК по биологии 7 класс) Многообразие ракообразных. Паукообразные и их роль в природе Обмен веществ и энергии. Терморегуляция
Обмен веществ и энергии. Терморегуляция Забота о потомстве у животных
Забота о потомстве у животных Презентация на тему Агроценоз
Презентация на тему Агроценоз  Вены большого круга кровообращения. Сосуды малого и коронарного кругов кровообращения
Вены большого круга кровообращения. Сосуды малого и коронарного кругов кровообращения Поражение глазодвигательного (III) нерва (n. oculomotorius)
Поражение глазодвигательного (III) нерва (n. oculomotorius) Опорно-двигательная система человека. Скелет
Опорно-двигательная система человека. Скелет Биосоциальная природа человека
Биосоциальная природа человека  Технология механизированных работ посева кукурузы на зерно
Технология механизированных работ посева кукурузы на зерно Многообразие и значение грибов
Многообразие и значение грибов Нервная система человека
Нервная система человека Медузы
Медузы Тип Губки. Тип Кишечнополостные
Тип Губки. Тип Кишечнополостные Тип Мшанки
Тип Мшанки Анатомия глаза, его придатков и глазницы
Анатомия глаза, его придатков и глазницы Аттестационная работа. Методическая разработка по выполнению проекта Необходимые и полезные овощи для нашего организма
Аттестационная работа. Методическая разработка по выполнению проекта Необходимые и полезные овощи для нашего организма