- Нуклеиновые кислоты

Содержание
- 2. Полимерные цепи нуклеиновых кислот построены из мономерных единиц – нуклеотидов, в связи с чем нуклеиновые кислоты
- 3. Мономерное звено представляет собой трёхкомпонентное образование, включающее: гетероциклическое основание, углеводный остаток, фосфатную группу.
- 4. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
- 5. Углеводы β-D-рибоза
- 6. Углеводы 2-дезокси-β-D-рибоза
- 7. Входящие в состав нуклеиновых кислот гетероциклические основания пиримидинового и пуринового рядов называют нуклеиновыми основаниями.
- 8. Заместители в гетероциклическом ядре нуклеиновых оснований: оксогруппа аминогруппа одновременно обе эти группы
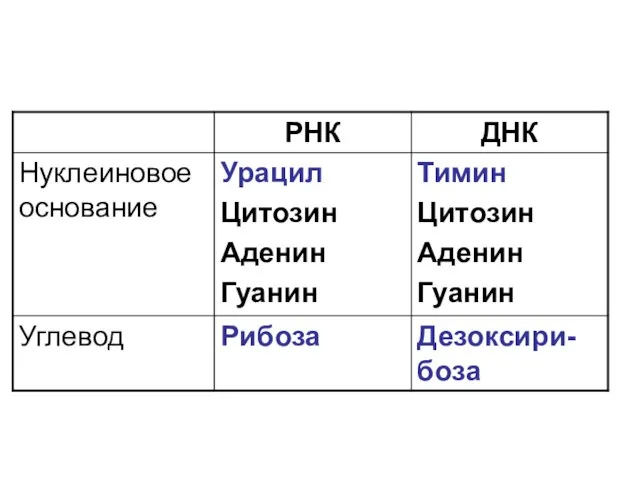
- 9. Нуклеиновые пиримидиновые основания Цитозин (Ц) Урацил (У) Тимин (Т)
- 10. Нуклеиновые пуриновые основания Аденин (А) Гуанин (Г)
- 11. Азотистое основание и углевод связаны между собой N-гликозидной связью. При этом N-гликозидная связь осуществляется между атомом
- 12. N-гликозиды нуклеиновых оснований с рибозой или дезоксирибозой – нуклеозиды. В зависимости от природы углеводного остатка различают
- 13. Общая формула нуклеозидов
- 15. Номенклатура нуклеозидов Цитозин + рибоза → цитидин Цитозин + дезоксирибоза → дезоксицитидин Аденин + рибоза →
- 16. аденозин дезоксиаденозин
- 17. цитидин дезоксицитидин
- 18. Нуклеозиды достаточно устойчивы к гидролизу в слабощелочной среде. В кислой среде они подвергаются гидролизу. При этом
- 19. Нуклеотиды - фосфаты нуклеозидов Реакция этерификации между фосфорной кислотой и нуклеозидом обычно осуществляется при С-5 или
- 20. аденозин-5’-фосфат аденозин-3’-фосфат
- 21. Номенклатура нуклеотидов
- 22. Аденозин-5'-монофосфат (АМФ) Аденозин-5'-дифосфат (АДФ) Аденозин-5'-трифосфат (АТФ)
- 23. циклический 3',5'-АМФ (цАМФ) является естественно встречающимся рибонуклеотидом (он образуется из АТФ в процессе реакции, катализируемой ферментом
- 25. ТЦА
- 26. Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями в непрерывную цепь полинуклеотида. Важной
- 27. ДНК в основном содержится в ядрах клеток, а РНК находится в рибосомах и в протоплазме клеток.
- 28. Вторичная структура ДНК Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи.
- 29. Дж. Уотсон, Ф. Крик 1953 г. Вторичная структура ДНК в виде двойной спирали Молекула ДНК состоит
- 33. Основания, располагающиеся внутри спирали, прочно упакованы и не контактируют с водой. Вода контактирует лишь с ОН
- 34. Т.е. первичная структура (нуклеотидная последовательность) одной цепи предопределяет первичную структуру второй цепи.
- 35. Правила Чаргаффа Количество пуриновых оснований равно количеству пиримидиновых оснований Количество аденина равно количеству тимина; количество гуанина
- 36. Роль комплементарных взаимодействий в осуществлении биологической функции ДНК Комплементарность цепей составляет химическую основу важнейшей функции ДНК
- 37. Однако нуклеотидная последовательность ДНК под действием различных факторов может подвергаться изменениям – мутациям. Мутация – изменение
- 38. Мутагены – вещества, вызывающие мутации: мутагены прямого действия, промутагены, которые сами по себе неактивны, но в
- 39. HNO2 Будет связывать цитозин
- 40. Третичная структура ДНК У всех живых организмов двухспиральные молекулы ДНК плотно упакованы с образованием сложных трехмерных
- 41. репликация
- 42. Синтез мРНК на матрице ДНК
- 43. тРНК
- 44. Аминокислоты, необходимые для синтеза белка, доставляются в рибосомы с помощью т-РНК. Каждая молекула т-РНК переносит только
- 46. Уровни структурной организации белковых макромолекул
- 47. Последовательность расположения аминокислотных остатков в одной или нескольких полипептидных цепях, составляющих молекулу белка, – это первичная
- 48. Кроме первичной, в белковых молекулах выделяют вторичную, третичную и четвертичную структуры. Под вторичной структурой белка подразумевают
- 49. Ключевую роль в стабилизации этой структуры играют водородные связи, которые в α–спирали образуются между карбонильным атомом
- 50. Вторичная структура белка (α-спираль)
- 51. В отличие от α–спирали β–структура образована за счёт межцепочечных водородных связей между соседними участками полипептидной цепи
- 52. Особенности вторичной структуры белка во многом определяются аминокислотным составом
- 53. Под третичной структурой белка (субъединицей) подразумевают пространственную ориентацию или способ укладки полипептидной цепи в определенном объеме,
- 54. Типы взаимодействий, стабилизирующие третичную структуру белка
- 55. а - электростатическое взаимодействие б - водородная связь в - гидрофобные взаимодействия неполярных групп г -
- 57. Под четвертичной структурой белка подразумевают ассоциированные между собой две или более субъединиц, ориентированных в пространстве. Четвертичная
- 59. Пространственная структура белковой молекулы способна нарушаться под влиянием изменения pH–среды, повышенной температуры, облучения УФ–светом и т.д.
- 60. Биологические функции белков 1. Строительная (структурная). Белки – основа протоплазмы любой клетки, основной структурный материал всех
- 61. 4. Транспортная. Белки крови транспортируют кислород, жирные кислоты, липиды, гормоны. Специальные белки переносят различные вещества через
- 62. 7. Опорная. Белки входят в состав костей скелета, сухожилий, суставов и т. д. 8. Рецепторная. Рецепторные
- 63. Классификация белков 1. По форме молекул различают фибриллярные (волокнистые) и глобулярные (корпускулярные) белки. Фибриллярные белки нерастворимы
- 64. Молекулы фибриллярных белков вытянуты в длину, нитеобразны и склонны группироваться одна около другой с образованием волокон.
- 65. Фибриллярные белки служат основным строительным материалом. К их числу относят следующие белки: кератин – в коже,
- 66. Глобулярные белки выполняют ряд функций, связанных с поддержанием и регуляцией жизненных процессов, - функций, требующих подвижности
- 67. 2. По степени сложности белки разделяют на простые и сложные. При гидролизе простых белков получаются только
- 68. К простым белкам относят: - альбумины – водорастворимые белки, составляют 50% всех белков плазмы крови человека,
- 69. - глютелины – содержатся в семенах злаков и бобовых растений; - гистоны – присутствуют в ядрах
- 70. К сложным белкам относят: - липопротеины = белок + липид. Образуются за счёт водородных связей и
- 72. Скачать презентацию

