- Введение в нейрофизиологию

Содержание
- 2. План лекции Введение История развития нейрофизиологии Основные процессы в нервной системе Нейрон – как структурно-функциональная единица
- 3. РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ
- 4. Крылова Н.В., Искренко И.А. Мозг и проводящие пути: Анатомия человека в схемах и рисунках: Атлас-пособие. Изд.
- 5. Достижения человечества существуют благодаря разуму, вместилищем которого является головной мозг. Все продукты цивилизации адаптированы и ограниченны
- 6. Введение Нейрофизиология относится к разделу физиологии, который изучает функции нервной системы, в том числе её единиц
- 7. Разделы нейрофизиологии Общая нейрофизиология - изучает закономерности функционирования нервной системы на разных уровнях. Возрастная нейрофизиология –это
- 8. История открытия и развитие представлений о биопотенциалах головного мозга
- 9. История развития нейрофизиологии Зависимость сознания от мозга утверждал еще отец медицины - Гиппократ. Ученики Гиппократа, следуя
- 10. Первые исследования около 620-550 до н.э. – Фалес Милетский - считается первооткрывателем статического электричества, производимого трением
- 11. История электрофизиологии началась с изучения влияния электричества на организм. В 18 веке физики, занимавшиеся изучением электрических
- 12. История развития нейрофизиологии Первые представления о рефлекторном принципе действия нервной системы человека были сформулированы в XVII
- 13. Начало электрофизиологии обычно связывают со знаменитыми опытами итальянского врача, анатома и физиолога Луиджи Гальвани (L. Galvani
- 15. Карло Маттеучи (Carlo Matteucci 1811 – 1868) в 1830-1840 годах показал, что в мышце всегда может
- 17. Опыт по электрофизиологии нервов и мышц лягушки [Du Bois-Reymond: Frog Experiment (1848)] Эмиль Дюбуа Реймон Первая
- 18. В 1875 году английский хирург и физиолог Ричард Кэтон на заседании Британской медицинской ассоциации впервые сообщил,
- 19. Василий Яковлевич Данилевский В 1875 г. В.Я. Данилевский изложил данные по изучению фоновой электрической активности обнаженного
- 20. Иван Михайлович Сеченов 1864 г. «Рефлексы головного мозга» В 1882 году И.М.Сеченов опубликовал работу “Гальванические явления
- 21. Иван Михайлович Сеченов 1829 – 1905 В 1863 г. опубликовал книгу «Рефлексы головного мозга», где приводил
- 22. Он обосновал возможность распространения принципа рефлекса как принципа организации поведения на всю работу головного мозга
- 23. В 1884 году Н.Е. Введенский в работе “Исследования над нервными центрами” применил телефонический метод для изучения
- 24. Б.Ф. Вериго В 1889 г. В журнале «Вестник клинической и судебной психиатрии и невропатологи» была опубликована
- 25. 1890 год - работа А. Бека, посвященная использованию метода регистрации электрической активности мозга для вопросов локализации
- 26. 1898 год - В.Е. Ларионов производит исследования электрических явлений в головном мозгу для доказательства существования открытых
- 27. 1900 год – работа С.А. Тривуса, который получил определенные изменения потенциалов при зрительных раздражениях. Впервые установил
- 28. 1912 год - обширное исследование П.Ю. Кауфмана, который в тщательных опытах на собаках показывает, что электрические
- 29. Первая запись биотоков головного мозга человека получена Гансом Бергером в 1925 году. Ганс Бергер Первая запись
- 30. Нейрофизиологические методы: Внутриклеточная регистрация активности нервных клеток (ПП, ПД) Внеклеточная регистрация нейронной активности (ПД, ЭЭГ) Гистологические
- 32. Регистрация импульсной активности нервных клеток Фотографии нейронов, окрашенных флуоресцентным красителем Подведение электрода к телу нейрона
- 33. Два подхода к изучению мозга Гемодинамика (метаболизм) – ПЭТ, ЯМРИ Электрофизиология – ЭЭГ, МЭГ
- 35. Основные ритмы ЭЭГ Альфа-ритм, 8-13 Гц 50-100 мкВ Дельта-ритм, 0.5-4 Гц 20-200 мкВ Бета-ритм, 14-30 Гц
- 38. Современные методы исследования мозга человека, позволяющие локализовать функции Компьютерная томография Ядерно-магнитный резонанс Магнитоэнцефалография Позитронно-эмиссионная томография
- 41. Компьютерная томография
- 42. Компьютерная томограмма
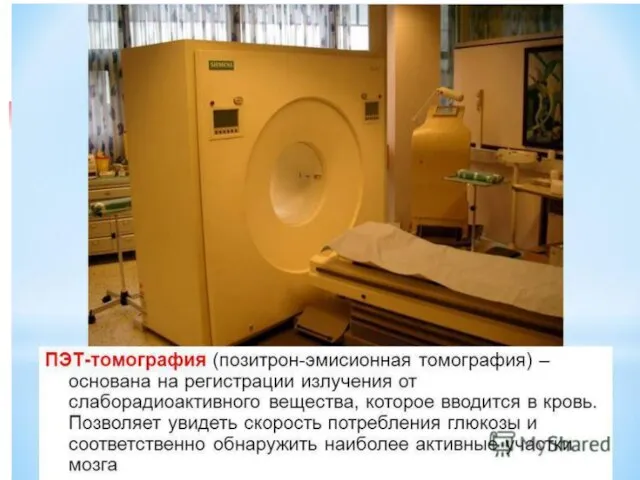
- 43. Позитронно-эмиссионная томография
- 45. Результаты ПЭТ-сканирования ПET - изображение мозга, показывающее зрительное и слуховое распознавание
- 46. Ядерно-магнитный резонанс
- 49. Ядерная магнитно-резонансная интроскопия (ЯМРИ, MRI)
- 50. Послойное сканирование ЯМРИ
- 51. Возникновение современной нейронауки На современном этапе функции нейрофизиологии построены на изучении интегративной деятельности нервной системы и
- 52. Задачи нейронауки Объяснить поведение человека в терминах активности мозга. Понять, как мозг управляет всеми миллионами отдельных
- 54. Молекулярный уровень рассматриваются вопросы морфологии и физиологические особенности нейронов, а также взаимосвязь нейронной организации мозга с
- 55. Клеточный уровень Оценка механизмов восприятия сигналов дендритами, сомами и аксонами нейронов, а также передача их с
- 56. Системный уровень Использование анатомических и физиологических знаний для изучения таких физиологических функций, как рефлексы, сенсорная интеграция,
- 57. Когнитивный уровень Оценка психологических функций с точки зрения формирования нервной системы.
- 58. Основные процессы в нервной системе Трансдукция. Превращение раздражения, внешнего по отношению к самой нервной системе, в
- 59. Основные процессы в нервной системе Моделирование. Построение нервной модели раздражения и/или раздражителя, которая заменяет сам раздражитель.
- 60. Виды модуляции 1. Активация (возбуждение). Повышение активности нервной структуры, повышение её возбуждения и/или возбудимости. Доминантное состояние.
- 61. Виды модуляции 3. Пластическая перестройка нервной структуры. Варианты пластических перестроек: 1) Сенситизация - улучшение передачи возбуждения.
- 62. Виды модуляции 4. Активация исполнительного органа для совершения действия. Таким способом нервная система обеспечивает рефлекторную ответную
- 65. Нейрон и его элементы
- 67. Нейрон – структурно-функциональная единица центральной-нервной системы. Структурно-функциональная организация нервной системы
- 68. Тело (сома) нейрона - центральная часть нейрона, обеспечивающая питанием все остальные части клетки. Строение нейрона
- 69. По отношению к отросткам тело клетки выполняют трофическую функцию -регулирует в них уровень обмена веществ. Вот
- 70. Аксон – одиночный, обычно длинный выходной отросток нейрона, служащий для быстрого проведения возбуждения. По аксону транспортируются
- 72. Строение нейрона Дендриты – древовидно-ветвящиеся отростки нейрона, его главное рецептивное поле, обеспечивающее сбор информации, которая поступает
- 73. Строение нейрона На дендритах имеются микроскопических размеров выросты (шипики), которые значительно увеличивают поверхность соприкосновения с другими
- 74. Виды нейронов А – униполярные: имеют один отросток – аксон. Может быть на промежуточной стадии дифференцировки
- 75. Виды нейронов В –биполярные клетки: имеют два отростка – аксон и дендрит. Г – мультиполярные: один
- 76. Виды нейронов По форме: Пирамидные, Веретенообразные, Звездчатые, Корзинчатые.
- 77. Виды нейронов По локализации: Центральные - лежат в пределах ЦНС. Периферические нейроны принадлежат периферической нервной системе.
- 78. Виды нейронов по выполняемой функции Афферентные (чувствительные) нейроны обеспечивают восприятие раздражения и передачу информации в ЦНС.
- 79. Виды нейронов по выполняемой функции Вставочные нейроны обеспечивают передачу информации внутри ЦНС. В зависимости от эффекта
- 80. Тела нейронов образуют серое вещество головного мозга: кора, подкорковые ядра Отростки нейронов – белое вещество головного
- 81. Глиальные клетки Глиальные клетки в совокупности называют нейроглией. Это клетки, окружающие нейроны и входящие вместе с
- 82. Анатомическое расположение клетки нейроглии в головном мозге; шванновские клетки в периферической нервной системе.
- 83. Функции нейроглии Опорная – поддерживает нервные клетки Изолирующая – препятствует переходу нервных импульсов с тела одного
- 84. Функции нейроглии Участие в регенерации. В отличие от нейронов, глиальные клетки сохраняют способность к делению в
- 85. Виды глиальных клеток Эпиндемоциты - выстилают полости внутри мозга и имеют цилии помогающие циркуляции спиномозговой жидкости.
- 86. Астроциты Функции Гомеостатическая (поддержание ионного и химического состава среды) Метаболическая (синтез и разложение веществ) Трофическая (влияние
- 87. Олигодендроциты Находятся преимущественно в белом веществе мозга, где они образуют миелин вокруг крупных аксонов.
- 88. Радиальная глия Участвует в миграции нейронов во время внутриутробного развития плода
- 89. Тело нейрона покрыто слоистой мембраной, которая представляет собой два слоя липидов с противоположной ориентацией, образующих матрикс,
- 91. Строение нейрона Белки мембраны: Интегральные – ионные каналы Поверхностные - рецепторы
- 92. Структуры, поддерживающие гомеостаз нейрона: Мембрана – отделяет внутриклеточную среду от внеклеточной и разделяет клетку на функциональные
- 93. Схема ионного канала
- 94. Функция канала
- 95. Основные функции мембраны Образует полупроницаемый, избирательный барьер между цитоплазмой и внешней средой Проницаемость мембраны управляется ее
- 96. Ионные каналы 1. Пассивные (для калия) 2. Управляемые: потенциалзависимые; хемозависимые; механические
- 98. Мембрана называется полупроницаемой, потому что: Мембрана ограничивает движение ионов какого-либо вида, т.е. одни ионы проходят через
- 99. Благодаря этим свойствам мембрана создает и поддерживает Химическую (концентрационную) асимметрию ионов по обе стороны мембраны. Электрический
- 100. В покое мембрана более проницаема для ионов К+
- 101. Концентрационный градиент Различная концентрация ионов является движущей силой для их диффузии в область меньшей концентрации. Катионы
- 102. Возникает и поддерживается МП МП = ПП = Пассивный К+ ток. МП регистрируют с помощью микроэлектродной
- 103. ГЕНЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ
- 104. Основной принцип функционирования центральной нервной системы – способность мембраны нейронов генерировать и передавать возбуждение. В формировании
- 105. Мембранный потенциал это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью мембраны.
- 106. Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому,
- 108. Потенциал покоя нейрона Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет
- 109. Потенциал покоя нейрона Снаружи — преобладают ионы натрия, кальция и хлора, внутри — ионы калия и
- 110. Под воздействием стимула происходит открытие натриевых каналов. По градиенту концентраций положительно заряженные ионы натрия устремляются на
- 111. Потенциал действия нейрона Исходную концентрацию ионов восстанавливает натрий-калиевый насос. Длительность ПД составляет около 1 мс. Амплитуда
- 112. Как распространяется возбуждение (потенциал действия) в нервной ткани? Развития современных представлений о биоэлектрических явления способствовали исследования
- 113. Согласно этой теории биоэлектрических потенциалы обусловлены неодинаковой различной проницаемостью для них поверхностной мембраны
- 118. Потенциал действия (ПД) или Нервный импульс
- 119. Потенциал действия нейрона
- 120. ПД и фазы
- 121. Распространение ПД по волокну
- 122. В немиелинизированных нервных волокнах возбуждение распространяется посредством локальных токов очень маленькими шажками. Для немиелинизированных волокон средняя
- 123. В миелинизированных волокнах, где поперечное сопротивление увеличивается за счет наличия дополнительной миелинизированной муфты с высоким сопротивлением,
- 125. Потенциал действия Возникновению потенциала действия предшествуют активные изменения мембранного потенциала, которые обусловлены повышением натриевой проницаемости мембраны.
- 127. В практической медицине используют эти фундаментальные знания Местные анестетики блокируют натриевые каналы. Натрий не входит в
- 128. Закономерности проведения возбуждения по нервным волокнам Двустороннее проведение возбуждения. Изолированное проведение возбуждения в отдельных нервных волокнах.
- 129. Контрольные вопросы
- 131. Кто впервые определил наличие токов в ЦНС А – Бергер Б – Декарт В – Кэтон
- 132. Чему равен потенциал покоя мембраны нейрона? А. -70 мВ Б . +20 мВ
- 134. Скачать презентацию