Полимерные цепи нуклеиновых кислот построены из мономерных единиц – нуклеотидов, в
Полимерные цепи нуклеиновых кислот построены из мономерных единиц – нуклеотидов, в

Мономерное звено представляет собой трёхкомпонентное образование, включающее:
гетероциклическое основание,
углеводный остаток,
фосфатную группу.
Мономерное звено представляет собой трёхкомпонентное образование, включающее:
гетероциклическое основание,
углеводный остаток,
фосфатную группу.

НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
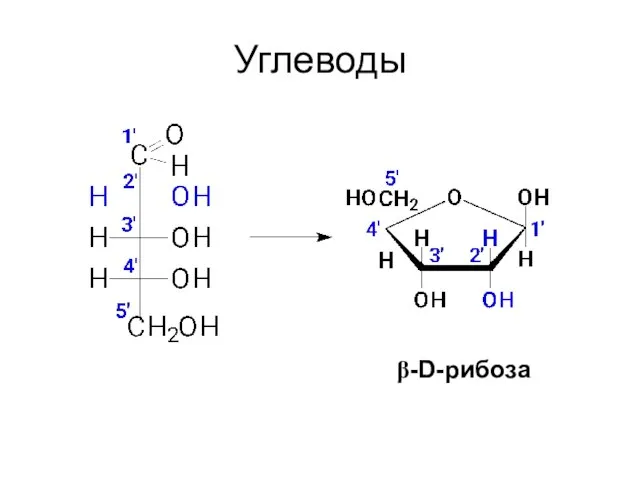
Углеводы
β-D-рибоза
Углеводы
β-D-рибоза
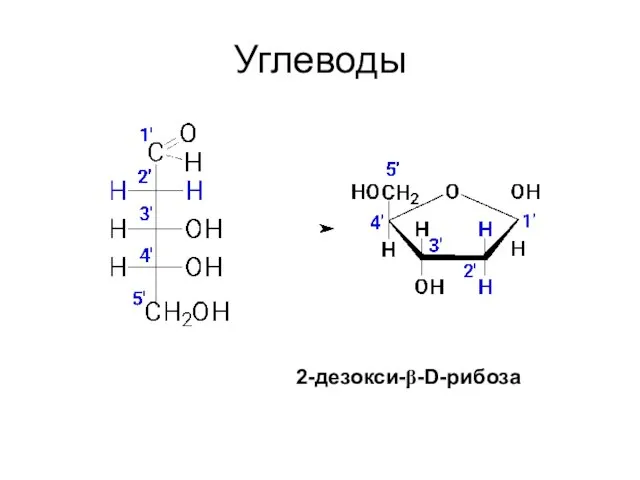
Углеводы
2-дезокси-β-D-рибоза
Углеводы
2-дезокси-β-D-рибоза

Входящие в состав нуклеиновых кислот гетероциклические основания пиримидинового и пуринового рядов
Входящие в состав нуклеиновых кислот гетероциклические основания пиримидинового и пуринового рядов

Заместители в гетероциклическом ядре нуклеиновых оснований:
оксогруппа
аминогруппа
одновременно обе эти группы
Заместители в гетероциклическом ядре нуклеиновых оснований:
оксогруппа
аминогруппа
одновременно обе эти группы
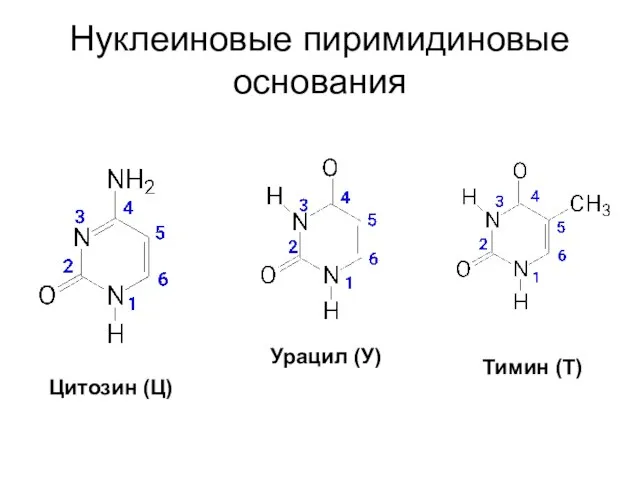
Нуклеиновые пиримидиновые основания
Цитозин (Ц)
Урацил (У)
Тимин (Т)
Нуклеиновые пиримидиновые основания
Цитозин (Ц)
Урацил (У)
Тимин (Т)
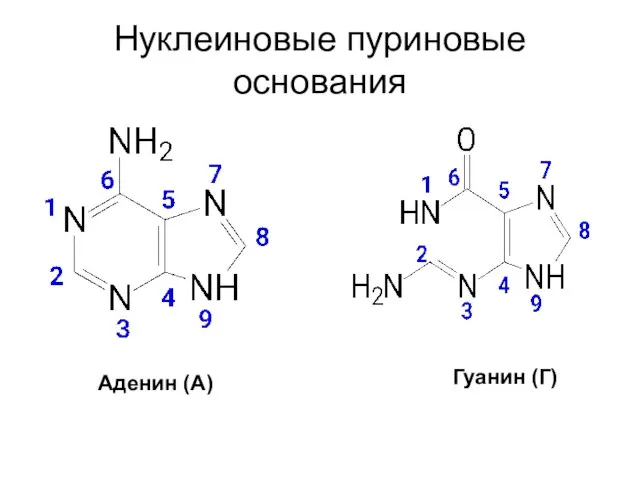
Нуклеиновые пуриновые основания
Аденин (А)
Гуанин (Г)
Нуклеиновые пуриновые основания
Аденин (А)
Гуанин (Г)

Азотистое основание и углевод связаны между собой N-гликозидной связью. При этом
Азотистое основание и углевод связаны между собой N-гликозидной связью. При этом

N-гликозиды нуклеиновых оснований с рибозой или дезоксирибозой – нуклеозиды.
В зависимости от
N-гликозиды нуклеиновых оснований с рибозой или дезоксирибозой – нуклеозиды.
В зависимости от
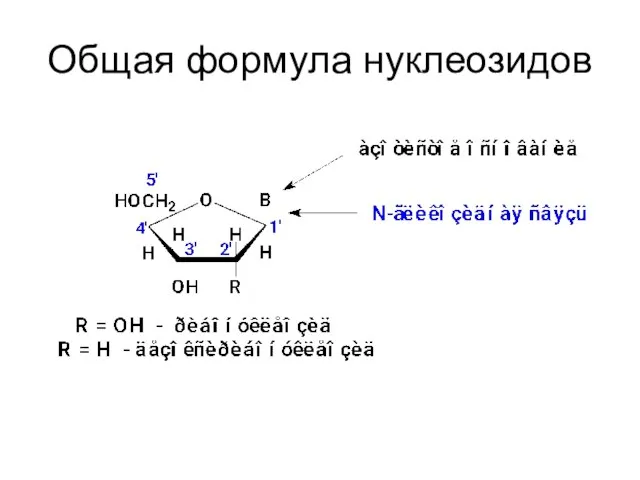
Общая формула нуклеозидов
Общая формула нуклеозидов

Номенклатура нуклеозидов
Цитозин + рибоза → цитидин
Цитозин + дезоксирибоза → дезоксицитидин
Аденин +
Номенклатура нуклеозидов
Цитозин + рибоза → цитидин
Цитозин + дезоксирибоза → дезоксицитидин
Аденин +
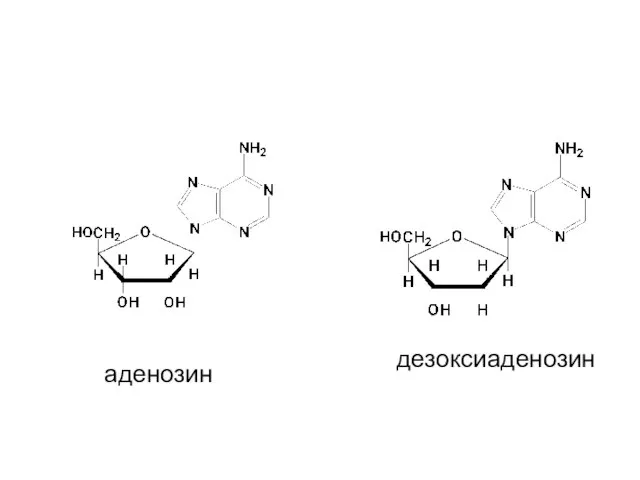
аденозин
дезоксиаденозин
аденозин
дезоксиаденозин
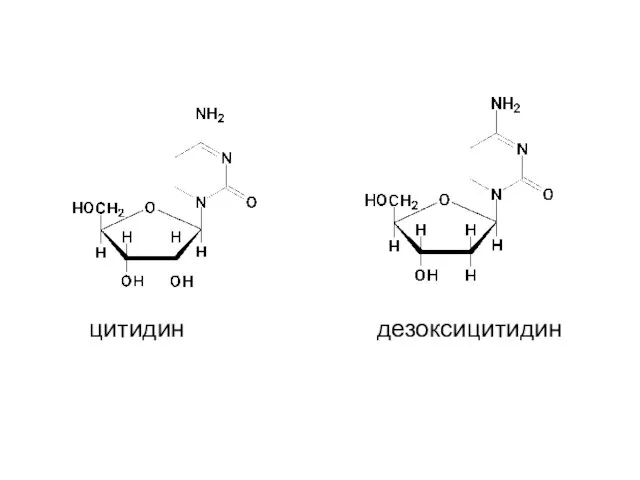
цитидин
дезоксицитидин
цитидин
дезоксицитидин

Нуклеозиды достаточно устойчивы к гидролизу в слабощелочной среде. В кислой среде
Нуклеозиды достаточно устойчивы к гидролизу в слабощелочной среде. В кислой среде