План лекции
Введение
История развития нейрофизиологии
Основные процессы в нервной системе
Нейрон – как структурно-функциональная
План лекции
Введение
История развития нейрофизиологии
Основные процессы в нервной системе
Нейрон – как структурно-функциональная

РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ

Крылова Н.В., Искренко И.А. Мозг и проводящие пути: Анатомия человека в
Крылова Н.В., Искренко И.А. Мозг и проводящие пути: Анатомия человека в

Достижения человечества существуют благодаря разуму, вместилищем которого является головной мозг.
Все продукты
Достижения человечества существуют благодаря разуму, вместилищем которого является головной мозг.
Все продукты

Введение
Нейрофизиология относится к разделу физиологии, который изучает функции нервной системы, в
Введение
Нейрофизиология относится к разделу физиологии, который изучает функции нервной системы, в

Разделы нейрофизиологии
Общая нейрофизиология - изучает закономерности функционирования нервной системы на разных
Разделы нейрофизиологии
Общая нейрофизиология - изучает закономерности функционирования нервной системы на разных

История открытия и развитие представлений о биопотенциалах головного мозга
История открытия и развитие представлений о биопотенциалах головного мозга
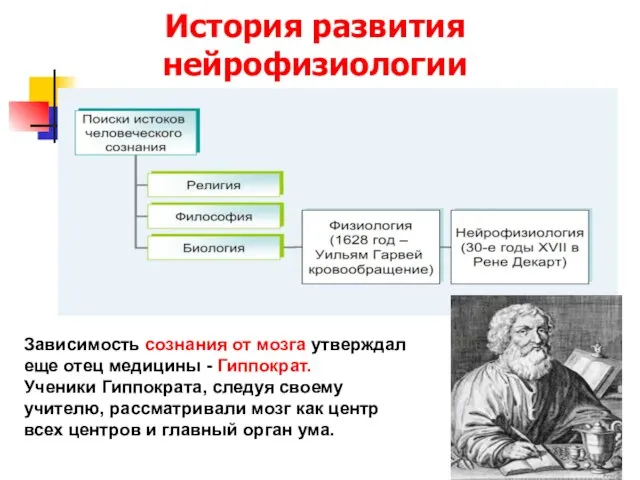
История развития нейрофизиологии
Зависимость сознания от мозга утверждал еще отец медицины -
История развития нейрофизиологии
Зависимость сознания от мозга утверждал еще отец медицины -

Первые исследования
около 620-550 до н.э. – Фалес Милетский - считается первооткрывателем
Первые исследования
около 620-550 до н.э. – Фалес Милетский - считается первооткрывателем
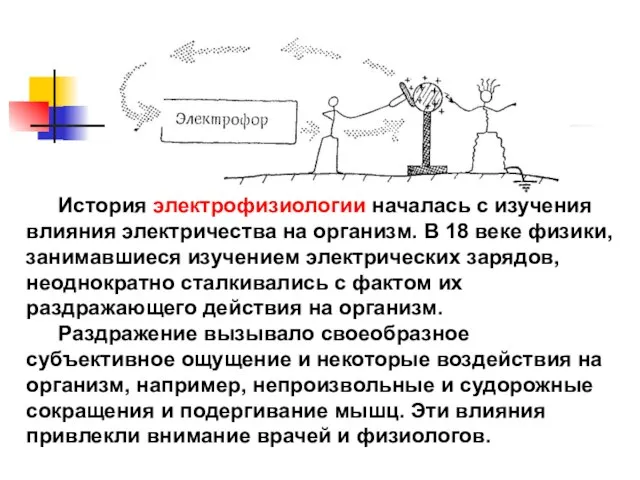
История электрофизиологии началась с изучения влияния электричества на организм. В 18
История электрофизиологии началась с изучения влияния электричества на организм. В 18

История развития нейрофизиологии
Первые представления о рефлекторном принципе действия нервной системы человека
История развития нейрофизиологии
Первые представления о рефлекторном принципе действия нервной системы человека

Начало электрофизиологии обычно связывают со знаменитыми опытами итальянского врача, анатома и
Начало электрофизиологии обычно связывают со знаменитыми опытами итальянского врача, анатома и
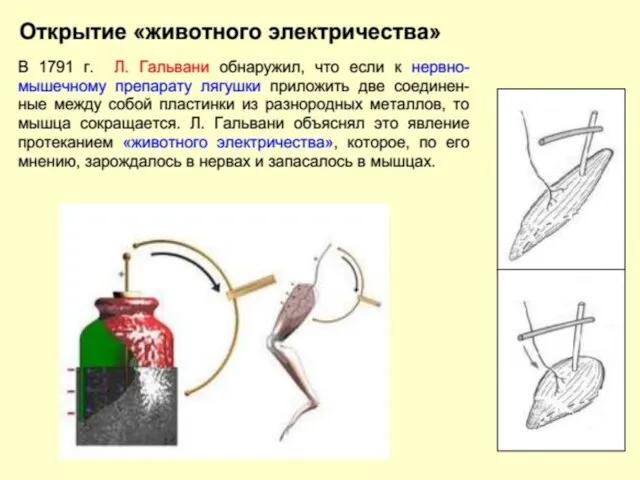

Карло Маттеучи (Carlo Matteucci 1811 – 1868) в 1830-1840 годах показал,
Карло Маттеучи (Carlo Matteucci 1811 – 1868) в 1830-1840 годах показал,


Опыт по электрофизиологии нервов и мышц лягушки [Du Bois-Reymond: Frog Experiment
Опыт по электрофизиологии нервов и мышц лягушки [Du Bois-Reymond: Frog Experiment

В 1875 году английский хирург и физиолог Ричард Кэтон на заседании
В 1875 году английский хирург и физиолог Ричард Кэтон на заседании

Василий Яковлевич
Данилевский
В 1875 г. В.Я. Данилевский изложил данные по изучению
Василий Яковлевич
Данилевский
В 1875 г. В.Я. Данилевский изложил данные по изучению

Иван Михайлович
Сеченов
1864 г. «Рефлексы головного мозга»
В 1882 году И.М.Сеченов опубликовал
Иван Михайлович
Сеченов
1864 г. «Рефлексы головного мозга»
В 1882 году И.М.Сеченов опубликовал

Иван Михайлович Сеченов
1829 – 1905
В 1863 г. опубликовал книгу
Иван Михайлович Сеченов
1829 – 1905
В 1863 г. опубликовал книгу

Он обосновал возможность распространения принципа рефлекса как принципа организации поведения на
Он обосновал возможность распространения принципа рефлекса как принципа организации поведения на

В 1884 году Н.Е. Введенский в работе “Исследования над нервными центрами”
В 1884 году Н.Е. Введенский в работе “Исследования над нервными центрами”