Нуклеотиды - фосфаты нуклеозидов
Реакция этерификации между фосфорной кислотой и нуклеозидом обычно
Нуклеотиды - фосфаты нуклеозидов
Реакция этерификации между фосфорной кислотой и нуклеозидом обычно
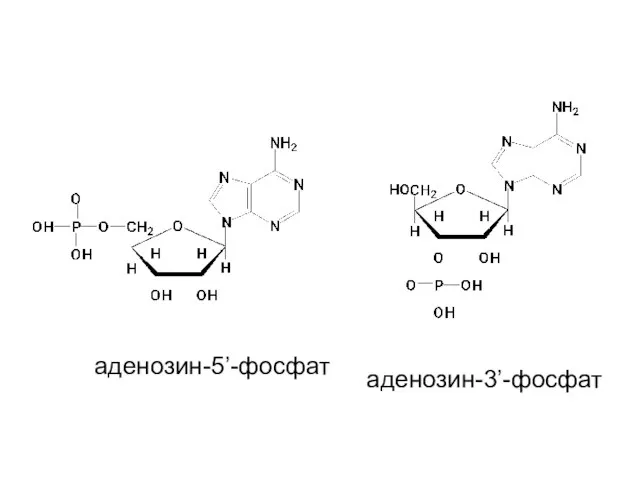
аденозин-5’-фосфат
аденозин-3’-фосфат
аденозин-5’-фосфат
аденозин-3’-фосфат
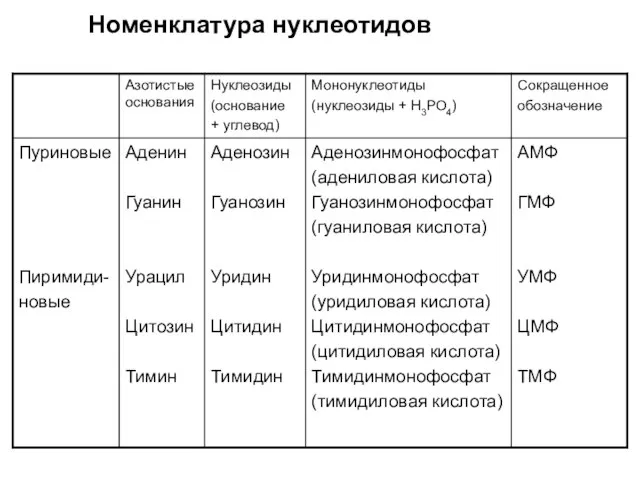
Номенклатура нуклеотидов
Номенклатура нуклеотидов
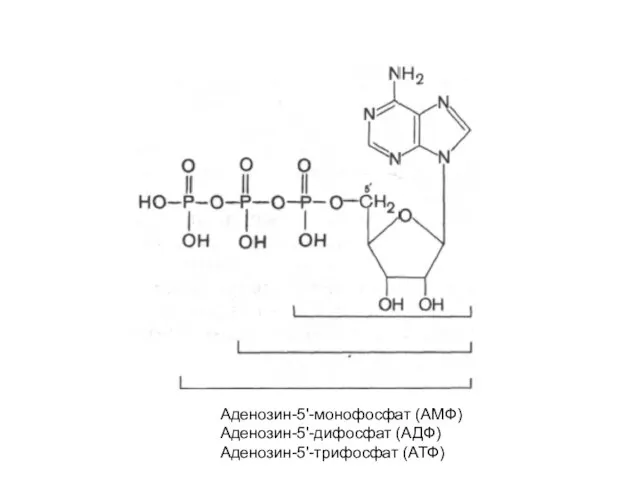
Аденозин-5'-монофосфат (АМФ)
Аденозин-5'-дифосфат (АДФ)
Аденозин-5'-трифосфат (АТФ)
Аденозин-5'-монофосфат (АМФ)
Аденозин-5'-дифосфат (АДФ)
Аденозин-5'-трифосфат (АТФ)

циклический 3',5'-АМФ (цАМФ) является естественно встречающимся рибонуклеотидом (он образуется из АТФ
циклический 3',5'-АМФ (цАМФ) является естественно встречающимся рибонуклеотидом (он образуется из АТФ
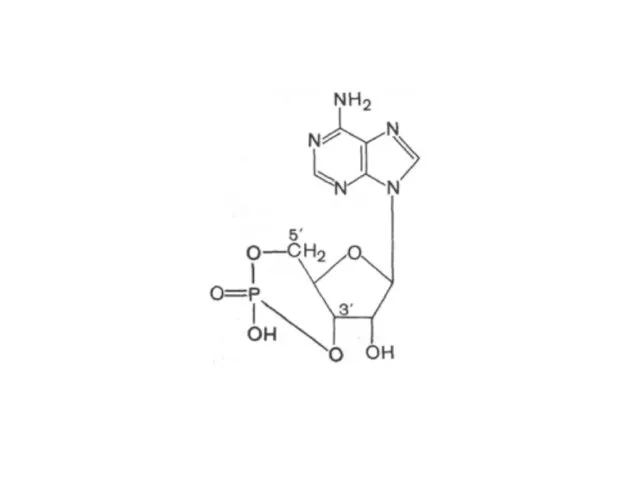
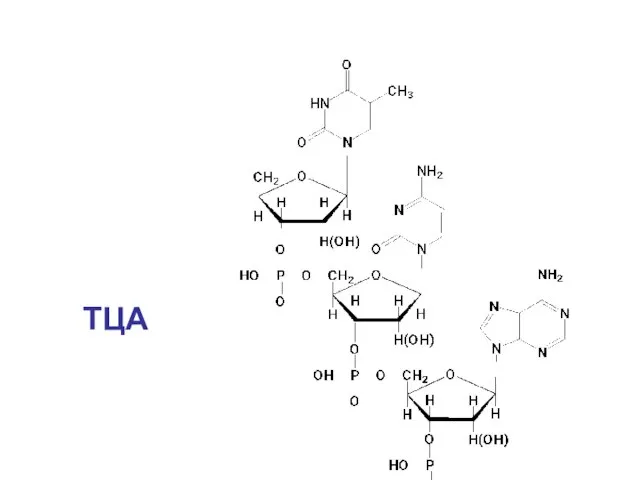
ТЦА
ТЦА

Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями
Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями

ДНК в основном содержится в ядрах клеток, а РНК находится в
ДНК в основном содержится в ядрах клеток, а РНК находится в

Вторичная структура ДНК
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи.
Вторичная структура ДНК
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи.
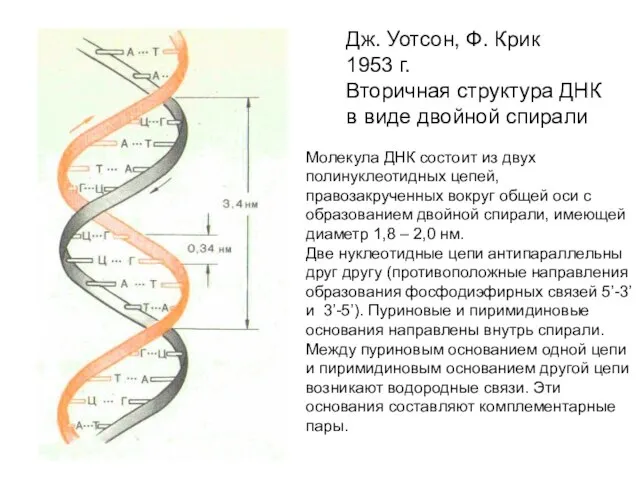
Дж. Уотсон, Ф. Крик
1953 г.
Вторичная структура ДНК
в виде двойной спирали
Молекула ДНК
Дж. Уотсон, Ф. Крик
1953 г.
Вторичная структура ДНК
в виде двойной спирали
Молекула ДНК
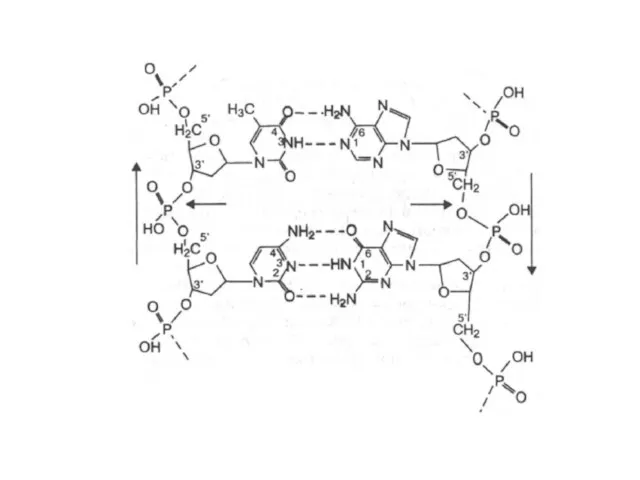
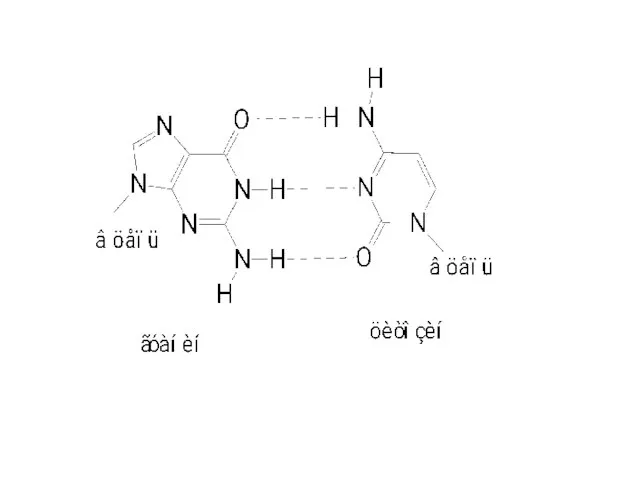

Основания, располагающиеся внутри спирали, прочно упакованы и не контактируют с водой.
Основания, располагающиеся внутри спирали, прочно упакованы и не контактируют с водой.
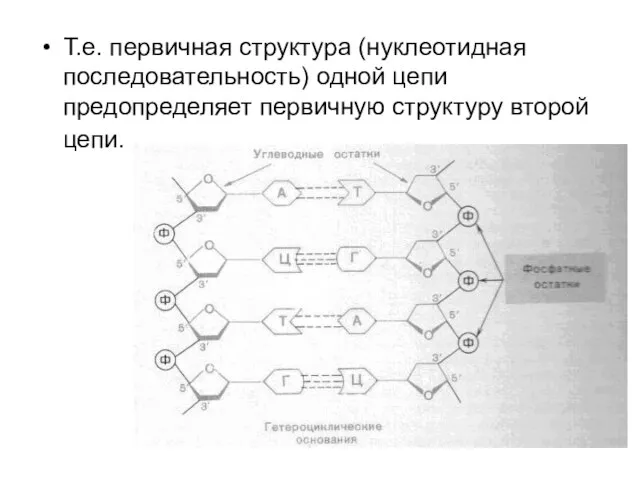
Т.е. первичная структура (нуклеотидная последовательность) одной цепи предопределяет первичную структуру второй
Т.е. первичная структура (нуклеотидная последовательность) одной цепи предопределяет первичную структуру второй

Правила Чаргаффа
Количество пуриновых оснований равно количеству пиримидиновых оснований
Количество аденина равно количеству
Правила Чаргаффа
Количество пуриновых оснований равно количеству пиримидиновых оснований
Количество аденина равно количеству

Роль комплементарных взаимодействий в осуществлении биологической функции ДНК
Комплементарность цепей составляет химическую
Роль комплементарных взаимодействий в осуществлении биологической функции ДНК
Комплементарность цепей составляет химическую

Однако нуклеотидная последовательность ДНК под действием различных факторов может подвергаться изменениям
Однако нуклеотидная последовательность ДНК под действием различных факторов может подвергаться изменениям