Б.Ф. Вериго
В 1889 г. В журнале «Вестник клинической и судебной психиатрии
Б.Ф. Вериго
В 1889 г. В журнале «Вестник клинической и судебной психиатрии

1890 год - работа А. Бека, посвященная использованию метода регистрации электрической
1890 год - работа А. Бека, посвященная использованию метода регистрации электрической

1898 год - В.Е. Ларионов производит исследования электрических явлений в головном
1898 год - В.Е. Ларионов производит исследования электрических явлений в головном
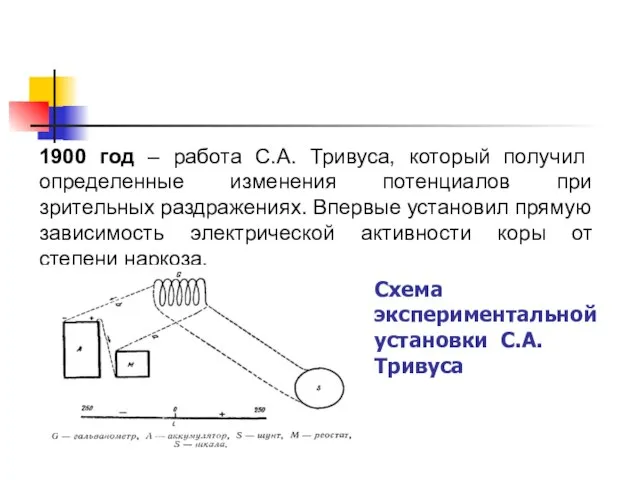
1900 год – работа С.А. Тривуса, который получил определенные изменения потенциалов
1900 год – работа С.А. Тривуса, который получил определенные изменения потенциалов

1912 год - обширное исследование П.Ю. Кауфмана, который в тщательных опытах
1912 год - обширное исследование П.Ю. Кауфмана, который в тщательных опытах

Первая запись биотоков головного мозга человека получена Гансом Бергером в 1925
Первая запись биотоков головного мозга человека получена Гансом Бергером в 1925

Нейрофизиологические методы:
Внутриклеточная регистрация активности нервных клеток (ПП, ПД)
Внеклеточная регистрация нейронной активности
Нейрофизиологические методы:
Внутриклеточная регистрация активности нервных клеток (ПП, ПД)
Внеклеточная регистрация нейронной активности
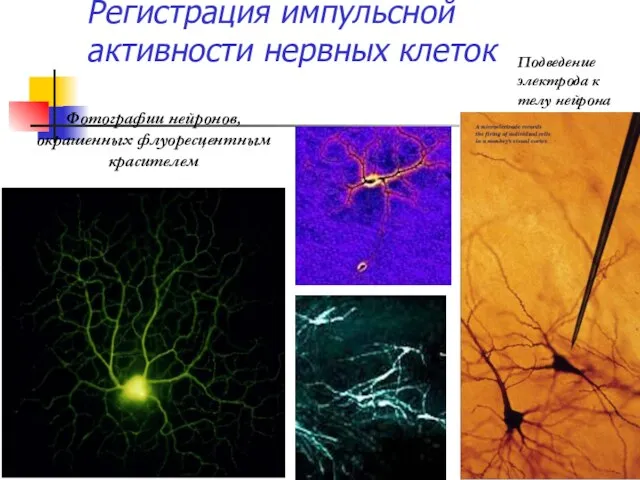
Регистрация импульсной активности нервных клеток
Фотографии нейронов, окрашенных флуоресцентным красителем
Подведение электрода к
Регистрация импульсной активности нервных клеток
Фотографии нейронов, окрашенных флуоресцентным красителем
Подведение электрода к
Два подхода к изучению мозга
Гемодинамика (метаболизм) – ПЭТ, ЯМРИ
Электрофизиология – ЭЭГ,
Два подхода к изучению мозга
Гемодинамика (метаболизм) – ПЭТ, ЯМРИ
Электрофизиология – ЭЭГ,

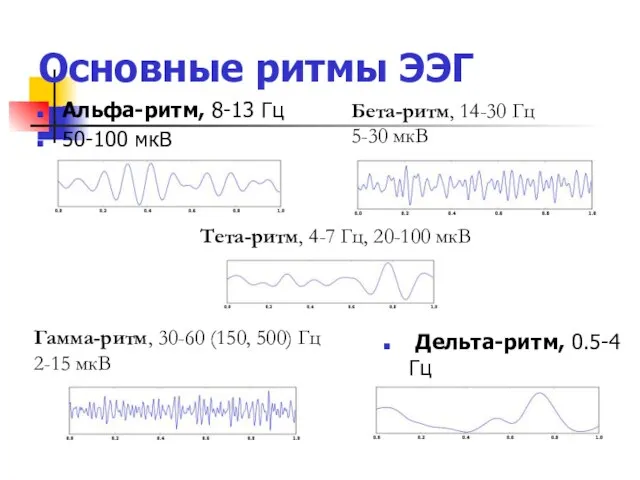
Основные ритмы ЭЭГ
Альфа-ритм, 8-13 Гц
50-100 мкВ
Дельта-ритм, 0.5-4 Гц
20-200 мкВ
Бета-ритм, 14-30
Основные ритмы ЭЭГ
Альфа-ритм, 8-13 Гц
50-100 мкВ
Дельта-ритм, 0.5-4 Гц
20-200 мкВ
Бета-ритм, 14-30


Современные методы исследования мозга человека, позволяющие локализовать функции
Компьютерная томография
Ядерно-магнитный резонанс
Магнитоэнцефалография
Позитронно-эмиссионная томография
Современные методы исследования мозга человека, позволяющие локализовать функции
Компьютерная томография
Ядерно-магнитный резонанс
Магнитоэнцефалография
Позитронно-эмиссионная томография


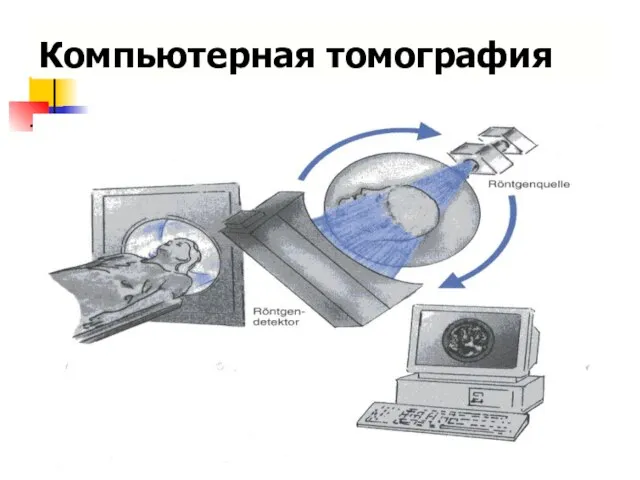
Компьютерная томография
Компьютерная томография
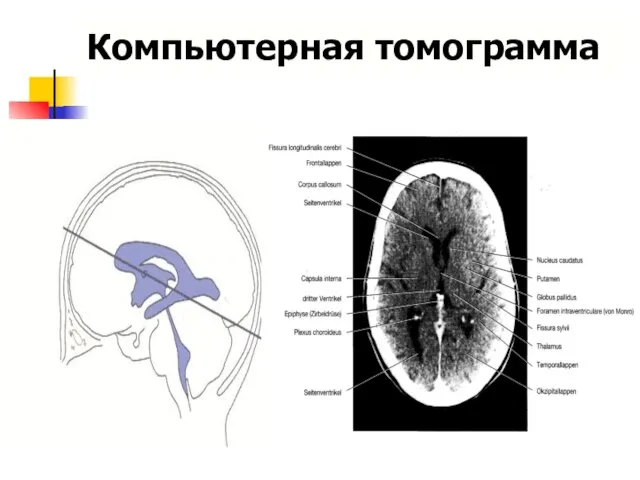
Компьютерная томограмма
Компьютерная томограмма
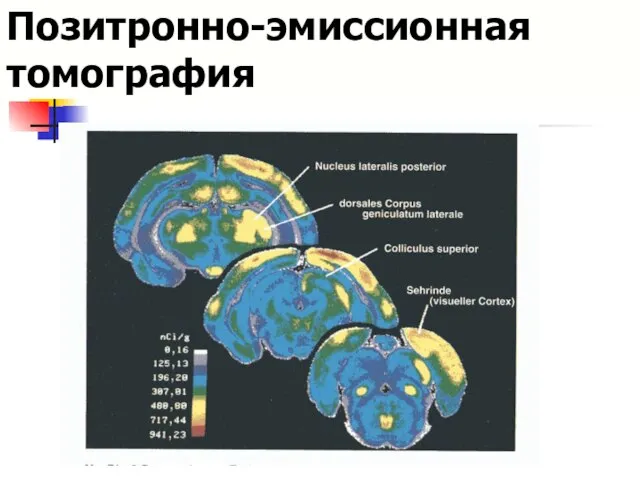
Позитронно-эмиссионная томография
Позитронно-эмиссионная томография
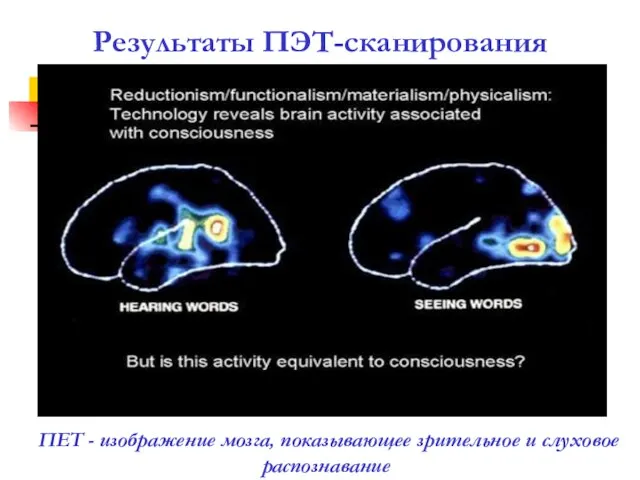
Результаты ПЭТ-сканирования
ПET - изображение мозга, показывающее зрительное и слуховое распознавание
Результаты ПЭТ-сканирования
ПET - изображение мозга, показывающее зрительное и слуховое распознавание
Ядерно-магнитный резонанс
Ядерно-магнитный резонанс

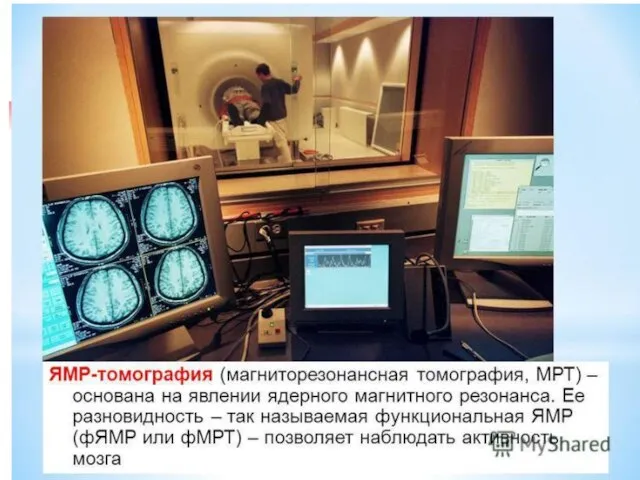
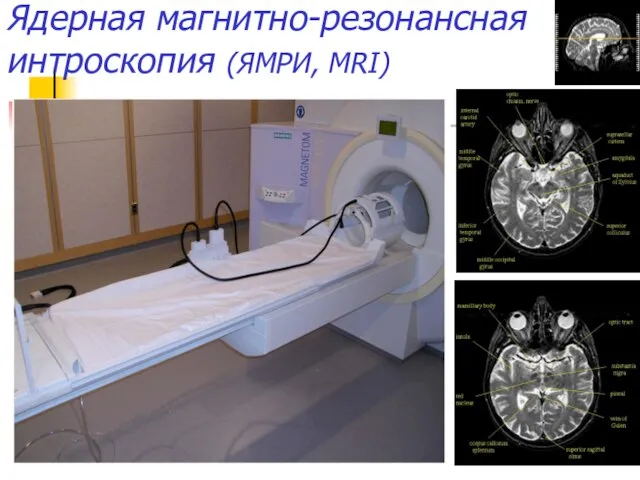
Ядерная магнитно-резонансная интроскопия (ЯМРИ, MRI)
Ядерная магнитно-резонансная интроскопия (ЯМРИ, MRI)
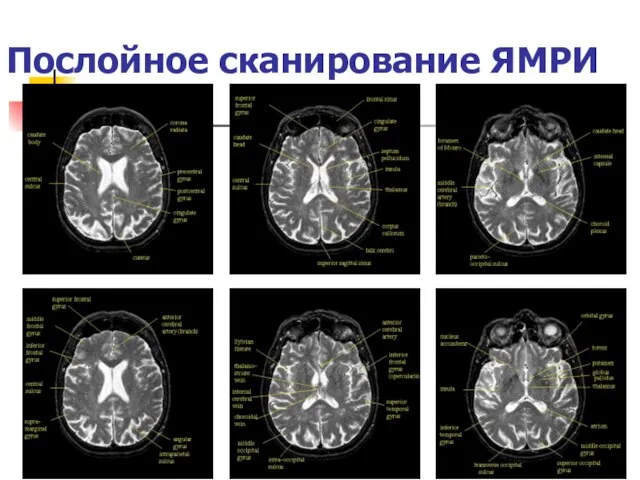
Послойное сканирование ЯМРИ
Послойное сканирование ЯМРИ
Возникновение современной нейронауки
На современном этапе функции нейрофизиологии построены на изучении интегративной
Возникновение современной нейронауки
На современном этапе функции нейрофизиологии построены на изучении интегративной

Задачи нейронауки
Объяснить поведение человека в терминах активности мозга.
Понять, как
Задачи нейронауки
Объяснить поведение человека в терминах активности мозга.
Понять, как
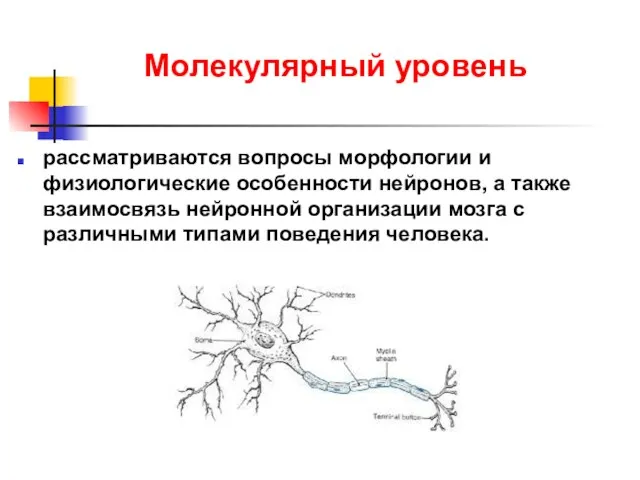
Молекулярный уровень
рассматриваются вопросы морфологии и физиологические особенности нейронов, а также взаимосвязь
Молекулярный уровень
рассматриваются вопросы морфологии и физиологические особенности нейронов, а также взаимосвязь

Клеточный уровень
Оценка механизмов восприятия сигналов дендритами, сомами и аксонами нейронов, а
Клеточный уровень
Оценка механизмов восприятия сигналов дендритами, сомами и аксонами нейронов, а
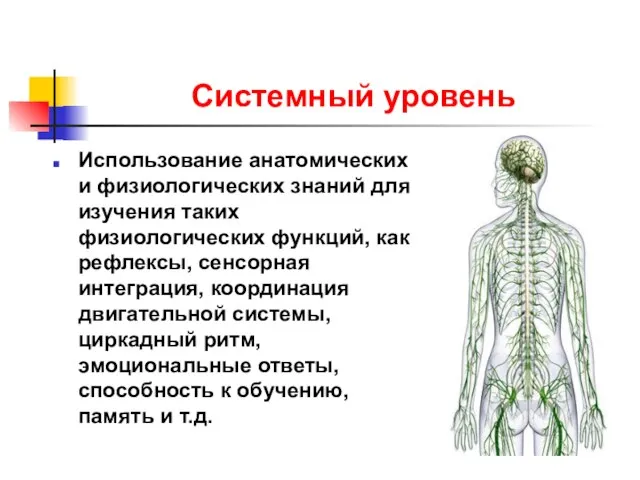
Системный уровень
Использование анатомических и физиологических знаний для изучения таких физиологических функций,
Системный уровень
Использование анатомических и физиологических знаний для изучения таких физиологических функций,

Когнитивный уровень
Оценка психологических функций с точки зрения формирования нервной системы.
Когнитивный уровень
Оценка психологических функций с точки зрения формирования нервной системы.

Основные процессы в нервной системе
Трансдукция. Превращение раздражения, внешнего по отношению к
Основные процессы в нервной системе
Трансдукция. Превращение раздражения, внешнего по отношению к

Основные процессы в нервной системе
Моделирование. Построение нервной модели раздражения и/или раздражителя,
Основные процессы в нервной системе
Моделирование. Построение нервной модели раздражения и/или раздражителя,

Виды модуляции
1. Активация (возбуждение). Повышение активности нервной структуры, повышение её
Виды модуляции
1. Активация (возбуждение). Повышение активности нервной структуры, повышение её

Виды модуляции
3. Пластическая перестройка нервной структуры.
Варианты пластических перестроек:
1) Сенситизация -
Виды модуляции
3. Пластическая перестройка нервной структуры. Варианты пластических перестроек: 1) Сенситизация -

Виды модуляции
4. Активация исполнительного органа для совершения действия.
Таким способом нервная
Виды модуляции
4. Активация исполнительного органа для совершения действия.
Таким способом нервная
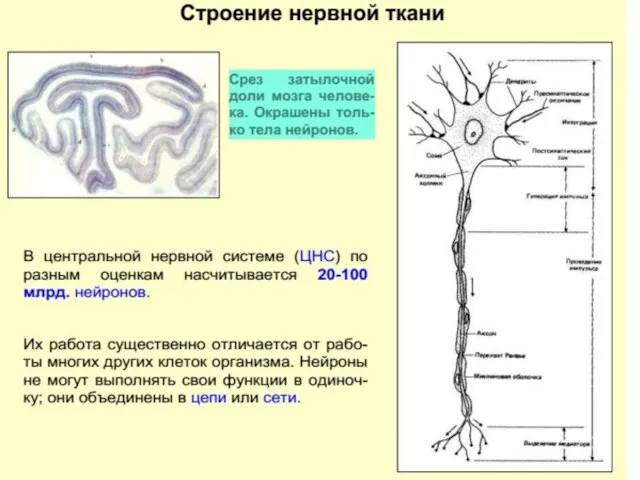
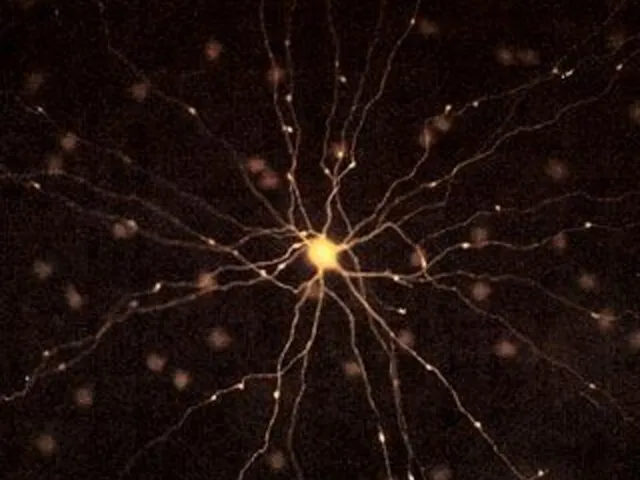
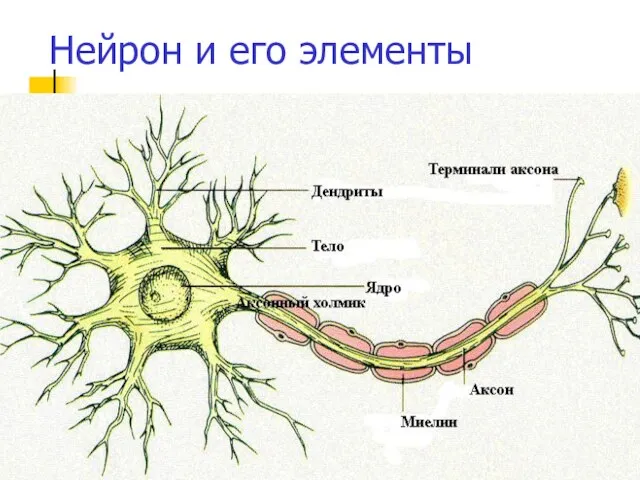
Нейрон и его элементы
Нейрон и его элементы
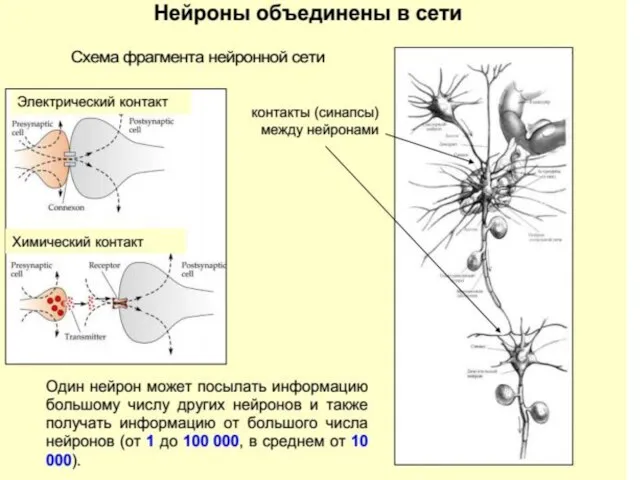