Мутагены – вещества, вызывающие мутации:
мутагены прямого действия,
промутагены, которые сами по себе
Мутагены – вещества, вызывающие мутации:
мутагены прямого действия,
промутагены, которые сами по себе
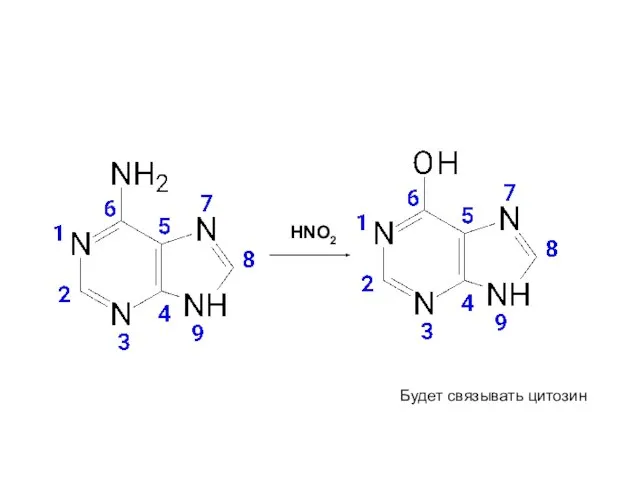
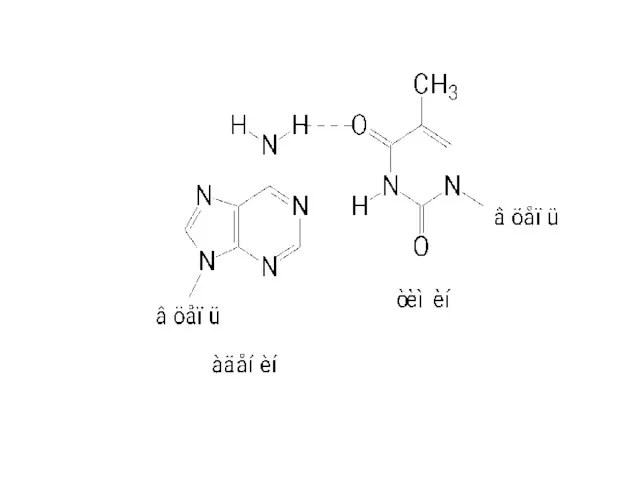
HNO2
Будет связывать цитозин
HNO2
Будет связывать цитозин

Третичная структура ДНК
У всех живых организмов двухспиральные молекулы ДНК плотно
Третичная структура ДНК
У всех живых организмов двухспиральные молекулы ДНК плотно
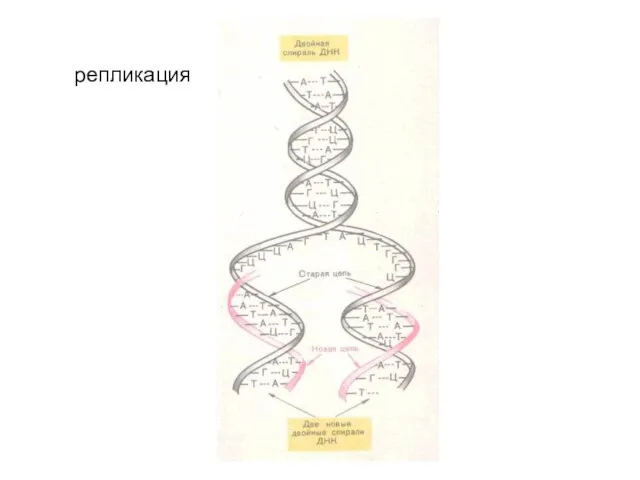
репликация
репликация
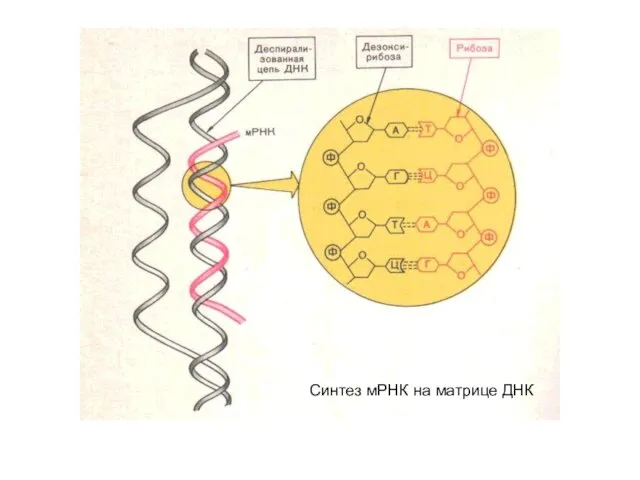
Синтез мРНК на матрице ДНК
Синтез мРНК на матрице ДНК
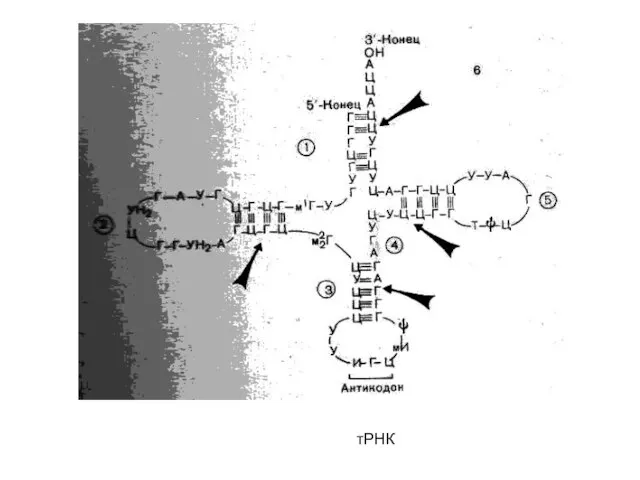
тРНК
тРНК

Аминокислоты, необходимые для синтеза белка, доставляются в рибосомы с помощью т-РНК. Каждая молекула
Аминокислоты, необходимые для синтеза белка, доставляются в рибосомы с помощью т-РНК. Каждая молекула


Уровни структурной организации белковых макромолекул
Уровни структурной организации белковых макромолекул

Последовательность расположения аминокислотных остатков в одной или нескольких полипептидных цепях, составляющих
Последовательность расположения аминокислотных остатков в одной или нескольких полипептидных цепях, составляющих

Кроме первичной, в белковых молекулах выделяют вторичную, третичную и четвертичную структуры.
Под
Кроме первичной, в белковых молекулах выделяют вторичную, третичную и четвертичную структуры.
Под

Ключевую роль в стабилизации этой структуры играют водородные связи, которые в
Ключевую роль в стабилизации этой структуры играют водородные связи, которые в
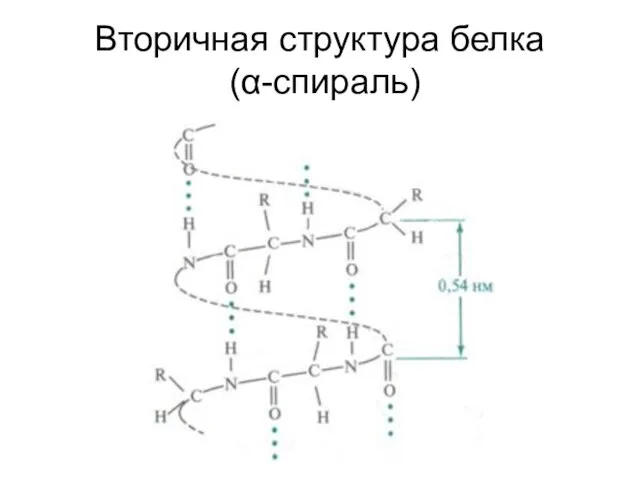
Вторичная структура белка
(α-спираль)
Вторичная структура белка
(α-спираль)
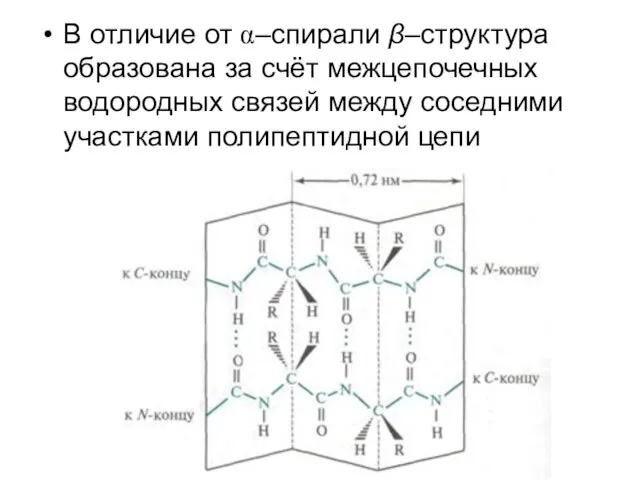
В отличие от α–спирали β–структура образована за счёт межцепочечных водородных связей
В отличие от α–спирали β–структура образована за счёт межцепочечных водородных связей

Особенности вторичной структуры белка во многом определяются аминокислотным составом
Особенности вторичной структуры белка во многом определяются аминокислотным составом

Под третичной структурой белка (субъединицей) подразумевают пространственную ориентацию или способ укладки
Под третичной структурой белка (субъединицей) подразумевают пространственную ориентацию или способ укладки
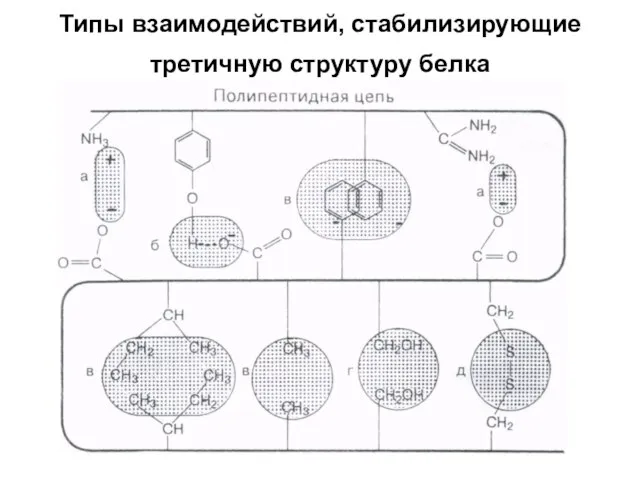
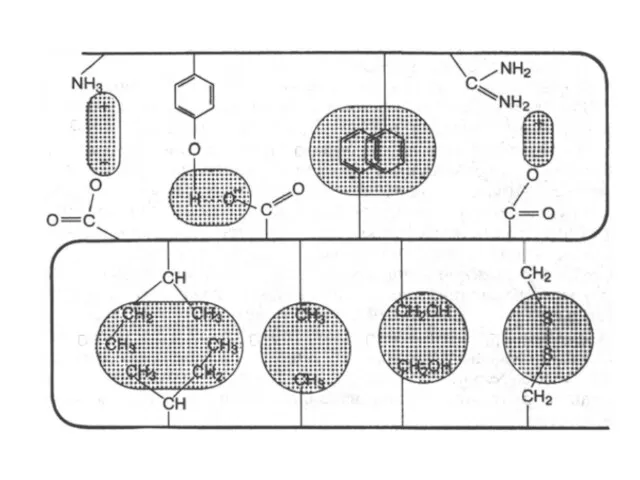
Типы взаимодействий, стабилизирующие третичную структуру белка
Типы взаимодействий, стабилизирующие третичную структуру белка

а - электростатическое взаимодействие
б - водородная связь
в - гидрофобные взаимодействия неполярных
а - электростатическое взаимодействие
б - водородная связь
в - гидрофобные взаимодействия неполярных