Нейрон – структурно-функциональная единица центральной-нервной системы.
Структурно-функциональная организация нервной системы
Нейрон – структурно-функциональная единица центральной-нервной системы.
Структурно-функциональная организация нервной системы
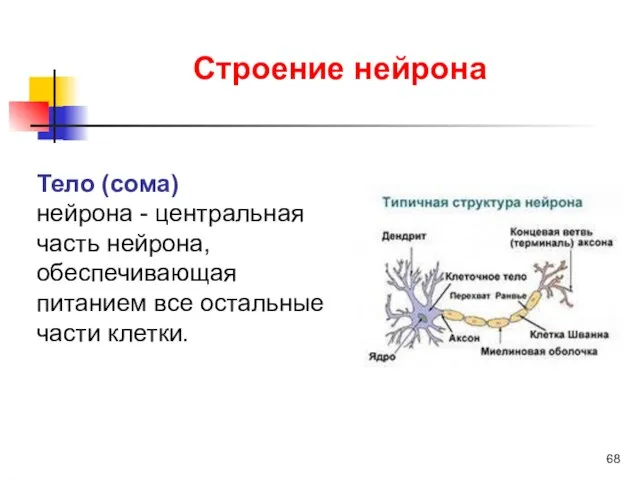
Тело (сома)
нейрона - центральная часть нейрона, обеспечивающая питанием все остальные
Тело (сома)
нейрона - центральная часть нейрона, обеспечивающая питанием все остальные

По отношению к отросткам тело клетки выполняют трофическую функцию -регулирует в
По отношению к отросткам тело клетки выполняют трофическую функцию -регулирует в

Аксон – одиночный, обычно длинный выходной отросток нейрона, служащий для быстрого
Аксон – одиночный, обычно длинный выходной отросток нейрона, служащий для быстрого
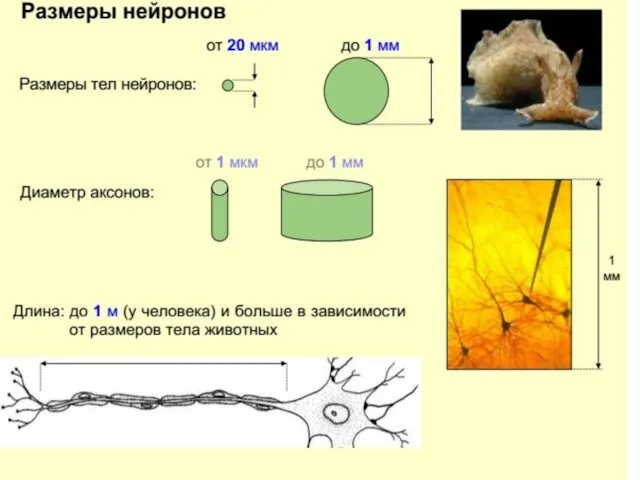

Строение нейрона
Дендриты – древовидно-ветвящиеся отростки нейрона, его главное рецептивное поле,
Строение нейрона
Дендриты – древовидно-ветвящиеся отростки нейрона, его главное рецептивное поле,

Строение нейрона
На дендритах имеются микроскопических размеров выросты (шипики), которые значительно увеличивают
Строение нейрона
На дендритах имеются микроскопических размеров выросты (шипики), которые значительно увеличивают
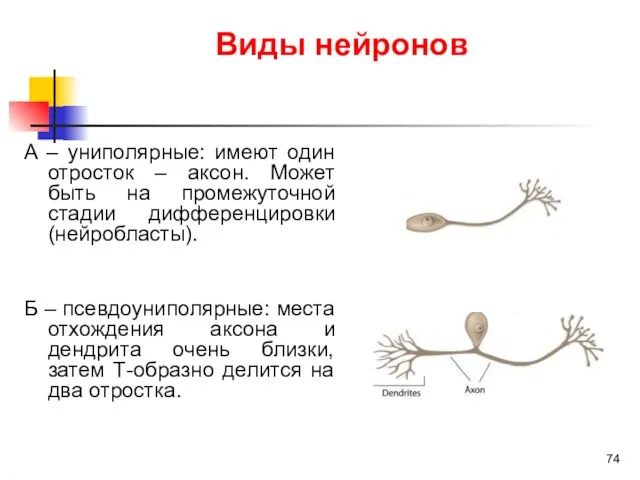
Виды нейронов
А – униполярные: имеют один отросток – аксон. Может быть
Виды нейронов
А – униполярные: имеют один отросток – аксон. Может быть
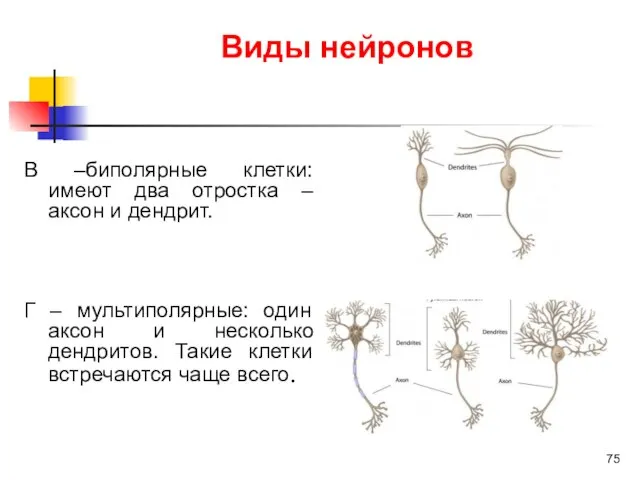
Виды нейронов
В –биполярные клетки: имеют два отростка – аксон и дендрит.
Г
Виды нейронов
В –биполярные клетки: имеют два отростка – аксон и дендрит.
Г

Виды нейронов
По форме:
Пирамидные,
Веретенообразные,
Звездчатые,
Корзинчатые.
Виды нейронов
По форме:
Пирамидные,
Веретенообразные,
Звездчатые,
Корзинчатые.

Виды нейронов
По локализации:
Центральные - лежат в пределах ЦНС.
Периферические нейроны
Виды нейронов
По локализации:
Центральные - лежат в пределах ЦНС.
Периферические нейроны
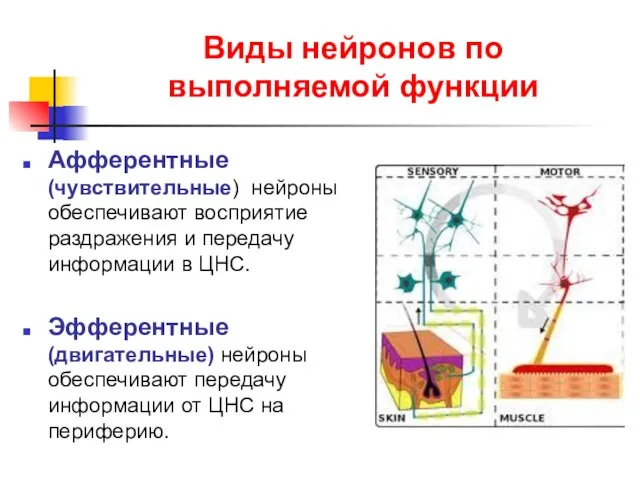
Виды нейронов по выполняемой функции
Афферентные (чувствительные) нейроны обеспечивают восприятие раздражения и
Виды нейронов по выполняемой функции
Афферентные (чувствительные) нейроны обеспечивают восприятие раздражения и