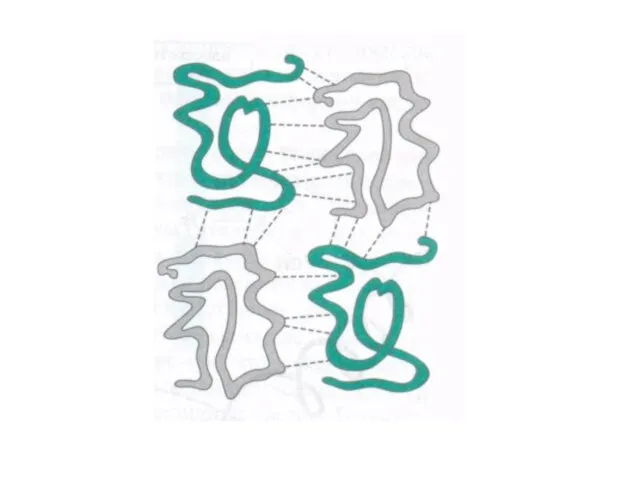
Под четвертичной структурой белка подразумевают ассоциированные между собой две или более
Под четвертичной структурой белка подразумевают ассоциированные между собой две или более

Пространственная структура белковой молекулы способна нарушаться под влиянием изменения pH–среды, повышенной
Пространственная структура белковой молекулы способна нарушаться под влиянием изменения pH–среды, повышенной

Биологические функции белков
1. Строительная (структурная). Белки – основа протоплазмы любой
Биологические функции белков
1. Строительная (структурная). Белки – основа протоплазмы любой

4. Транспортная. Белки крови транспортируют кислород, жирные кислоты, липиды, гормоны. Специальные
4. Транспортная. Белки крови транспортируют кислород, жирные кислоты, липиды, гормоны. Специальные

7. Опорная. Белки входят в состав костей скелета, сухожилий, суставов и
7. Опорная. Белки входят в состав костей скелета, сухожилий, суставов и

Классификация белков
1. По форме молекул различают фибриллярные (волокнистые) и глобулярные
Классификация белков
1. По форме молекул различают фибриллярные (волокнистые) и глобулярные

Молекулы фибриллярных белков вытянуты в длину, нитеобразны и склонны группироваться одна
Молекулы фибриллярных белков вытянуты в длину, нитеобразны и склонны группироваться одна

Фибриллярные белки служат основным строительным материалом. К их числу относят следующие
Фибриллярные белки служат основным строительным материалом. К их числу относят следующие

Глобулярные белки выполняют ряд функций, связанных с поддержанием и регуляцией жизненных
Глобулярные белки выполняют ряд функций, связанных с поддержанием и регуляцией жизненных

2. По степени сложности белки разделяют на простые и сложные. При
2. По степени сложности белки разделяют на простые и сложные. При

К простым белкам относят:
- альбумины – водорастворимые белки, составляют 50% всех
К простым белкам относят:
- альбумины – водорастворимые белки, составляют 50% всех

- глютелины – содержатся в семенах злаков и бобовых растений;
- гистоны
- глютелины – содержатся в семенах злаков и бобовых растений;
- гистоны

К сложным белкам относят:
- липопротеины = белок + липид. Образуются за
К сложным белкам относят:
- липопротеины = белок + липид. Образуются за
 Обмен белков. Общие пути обмена аминокислот
Обмен белков. Общие пути обмена аминокислот Умные материалы
Умные материалы Свойства металла. Подготовительная группа 2
Свойства металла. Подготовительная группа 2 Химическая кинетика и катализ. Смещение химического равновесия
Химическая кинетика и катализ. Смещение химического равновесия Методы выращивания GaAs. Сравнение
Методы выращивания GaAs. Сравнение Хроматография как метод разделения и обнаружения в качественном анализе Выполнила: студентка 2 курса Акаева Дарья
Хроматография как метод разделения и обнаружения в качественном анализе Выполнила: студентка 2 курса Акаева Дарья  Внеклассное мероприятие: Свойства активированного угля
Внеклассное мероприятие: Свойства активированного угля Нефть: состав, основные физико-химические свойства. Классификация нефтей
Нефть: состав, основные физико-химические свойства. Классификация нефтей Массасы 4,2 г көміртек (IV) оксиді сумен әрекеттескенде қанша грамм көмір қышқылы (Н2СО3) түзілетінін есепте
Массасы 4,2 г көміртек (IV) оксиді сумен әрекеттескенде қанша грамм көмір қышқылы (Н2СО3) түзілетінін есепте Сера и ее аллотропные модификации. (9 класс)
Сера и ее аллотропные модификации. (9 класс) Типы химических реакций
Типы химических реакций  Ионно-координационная полимеризация
Ионно-координационная полимеризация Технология получения полиуретанов
Технология получения полиуретанов Коррозия металлов
Коррозия металлов Известные химики
Известные химики Проблемы загрязнения воды Ташбаев Далерхан 7 «А»
Проблемы загрязнения воды Ташбаев Далерхан 7 «А» Качественные и количественные методы обработки хроматограмм
Качественные и количественные методы обработки хроматограмм Оксиды углерода
Оксиды углерода «Углерод». Презентация к открытому уроку химии 9 класс. Выполнила: Отряскина Т. А.
«Углерод». Презентация к открытому уроку химии 9 класс. Выполнила: Отряскина Т. А.  Презентация по Химии "Галогени" - скачать смотреть бесплатно
Презентация по Химии "Галогени" - скачать смотреть бесплатно Химическая кинетика
Химическая кинетика Fiber. Iron. Calcium
Fiber. Iron. Calcium Свойства растворов и гетерогенных систем
Свойства растворов и гетерогенных систем Небезпечні хімічні речовини Підготувала: учениця 8 – А класу Кіровоградського НВК №34 Подколзіна Анна
Небезпечні хімічні речовини Підготувала: учениця 8 – А класу Кіровоградського НВК №34 Подколзіна Анна  Агрегатные состояния вещества
Агрегатные состояния вещества Основания. Состав оснований
Основания. Состав оснований Презентация по Химии "Кинетика химических реакций.Химическое равновесие" - скачать смотреть
Презентация по Химии "Кинетика химических реакций.Химическое равновесие" - скачать смотреть  Гидролиз органических веществ
Гидролиз органических веществ