Виды нейронов по выполняемой функции
Вставочные нейроны обеспечивают передачу информации внутри ЦНС.
В
Виды нейронов по выполняемой функции
Вставочные нейроны обеспечивают передачу информации внутри ЦНС.
В
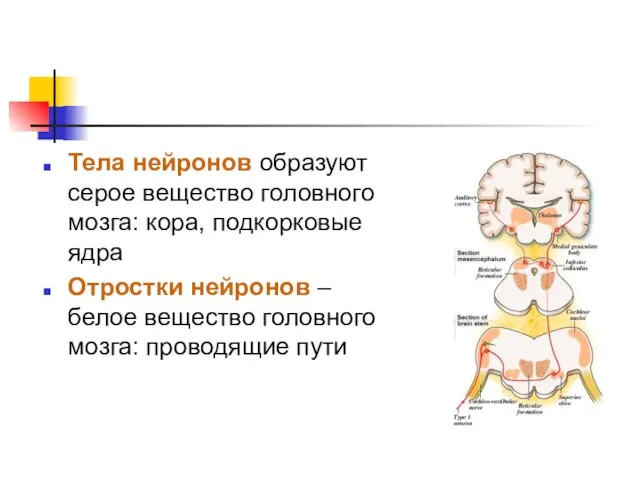
Тела нейронов образуют серое вещество головного мозга: кора, подкорковые ядра
Отростки нейронов
Тела нейронов образуют серое вещество головного мозга: кора, подкорковые ядра
Отростки нейронов
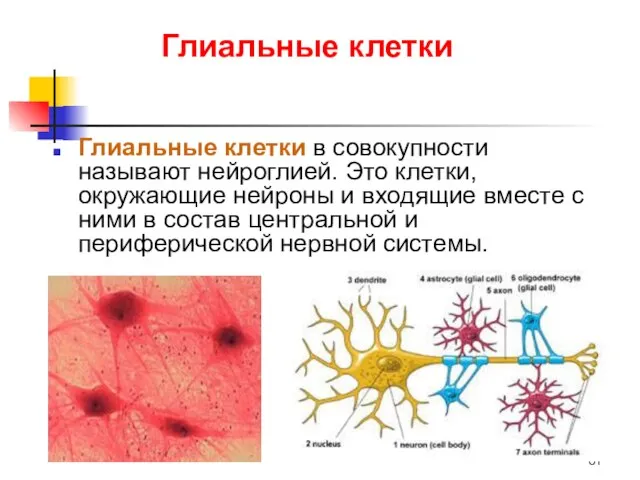
Глиальные клетки
Глиальные клетки в совокупности называют нейроглией. Это клетки, окружающие нейроны
Глиальные клетки
Глиальные клетки в совокупности называют нейроглией. Это клетки, окружающие нейроны

Анатомическое расположение
клетки нейроглии в головном мозге;
шванновские клетки в периферической нервной системе.
Анатомическое расположение
клетки нейроглии в головном мозге;
шванновские клетки в периферической нервной системе.

Функции нейроглии
Опорная – поддерживает нервные клетки
Изолирующая – препятствует переходу нервных импульсов
Функции нейроглии
Опорная – поддерживает нервные клетки
Изолирующая – препятствует переходу нервных импульсов

Функции нейроглии
Участие в регенерации. В отличие от нейронов, глиальные клетки сохраняют
Функции нейроглии
Участие в регенерации. В отличие от нейронов, глиальные клетки сохраняют

Виды глиальных клеток
Эпиндемоциты - выстилают полости внутри мозга и имеют цилии
Виды глиальных клеток
Эпиндемоциты - выстилают полости внутри мозга и имеют цилии
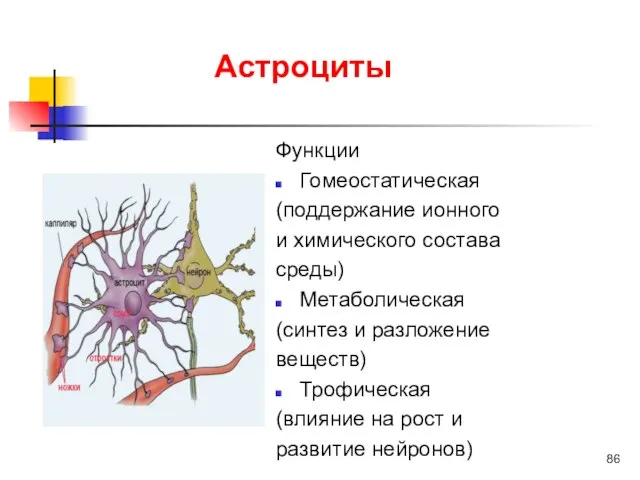
Астроциты
Функции
Гомеостатическая
(поддержание ионного
и химического состава
среды)
Метаболическая
(синтез и разложение
веществ)
Трофическая
(влияние на рост и
развитие нейронов)
Астроциты
Функции
Гомеостатическая
(поддержание ионного
и химического состава
среды)
Метаболическая
(синтез и разложение
веществ)
Трофическая
(влияние на рост и
развитие нейронов)
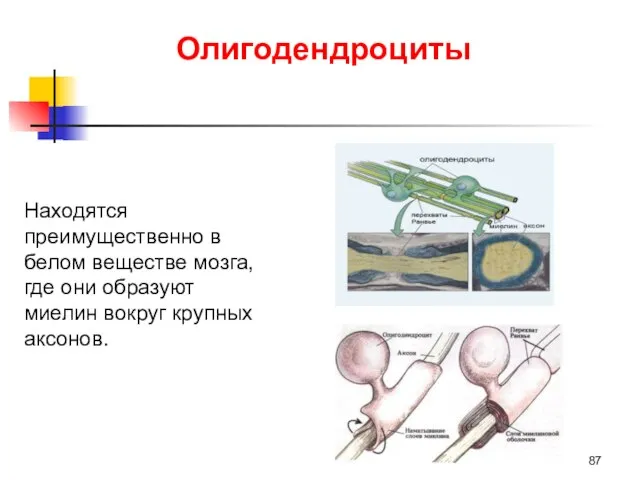
Олигодендроциты
Находятся преимущественно в белом веществе мозга, где они образуют миелин вокруг
Олигодендроциты
Находятся преимущественно в белом веществе мозга, где они образуют миелин вокруг
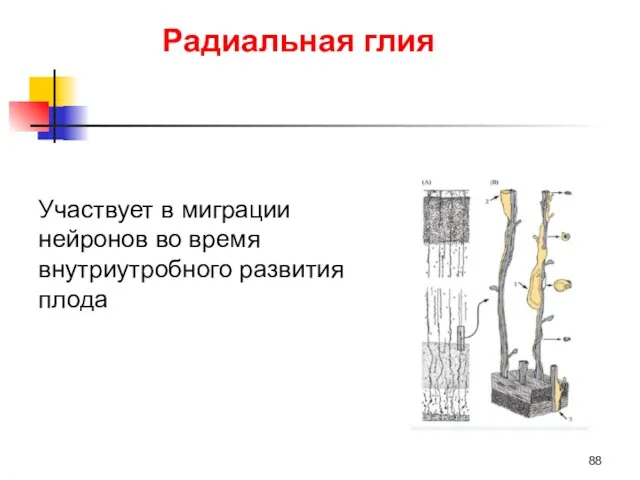
Радиальная глия
Участвует в миграции нейронов во время внутриутробного развития плода
Радиальная глия
Участвует в миграции нейронов во время внутриутробного развития плода
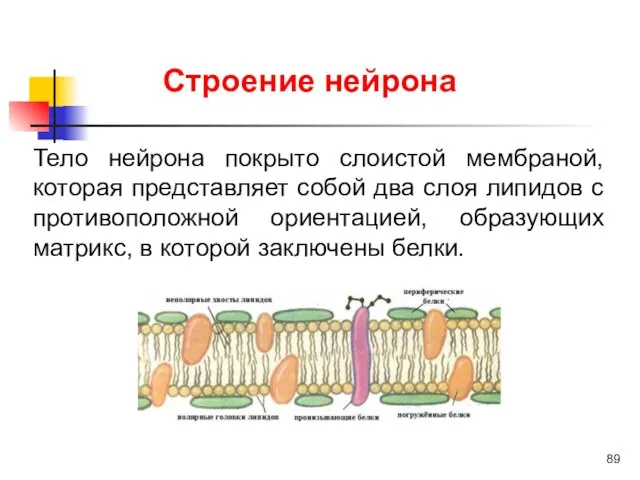
Тело нейрона покрыто слоистой мембраной, которая представляет собой два слоя липидов
Тело нейрона покрыто слоистой мембраной, которая представляет собой два слоя липидов
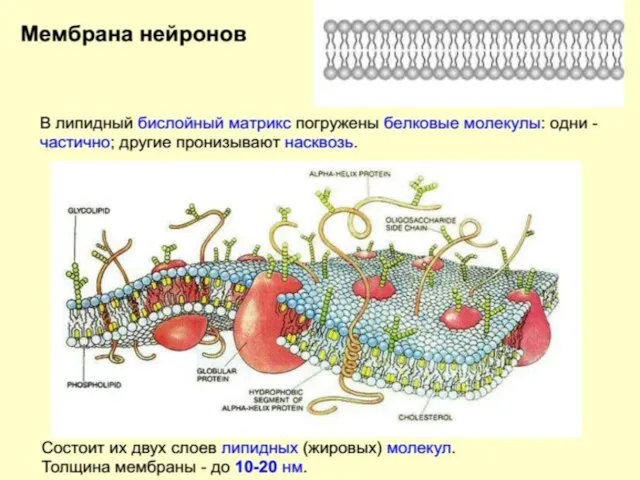

Строение нейрона
Белки мембраны:
Интегральные – ионные каналы
Поверхностные - рецепторы
Строение нейрона
Белки мембраны:
Интегральные – ионные каналы
Поверхностные - рецепторы

Структуры, поддерживающие гомеостаз нейрона:
Мембрана – отделяет внутриклеточную среду от внеклеточной и
Структуры, поддерживающие гомеостаз нейрона:
Мембрана – отделяет внутриклеточную среду от внеклеточной и
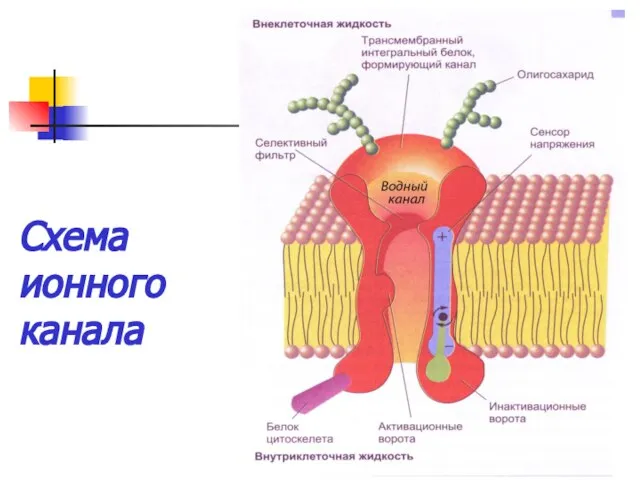
Схема ионного канала
Схема ионного канала
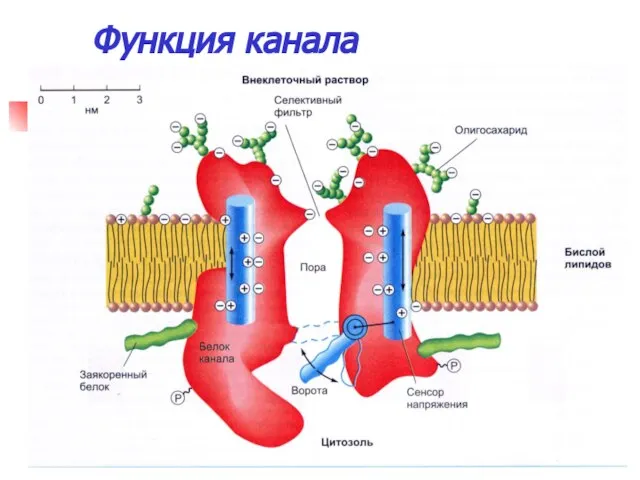
Функция канала
Функция канала

Основные функции мембраны
Образует полупроницаемый, избирательный барьер между цитоплазмой и внешней средой
Основные функции мембраны
Образует полупроницаемый, избирательный барьер между цитоплазмой и внешней средой

Ионные каналы
1. Пассивные (для калия)
2. Управляемые:
потенциалзависимые;
хемозависимые;
механические
Ионные каналы
1. Пассивные (для калия)
2. Управляемые:
потенциалзависимые;
хемозависимые;
механические
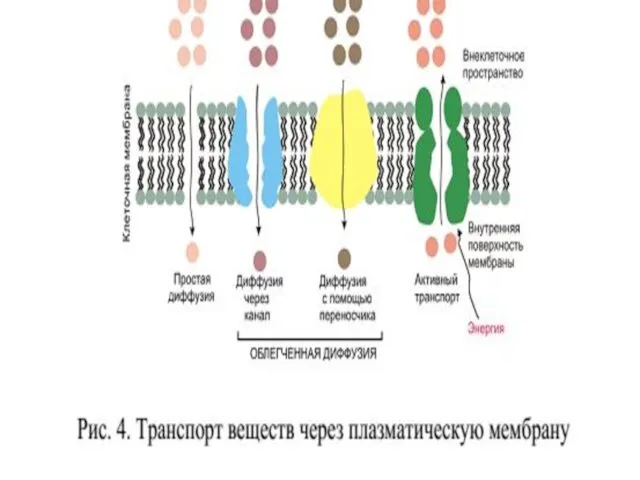

Мембрана называется полупроницаемой, потому что:
Мембрана ограничивает движение ионов какого-либо вида, т.е.
Мембрана называется полупроницаемой, потому что:
Мембрана ограничивает движение ионов какого-либо вида, т.е.

Благодаря этим свойствам мембрана создает и поддерживает
Химическую (концентрационную) асимметрию ионов по
Благодаря этим свойствам мембрана создает и поддерживает
Химическую (концентрационную) асимметрию ионов по
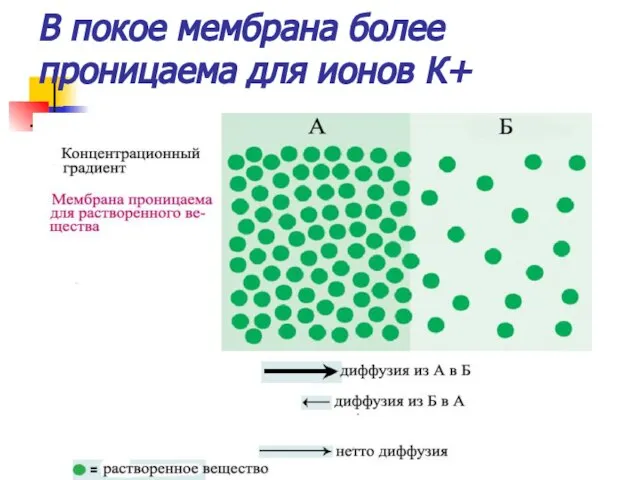
В покое мембрана более проницаема для ионов К+
В покое мембрана более проницаема для ионов К+

Концентрационный градиент
Различная концентрация ионов является движущей силой для их диффузии в
Концентрационный градиент
Различная концентрация ионов является движущей силой для их диффузии в

Возникает и поддерживается МП
МП = ПП = Пассивный К+ ток.
МП регистрируют
Возникает и поддерживается МП
МП = ПП = Пассивный К+ ток.
МП регистрируют

ГЕНЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ
ГЕНЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ

Основной принцип функционирования центральной нервной системы – способность мембраны нейронов генерировать
Основной принцип функционирования центральной нервной системы – способность мембраны нейронов генерировать

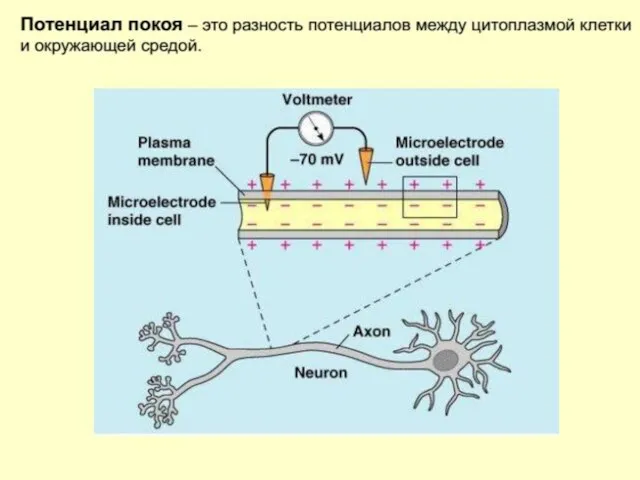
Мембранный потенциал
это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью
Мембранный потенциал
это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью

Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению
Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению
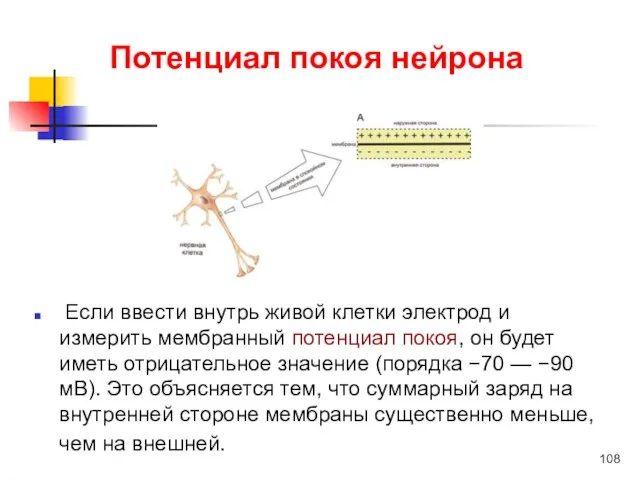
Потенциал покоя нейрона
Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя,
Потенциал покоя нейрона
Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя,
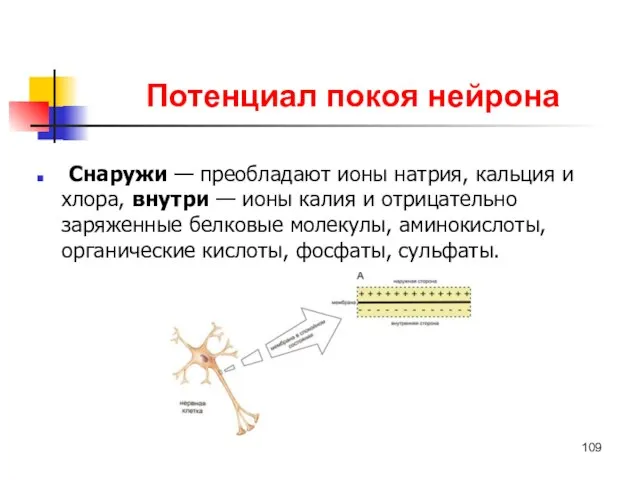
Потенциал покоя нейрона
Снаружи — преобладают ионы натрия, кальция и хлора, внутри — ионы калия и отрицательно
Потенциал покоя нейрона
Снаружи — преобладают ионы натрия, кальция и хлора, внутри — ионы калия и отрицательно
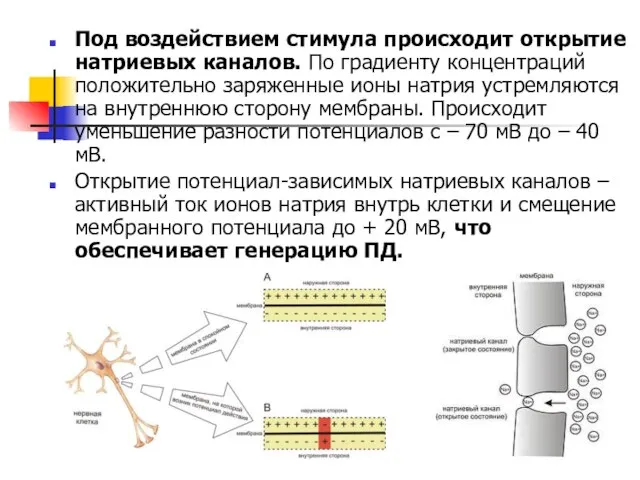
Под воздействием стимула происходит открытие натриевых каналов. По градиенту концентраций положительно
Под воздействием стимула происходит открытие натриевых каналов. По градиенту концентраций положительно

Потенциал действия нейрона
Исходную концентрацию ионов восстанавливает натрий-калиевый насос.
Длительность ПД составляет около
Потенциал действия нейрона
Исходную концентрацию ионов восстанавливает натрий-калиевый насос.
Длительность ПД составляет около

Как распространяется возбуждение (потенциал действия) в нервной ткани?
Развития современных представлений
Как распространяется возбуждение (потенциал действия) в нервной ткани?
Развития современных представлений

Согласно этой теории биоэлектрических потенциалы обусловлены неодинаковой различной проницаемостью для них
Согласно этой теории биоэлектрических потенциалы обусловлены неодинаковой различной проницаемостью для них

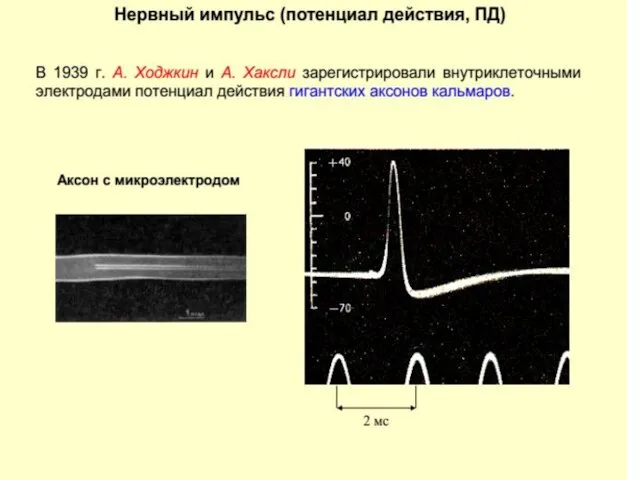
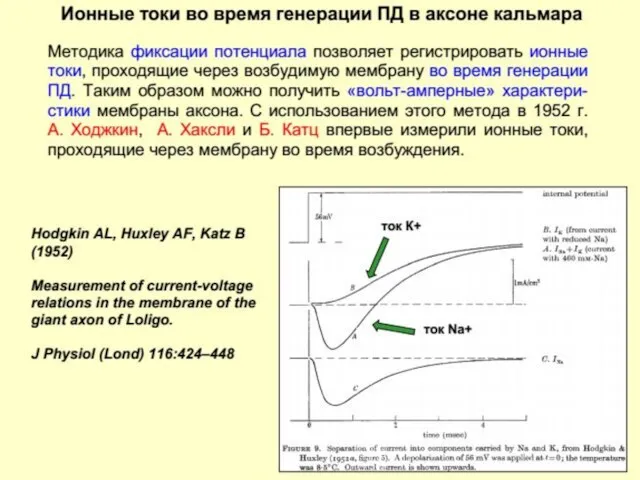

Потенциал действия (ПД)
или
Нервный импульс
Потенциал действия (ПД)
или
Нервный импульс
Потенциал действия нейрона
Потенциал действия нейрона
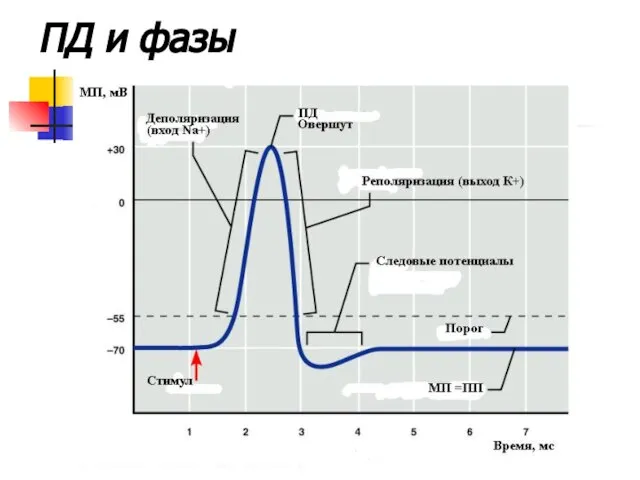
ПД и фазы
ПД и фазы
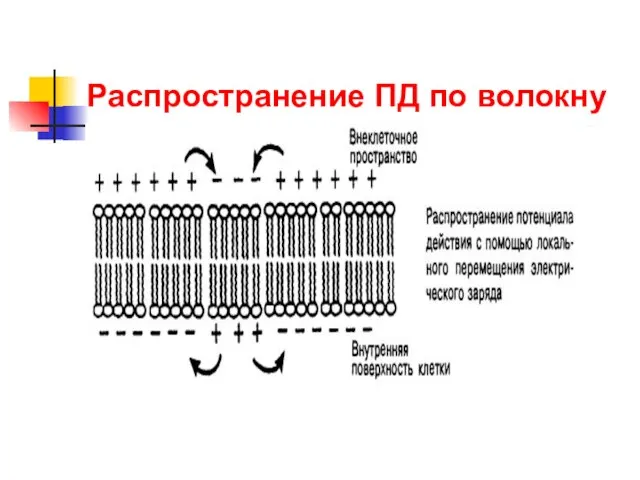
Распространение ПД по волокну
Распространение ПД по волокну

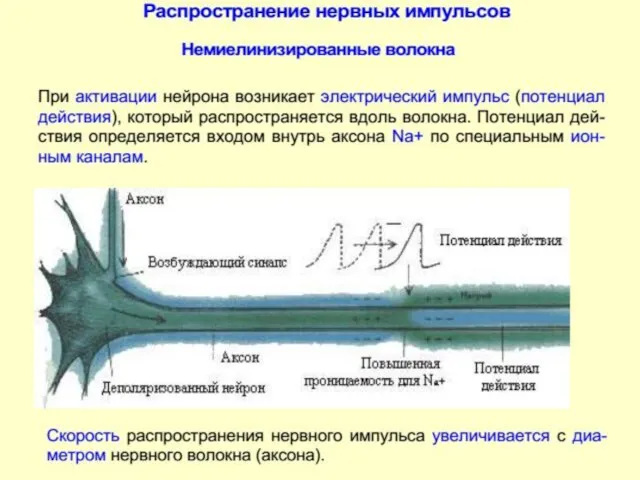
В немиелинизированных нервных волокнах возбуждение распространяется посредством локальных токов очень маленькими
В немиелинизированных нервных волокнах возбуждение распространяется посредством локальных токов очень маленькими

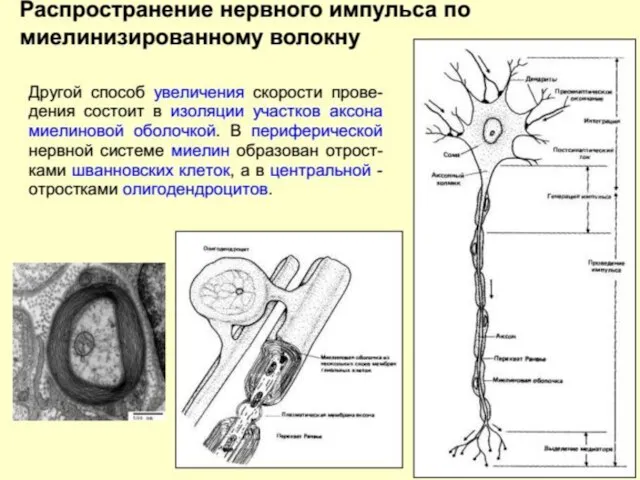
В миелинизированных волокнах, где поперечное сопротивление увеличивается за счет наличия дополнительной
В миелинизированных волокнах, где поперечное сопротивление увеличивается за счет наличия дополнительной

Потенциал действия
Возникновению потенциала действия предшествуют активные изменения мембранного потенциала, которые обусловлены
Потенциал действия
Возникновению потенциала действия предшествуют активные изменения мембранного потенциала, которые обусловлены

В практической медицине используют эти фундаментальные знания
Местные анестетики блокируют натриевые каналы.
Натрий
В практической медицине используют эти фундаментальные знания
Местные анестетики блокируют натриевые каналы.
Натрий

Закономерности проведения возбуждения по нервным волокнам
Двустороннее проведение возбуждения.
Изолированное проведение возбуждения в
Закономерности проведения возбуждения по нервным волокнам
Двустороннее проведение возбуждения.
Изолированное проведение возбуждения в

Контрольные вопросы
Контрольные вопросы


Кто впервые определил наличие токов в ЦНС
А – Бергер
Б – Декарт
В
Кто впервые определил наличие токов в ЦНС
А – Бергер
Б – Декарт
В

Чему равен потенциал покоя мембраны нейрона?
А. -70 мВ
Б . +20 мВ
Чему равен потенциал покоя мембраны нейрона?
А. -70 мВ
Б . +20 мВ
 Железы внутренней секреции
Железы внутренней секреции Зубы и яды
Зубы и яды Систематика бактерий. Критерии систематики. Структурно-функциональные подсистемы клетки
Систематика бактерий. Критерии систематики. Структурно-функциональные подсистемы клетки Презентация на тему "Корень" - скачать бесплатно презентации по Биологии
Презентация на тему "Корень" - скачать бесплатно презентации по Биологии Семейство Moraxellaceae. Род Moraxella. Вид M. catarrhalis
Семейство Moraxellaceae. Род Moraxella. Вид M. catarrhalis Обозначение методов. Три составных части любого обозначения
Обозначение методов. Три составных части любого обозначения ДРОЖЖИ
ДРОЖЖИ Сенсорные системы: общие принципы организации, разнообразие рецепторов, кодировка количества и качества сигналов
Сенсорные системы: общие принципы организации, разнообразие рецепторов, кодировка количества и качества сигналов Генетика человека
Генетика человека Эпителиальная ткань
Эпителиальная ткань Витамины - презентация
Витамины - презентация Методы разведения. Чистопородное разведение
Методы разведения. Чистопородное разведение Познавательно-исследовательский проект Тайны воды
Познавательно-исследовательский проект Тайны воды АТМОСФЕРНОЕ ДАВЛЕНИЕ Как атмосферное давление влияет на организм человека?
АТМОСФЕРНОЕ ДАВЛЕНИЕ Как атмосферное давление влияет на организм человека? Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Врождённое поведение человека
Врождённое поведение человека Презентация на тему "Клеточные структуры" - скачать презентации по Биологии
Презентация на тему "Клеточные структуры" - скачать презентации по Биологии Адаптация. Стресс
Адаптация. Стресс Скелет человека. Соединения костей. 8 класс
Скелет человека. Соединения костей. 8 класс Тема урока: Вирусы. Строение и классификация.
Тема урока: Вирусы. Строение и классификация.  Сон. Функции и фазы сна
Сон. Функции и фазы сна Живородящая ящерица и её содержание в неволе
Живородящая ящерица и её содержание в неволе Овечий рунец
Овечий рунец Презентация на тему "Строение растения кукурузы" - скачать презентации по Биологии
Презентация на тему "Строение растения кукурузы" - скачать презентации по Биологии Внутренняя среда организма
Внутренняя среда организма  В мире насекомых
В мире насекомых Физиология высшей нервной деятельности
Физиология высшей нервной деятельности Эволюция кровеносной системы
Эволюция кровеносной системы